
The Effects of Different Management Methods on Restored Grasslands in Potential Temperate Forest Zones

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- How close are the grasslands developed in the shrub-cut areas to natural habitats?
- Is mowing or grazing more effective in reducing shrubs?
- Which is the best management method resulting in greater biodiversity? On what level are they useful for grassland management, and how much livestock can they keep?
2. Materials and Methods
- Systematic mowing until 2013. Mosaic-like vegetation, in which Festuco ovinae-Nardetum Dostál 1933 and Festucetum rubrae-Cynosuretum Tx. 1940 patches are the most common.
- Shrub was cut. Festucetum rubrae-Cynosuretum Tx. 1940 typicum patches are the most common, but Helictotron pubescens facies is also present. After shrub cutting, the area was abandoned.
- Shrub was cut. Festucetum rubrae-Cynosuretum Tx. 1940 is the most common taxa. In 2013, the area was divided to two parts (IIIa and IIIb).
- IIIa
- Intensive grazing (3–4 animals per hectare or stocking density, which was 1.3 animal unit (AU)/ha) was conducted with Carpatian Braunvieh (Borzderes) cattle and Ratzka sheep.
- IIIb
- The grazing was in extensive utilization, but stocking density was 0.5 AU/ha less intensive, due to periodic, less frequent use of the area.
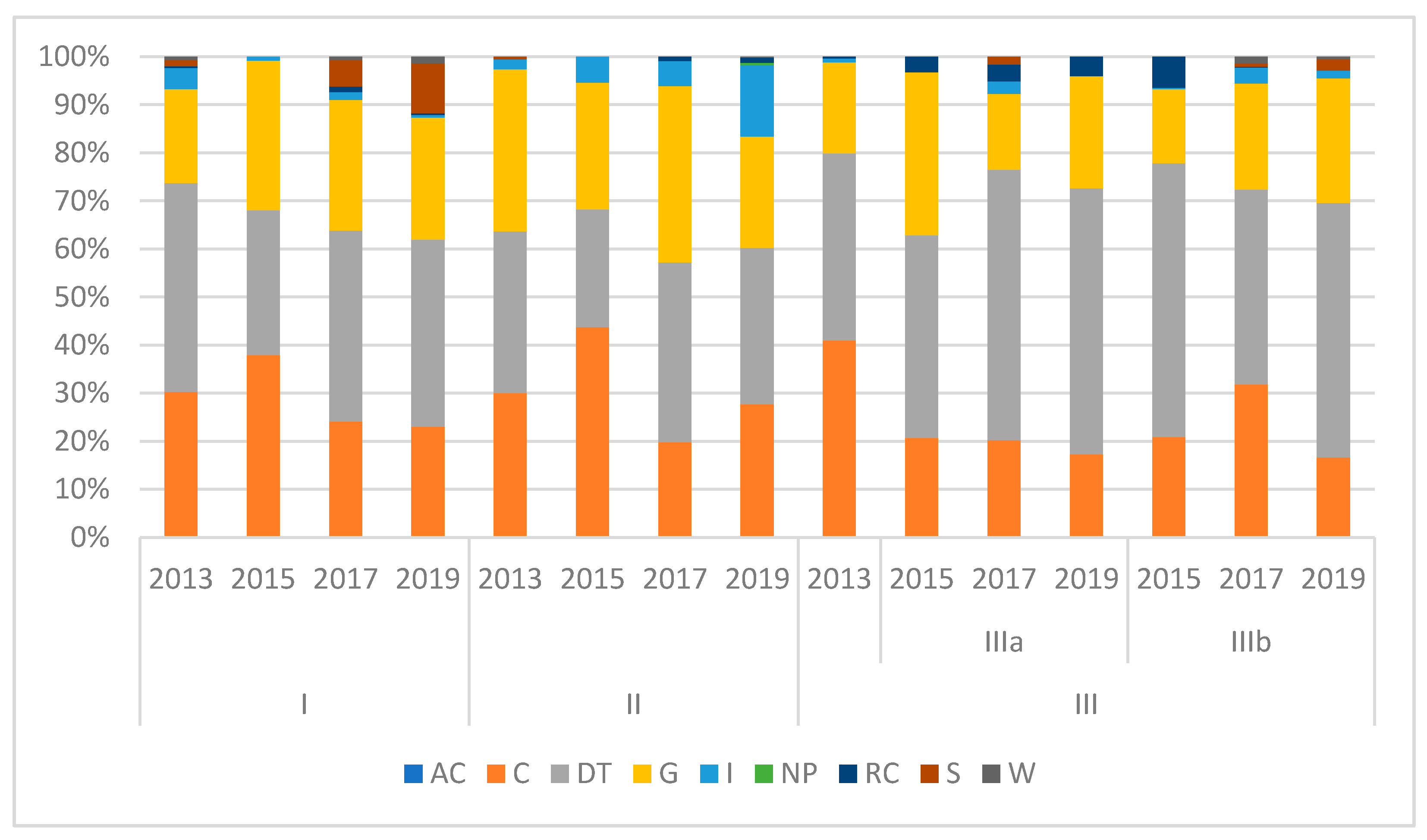
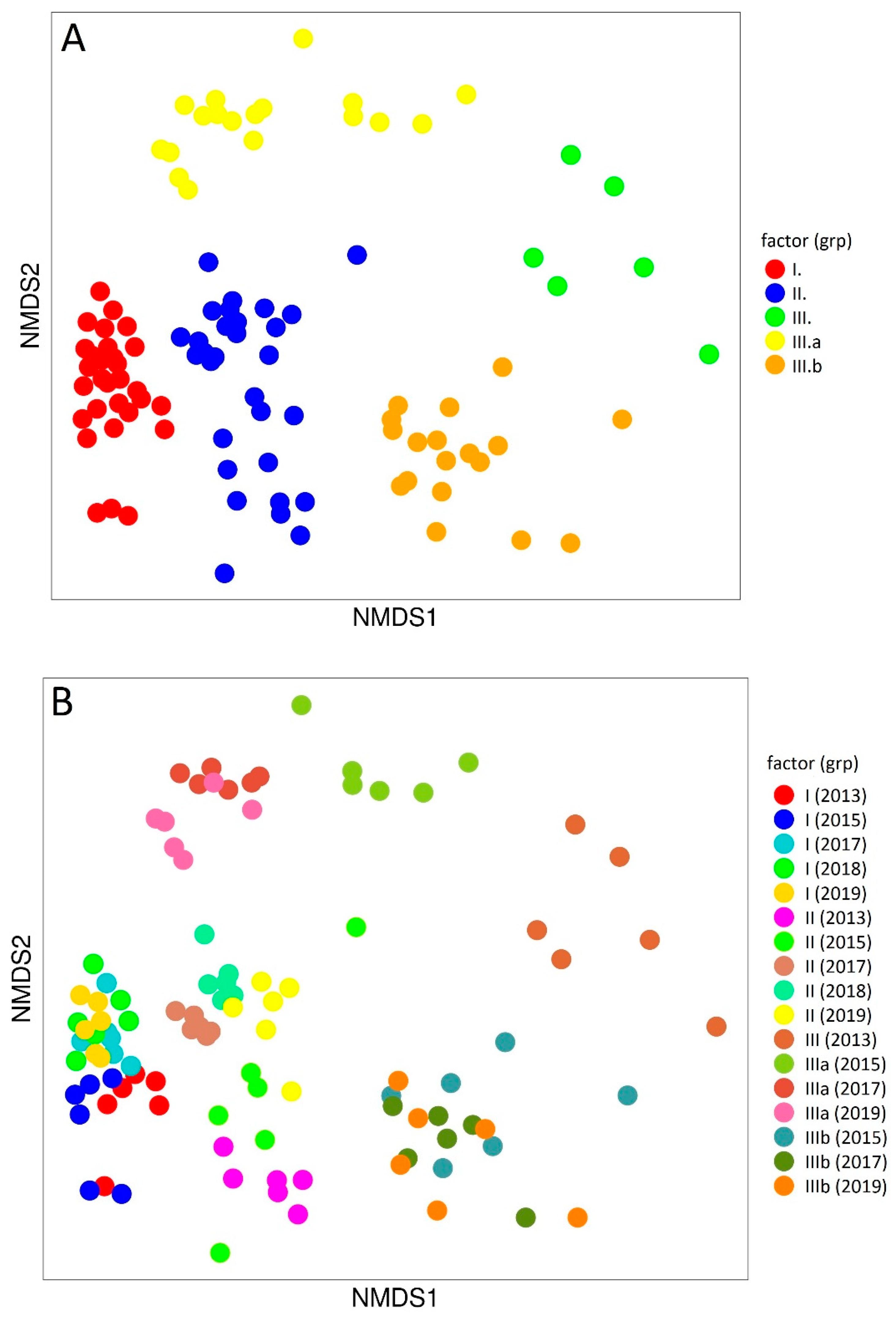
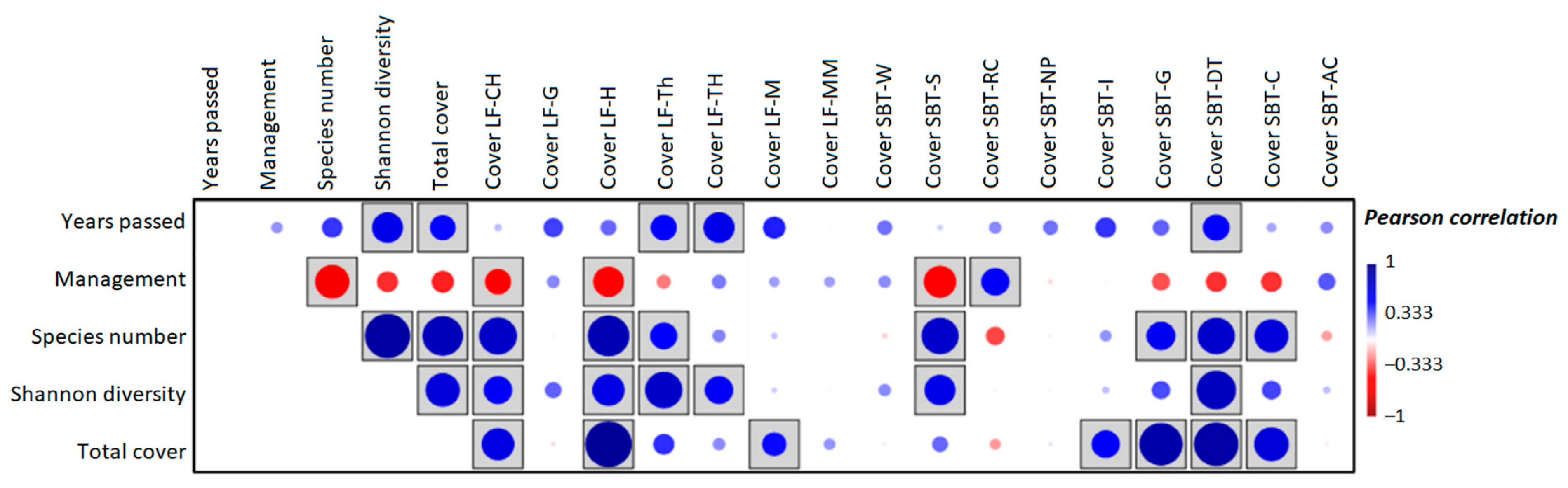
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pärtel, M.; Bruun, H.H.; Sammul, M. Biodiversity in temperate European grasslands: Origin and conservation. In Integrating Efficient Grassland Farming and Biodiversity, Proceedings of the 13th International Occasional Symposium of the European Grassland Federation, Tartu, Estonia, 29–31 August 2005; European Grassland Federation: Zurich, Switzerland, 2005; Volume 10, pp. 1–14. [Google Scholar]
- Owen, P. (Ed.) LIFE and Europe’s Grasslands: Restoring a Forgotten Habitat; European Commission, Environment Directorate-General: New York, NY, USA, 2008; 56p. [Google Scholar]
- Valkó, O.; Zmihorski, M.; Biurrun, I.; Loos, J.; Labadessa, R.; Venn, S. Ecology and conservation of steppes and semi-natural grasslands. Hacquetia 2016, 15, 5–14. [Google Scholar] [CrossRef]
- Kricsfalusy, V.V. Mountain grasslands of high conservation value in the Eastern Carpathians: Syntaxonomy, biodiversity, protection and management. Thaiszia J. Bot. 2013, 23, 67–112. [Google Scholar]
- Baráth, N.; Penksza, K. A Szénások Európa diplomás terület tájváltozásának és természeti állapotának vizsgálata, különös tekintettel a gyepekre. Tájökológiai Lapok 2012, 10, 361–370. [Google Scholar]
