
Evaluation of Climate Change Impacts on the Global Distribution of the Calliphorid Fly Chrysomya albiceps Using GIS

 ,
, 
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
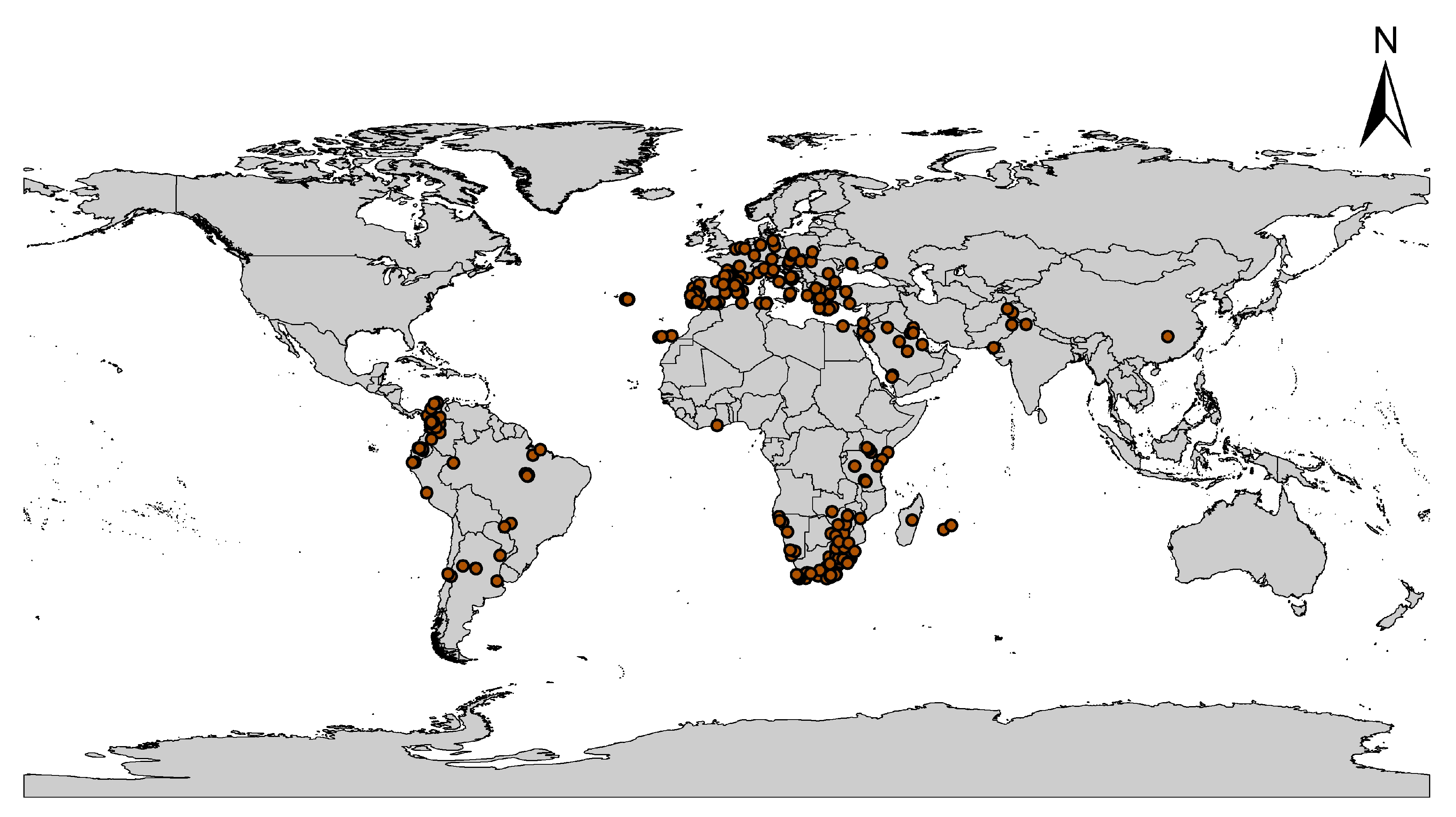
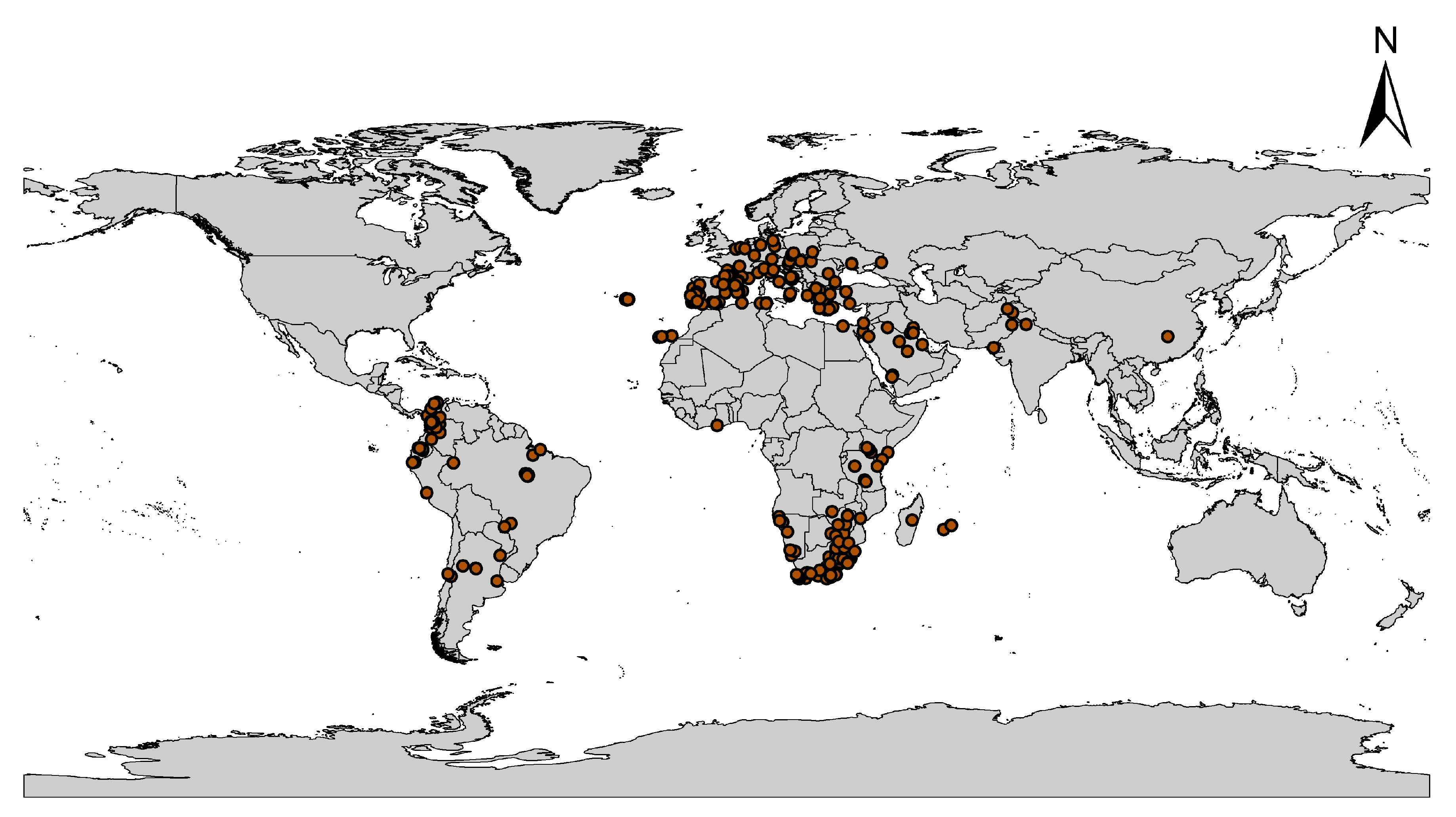
2.1. Global Occurrence Data of C. albiceps
2.2. Environmental Variables
2.3. Habitat Suitability Modeling
2.4. Model Interpretation and Evaluation
3. Results
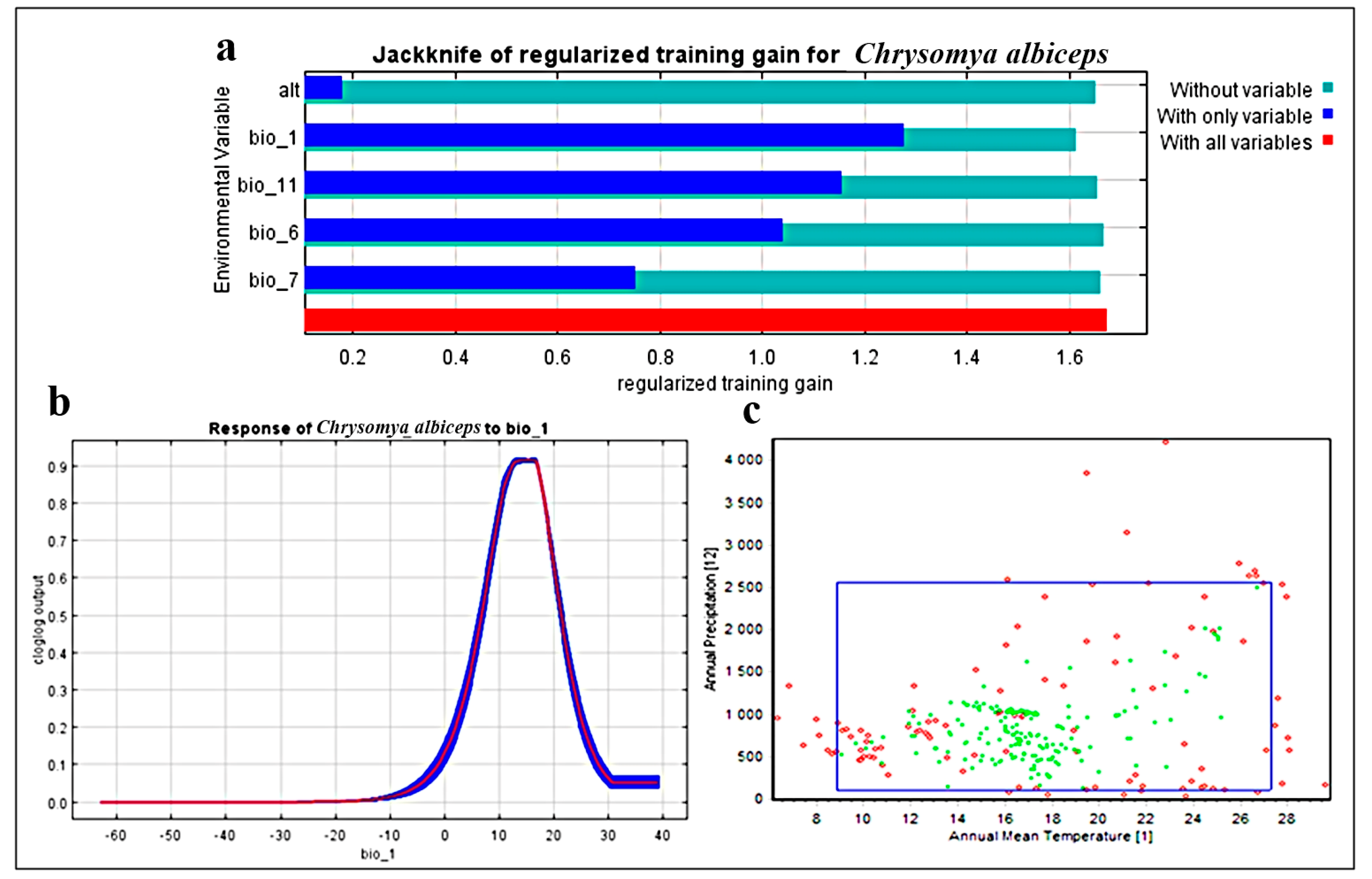
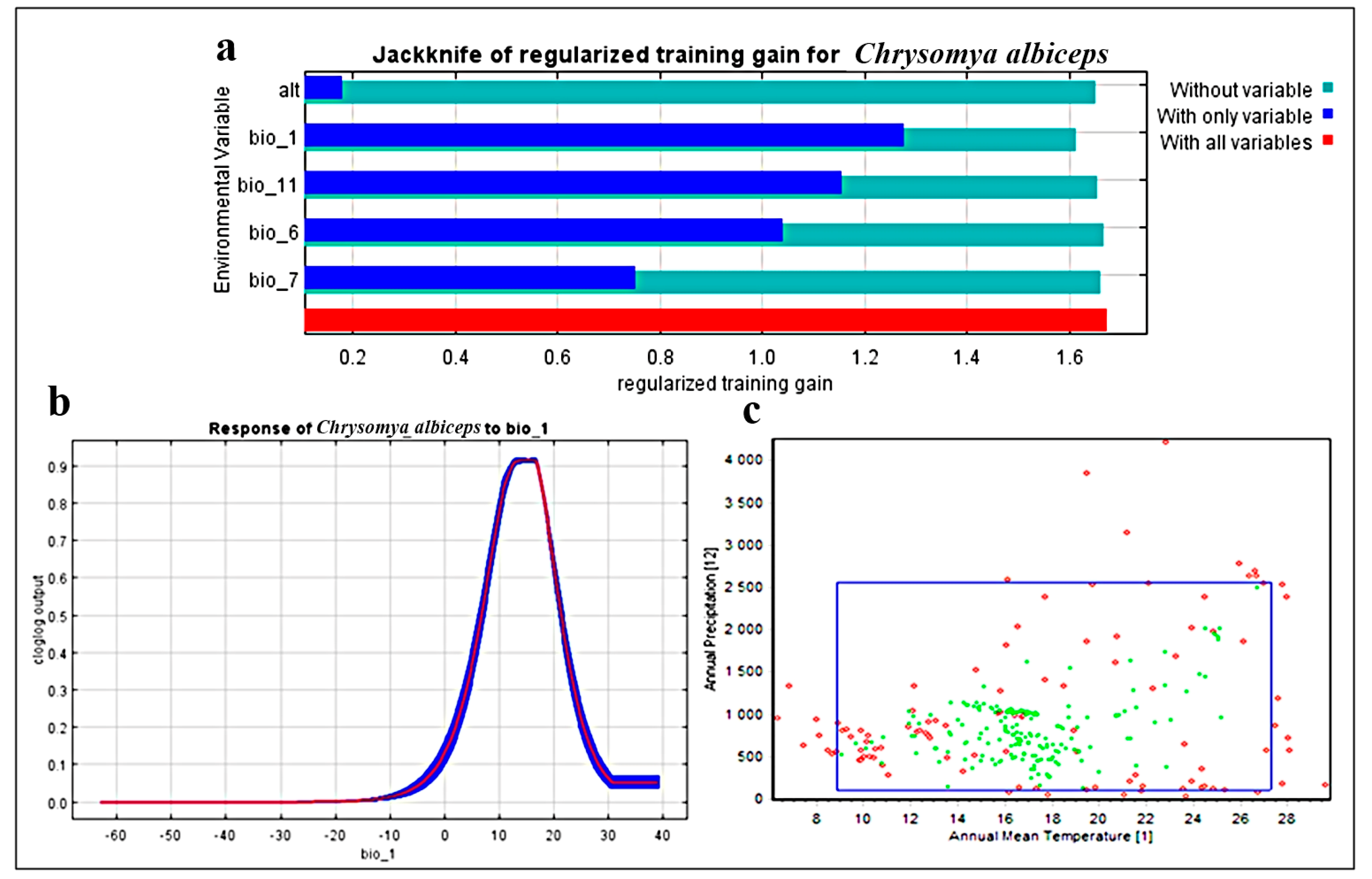
3.1. Model Performance and Effects of Environmental Variables
3.2. Two-Dimensional Niche Analysis
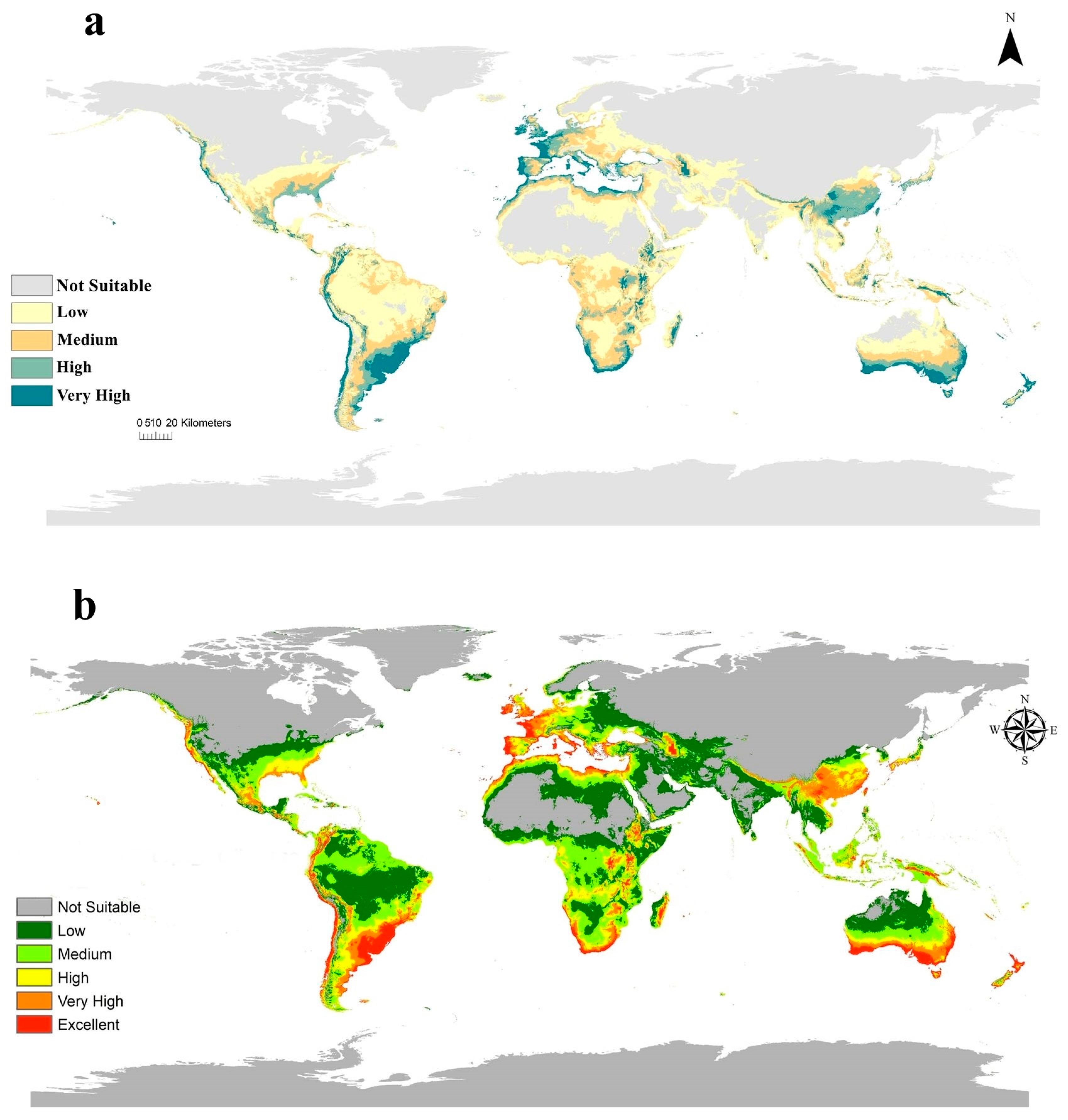
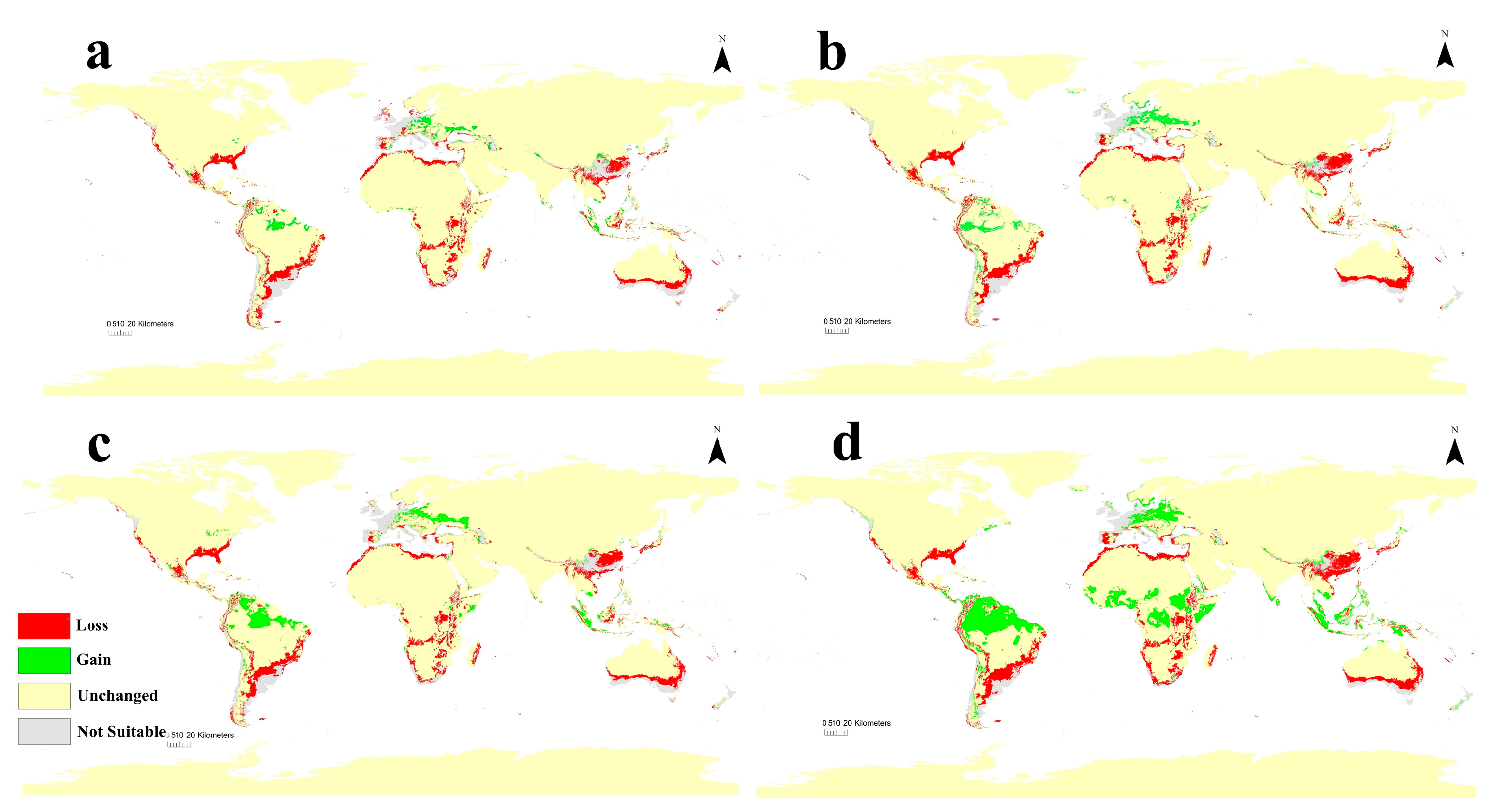
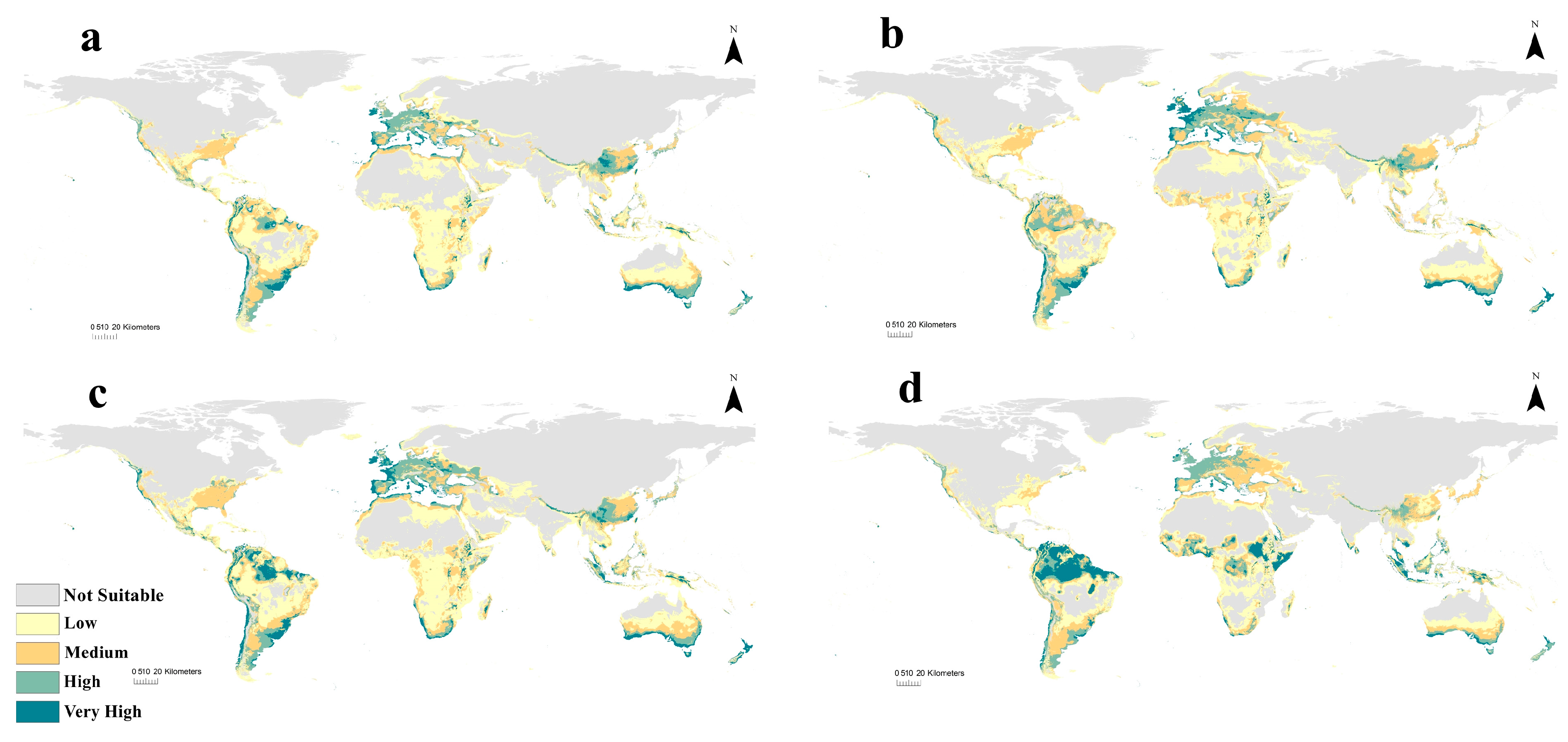
3.3. Current Potential Distribution of C. albiceps
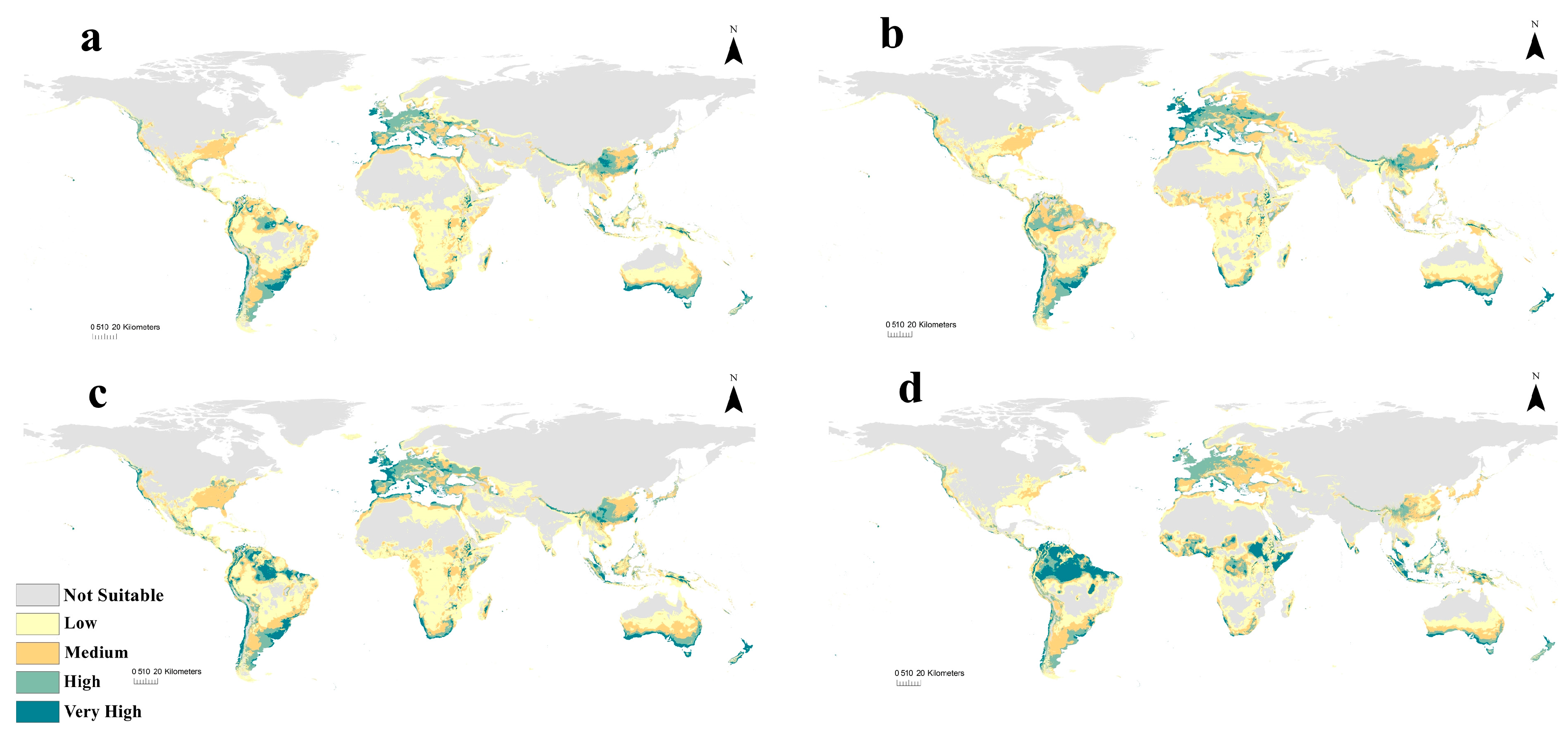
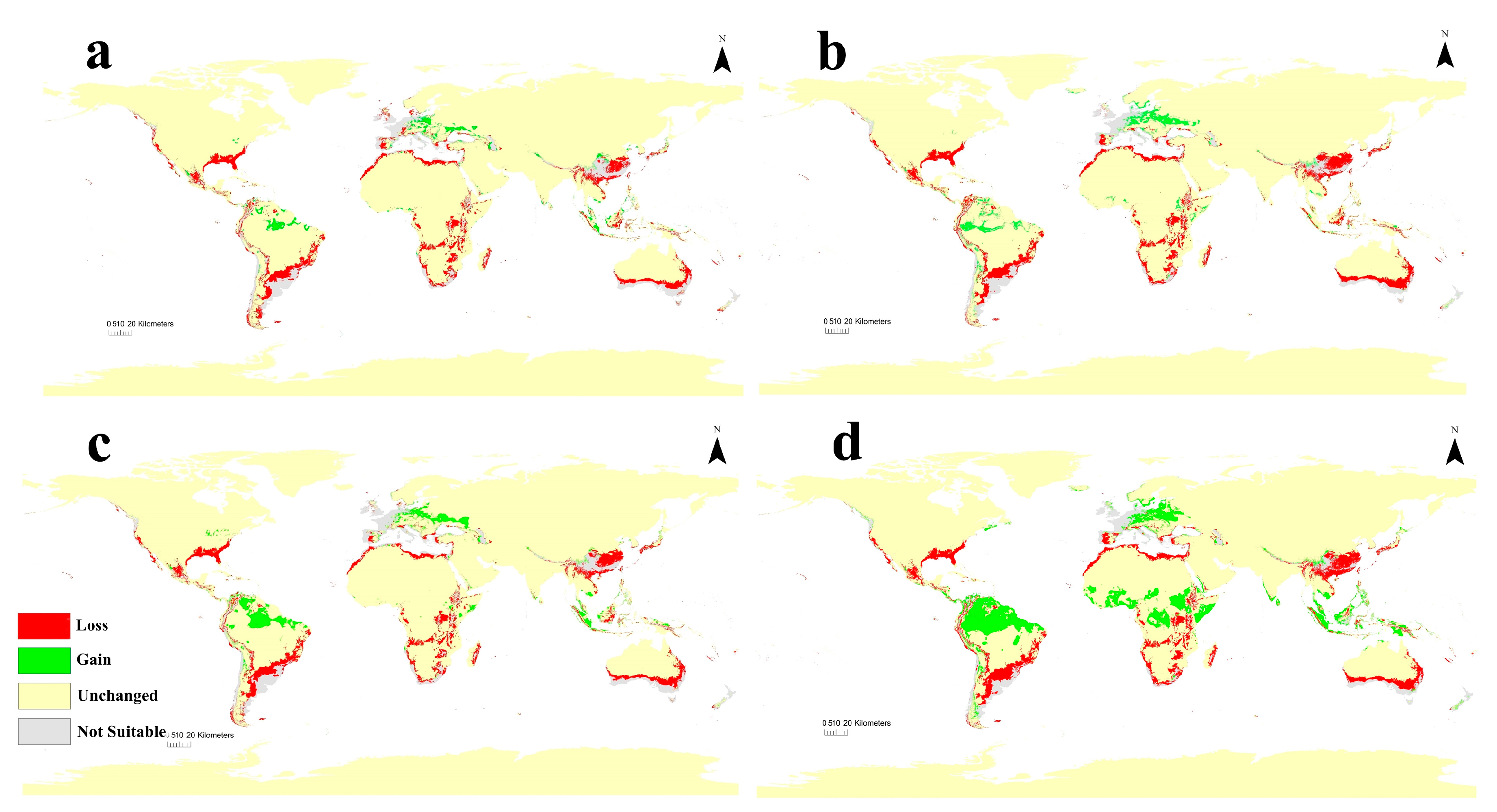
3.4. Future Potential Distribution of C. albiceps
