
Genetic Variation in Gaultheria nummularioides (Ericaceae: Gaultherieae) from the Sky Islands of the Himalaya-Hengduan Mountains

Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Sequencing
2.3. Phylogenetic Reconstruction
2.4. Phylogeographic Analyses
2.5. Population Divergence Estimation
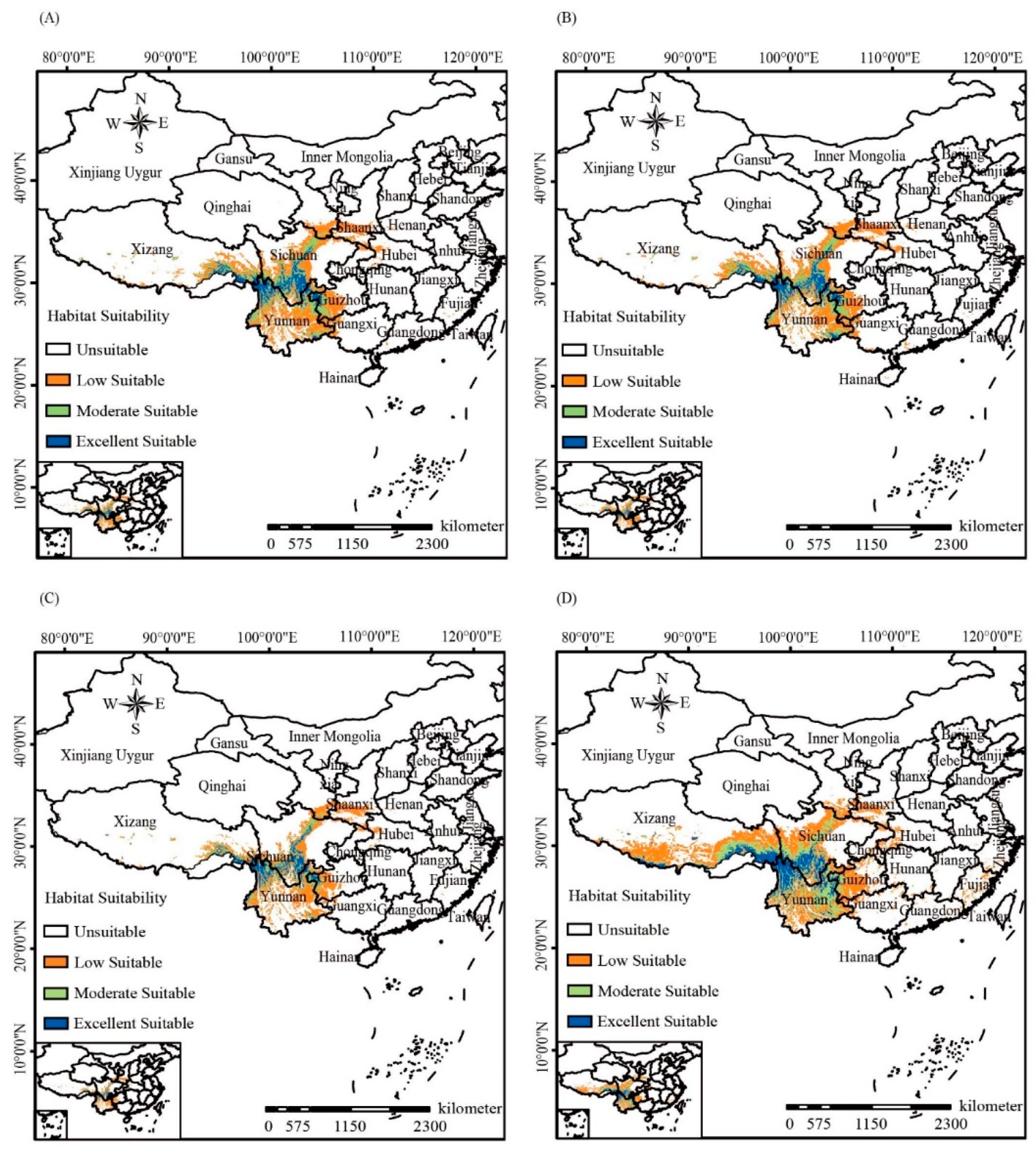
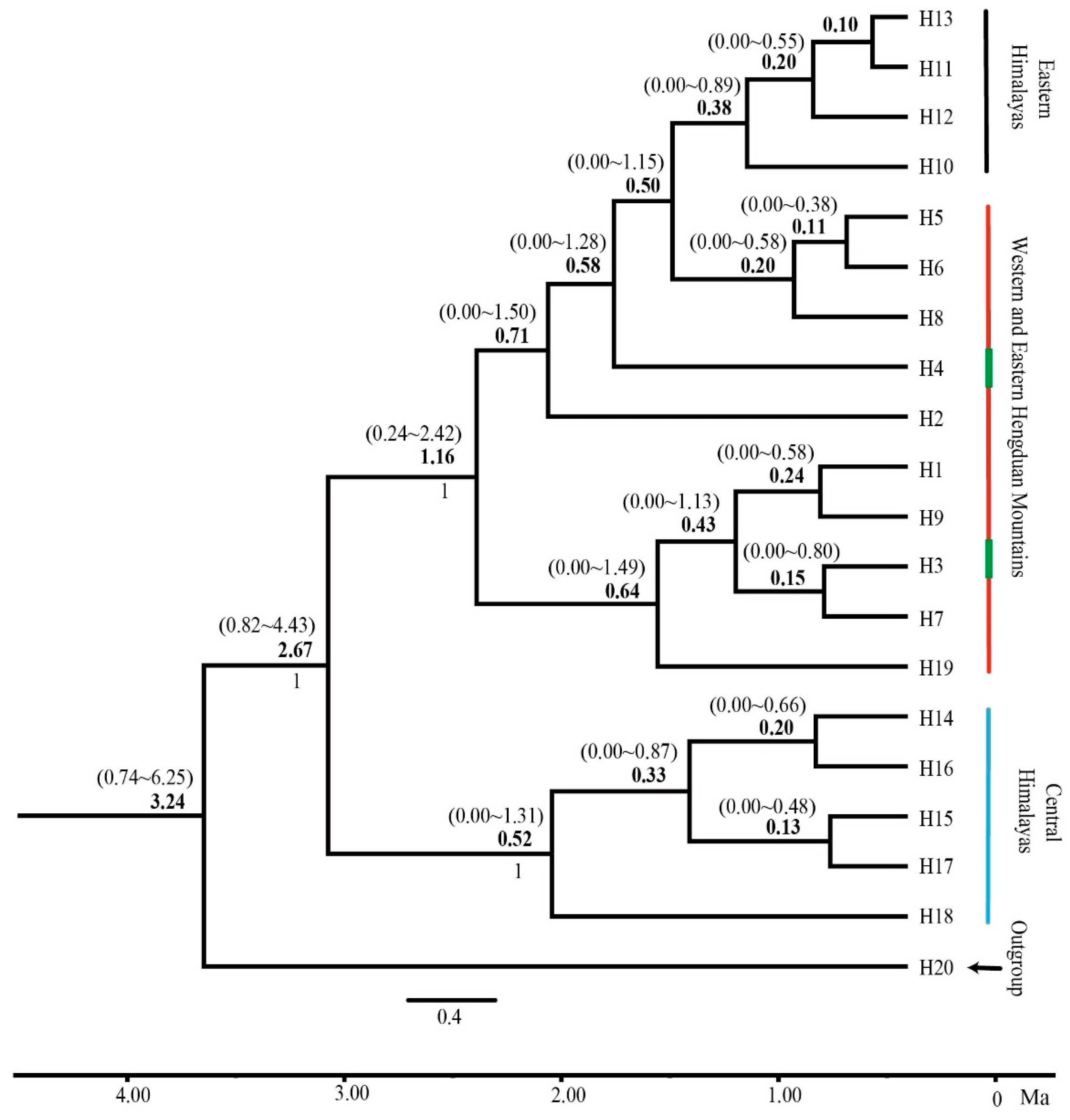
3. Results
3.1. Phylogenetic Tree Reconstruction
3.2. Genetic Diversity
3.3. Phylogeographic Analyses
3.4. Divergence of Population Haplotypes
4. Discussion
4.1. Genetic Differentiation within and among Populations
4.2. Genetic Divergence Correlated with Pleistocene Glacial Cycle-Induced Climatic Shifts and Monsoons in Montane Habitats
4.3. Morphological Variation among Various Populations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heald, W.F. Sky islands in Arizona. Nat. Hist. 1951, 60, 95–96. [Google Scholar]
- He, K.; Jiang, X.L. Sky islands of southwest China. I: An overview of phylogeographic patterns. Chin. Sci. Bull. 2014, 59, 585–597. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- DeBano, L.F. The Madrean Sky Island Archipelago: A Planetary Overview. In Biodiversity and Management of the Madrean Archipelago: The Sky Islands of Southwestern United States and Northwestern Mexico, September 19–23, 1994; University of Arizona: Tucson, AZ, USA, 1995. [Google Scholar]
- He, K.; Gutierrez, E.E.; Heming, N.M.; Koepfli, K.P.; Wan, T.; He, S.W.; Jin, W.; Liu, S.Y.; Jiang, X.L. Cryptic phylogeographic history sheds light on the generation of species diversity in sky-island mountains. J. Biogeogr. 2019, 46, 2232–2247. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000; pp. viii + 447. [Google Scholar]
- Luo, D.; Xu, B.; Rana, S.K.; Li, Z.M.; Sun, H. Phylogeography of rare fern Polystichum glaciale endemic to the subnival zone of the Sino-Himalaya. Plant Syst. Evol. 2018, 304, 485–499. [Google Scholar] [CrossRef]
- Luo, D.; Xu, B.; Li, Z.M.; Sun, H. The ‘Ward Line-Mekong-Salween Divide’ is an important floristic boundary between the eastern Himalaya and Hengduan Mountains: Evidence from the phylogeographical structure of subnival herbs Marmoritis complanatum (Lamiaceae). Bot. J. Linn. Soc. 2017, 185, 482–496. [Google Scholar] [CrossRef]
- Luo, D.; Yue, J.P.; Sun, W.G.; Xu, B.; Li, Z.M.; Comes, H.P.; Sun, H. Evolutionary history of the subnival flora of the Himalaya-Hengduan Mountains: First insights from comparative phylogeography of four perennial herbs. J. Biogeogr. 2016, 43, 31–43. [Google Scholar] [CrossRef]
- Li, X.W.; Jie, L. The Tanaka-Kaiyong line—An important floristic line for the study of the flora of East Asia. Ann. Mo. Bot. Gard. 1997, 84, 888–892. [Google Scholar]
- Zhang, Q.; Chiang, T.Y.; George, M.; Liu, J.Q.; Abbott, R.J. Phylogeography of the Qinghai-Tibetan Plateau endemic Juniperus przewalskii (Cupressaceae) inferred from chloroplast DNA sequence variation. Mol. Ecol. 2005, 14, 3513–3524. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.S.; Li, Y.F.; Ding, X.; Wang, X.Q. Extensive population expansion of Pedicularis longiflora (Orobanchaceae) on the Qinghai-Tibetan Plateau and its correlation with the Quaternary climate change. Mol. Ecol. 2008, 17, 5135–5145. [Google Scholar] [CrossRef]