- Erdős, L.; Cserhalmi, D.; Bátori, Z.; Kiss, T.; Morschhauser, T.; Benyhe, B.; Dénes, A. Shrub encroachment in a wooded-steppemosaic: Combining GIS methods with landscape historical analysis. Appl. Ecol. Environ. Res. 2013, 11, 371–384. [Google Scholar] [CrossRef]
- Erdős, L.; Bátori, Z.; Tölgyesi, C.; Körmöczi, L. The moving split window (MSW) analysis in vegetation science—An overview. Appl. Ecol. Environ. Res. 2014, 12, 787–805. [Google Scholar] [CrossRef]
- Erdős, L.; Tölgyesi, C.; Dénes, A.; Darányi, N.; Fodor, A.; Bátori, Z.; Tolnay, D. Comparative analysis of the natural and semi-natural plant communities of Mt Nagy and other parts of the Villány Mts (south Hungary). Thaiszia J. Bot. 2014, 24, 1–21. [Google Scholar]
- Török, P.; Arany, A.; Prommer, M.; Valkó, O.; Balogh, A.; Vida, E.; Tóthmérész, B.; Matus, G. Újrakezdett kezelés hatása fokozottan védett kékperjés láprét fitomasszájára, faj- és virággazdagságára. Természetvédelmi Közlemények 2007, 13, 187–198. [Google Scholar]
- Török, P.; Valkó, O.; Deák, B.; Kelemen, A.; Tóthmérész, B. Traditional cattle grazing in a mosaic alkali landscape: Effects on grassland biodiversity along a moisture gradient. PLoS ONE 2014, 9, e97095. [Google Scholar] [CrossRef]
- Valkó, O.; Török, P.; Matus, G.; Tóthmérész, B. Is regular mowing the most appropriate and cost-effective management maintaining diversity and biomass of target forbs in mountain hay meadows? Flora 2012, 207, 303–309. [Google Scholar] [CrossRef]
- Ausden, M. Habitat Management for Conservation; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Jones-Walters, L.M. A new approach to the management of chalk grassland with particular reference to the integration of conservation measures for invertebrates. In Calcareous Grasslands—Ecology and Management; Hillier, S.H., Walton, D.W.H., Wells, D.A., Eds.; Bluntisham Books: Huntingdon, UK, 1990; pp. 67–73. [Google Scholar]
- Ward, L.K. Management of grassland-scrub mosaics. In Calcareous Grasslands—Ecology and Management; Hillier, S.H., Walton, D.W.H., Wells, D.A., Eds.; Bluntisham Books: Huntingdon, UK, 1990; pp. 134–139. [Google Scholar]
- Marriott, C.A.; Fothergill, M.; Jeangros, B.; Scotton, M.; Louault, F. Long-term impacts of extensification of grassland managementon biodiversity and productivity in upland areas. A review. Agronomie 2004, 24, 447–462. [Google Scholar] [CrossRef]
- Archer, S.; Schimel, D.S.; Holland, E.A. Mechanisms of shrubland expansion: Land use, climate or CO2. Clim. Chang. 1995, 29, 91–99. [Google Scholar] [CrossRef]
