
Rock Refuges Are Strongly Associated with Increased Urban Occupancy in the Western Fence Lizard, Sceloporus occidentalis


Abstract
1. Introduction
2. Materials and Methods
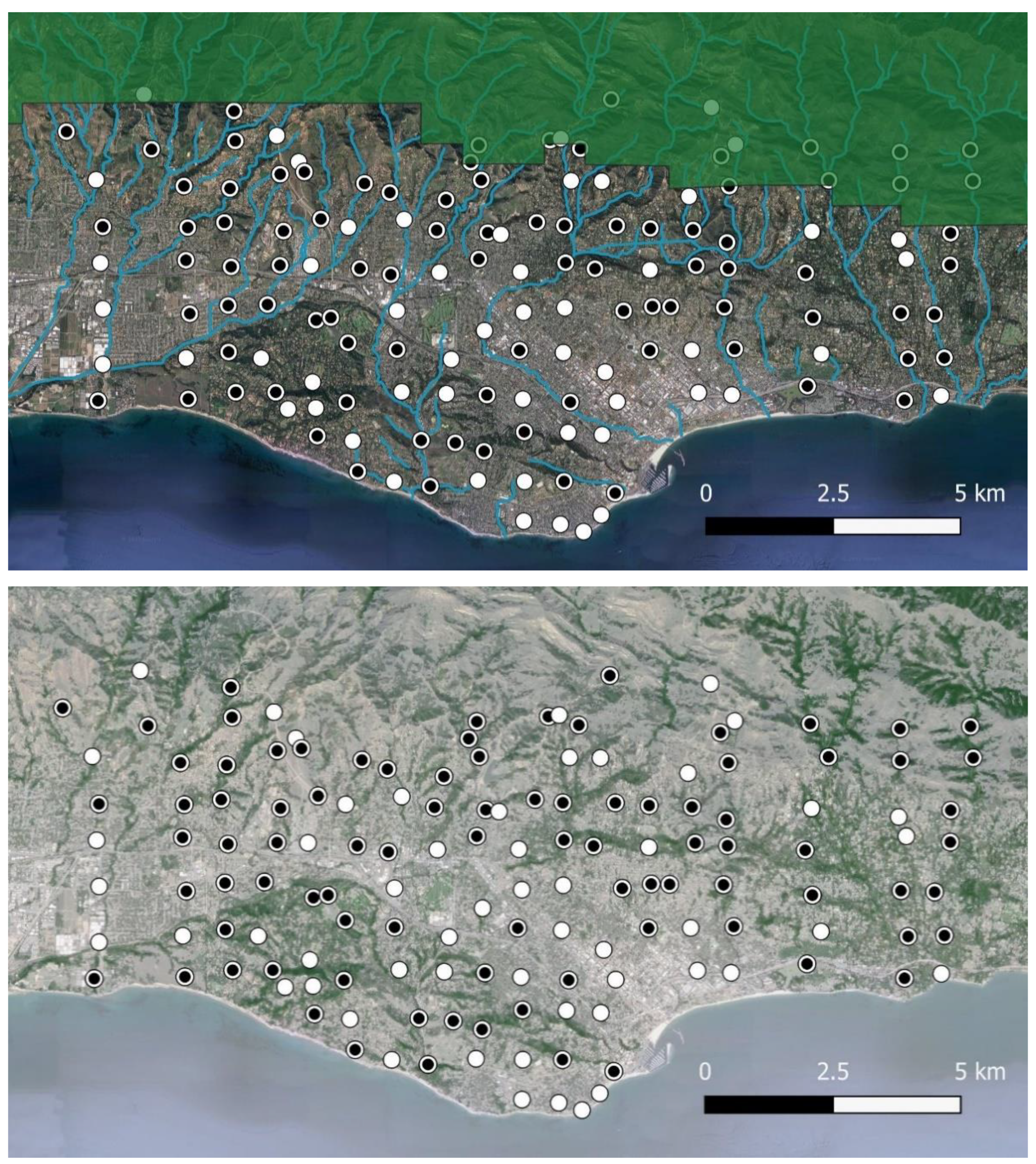
2.1. Study Area
2.2. Field Sampling
2.3. Statistical Analysis
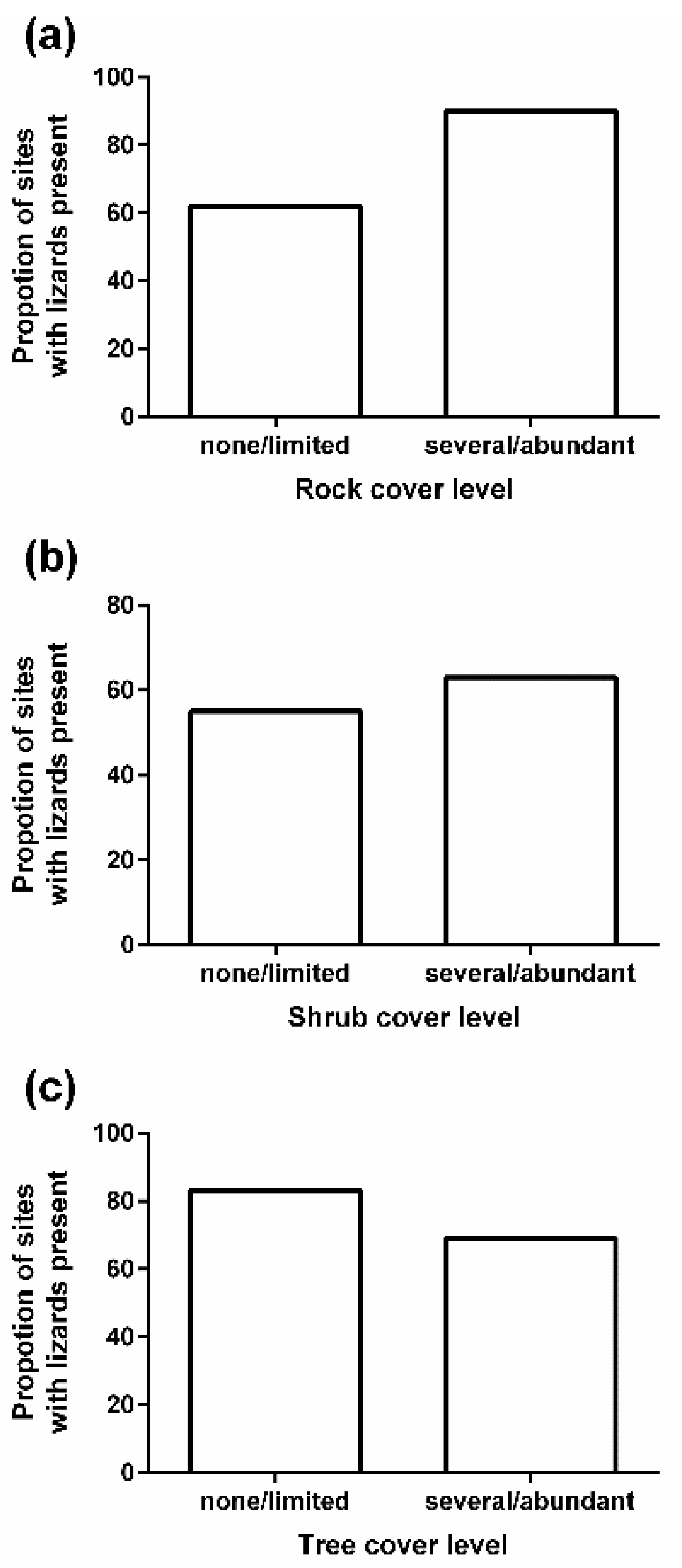
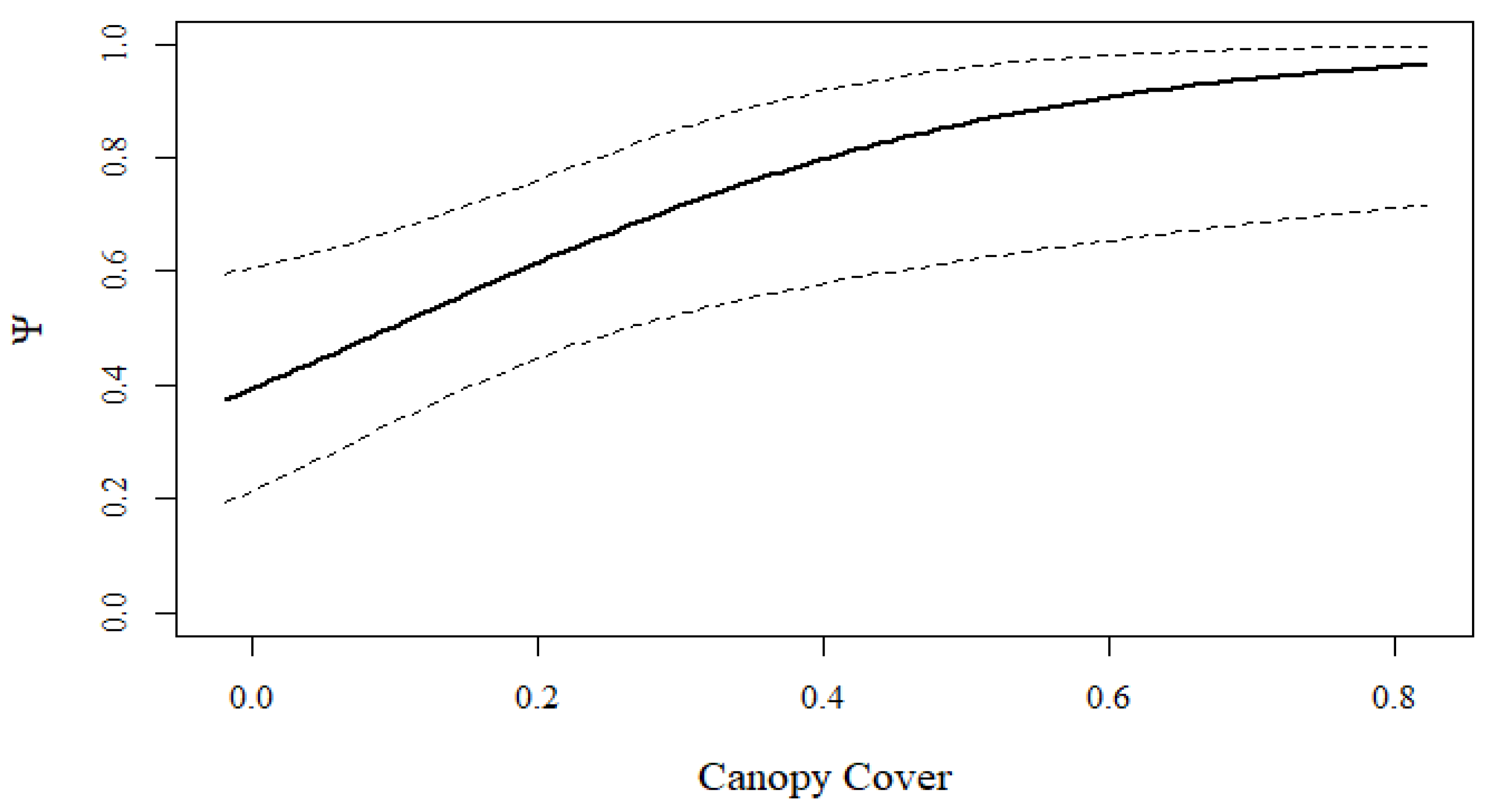
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brook, B.W.; Sodhi, N.S.; Bradshaw, C.J.A. Synergies among Extinction Drivers under Global Change. Trends Ecol. Evol. 2008, 23, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Crowl, T.A.; Crist, T.O.; Parmenter, R.R.; Belovsky, G.; Lugo, A.E. The Spread of Invasive Species and Infectious Disease as Drivers of Ecosystem Change. Front. Ecol. Environ. 2008, 6, 238–246. [Google Scholar] [CrossRef]
- Kendall, R.J.; Lacher, T.E.; Cobb, G.C.; Cox, S.B. (Eds.) Wildlife Toxicology: Emerging Contaminant and Biodiversity Issues; CRC Press: Boca Raton, FL, USA, 2010; ISBN 978-0-429-15018-0. [Google Scholar]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Niemelä, J.; Breuste, J.H.; Guntenspergen, G.; McIntyre, N.E.; Elmqvist, T.; James, P. Urban Ecology: Patterns, Processes, and Applications; Oxford University Press: Oxford, UK, 2011; ISBN 978-0-19-161323-4. [Google Scholar]
- Elmqvist, T.; Fragkias, M.; Goodness, J.; Güneralp, B.; Marcotullio, P.J.; McDonald, R.I.; Parnell, S.; Schewenius, M.; Sendstad, M.; Seto, K.C. Urbanization, Biodiversity and Ecosystem Services: Challenges and Opportunities: A Global Assessment; Springer Nature: Heidelberg, Germany, 2013. [Google Scholar]
- Ditchkoff, S.S.; Saalfeld, S.T.; Gibson, C.J. Animal Behavior in Urban Ecosystems: Modifications Due to Human-Induced Stress. Urban Ecosyst. 2006, 9, 5–12. [Google Scholar] [CrossRef]