- Yang, F.S.; Qin, A.L.; Li, Y.F.; Wang, X.Q. Great genetic differentiation among populations of Meconopsis integrifolia and its implication for plant speciation in the Qinghai-Tibetan Plateau. PLoS ONE 2012, 7, e37196. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.P.; Mateo, R.G.; Liu, J.Q.; Suchan, T.; Alvarez, N.; Guisan, A.; Conti, E.; Salamin, N. Genetic consequences of Quaternary climatic oscillations in the Himalayas: Primula tibetica as a case study based on restriction site-associated DNA sequencing. New Phytol. 2017, 213, 1500–1512. [Google Scholar] [CrossRef]
- Lin, N.; Deng, T.; Moore, M.J.; Sun, Y.X.; Huang, X.H.; Sun, W.G.; Luo, D.; Wang, H.C.; Zhang, J.W.; Sun, H. Phylogeography of Parasyncalathium souliei (Asteraceae) and its potential application in delimiting phylogeoregions in the Qinghai-Tibet Plateau (QTP)-Hengduan Mountains (HDM) Hotspot. Front. Genet. 2018, 9, 171. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.S.; Wang, B.X.; Gao, L.; Xiong, H.B.; Tang, W. Ecological study on molecular phylogeography of Quercus aquifolioides, an endemic oak in SW China. Ekoloji 2019, 28, 899–914. [Google Scholar]
- Fritsch, P.W.; Lu, L.; Bush, C.M.; Cruz, B.C.; Kron, K.A.; Li, D.Z. Phylogenetic analysis of the Wintergreen Group (Ericaceae) based on six genic regions. Syst. Bot. 2011, 36, 990–1003. [Google Scholar] [CrossRef]
- Kron, K.A.; Judd, W.S.; Stevens, P.F.; Crayn, D.M.; Anderberg, A.A.; Gadek, P.A.; Quinn, C.J.; Luteyn, J.L. Phylogenetic classification of Ericaceae: Molecular and morphological evidence. Bot. Rev. 2002, 68, 335–423. [Google Scholar] [CrossRef]
- Middleton, D.J.; Wilcock, C.C. A critical examination of the status of Pernettya as a genus distinct from Gaultheria. Edinb. J. Bot. 1990, 47, 291–301. [Google Scholar] [CrossRef]
- Middleton, D.J. Infrageneric classification of the genus Gaultheria L (Ericaceae). Bot. J. Linn. Soc. 1991, 106, 229–258. [Google Scholar] [CrossRef]
- Lu, L.; Fritsch, P.W.; Bush, C.M.; Wang, H.; Kron, K.A.; Li, D.Z. Allopolyploidy in the Wintergreen Group of tribe Gaultherieae (Ericaceae) inferred from low-copy nuclear genes. Nord. J. Bot. 2019, 37, e02077. [Google Scholar] [CrossRef]
- Fritsch, P.W.; Zhou, L.; Lu, L.; Bartholomew, B. The flowering plant genus Gaultheria (Ericaceae) in the Gaoligong Shan, along the border region of China and Myanmar. Proc. Calif. Acad. Sci. 2008, 59, 147–214. [Google Scholar]
- Lu, L.; Fritsch, P.W.; Cruz, B.C.; Wang, H.; Li, D.Z. Reticulate evolution, cryptic species, and character convergence in the core East Asian clade of Gaultheria (Ericaceae). Mol. Phylogenet. Evol. 2010, 57, 364–379. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.Z.; Stevens, P.F. Gaultheria. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2005; Volume 14, pp. 464–475. [Google Scholar]