- Callaway, R.M.; Davis, F.W. Recruitment of Quercusagrifolia in central California: The importance of shrub dominated patches. J. Veg. Sci. 1998, 9, 647–656. [Google Scholar] [CrossRef]
- Barnes, P.W.; Archer, S. Tree-shrub interactions in asubtropical savanna parkland: Competition or facilitation? J. Veg. Sci. 1999, 10, 525–536. [Google Scholar] [CrossRef]
- Rousset, O.; Lepart, J. Shrub facilitation of Quercushumilis regeneration in succession on calcareous grasslands. J. Veg. Sci. 1999, 10, 493–502. [Google Scholar] [CrossRef]
- Michielsen, M.; Szemán, L.; Fenesi, A.; Nijs, I.; Ruprecht, E. Resprouting of woody species encroaching temperate European grasslands after cutting and burning. Appl. Veg. Sci. 2017, 20, 388–396. [Google Scholar] [CrossRef]
- Hmielowski, T.L.; Robertson, K.M.; Platt, W.J. Influence of season and method of topkill on resprouting characteristics and biomass of Quercus nigra saplings from a southeastern U.S. pine-grassland ecosystem. Plant Ecol. 2014, 215, 1221–1231. [Google Scholar] [CrossRef]
- Baráz, C. (Ed.) A Mátrai Tájvédelmi Körzet—Heves és Nógrád határán; Bükki Nemzeti Park Igazgatóság: Eger, Hungary, 2001; pp. 56–78. [Google Scholar]
- Baráz, C.; Schmotzer, A. (Eds.) A Bükki Nemzeti Park Igazgatóság Működési Területe; Bükki Nemzeti Park Igazgatóság: Eger, Hungary, 2010. [Google Scholar]
- Simberloff, D. A succession of paradigms in ecology: Essentialism tomaterialism and probablism. In Conceptual Issues in Ecology; Saarinen, E., Ed.; Reidel (Kluwer): Boston, MA, USA, 1982; pp. 63–99. [Google Scholar]
- White, P.S. Pattern, process, and natural disturbance in vegetation. Bot. Rev. 1979, 45, 229–299. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Thompson, J.N. Patch dynamics and the design of nature reserves. Biol. Conversat. 1978, 13, 27–37. [Google Scholar] [CrossRef]
- Whittaker, R.H.; Levin, S.A. The role of mosaic phenomena in natural communities. Theor. Popul. Biol. 1977, 12, 117–139. [Google Scholar] [CrossRef]
- Standovár, T.; Primack, R. A Természetvédelmi Biológiai Alapjai; Nemzeti Tankönyvkiadó: Budapest, Hungary, 2001; pp. 58–96. [Google Scholar]
- Ascasco, J.; Reiné, R. Temporal Variations in the Production—Quality and Optimal Cutting Date of Hay Meadows in the Central Pyrenees (Spain). Agronomy 2022, 12, 918. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Végvári, Z.; Hartel, T.; Schmotzer, A.; Kapocsi, I.; Tóthmérész, B. Grassland fires in Hungary—A problem or a potential alternative management tool? Appl. Ecol. Environ. Res. 2014, 12, 267–283. [Google Scholar] [CrossRef]
- Dengler, J.; Janišová, M.; Török, P.; Wellstein, C. Biodiversity of Palaearctic grasslands: A synthesis. Agric. Ecosyst. Environ. 2014, 182, 1–14. [Google Scholar] [CrossRef]