- Shochat, E.; Warren, P.S.; Faeth, S.H.; McIntyre, N.E.; Hope, D. From Patterns to Emerging Processes in Mechanistic Urban Ecology. Trends Ecol. Evol. 2006, 21, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.D.; Cleeton, S.H.; Lyons, T.P.; Miller, J.R. Urbanization and the Predation Paradox: The Role of Trophic Dynamics in Structuring Vertebrate Communities. BioScience 2012, 62, 809–818. [Google Scholar] [CrossRef]
- Forman, R.T.T. Urban Ecology: Science of Cities; Cambridge University Press: New York, NY, USA, 2014; ISBN 978-1-107-00700-0. [Google Scholar]
- McDonnell, M.J.; Hahs, A.K. The Future of Urban Biodiversity Research: Moving beyond the ‘Low-Hanging Fruit’. Urban Ecosyst. 2013, 16, 397–409. [Google Scholar] [CrossRef]
- Ikin, K.; Le Roux, D.S.; Rayner, L.; Villaseñor, N.R.; Eyles, K.; Gibbons, P.; Manning, A.D.; Lindenmayer, D.B. Key Lessons for Achieving Biodiversity-Sensitive Cities and Towns. Ecol. Manag. Restor. 2015, 16, 206–214. [Google Scholar] [CrossRef]
- Ouyang, J.Q.; Isaksson, C.; Schmidt, C.; Hutton, P.; Bonier, F.; Dominoni, D. A New Framework for Urban Ecology: An Integration of Proximate and Ultimate Responses to Anthropogenic Change. Integr. Comp. Biol. 2018, 58, 915–928. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Warren, P.S. Urban Bird Ecology and Conservation; University of California Press: Berkeley, CA, USA, 2012; ISBN 978-0-520-95389-5. [Google Scholar]
- Gil, D.; Brumm, H. Avian Urban Ecology; Oxford University Press: Oxford, UK, 2014; ISBN 978-0-19-966158-9. [Google Scholar]
- Murgui, E.; Hedblom, M. Ecology and Conservation of Birds in Urban Environments; Springer: Heidelberg, Germany, 2017; ISBN 978-3-319-43314-1. [Google Scholar]
- Boal, C.W.; Dykstra, C.R. Urban Raptors: Ecology and Conservation of Birds of Prey in Cities; Island Press: Washington, DC, USA, 2018; ISBN 978-1-61091-840-4. [Google Scholar]
- Böhm, M.; Collen, B.; Baillie, J.E.M.; Bowles, P.; Chanson, J.; Cox, N.; Hammerson, G.; Hoffmann, M.; Livingstone, S.R.; Ram, M.; et al. The Conservation Status of the World’s Reptiles. Biol. Conserv. 2013, 157, 372–385. [Google Scholar] [CrossRef]