- Slentz, S.; Boyd, A.E.; McDade, L.A. Morphological differentiation among Madrean sky island populations of Castilleja austromontana (Scrophulariaceae). Madroño 1999, 46, 100–111. [Google Scholar]
- Mayr, E. Change of Genetic Environment and Evolution; Allen & Unwin: London, UK, 1954. [Google Scholar]
- Wright, S. Isolation by distance. Genetics 1943, 28, 114–138. [Google Scholar] [CrossRef] [PubMed]
- DeChaine, E.G.; Martin, A.P. Marked genetic divergence among sky island populations of Sedum lanceolatum (Crassulaceae) in the Rocky Mountains. Am. J. Bot. 2005, 92, 477–486. [Google Scholar] [CrossRef]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef]
- Hewitt, G.M. The structure of biodiversity—Insights from molecular phylogeography. Front. Zool. 2004, 1, 4. [Google Scholar] [CrossRef]
- Slatkin, M. Gene flow and the geographic structure of natural-populations. Science 1987, 236, 787–792. [Google Scholar] [CrossRef]
- Manel, S.; Holderegger, R. Ten years of landscape genetics. Trends Ecol. Evol. 2013, 28, 614–621. [Google Scholar] [CrossRef]
- Wiens, J.J.; Camacho, A.; Goldberg, A.; Jezkova, T.; Kaplan, M.E.; Lambert, S.M.; Miller, E.C.; Streicher, J.W.; Walls, R.L. Climate change, extinction, and Sky Island biogeography in a montane lizard. Mol. Ecol. 2019, 28, 2610–2624. [Google Scholar] [CrossRef]
- Hirao, A.S.; Shimono, Y.; Narita, K.; Wada, N.; Kudo, G. Ecotypic divergences of the alpine herb Potentilla matsumurae adapted to fellfield-snowbed habitats across a series of mountain sky islands. Am. J. Bot. 2019, 106, 772–787. [Google Scholar] [CrossRef]
- Fritsch, P.W.; Manchester, S.R.; Stone, R.D.; Cruz, B.C.; Almeda, F. Northern Hemisphere origins of the amphi-Pacific tropical plant family Symplocaceae. J. Biogeogr. 2015, 42, 891–901. [Google Scholar] [CrossRef]
- Huang, J.C.; Ge, X.J.; Sun, M. Modified CTAB protocol using a silica matrix for isolation of plant genomic DNA. Biotechniques 2000, 28, 432–434. [Google Scholar] [CrossRef] [PubMed]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Fritsch, P.W.; Ma, P.F.; Wang, H.; Lu, L.; Li, D.Z. Plastid phylogenomics and adaptive evolution of Gaultheria series Trichophyllae (Ericaceae), a clade from sky islands of the Himalaya-Hengduan Mountains. Mol. Phylogenet. Evol. 2017, 110, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Aubriot, X.; Soulebeau, A.; Haevermans, T.; Schatz, G.E.; Cruaud, C.; Lowry, P.P. Molecular phylogenetics of Sarcolaenaceae (Malvales), Madagascar’s largest endemic plant family. Bot. J. Linn. Soc. 2016, 182, 729–743. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Pons, O.; Petit, R.J. Measuring and testing genetic differentiation with ordered versus unordered alleles. Genetics 1996, 144, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Masatoshi, N. Molecular Population Genetics and Evolution; North-Holland Publishing Company: Amsterdam, The Netherlands, 1975. [Google Scholar]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Beaumont, M.A. Adaptation and speciation: What can Fst tell us? Trends Ecol. Evol. 2005, 20, 435–440. [Google Scholar] [CrossRef]