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fei, S.; Desprez, J.M.; Potter, K.M.; Jo, I.; Knott, J.A.; Oswalt, C.M. Divergence of species responses to climate change. Sci. Adv. 2017, 3, e1603055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letcher, T.M. Why do we have global warming? In Managing Global Warming; Academic Press: Cambridge, MA, USA, 2019; pp. 3–15. [Google Scholar]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Pol, G.; Crotta, M.; Taylor, R.A. Modelling the temperature suitability for the risk of West Nile Virus establishment in European Culex pipiens populations. Transbound. Emerg. Dis. 2022, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, L.J.; Hughes, L.; Pitman, A. Why is the choice of future climate scenarios for species distribution modelling important? Ecol. Lett. 2008, 11, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Van Der Putten, W.H.; Macel, M.; Visser, M.E. Predicting species distribution and abundance responses to climate change: Why it is essential to include biotic interactions across trophic levels. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2025–2034. [Google Scholar] [CrossRef]
- Hosni, E.M.; Kenawy, M.A.; Nasser, M.G.; Al-Ashaal, S.A.; Rady, M.H. A Brief Review of Myiasis with Special Notes on the Blow Flies’ Producing Myiasis (F.: Calliphoridae). Egypt. Acad. J. Biol. Sci. E Med. Entomol. Parasitol. 2019, 11, 25–32. [Google Scholar] [CrossRef]
- Verves, Y.G. The new faunistic data on Calliphoridae and Sarcophagidae (Diptera) of the Republic of Seychelles. Phelsuma 2007, 15, 71–81. [Google Scholar]
- Nasser, M.G.; Hosni, E.M.; Kenawy, M.A.; Alharbi, S.A.; Almoallim, H.S.; Rady, M.H.; Merdan, B.A.; Pont, A.C.; Al-Ashaal, S.A. Evolutionary profile of the family Calliphoridae, with notes on the origin of myiasis. Saudi J. Biol. Sci. 2021, 28, 2056–2066. [Google Scholar] [CrossRef]