- McCallum, M.L. Vertebrate Biodiversity Losses Point to a Sixth Mass Extinction. Biodiv. Conserv. 2015, 24, 2497–2519. [Google Scholar] [CrossRef]
- Tonini, J.F.R.; Beard, K.H.; Ferreira, R.B.; Jetz, W.; Pyron, R.A. Fully-Sampled Phylogenies of Squamates Reveal Evolutionary Patterns in Threat Status. Biol. Conserv. 2016, 204, 23–31. [Google Scholar] [CrossRef]
- Zipkin, E.F.; DiRenzo, G.V.; Ray, J.M.; Rossman, S.; Lips, K.R. Tropical Snake Diversity Collapses after Widespread Amphibian Loss. Science 2020, 367, 814–816. [Google Scholar] [CrossRef] [PubMed]
- Gardner, T.A.; Barlow, J.; Peres, C.A. Paradox, presumption and pitfalls in conservationbiology: The importance of habitat change for amphibians and reptiles. Biol. Conserv. 2007, 138, 166–179. [Google Scholar] [CrossRef]
- Keinath, D.A.; Doak, D.F.; Hodges, K.E.; Prugh, L.R.; Fagan, W.; Sekercioglu, C.H.; Buchart, S.H.M.; Kauffman, M. A Global Analysis of Traits Predicting Species Sensitivity to Habitat Fragmentation. Glob. Biol. Biogeogr. 2017, 26, 115–127. [Google Scholar] [CrossRef]
- Doherty, T.S.; Balouch, S.; Bell, K.; Burns, T.J.; Feldman, A.; Fist, C.; Garvey, T.F.; Jessop, T.S.; Meiri, S.; Driscoll, D.A. Reptile Responses to Anthropogenic Habitat Modification: A Global Meta-Analysis. Glob. Biol. Biogeogr. 2020, 29, 1265–1279. [Google Scholar] [CrossRef]
- Mancera, K.; Murray, P.; Lisle, A.; Dupont, C.; Faucheux, F.; Phillips, C. The Effects of Acute Exposure to Mining Machinery Noise on the Behaviour of Eastern Blue-Tongued Lizards (Tiliqua scincoides). Anim. Welf. 2017, 26, 11–24. [Google Scholar] [CrossRef]
- Kolbe, J.J.; VanMiddlesworth, P.; Battles, A.C.; Stroud, J.T.; Buffum, B.; Forman, R.T.T.; Losos, J.B. Determinants of Spread in an Urban Landscape by an Introduced Lizard. Landsc. Ecol. 2016, 31, 1795–1813. [Google Scholar] [CrossRef]
- Mohanty, N.P.; Joshi, M.; Thaker, M. Urban Lizards Use Sleep Sites That Mirror the Structural, Thermal, and Light Properties of Natural Sites. Behav. Ecol. Sociobiol. 2021, 75, 166. [Google Scholar] [CrossRef]