- Miguel, I.; Iriondo, M.; Garnery, L.; Sheppard, W.S.; Estonba, A. Gene flow within the M evolutionary lineage of Apis mellifera: Role of the Pyrenees, isolation by distance and post-glacial re-colonization routes in the western Europe. Apidologie 2007, 38, 141–155. [Google Scholar] [CrossRef]
- Smouse, P.E.; Long, J.C.; Sokal, R.R. Multiple-regression and correlation extensions of the Mantel test of matrix correspondence. Syst. Zool. 1986, 35, 627–632. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [PubMed]
- Slatkin, M.; Hudson, R.R. Pairwise comparisons of mitochondrial DNA sequences in stable and exponentially growing populations. Genetics 1991, 129, 555–562. [Google Scholar] [PubMed]
- Harpending, H.C. Signature of ancient population-growth in a low-resolution mitochondrial DNA mismatch distribution. Hum. Biol. 1994, 66, 591–600. [Google Scholar]
- Kawabe, A.; Yamane, K.; Miyashita, N.T. DNA polymorphism at the cytosolic phosphoglucose isomerase (PgiC) locus of the wild plant Arabidopsis thaliana. Genetics 2000, 156, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.H.; Li, W.H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.E.; Pennington, R.T.; Pennington, T.D.; Hollingsworth, P.M. Rapid diversification of a species-rich genus of neotropical rain forest trees. Science 2001, 293, 2242–2245. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Meng, F.Y.; Zhou, G.F.; Li, Y.F.; Wang, B.; Lu, H. Assessing the suitable cultivation areas for Scutellaria baicalensis in China using the Maxent model and multiple linear regression. Biochem. Syst. Ecol. 2020, 90. [Google Scholar] [CrossRef]
- Lu, C.Y.; Gu, W.; Dai, A.H.; Wei, H.Y. Assessing habitat suitability based on geographic information system (GIS) and fuzzy: A case study of Schisandra sphenanthera Rehd. et Wils. in Qinling Mountains, China. Ecol. Modell. 2012, 242, 105–115. [Google Scholar] [CrossRef]
- Mousazade, M.; Ghanbarian, G.; Pourghasemi, H.R.; Safaeian, R.; Cerda, A. Maxent data mining technique and its comparison with a bivariate statistical model for predicting the potential distribution of Astragalus fasciculifolius Boiss. in Fars, Iran. Sustainability 2019, 11, 3452. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.C.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jimenez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Global Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.Q.; Zhang, L.; Wang, J.N.; Wang, W.W.; Niyati, N.; Guo, Y.L.; Wang, X.F. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. Sci. Total Environ. 2021, 755, 142548. [Google Scholar] [CrossRef] [PubMed]