- Kiss, T.; Lévai, P.; Ferencz, Á.; Szentes, S.; Hufnagel, L.; Nagy, A.; Balogh, Á.; Pintér, O.; Saláta, D.; Házi, J.; et al. Change of composition and diversity of species and grassland management between different grazing intensity—In Pannonian dry and wet grasslands. Appl. Ecol. Environ. Res. 2011, 9, 197–230. [Google Scholar] [CrossRef]
- Klimeš, L.; Jongepierova, I.; Jongepier, J.W. Effect of mowing on a previously abandoned meadow: Ten year experiment. Priroda 2000, 17, 7–24. [Google Scholar]
- Házi, J.; Bartha, S.; Szentes, S.; Penksza, K. Seminatural grassland management by mowing of Calamagrostis epigejos in Hungary. Plant Biosyst. 2010, 145, 699–707. [Google Scholar] [CrossRef]
- Szirmai, O.; Saláta, D.; Benedek, L.K.; Czóbel, S. Investigation of the Secondary Succession of Abandoned Areas from Different Cultivation in the Pannonian Biogeographic Region. Agronomy 2022, 12, 773. [Google Scholar] [CrossRef]
- Házi, J.; Penksza, K.; Bartha, S.; Hufnagel, L.; Tóth, A.; Gyuricza, C.; Szentes, S. Cut mowing and grazing Effects with grey cattle on plant species composition in case of Pannon wet grasslands. Appl. Ecol. Environ. Res. 2012, 10, 223–231. [Google Scholar] [CrossRef]
- Kelemen, A.; Török, P.; Valkó, O.; Deák, B.; Miglécz, T.; Tóth, K.; Ölvedi, T.; Tóthmérész, B. Sustaining recovered grasslands is not likely without proper management: Vegetation changes and large-scale evidences after cessation of mowing. Biodivers. Conserv. 2014, 23, 741–751. [Google Scholar] [CrossRef]
- Maccherini, S.; Santi, E.; Torri, D. Germinable Soil Seed Bank in Biancana Badlands. Diversity 2019, 11, 223. [Google Scholar] [CrossRef]
- Valkó, O.; Török, P.; Tóthmérész, B.; Matus, G. Restoration potential in seed banks of acidic fen and dry- mesophilous meadows: Can restoration be based on local seed banks? Restor. Ecol. 2011, 19, 9–15. [Google Scholar] [CrossRef]
- Guimaräes-Steinicke, C.; Weigelt, A.; Ebeling, A.; Eisenhauer, N.; Wirth, C. Diversity Effects on Canopy Structure Change throughout a Growing Season in Experimental Grassland Communities. Remote Sens. 2022, 14, 1557. [Google Scholar] [CrossRef]
- Zhao, J.; Cao, J.; Che, Z.; Guo, Y.; Ma, C. Contribution of Sheep Grazing to Plant Diversity in Natural Grasslands. Diversity 2022, 14, 446. [Google Scholar] [CrossRef]
- Török, P.; Arany, I.; Prommer, M.; Valkó, O.; Balogh, A.; Vida, E.; Tóthmérész, B.; Matus, G. Vegetation, phytomass and seed bank of strictly protected hay-making Molinion meadows in Zemplén Mountains (Hungary) after restored management. Thaiszia J. Bot. 2009, 19, 67–77. [Google Scholar]