- Kechnebbou, M.; Martín, J.; Chammem, M.; Arregui, L.; Nouira, S. Roads and Urban Areas as Physiological Stressors of Spiny-Tailed Lizards, Uromastyx acanthinura. J. Arid. Environ. 2019, 170, 103997. [Google Scholar] [CrossRef]
- Wotherspoon, D.; Burgin, S. The Impact on Native Herpetofauna Due to Traffic Collision at the Interface between a Suburban Area and the Greater Blue Mountains World Heritage Area: An Ecological Disaster? Austral. Zool. 2011, 35, 1040–1046. [Google Scholar] [CrossRef]
- Germaine, S.S.; Wakeling, B.F. Lizard Species Distributions and Habitat Occupation along an Urban Gradient in Tucson, Arizona, USA. Biol. Conserv. 2001, 97, 229–237. [Google Scholar] [CrossRef]
- French, S.S.; Fokidis, H.B.; Moore, M.C. Variation in Stress and Innate Immunity in the Tree Lizard (Urosaurus ornatus) across an Urban–Rural Gradient. J. Comp. Physiol. B 2008, 178, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Ackley, J.W.; Wu, J.; Angilletta, M.J.; Myint, S.W.; Sullivan, B. Rich Lizards: How Affluence and Land Cover Influence the Diversity and Abundance of Desert Reptiles Persisting in an Urban Landscape. Biol. Conserv. 2015, 182, 87–92. [Google Scholar] [CrossRef]
- Beninde, J.; Feldmeier, S.; Werner, M.; Peroverde, D.; Schulte, U.; Hochkirch, A.; Veith, M. Cityscape Genetics: Structural vs. Functional Connectivity of an Urban Lizard Population. Mol. Ecol. 2016, 25, 4984–5000. [Google Scholar] [CrossRef]
- Kolbe, J.J.; Moniz, H.A.; Lapiedra, O.; Thawley, C.J. Bright Lights, Big City: An Experimental Assessment of Short-Term Behavioral and Performance Effects of Artificial Light at Night on Anolis Lizards. Urban Ecosyst. 2021, 24, 1035–1045. [Google Scholar] [CrossRef]
- Hall, J.M.; Warner, D.A. Body Size and Reproduction of a Non-Native Lizard Are Enhanced in an Urban Environment. Biol. J. Linn. Soc. 2017, 122, 860–871. [Google Scholar] [CrossRef]