- Marmion, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. The performance of state-of-the-art modelling techniques depends on geographical distribution of species. Ecol. Modell. 2009, 220, 3512–3520. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Fritsch, P.W.; Matzke, N.J.; Wang, H.; Kron, K.A.; Li, D.Z.; Wiens, J.J. Why is fruit colour so variable? Phylogenetic analyses reveal relationships between fruit-colour evolution, biogeography and diversification. Global Ecol. Biogeogr. 2019, 28, 891–903. [Google Scholar] [CrossRef]
- Xiang, Q.Y.; Thomas, D.T.; Xiang, Q.P. Resolving and dating the phylogeny of Cornales—Effects of taxon sampling, data partitions, and fossil calibrations. Mol. Phylogenet. Evol. 2011, 59, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J. Tracer, V1.6. Available online: http://tree.bio.ed.ac.uk/software/tracer/ (accessed on 4 July 2022).
- Tian, B.; Zhou, Z.L.; Du, F.K.; He, C.Z.; Xin, P.Y.; Ma, H.C. The Tanaka Line shaped the phylogeographic pattern of the cotton tree (Bombax ceiba) in southwest China. Biochem. Syst. Ecol. 2015, 60, 150–157. [Google Scholar] [CrossRef]
- Yue, L.L.; Chen, G.; Sun, W.B.; Sun, H. Phylogeography of Buddleja crispa (Buddlejaceae) and its correlation with drainage system evolution in Southwestern China. Am. J. Bot. 2012, 99, 1726–1735. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, W.B.; Wang, Z.L.; Guan, K.Y.; Yang, J.B. Isolation and characterization of microsatellite loci for Hibiscus aridicola (Malvaceae), an endangered plant endemic to the dry-hot valleys of Jinsha River in Southwest China. Int. J. Mol. Sci. 2011, 12, 5698–5704. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, D.L.A.; Balslev, H.; Hansen, M.M.; Sklenar, P.; Romoleroux, K. Low genetic variation and high differentiation across sky island populations of Lupinus alopecuroides (Fabaceae) in the northern Andes. Alpine Bot. 2016, 126, 135–142. [Google Scholar] [CrossRef]
- Sekar, S.; Karanth, P. Flying between Sky Islands: The effect of naturally fragmented habitat on butterfly population structure. PLoS ONE 2013, 8, e71573. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Tan, K.; Ren, M.X. Effects of monsoon on distribution patterns of tropical plants in Asia. Chin. J. Plant Ecol. 2017, 10, 1103–1112. [Google Scholar]
- Roy, A.B.; Purohit, R. Chapter 17—The Himalayas: Evolution through collision. In Indian Shield; Roy, A.B., Purohit, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 311–327. [Google Scholar]
- Zhao, L.F.; Li, L.; Liao, J.; Dong, S.X.; Liang, Y.L.; Gao, R. Shear-wave velocity reveals heterogeneous geometry of the main Himalayan thrust system and deep structure beneath the Nepal Himalayas. Geochem. Geophys. Geosyst. 2022, 23, e2021GC010263. [Google Scholar] [CrossRef]
- Garg, P.K.; Shukla, A.; Jasrotia, A.S. On the strongly imbalanced state of glaciers in the Sikkim, eastern Himalaya, India. Sci. Total Environ. 2019, 691, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Bhattari, S.; Chhetri, D.B. Ecosystem Profile Eastern Himalayas Region; WWF-US, Asia Program: Washington, DC, USA, 2005; pp. 1–97. Available online: https://www.cepf.net/sites/default/files/final.ehimalayas.ep_.pdf (accessed on 4 July 2022).