- Török, P.; Deák, B.; Vida, E.; Valkó, O.; Lengyel, S.; Tóthmérész, B. Restoring grassland biodiversity: Sowing lowdiversity seed mixtures can lead to rapid favourable changes. Biol. Conserv. 2010, 143, 806–812. [Google Scholar] [CrossRef]
- Török, P.; Kelemen, A.; Valkó, O.; Deák, B.; Lukács, B.; Tóthmérész, B. Lucerne-dominated fields recover native grass diversity without intensive management actions. J. Appl. Ecol. 2011, 48, 257–264. [Google Scholar] [CrossRef]
- Komarek, L. A Dél-Alföldi Régió erdősültségének alakulása a rendszerváltozás utáni időszakban. ÖKO—Ökológia-Környezetgazdálkodás-Társadalom 2005, 13, 113–119. [Google Scholar]
- Komarek, L. A hazai erdőgazdálkodás néhány indikátorának alakulása, különös tekintettel napjainkra. A Földrajz Tanítása—Módszertani Folyóirat 2007, 15, 10–19. [Google Scholar]
- Komarek, L. Spatial and temporal changes in some indicators of Hungarian forest management. Infrastruct. Ecol. Rural. Areas 2015, 11, 441–456. [Google Scholar]
- Komarek, L. Hungarian forest management tendencies at the beginning of the XXI century. Russ. J. Agric. Socio-Econ. Sci. 2018, 78, 7–18. [Google Scholar] [CrossRef]
- Morris, M.G. The effects of structure and its dynamics on the ecology and conservation of arthropods in British grasslands. Biol. Conserv. 2000, 95, 129–142. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Kelemen, A.; Miglécz, T.; Szabó, S.; Szabó, G.; Tóthmérész, B. Micro-topographic heterogeneity increases plant diversity in old stages of restored grasslands. Basic Appl. Ecol. 2015, 16, 291–299. [Google Scholar] [CrossRef]
- Ryser, P.; Langenauer, R.; Gigon, A. Species richness and vegetation structure in a limestone grassland after 15 years management with six biomass removal regimes. Folia Geobot. Phytotaxon. 1995, 30, 157–167. [Google Scholar] [CrossRef]
- Fiala, K.; Holub, P.; Sedláková, I.; Tůma, I.; Záhora, J.; Tesařová, M. Reasons and consequences of expansion of Calamagrostis epigejos in alluvial meadows of landscape affected by water control measures. Ekológia 2003, 22 (Suppl. 2), 242–252. [Google Scholar]
- Wang, X.; Jiang, L.; Yang, X.; Shi, Z.; Yu, P. Does Shrub Encroachment Indicate Ecosystem Degradation? A Perspective Based on the Spatial Patterns of Woody Plants in a Temperate Savanna-Like Ecosystem of Inner Mongolia, China. Forests 2020, 11, 1248. [Google Scholar]
- Virágh, K.; Horváth, A.; Bartha, S.; Somodi, I. A multiscale methodological approach novel in monitoring the effectiveness of grassland management. Community Ecol. 2008, 9, 237–246. [Google Scholar] [CrossRef]