- Alahmed, A.M.; Nasser, M.G.; Sallam, M.F.; Dawah, H.; Kheir, S.; AlAshaal, S.A. Two new records of flies causing myiasis from Saudi Arabia with a survey of flies parasitizing goats and sheep in Jazan Region. Trop. Biomed. 2020, 37, 499–512. [Google Scholar]
- Greenberg, B. Flies and Diseases. Vol. I. Ecology, Classification and Biotic Associations; Princeton University Press: Precenton, NJ, USA, 1971; p. 856. [Google Scholar]
- Greenberg, B. Flies and Diseases. Vol. II. Biology and Disease Transmission; Princeton University Press: Precenton, NJ, USA, 1973; p. 447. [Google Scholar]
- Zumpt, F. Myiasis in Man and Animals in the Old World. A Textbook for Physicians, Veterinarians and Zoologists; Butterworths: London, UK, 1965; p. 267. [Google Scholar]
- Ivorra, T.; Martínez-Sánchez, A.; Rojo, S. Coexistence and intraguild competition of Chrysomya albiceps and Lucilia sericata larvae: Case reports and experimental studies applied to forensic entomology. Acta Trop. 2022, 226, 106233. [Google Scholar] [CrossRef]
- Iloba, B.N.; Odigie, O.O. Arthropod succession on buried carrion of the African giant. Afr. Sci. 2021, 7, 53–59. [Google Scholar]
- Carmo, R.F.; Barbosa, T.M.; Torris, A.F.; Bezerra, M.-A.S.; Vasconcelos, S.D. Diversity of sarcosaprophagous Diptera (Calliphoridae, Sarcophagidae) in organic and conventional mango plantations in the Brazilian semi-arid region. Rev. Bras. De Èntomol. 2021, 65, 1–5. [Google Scholar] [CrossRef]
- Dag, A.; Gazit, S. Mango pollinators in Israel. J. Appl. Hortic. 2000, 2, 39–43. [Google Scholar] [CrossRef]
- Siqueira, M.F.D.; Peterson, A.T. Consequences of global climate change for geographic distributions of cerrado tree species. Biota Neotrop. 