- Zhang, T.G.; Wang, W.C.; Gao, T.G.; An, B.S.; Yao, T.D. An integrative method for identifying potentially dangerous glacial lakes in the Himalayas. Sci. Total Environ. 2022, 806, 150442. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.J.; Lai, Z.P.; Zhou, S.Z.; Zeng, L.H. Timing of glacier fluctuations and trigger mechanisms in eastern Qinghai-Tibetan Plateau during the late Quaternary. Quat. Res. 2014, 81, 464–475. [Google Scholar] [CrossRef]
- Bolch, T.; Kulkarni, A.; Kaab, A.; Huggel, C.; Paul, F.; Cogley, J.G.; Frey, H.; Kargel, J.S.; Fujita, K.; Scheel, M.; et al. The state and fate of Himalayan glaciers. Science 2012, 336, 310–314. [Google Scholar] [CrossRef]
- Sgorbati, S.; Labra, M.; Grugni, E.; Barcaccia, G.; Galasso, G.; Boni, U.; Mucciarelli, M.; Citterio, S.; Iramategui, A.B.; Gonzales, L.V.; et al. A survey of genetic diversity and reproductive biology of Puya raimondii (Bromeliaceae), the endangered queen of the Andes. Plant Biol. 2004, 6, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Salerno, P.E.; Senaris, J.C.; Rojas-Runjaic, F.J.M.; Cannatella, D.C. Recent evolutionary history of Lost World endemics: Population genetics, species delimitation, and phylogeography of sky-island treefrogs. Mol. Phylogenet. Evol. 2015, 82, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Spurgin, L.G.; Illera, J.C.; Jorgensen, T.H.; Dawson, D.A.; Richardson, D.S. Genetic and phenotypic divergence in an island bird: Isolation by distance, by colonization or by adaptation? Mol. Ecol. 2014, 23, 1028–1039. [Google Scholar] [CrossRef] [PubMed]
- Hoeck, P.E.A.; Bollmer, J.L.; Parker, P.G.; Keller, L.F. Differentiation with drift: A spatio-temporal genetic analysis of Galapagos mockingbird populations (Mimus spp.). Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Beheregaray, L.B.; Ciofi, C.; Caccone, A.; Gibbs, J.P.; Powell, J.R. Genetic divergence, phylogeography and conservation units of giant tortoises from Santa Cruz and Pinzon, Galapagos Islands. Conserv. Genet. 2003, 4, 31–46. [Google Scholar] [CrossRef]
- Carson, H.L. Increased genetic variance after a population bottleneck. Trends Ecol. Evol. 1990, 5, 228–230. [Google Scholar] [CrossRef]
- Carson, H.L.; Templeton, A.R. Genetic revolutions in relation to speciation phenomena—The founding of new populations. Annu. Rev. Ecol. Syst. 1984, 15, 97–131. [Google Scholar] [CrossRef]
- Schaal, B.A.; Hayworth, D.A.; Olsen, K.M.; Rauscher, J.T.; Smith, W.A. Phylogeographic studies in plants: Problems and prospects. Mol. Ecol. 1998, 7, 465–474. [Google Scholar] [CrossRef]
- Higashi, H.; Sakaguchi, S.; Ikeda, H.; Isagi, Y.; Setoguchi, H. Multiple introgression events and range shifts in Schizocodon (Diapensiaceae) during the Pleistocene. Bot. J. Linn. Soc. 2013, 173, 46–63. [Google Scholar] [CrossRef]
- Gugger, P.F.; Sugita, S.; Cavender-Bares, J. Phylogeography of Douglas-fir based on mitochondrial and chloroplast DNA sequences: Testing hypotheses from the fossil record. Mol. Ecol. 2010, 19, 1877–1897. [Google Scholar] [CrossRef] [PubMed]
- Dixon, C.J.; Schonswetter, P.; Schneeweiss, G.M. Traces of ancient range shifts in a mountain plant group (Androsace halleri complex, Primulaceae). Mol. Ecol. 2007, 16, 3890–3901. [Google Scholar] [CrossRef]
- Chen, J.H.; Huang, Y.; Brachi, B.; Yun, Q.Z.; Zhang, W.; Lu, W.; Li, H.N.; Li, W.Q.; Sun, X.D.; Wang, G.Y.; et al. Genome-wide analysis of Cushion willow provides insights into alpine plant divergence in a biodiversity hotspot. Nat. Commun. 2019, 10, 5230. [Google Scholar] [CrossRef]
- Gao, Q.B.; Zhang, F.Q.; Xing, R.; Gornall, R.J.; Fu, P.C.; Li, Y.; Gengji, Z.M.; Chen, S.L. Phylogeographic study revealed microrefugia for an endemic species on the Qinghai-Tibetan Plateau: Rhodiola chrysanthemifolia (Crassulaceae). Plant Syst. Evol. 2016, 302, 1179–1193. [Google Scholar] [CrossRef]