- Leung, T.K.C.; So, K.Y.K.; Shum, B.T.W.; Hau, B.C.H. Optimal Mowing Regime in Enhancing Biodiversity in Seasonal Floodplains along Engineered Channels. Sustainability 2022, 14, 4002. [Google Scholar] [CrossRef]
- Sendžikaite, J.; Pakalnis, R. Extensive use of sown meadows—A tool for restoration of botanical diversity. J. Environ. Eng. Landsc. Manag. 2006, 14, 149–158. [Google Scholar] [CrossRef]
- Saláta, D.; Wichmann, B.; Házi, J.; Falusi, E.; Penksza, K. Botanikai összehasonlító vizsgálat a cserépfalui és az erdőbényei fás legelőn. AWETH 2011, 7, 234–262. [Google Scholar]
- Stampfli, A.; Zeiter, M. Plant species decline due to abandonment of meadows cannot easily be reversed by mowing. A case study from the southern Alps. J. Veg. Sci. 1999, 10, 151–164. [Google Scholar]
- Billeter, R.; Peintinger, M.; Diemer, M. Restoration of montane fen meadows by mowing remains possible after 4–35 years of abandonment. Acta Bot. Helv. 2007, 117, 1–13. [Google Scholar] [CrossRef]
- Gerard, M.; El Kahloun, M.; Rymen, J.; Beauchard, O.; Meire, P. Importance of mowing and flood frequency in promoting species richness in restored floodplains. J. Appl. Ecol. 2008, 45, 1780–1789. [Google Scholar] [CrossRef]
- Kelemen, A.; Török, P.; Deák, B.; Valkó, O.; Lukács, B.A.; Lengyel, S.; Tóthmérész, B. Spontán gyepregeneráció extenzíven kezelt lucernásokban. Tájökológiai Lapok 2010, 8, 33–44. [Google Scholar]
- Rodríguez-Rojo, M.P.; Roig, S.; López-Carrasco, C.; García, M.M.R.; Sánchez-Mata, D. Which Factors Favour Biodiversity in Iberian Dehesas? Sustainability 2022, 14, 2345. [Google Scholar] [CrossRef]
- Bogunovic, I.; Kljak, K.; Dugan, I.; Grbeša, D.; Telak, L.J.; Duvnjak, M.; Kisic, I.; Solomun, M.K.; Pereira, P. Grassland Management Impact on Soil Degradation and Herbage Nutritional Value in a Temperate Humid Environment. Agriculture 2022, 12, 921. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, Y.; Li, J.; Zheng, L. Degradation or Restoration? The Temporal-Spatial Evolution of Ecosystem Services and Its Determinants in the Yellow River Basin, China. Land 2022, 11, 863. [Google Scholar] [CrossRef]
- Bonanomi, G.; Caporaso, S.; Allegrezza, M. Short-term effects of nitrogen enrichment, litter removal and cutting on a Mediterranean grassland. Acta Oecol. 2006, 30, 419–425. [Google Scholar] [CrossRef]
- Halász, A.; Nagy, G.; Tasi, J.; Bajnok, M.; Mikoné, J.E. Weather regulated cattle behaviour on rangeland. Appl. Ecol. Environ. Res. 2016, 14, 149–158. [Google Scholar] [CrossRef]