2003, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Gilman, S.E.; Urban, M.C.; Tewksbury, J.; Gilchrist, G.W.; Holt, R.D. A framework for community interactions under climate change. Trends Ecol. Evol. 2010, 25, 325–331. [Google Scholar] [CrossRef]
- Cox, J.S.H. The Role of Geographic Information Systems and Spatial Analysis in Area-Wide Vector Control Programm. In Area-wide Control of Insect Pests; Springer: Dordrecht, The Netherlands, 2007; pp. 199–209. [Google Scholar]
- Naeem, M.; Alahmed, A.M.; Kheir, S.M.; Sallam, M.F. Spatial distribution modeling of Stegomyia aegypti and Culex tri-taeniorhynchus (Diptera: Culicidae) in Al-bahah Province, Kingdom of Saudi Arabia. Trop. Biomed. 2016, 33, 295–310. [Google Scholar]
- Hosni, E.M.; Nasser, M.; Al-Ashaal, S.; Rady, M.H.; Kenawy, M.A. Modeling current and future global distribution of Chrysomya bezziana under changing climate. Sci. Rep. 2020, 10, 4947. [Google Scholar] [CrossRef]
- Abou-Shaara, H.; Alashaal, S.A.; Hosni, E.M.; Nasser, M.G.; Ansari, M.J.; Alharbi, S.A. Modeling the Invasion of the Large Hive Beetle, Oplostomusfuligineus, into North Africa and South Europe under a Changing Climate. Insects 2021, 12, 275. [Google Scholar] [CrossRef]
- Escobar, L.E.; Lira-Noriega, A.; Medina-Vogel, G.; Peterson, A.T. Potential for spread of the white-nose fungus (Pseudogym-noascus destructans) in the Americas: Use of Maxent and Niche A to assure strict model transference. Geospat. Health 2014, 9, 221–229. [Google Scholar] [CrossRef]
- Hosni, E.M.; Nasser, M.; Al-Khalaf, A.A.; Al-Shammery, K.A.; Al-Ashaal, S.; Soliman, D. Invasion of the Land of Samurai: Potential Spread of Old-World Screwworm to Japan under Climate Change. Diversity 2022, 14, 99. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 20 March 2020).