- Rana, H.K.; Luo, D.; Rana, S.K.; Sun, H. Geological and climatic factors affect the population genetic connectivity in Mirabilis himalaica (Nyctaginaceae): Insight From phylogeography and dispersal corridors in the Himalaya-Hengduan biodiversity hotspot. Front. Plant Sci. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.K.; Luo, D.; Rana, H.K.; O’Neill, A.R.; Sun, H. Geoclimatic factors influence the population genetic connectivity of Incarvillea arguta (Bignoniaceae) in the Himalaya-Hengduan Mountains biodiversity hotspot. J. Syst. Evol. 2021, 59, 151–168. [Google Scholar] [CrossRef]
- Jia, Y.; Milne, R.I.; Zhu, J.; Gao, L.M.; Zhu, G.F.; Zhao, G.F.; Liu, J.; Li, Z.H. Evolutionary legacy of a forest plantation tree species (Pinus armandii): Implications for widespread afforestation. Evol. Appl. 2020, 13, 2646–2662. [Google Scholar] [CrossRef]
- Wang, Z.W.; Chen, S.T.; Nie, Z.L.; Zhang, J.W.; Zhou, Z.; Deng, T.; Sun, H. Climatic factors drive population divergence and demography: Insights based on the phylogeography of a riparian plant species endemic to the Hengduan Mountains and adjacent regions. PLoS ONE 2015, 10, e0145014. [Google Scholar] [CrossRef]
- Shepard, D.B.; Burbrink, F.T. Lineage diversification and historical demography of a sky island salamander, Plethodon ouachitae, from the Interior Highlands. Mol. Ecol. 2008, 17, 5315–5335. [Google Scholar] [CrossRef]
- Wiens, J.J.; Graham, C.H. Niche conservatism: Integrating evolution, ecology, and conservation biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef]
- Wiens, J.J. The role of morphological data in phylogeny reconstruction. Syst. Biol. 2004, 53, 653–661. [Google Scholar] [CrossRef]
- Hewitt, G.M. Some genetic consequences of ice ages, and their role in divergence and speciation. Biol. J. Linn. Soc. 1996, 58, 247–276. [Google Scholar] [CrossRef]
- Liang, Q.L.; Xu, X.T.; Mao, K.S.; Wang, M.C.; Wang, K.; Xi, Z.X.; Liu, J.Q. Shifts in plant distributions in response to climate warming in a biodiversity hotspot, the Hengduan Mountains. J. Biogeogr. 2018, 45, 1334–1344. [Google Scholar] [CrossRef]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.E.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction debt of high-mountain plants under twenty-first-century climate change. Nat. Clim. Chang. 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Lande, R. Risks of population extinction from demographic and environmental stochasticity and random catastrophes. Am. Nat. 1993, 142, 911–927. [Google Scholar] [CrossRef] [PubMed]
- Elsen, P.R.; Tingley, M.W. Global mountain topography and the fate of montane species under climate change. Nat. Clim. Chang. 2015, 5, 772–776. [Google Scholar] [CrossRef]
- Wang, Y.H.; Lu, L.; Wang, Y.H.; Wang, H. Evaluation and utilization of germplasm resources of Gaultheria L. in China. J. Yunnan Univ. Nat. Sci. Ed. 2013, 35, 378–389+410. [Google Scholar]
- Fave, M.J.; Johnson, R.A.; Cover, S.; Handschuh, S.; Metscher, B.D.; Muller, G.B.; Gopalan, S.; Abouheif, E. Past climate change on Sky Islands drives novelty in a core developmental gene network and its phenotype. BMC Evol. Biol. 2015, 15, 183. [Google Scholar] [CrossRef]
- Boyd, A. Morphological analysis of Sky Island populations of Macromeria viridiflora (Boraginaceae). Syst. Bot. 2002, 27, 116–126. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Sets | Groups | HS | HT | GST | NST | Hd | π | r2 |
|---|---|---|---|---|---|---|---|---|
| Grouping set I based on geographical distance | CH | 0.479 (0.043) | 0.501 (0.045) | 0.044 (NC) | 0.107 (NC) | 0.493 | 0.001 | - |
| EH + WHM + EHM | 0.297 (0.081) | 0.801 (0.063) | 0.630 (0.101) | 0.706 (0.092) | 0.786 | 0.002 | - | |
| Grouping set II based on geographical distance | CH | 0.479 (0.043) | 0.501 (0.045) | 0.044 (NC) | 0.107 (NC) | 0.493 | 0.001 | - |
| EH + WHM | 0.398 (0.092) | 0.758 (0.081) | 0.475 (0.109) | 0.600 (0.104) | 0.754 | 0.002 | - | |
| EHM | 0.044 (0.044) | 0.667 (0.250) | 0.933 (NC) | 0.968 (NC) | 0.475 | 0.002 | - | |