- Pápay, G.; Kiss, O.; Fehér, Á.; Szabó, G.; Zimmermann, Z.; Hufnágel, L.; S.-Falusi, E.; Járdi, I.; Saláta, D.; Szemethy, L.; et al. Impact of shrub cover and wild ungulate browsing on the vegetation of restored mountain hay meadows. Tuexenia 2020, 40, 445–457. [Google Scholar]
- Katona, K.; Fehér, Á.; Szemethy, L.; Saláta, D.; Pápay, G.; S.-Falusi, E.; Kerényi-Nagy, V.; Szabó, G.; Wichmann, B.; Penksza, K. Vadrágás szerepe a mátrai hegyvidéki gyepek becserjésedésének lassításában. Gyepgazdálkodási Közlemények 2016, 14, 29–35. [Google Scholar]
- Penksza, K.; Fehér, Á.; Szemethy, L.; Saláta, D.; Pápay, G.; S.-Falusi, E.; Kerényi-Nagy, V.; Szabó, G.; Wichmann, B.; Katona, K. Gyepregeneráció és vadrágás vizsgálata cserjeirtás után parádóhutai (Mátra) mintaterületeken. Gyepgazdálkodási Közlemények 2016, 14, 36–41. [Google Scholar]
- Vasa, L.; Gyuricza, C.; Penksza, K. Methods of economical evaluation of grasslands under extreme climatic conditions based on plant sociological samples. Actual Probl. Econ. 2013, 145, 251–260. [Google Scholar]
- Láng, S. A Mátra és a Börzsöny Természeti Földrajza; Akadémiai Kiadó: Budapest, Hungary, 1955; pp. 32–45. [Google Scholar]
- Borhidi, A.; Kevey, B.; Lendvai, G. Plant Communities of Hungary; Academic Press: Budapest, Hungary, 2012; pp. 123–143. [Google Scholar]
- Országos Vadgazdálkodási Adattár. Available online: http://www.ova.info.hu (accessed on 12 December 2021).
- Katona, K.; Kiss, M.; Bleier, N.; Székely, J.; Nyeste, M.; Kovács, V.; Terhes, A.; Fodor, Á.; Olajos, T.; Rasztovits, E.; et al. Ungulate browsing shapes climate change impacts on forest biodiversity in Hungary. Biodivers. Conserv. 2013, 22, 1167–1180. [Google Scholar] [CrossRef][Green Version]
- Katona, K.; Kiss, M.; Bleier, N.; Székely, J.; Nyeste, M.; Kovács, V.; Terhes, A.; Fodor, Á.; Olajos, T.; Szemethy, L. Növényevő nagy vadak rágáspreferenciái, mint a táplálkozási igények indikátorai. Vadbiológia 2013, 15, 63–71. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie; Spinger: Vienna, Austria; Berlin, Germany, 1964; pp. 23–76. [Google Scholar]
- Plants of the World Online. Available online: https://powo.science.kew.org/ (accessed on 27 June 2022).
- Simon, T. A Magyarországi Edényes Flóra Határozója; Nemzeti Tankönyvkiadó: Budapest, Hungary, 2000; pp. 698–789. [Google Scholar]
- Borhidi, A. Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in the Hungarian flora. Acta Bot. Hung. 1995, 39, 97–181. [Google Scholar]
- Raunkiær, C. Life Form of Plants and Statistical Plant Geography; Clarendon Press: Oxford, UK, 1934; pp. 23–97. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.5-7; R Core Team: Vienna, Austria, 2020; Available online: https://CRAN.R-project.org/package=vegan (accessed on 3 January 2022).