- Lapiedra, O. Urban Behavioral Ecology: Lessons from Anolis Lizards. Integr. Comp. Biol. 2018, 58, 939–947. [Google Scholar] [CrossRef]
- Sparkman, A.; Howe, S.; Hynes, S.; Hobbs, B.; Handal, K. Parallel Behavioral and Morphological Divergence in Fence Lizards on Two College Campuses. PLoS ONE 2018, 13, e0191800. [Google Scholar] [CrossRef]
- Winchell, K.M.; Carlen, E.J.; Puente-Rolón, A.R.; Revell, L.J. Divergent Habitat Use of Two Urban Lizard Species. Ecol. Evol. 2018, 8, 25–35. [Google Scholar] [CrossRef]
- Batabyal, A.; Thaker, M. Lizards from Suburban Areas Learn Faster to Stay Safe. Biol. Lett. 2019, 15, 20190009. [Google Scholar] [CrossRef] [PubMed]
- Tiatragul, S.; Hall, J.M.; Warner, D.A. Nestled in the City Heat: Urban Nesting Behavior Enhances Embryo Development of an Invasive Lizard. J. Urban Ecol. 2020, 6, juaa001. [Google Scholar] [CrossRef]
- Delaney, K.S.; Busteed, G.; Fisher, R.N.; Riley, S.P.D. Reptile and Amphibian Diversity and Abundance in an Urban Landscape: Impacts of Fragmentation and the Conservation Value of Small Patches. Ichthyol. Herpetol. 2021, 109, 424–435. [Google Scholar] [CrossRef]
- Putman, B.J.; Williams, R.; Li, E.; Pauly, G.B. The Power of Community Science to Quantify Ecological Interactions in Cities. Sci. Rep. 2021, 11, 3069. [Google Scholar] [CrossRef]
- French, S.S.; Webb, A.C.; Hudson, S.B.; Virgin, E.E. Town and Country Reptiles: A Review of Reptilian Responses to Urbanization. Integr. Comp. Biol. 2018, 58, 948–966. [Google Scholar] [CrossRef]
- Putman, B.J.; Samia, D.S.; Cooper, W.E.; Blumstein, D.T. Impact of Human-Induced Environmental Changes on Lizard Behavior: Insights from Urbanization. In Behavior of Lizards; CRC Press: Boca Raton, FL, USA, 2019; pp. 375–396. [Google Scholar]
- Putman, B.J.; Tippie, Z.A. Big City Living: A Global Meta-Analysis Reveals Positive Impact of Urbanization on Body Size in Lizards. Front. Ecol. Evol. 2020, 8, 580745. [Google Scholar] [CrossRef]