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.H.; Graham, C.P.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Kessler, W.H.; Ganser, C.; Glass, G.E. Modeling the Distribution of Medically Important Tick Species in Florida. Insects 2019, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Mulieri, P.R.; Patitucci, L.D. Using ecological niche models to describe the geographical distribution of the myiasis-causing Cochliomyia hominivorax (Diptera: Calliphoridae) in southern South America. Parasitol. Res. 2019, 118, 1077–1086. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Kondratev, K.I.; Kondrat’ev, K.I.; Kondratyev, K.Y.; Kondrat’ev, K.J.; Krapivin, V.F.; Varotsos, C.; Barōtsos, K. Global Carbon Cycle and Climate Change; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Liu, Y.; Shi, J. Predicting the Potential Global Geographical Distribution of Two Icerya Species under Climate Change. Forests 2020, 11, 684. [Google Scholar] [CrossRef]
- Sax, D.F.; Early, R.; Bellemare, J. Niche syndromes, species extinction risks, and management under climate change. Trends Ecol. Evol. 2013, 28, 517–523. [Google Scholar] [CrossRef] [Green Version]
- Hannah, L.; Thomas, E.L. Biodiversity and Climate Change. In Climate Change and Biodiversity; Yale University: London, UK, 2004; Volume 3. [Google Scholar]
- Alotaibi, F.; Alkuriji, M.; AlReshaidan, S.; Alajmi, R.; Metwally, D.M.; Almutairi, B.; Alorf, M.; Haddadi, R.; Ahmed, A. Body Size and Cuticular Hydrocarbons as Larval Age Indicators in the Forensic Blow Fly, Chrysomya albiceps (Diptera: Calliphoridae). J. Med Èntomol. 2021, 58, 1048–1055. [Google Scholar] [CrossRef]
- Naeem, M.; Yuan, X.; Huang, J.; An, J. Habitat suitability for the invasion of Bombus terrestris in East Asian countries: A case study of spatial overlap with local Chinese bumblebees. Sci. Rep. 2018, 8, 11035. [Google Scholar] [CrossRef] [PubMed]
- Hosni, E.M.; Al-Khalaf, A.A.; Nasser, M.G.; Abou-Shaara, H.F.; Radwan, M.H. Modeling the Potential Global Distribution of Honeybee Pest, Galleria mellonella under Changing Climate. Insects 2022, 13, 484. [Google Scholar] [CrossRef] [PubMed]
- Wallman, J.; Donnellan, S. The utility of mitochondrial DNA sequences for the identification of forensically important blowflies (Diptera: Calliphoridae) in southeastern Australia. Forensic Sci. Int. 2001, 120, 60–67. [Google Scholar] [CrossRef]
- Kourgialas, N.N.; Dokou, Z. Water management and salinity adaptation approaches of Avocado trees: A review for hot-summer Mediterranean climate. Agric. Water Manag. 2021, 252, 106923. [Google Scholar] [CrossRef]
- Daniele, B.-C.; Barbara, S.; Isabel, B.; Alberto, G. Economic risk assessment of the quality labels and productive efficiency strategies in Spanish extensive sheep farms. Agric. Syst. 2021, 191, 103169. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosni, E.M.; Al-Khalaf, A.A.; Naguib, R.M.; Afify, A.E.; Abdalgawad, A.A.; Faltas, E.M.; Hassan, M.A.; Mahmoud, M.A.; Naeem, O.M.; Hassan, Y.M.; et al. Evaluation of Climate Change Impacts on the Global Distribution of the Calliphorid Fly Chrysomya albiceps Using GIS. Diversity 2022, 14, 578. https://doi.org/10.3390/d14070578
Hosni EM, Al-Khalaf AA, Naguib RM, Afify AE, Abdalgawad AA, Faltas EM, Hassan MA, Mahmoud MA, Naeem OM, Hassan YM, et al. Evaluation of Climate Change Impacts on the Global Distribution of the Calliphorid Fly Chrysomya albiceps Using GIS. Diversity. 2022; 14(7):578. https://doi.org/10.3390/d14070578
Chicago/Turabian StyleHosni, Eslam M., Areej A. Al-Khalaf, Randa M. Naguib, Abdalrahman E. Afify, Ashraf A. Abdalgawad, Ehab M. Faltas, Mohamed A. Hassan, Mohamed A. Mahmoud, Omar M. Naeem, Yossef M. Hassan, and et al. 2022. "Evaluation of Climate Change Impacts on the Global Distribution of the Calliphorid Fly Chrysomya albiceps Using GIS" Diversity 14, no. 7: 578. https://doi.org/10.3390/d14070578
APA StyleHosni, E. M., Al-Khalaf, A. A., Naguib, R. M., Afify, A. E., Abdalgawad, A. A., Faltas, E. M., Hassan, M. A., Mahmoud, M. A., Naeem, O. M., Hassan, Y. M., & Nasser, M. G. (2022). Evaluation of Climate Change Impacts on the Global Distribution of the Calliphorid Fly Chrysomya albiceps Using GIS. Diversity, 14(7), 578. https://doi.org/10.3390/d14070578