| Grouping set III based on geographical trend of mountains | CH + EH | 0.440 (0.110) | 0.773 (0.083) | 0.431 (0.162) | 0.909 (0.033) | 0.721 | 0.007 | - |
| WHM + EHM | 0.284 (0.083) | 0.747 (0.076) | 0.620 (0.112) | 0.671 (0.107) | 0.733 | 0.002 | - | |
| All populations | Overall | 0.326 (0.065) | 0.860 (0.041) | 0.620 (0.075) | 0.890 (0.032) | 0.845 | 0.007 | 0.086 * |
| Group Sets | Source of Variation | Sum of Squares (SS) | Variance Components | Percentage of Variation | Fixation Index |
|---|---|---|---|---|---|
| Grouping set I CH, EH + WHM + EHM | Among groups | 399.29 | 3.562 Va | 85.30 | FCT = 0.853 * |
| Among populations within groups | 100.06 | 0.398 Vb | 9.53 | FSC = 0.648 * | |
| Within populations | 54.68 | 0.216 Vc | 5.17 | FST = 0.948 * | |
| Grouping set II CH, EH + WHM, EHM | Among groups | 356.57 | 2.338 Va | 80.55 | FCT = 0.806 * |
| Among populations within groups | 82.79 | 0.348 Vb | 12.00 | FSC = 0.617 * | |
| Within populations | 54.67 | 0.216 Vc | 7.44 | FST = 0.926 * | |
| Grouping set III CH + EH, WHM + EHM | Among groups | 195.08 | 1.495 Va | 55.30 | FCT = 0.553 * |
| Among populations within groups | 244.27 | 0.992 Vb | 36.71 | FSC = 0.821 * | |
| Within populations | 54.67 | 0.216 Vc | 7.99 | FST = 0.920 * |
| Group Sets | Groups | Neutrality Tests | Mismatch Analysis | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| D | p Value | Fu’s FS | p Value | SSD | p Value | HRag | p Value | Tau | ||
| Grouping set I based on geographical distance | CH | −1.12 | 0.092 | −2.689 | 0.054 | 0.009 | 0.060 | 0.141 | 0.180 | 0.680 |
| EH + WHM + EHM | −0.507 | 0.320 | −4.952 | 0.060 | 0.006 * | 0.030 | 0.078 * | 0.010 | 1.463 | |
| Grouping set II based on geographical distance | CH | −1.12 | 0.092 | −2.689 | 0.054 | 0.009 | 0.060 | 0.141 | 0.180 | 0.680 |
| EH + WHM | −0.411 | 0.348 | −4.527 | 0.055 | 0.012 * | 0.010 | 0.076 * | 0.030 | 1.469 | |
| EHM | 0.850 | 0.673 | 1.875 | 0.919 | 0.185 * | 0.040 | 0.592 * | 0.010 | 1.984 | |
| Grouping set III based on geographical trend of mountains | CH + EH | 1.100 | 0.799 | 2.596 | 0.908 | 0.112 | 0.050 | 0.119 | 0.280 | 11.029 |
| WHM + EHM | −0.158 | 0.438 | −2.542 | 0.168 | 0.006 * | 0.030 | 0.089 * | 0.000 | 1.240 | |
| All populations | Overall | 0.185 | 0.581 | −1.078 | 0.418 | 0.043 | 0.400 | 0.047 | 0.620 | 9.771 |
| Variable | Description | Contribution (%) | Permutation Importance |
|---|---|---|---|
| Bio2 | Mean diurnal range | 5.6 | 6.5 |
| Bio3 | Isothermality | 14.2 | 8.1 |
| Bio6 | Minimum temperature of coldest month | 27.3 | 26.5 |
| Bio7 | Temperature annual range | 44.5 | 55.5 |
| Bio13 | Precipitation of the wettest month | 3.8 | 0.8 |
| Bio14 | Precipitation of the driest month | 4.5 | 2.6 |
| Bio15 | Coefficient of variation of precipitation seasonality | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Cheng, X.; Fritsch, P.W.; Li, Y.; Yang, S.; Lu, L. Genetic Variation in Gaultheria nummularioides (Ericaceae: Gaultherieae) from the Sky Islands of the Himalaya-Hengduan Mountains. Diversity 2022, 14, 652. https://doi.org/10.3390/d14080652
Zhang J, Cheng X, Fritsch PW, Li Y, Yang S, Lu L. Genetic Variation in Gaultheria nummularioides (Ericaceae: Gaultherieae) from the Sky Islands of the Himalaya-Hengduan Mountains. Diversity. 2022; 14(8):652. https://doi.org/10.3390/d14080652
Chicago/Turabian StyleZhang, Jing, Xiaojuan Cheng, Peter W. Fritsch, Yirong Li, Shuda Yang, and Lu Lu. 2022. "Genetic Variation in Gaultheria nummularioides (Ericaceae: Gaultherieae) from the Sky Islands of the Himalaya-Hengduan Mountains" Diversity 14, no. 8: 652. https://doi.org/10.3390/d14080652
APA StyleZhang, J., Cheng, X., Fritsch, P. W., Li, Y., Yang, S., & Lu, L. (2022). Genetic Variation in Gaultheria nummularioides (Ericaceae: Gaultherieae) from the Sky Islands of the Himalaya-Hengduan Mountains. Diversity, 14(8), 652. https://doi.org/10.3390/d14080652