- Kitchener, D.J.; Chapman, A.; Dell, J.; Muir, B.G.; Palmer, M. Lizard Assemblage and Reserve Size and Structure in the Western Australian Wheatbelt—Some Implications for Conservation. Biol. Conserv. 1980, 17, 25–62. [Google Scholar] [CrossRef]
- James, C. Response of Vertebrates to Fenceline Contrasts in Grazing Intensity in Semi-Arid Woodlands of Eastern Australia. Austral. Ecol. 2003, 28, 137–151. [Google Scholar] [CrossRef]
- Jellinek, S.; Driscoll, D.A.; Kirkpatrick, J.B. Environmental and Vegetation Variables Have a Greater Influence than Habitat Fragmentation in Structuring Lizard Communities in Remnant Urban Bushland. Austral. Ecol. 2004, 29, 294–304. [Google Scholar] [CrossRef]
- Castellano, M.J.; Valone, T.J. Effects of Livestock Removal and Perennial Grass Recovery on the Lizards of a Desertified Arid Grassland. J. Arid. Environ. 2006, 66, 87–95. [Google Scholar] [CrossRef]
- Wilgers, D.J.; Horne, E.A.; Sandercock, B.K.; Volkmann, A.W. Effects of Rangeland Management on Community Dynamics of the Herpetofauna of the Tallgrass Prairie. Herpetologica 2006, 62, 378–388. [Google Scholar] [CrossRef]
- Dibner, R.R.; Doak, D.F.; Murphy, M. Discrepancies in Occupancy and Abundance Approaches to Identifying and Protecting Habitat for an At-Risk Species. Ecol. Evol. 2017, 7, 5692–5702. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, T.S.; Folt, B.; Hall, J.M. Dumpsters and Other Anthropogenic Structures as Habitat for Invasive African Rock Agama Lizards in Florida. Biol. Invasions 2021, 23, 2689–2693. [Google Scholar] [CrossRef]
- Downes, S.; Shine, R. Heat, Safety or Solitude? Using Habitat Selection Experiments to Identify a Lizard’s Priorities. Anim. Behav. 1998, 55, 1387–1396. [Google Scholar] [CrossRef] [PubMed]
- Monasterio, C.; Salvador, A.; Díaz, J.A. Altitude and Rock Cover Explain the Distribution and Abundance of a Mediterranean Alpine Lizard. J. Herpetol. 2010, 44, 158–163. [Google Scholar] [CrossRef]
- van Heezik, Y.; Ludwig, K. Proximity to Source Populations and Untidy Gardens Predict Occurrence of a Small Lizard in an Urban Area. Landsc. Urban Plan. 2012, 104, 253–259. [Google Scholar] [CrossRef]
- Stebbins, R.C. Peterson Field Guide to Western Reptiles & Amphibians, 3rd ed.; Houghton Mifflin Company: Boston, MA, USA, 2003. [Google Scholar]
- Marcellini, D.; MacKey, J.P. Habitat Preferences of the Lizards, Sceloporus occidentalis and S. Graciosus (Lacertilia, Iguanidae). Herepetologica 1970, 26, 51–56. [Google Scholar]
- Rose, B.R. Habitat and Prey Selection of Sceloporus occidentalis and Sceloporus graciosus. Ecology 1976, 57, 531–541. [Google Scholar] [CrossRef]
- Grolle, E.K.; Lopez, M.C.; Gerson, M.M. Flight Initiation Distance Differs between Populations of Western Fence Lizards (Sceloporus occidentalis) at a Rural and an Urban Site. Bull. South. Calif. Acad. Sci. 2014, 113, 42–46. [Google Scholar] [CrossRef][Green Version]
- Putman, B.J.; Gasca, M.; Blumstein, D.T.; Pauly, G.B. Downsizing for Downtown: Limb Lengths, Toe Lengths, and Scale Counts Decrease with Urbanization in Western Fence Lizards (Sceloporus occidentalis). Urban Ecosyst. 2019, 22, 1071–1081. [Google Scholar] [CrossRef]
- Climate of Santa Barbara, California. Available online: https://repository.library.noaa.gov/view/noaa/14480 (accessed on 25 January 2022).
- Parker, V.T. Chaparral of California. In Encyclopedia of the World’s Biomes; Goldstein, M.I., DellaSala, D.A., Eds.; Elsevier: Oxford, UK, 2020; pp. 457–472. ISBN 978-0-12-816097-8. [Google Scholar]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Andrew Royle, J.; Langtimm, C.A. Estimating Site Occupancy Rates When Detection Probabilities Are Less than One. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Hines, J. PRESENCE 3.1 Software to Estimate Patch Occupancy and Related Parameters. 2006. Available online: http://www.mbr-pwrc.usgs.gov/software/presence.html (accessed on 1 September 2021).
- Smith, G.T.; Arnold, G.W.; Sarre, S.; Abensperg-Traun, M.; Steven, D.E. The Effect of Habitat Fragmentation and Livestock Grazing on Animal Communities in Remnants of Gimlet Eucalyptus salubris Woodland in the Western Australian Wheatbelt. II. Lizards. J. Appl. Ecol. 1996, 33, 1302–1310. [Google Scholar] [CrossRef]
- MacNally, R.; Brown, G.W. Reptiles and Habitat Fragmentation in the Box-Ironbark Forests of Central Victoria, Australia: Predictions, Compositional Change and Faunal Nestedness. Oecologia 2001, 128, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Delaney, K.S.; Riley, S.P.D.; Fisher, R.N. A Rapid, Strong, and Convergent Genetic Response to Urban Habitat Fragmentation in Four Divergent and Widespread Vertebrates. PLoS ONE 2010, 5, e12767. [Google Scholar] [CrossRef] [PubMed]
- Santos, T.; Díaz, J.A.; Pérez-Tris, J.; Carbonell, R.; Tellería, J.L. Habitat Quality Predicts the Distribution of a Lizard in Fragmented Woodlands Better than Habitat Fragmentation. Anim. Cons. 2008, 11, 46–56. [Google Scholar] [CrossRef]
- Ackley, J.W.; Angilletta, M.J.; DeNardo, D.; Sullivan, B.; Wu, J. Urban Heat Island Mitigation Strategies and Lizard Thermal Ecology: Landscaping Can Quadruple Potential Activity Time in an Arid City. Urban Ecosyst. 2015, 18, 1447–1459. [Google Scholar] [CrossRef]
- Somaweera, R.; Wijayathilaka, N.; Bowatte, G.; Meegaskumbura, M. Conservation in a Changing Landscape: Habitat Occupancy of the Critically Endangered Tennent’s Leaf-Nosed Lizard (Ceratophora tennentii) in Sri Lanka. J. Nat. Hist. 2015, 49, 1961–1985. [Google Scholar] [CrossRef]
- Koenig, J.; Shine, R.; Shea, G. The Ecology of an Australian Reptile Icon: How Do Blue-Tongued Lizards (Tiliqua scincoides) Survive in Suburbia? Wildl. Res. 2001, 28, 214–227. [Google Scholar] [CrossRef]
- Grover, M.C. Microhabitat Use and Thermal Ecology of Two Narrowly Sympatric Sceloporus (Phrynosomatidae) Lizards. J. Herpetol. 1996, 30, 152–160. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Model | AICc | ∆AICc | ω |
|---|---|---|---|
| 1 rock cover + tree cover + land type | 511.05 | 0 | 0.2124 |
| 2 rock cover + land type | 511.34 | 0.29 | 0.1838 |
| 3 rock cover + tree cover + canopy cover | 512.29 | 1.24 | 0.1143 |
| 4 rock cover + shrub cover + tree cover + land type | 512.3 | 1.25 | 0.1137 |
| 5 rock cover + shrub cover + land type | 512.67 | 1.62 | 0.0945 |
| 6 rock cover + canopy cover | 513.23 | 2.18 | 0.0714 |
| 7 rock cover + tree cover | 513.54 | 2.49 | 0.0612 |
| 8 rock cover + shrub cover + tree cover + canopy cover | 513.75 | 2.7 | 0.0551 |
| 9 rock cover | 513.76 | 2.71 | 0.0548 |
| 10 tree cover + canopy cover | 514.54 | 3.49 | 0.0371 |
| 1 group, constant p (null model) | 520.58 | 9.53 | 0.0018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rurik, A.J.; Wilmoth, S.C.; Dayton, K.E.; Sparkman, A.M. Rock Refuges Are Strongly Associated with Increased Urban Occupancy in the Western Fence Lizard, Sceloporus occidentalis. Diversity 2022, 14, 655. https://doi.org/10.3390/d14080655
Rurik AJ, Wilmoth SC, Dayton KE, Sparkman AM. Rock Refuges Are Strongly Associated with Increased Urban Occupancy in the Western Fence Lizard, Sceloporus occidentalis. Diversity. 2022; 14(8):655. https://doi.org/10.3390/d14080655
Chicago/Turabian StyleRurik, Alexander J., Seth C. Wilmoth, Kendra E. Dayton, and Amanda M. Sparkman. 2022. "Rock Refuges Are Strongly Associated with Increased Urban Occupancy in the Western Fence Lizard, Sceloporus occidentalis" Diversity 14, no. 8: 655. https://doi.org/10.3390/d14080655
APA StyleRurik, A. J., Wilmoth, S. C., Dayton, K. E., & Sparkman, A. M. (2022). Rock Refuges Are Strongly Associated with Increased Urban Occupancy in the Western Fence Lizard, Sceloporus occidentalis. Diversity, 14(8), 655. https://doi.org/10.3390/d14080655