- Wickham, H. ggplot2: Version 2.0 Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Hammer, Ø. PAST—PAleontological STatictics Version 4.07 Reference Manual; University of Oslo: Oslo, Norway, 2021; 293p. [Google Scholar]
- Weigl, P.D.; Knowles, T.W. Temperate mountain grasslands: A climate-herbivore hypothesis for origins and persistence. Biol. Rev. 2014, 89, 466–476. [Google Scholar] [CrossRef]
- Kovács-Hostyánszki, A.; Elek, E.; Balázs, K.; Centeri, C.; S.-Falusi, E.; Jeanneret, P.; Penksza, K.; Podmaniczky, L.; Szalkovszki, O.; Báldi, A. Earthworms, spiders and bees as indicators of habitat and management in a low-input farming region—A whole farm approach. Ecol. Indic. 2013, 33, 111–120. [Google Scholar] [CrossRef]
- Tälle, M.; Deák, B.; Poschlod, P.; Valkó, O.; Westerberg, L.; Milberg, P. Grazing vs. mowing: A meta-analysis of biodiversity benefits for grassland management. Agric. Ecosyst. Environ. 2016, 15, 200–212. [Google Scholar] [CrossRef]
- Török, P.; Valkó, O.; Deák, B.; Kelemen, A.; Tóth, E.; Tóthmérész, B. Managing for composition or species diversity?—Pastoral and year-round grazing systems in alkali grasslands. Agric. Ecosyst. Environ. 2016, 234, 23–30. [Google Scholar] [CrossRef]
- Szabó, M.; Kenéz, Á.; Saláta, D.; Szemán, L.; Malatinszky, Á. Studies on botany and environmental management relations on a wooded pasture between Pénzesgyőr and Hárskút villages. Cereal Res. Commun. 2007, 35, 1133–1136. [Google Scholar] [CrossRef]
- Vida, E.; Török, P.; Deák, B.; Tóthmérész, B. Gyepek létesítése mezőgazdasági művelés alól kivont területeken: A gyepesítés módszereinek áttekintése. Bot. Közlemények 2008, 95, 101–113. [Google Scholar]
- Huhta, A.P.; Rautio, P.; Tuomi, J.; Laine, K. Restorative mowing on an abandoned semi-natural meadow: Short-term and predicted long-term effects. J. Veg. Sci. 2001, 12, 677–686. [Google Scholar] [CrossRef]
- Beltman, B.; van den Broek, T.; Martin, W.; Ten Cate, M.; Güsewell, S. Impact of mowing regime on species richness and biomass of a limestone hay meadow in Ireland. Bull. Geobot. Inst. ETH 2003, 69, 17–30. [Google Scholar]
- Losvik, M. Plant species diversity ina n old, traditionally managed hay meadow compared to abandoned meadows in southwest Norway. Nord. J. Bot. 1999, 19, 473–487. [Google Scholar] [CrossRef]
- Willems, J.H.; Peet, R.K.; Bik, L. Changes in chalk-grassland structure and species richness resulting from selective nutrient additions. J. Veg. Sci. 1993, 4, 203–212. [Google Scholar] [CrossRef]
- Willems, J.H. Species composition and above ground phytomass in chalk grassland with different management. Vegetatio 1983, 52, 171–180. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Lasanta, T.; Romo, A. Analysis of spatial and temporal evolution of vegetation cover in the spanish central pyrenees: Role of human management. Environ. Manag. 2004, 34, 802–818. [Google Scholar] [CrossRef] [PubMed]
- Kozak, J. Forest cover change in the western carpathians in the past 180 years: A case study in the Orawa region in Poland. Mt. Res. Dev. 2003, 23, 369–375. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Lindenmayer, D.; Guariguata, M.R.; Crouzeilles, R.; Rey Benayas, J.M.; Lazos Chavero, E. Fostering natural forest regeneration on former agricultural land through economic and policy interventions. Environ. Res. Lett. 2020, 15, 043002. [Google Scholar] [CrossRef]
- Pérez-Luque, A.J.; Bonet-García, F.J.; Zamora, R. Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region. Forests 2021, 12, 1584. [Google Scholar] [CrossRef]
- Archer, S. Assessing and interpreting grass-woody plant dynamics. The ecology and dynamics in a continental north-west European heathland. J. Appl. Ecol. 1996, 37, 415–431. [Google Scholar]
- Kelemen, A.; Tölgyesi, C.; Kun, R.; Molnár, Z.; Vadász, C.; Tóth, K. Positive small-scale effects of shrubs on diversity and flowering in pastures. Tuexenia 2017, 37, 399–413. [Google Scholar]
- Hasan, S.S.; Zhen, L.; Miah, M.G.; Ahamed, T.; Samie, A. Impact of land use change on ecosystem services: A review. Environ. Dev. 2020, 34, 100527. [Google Scholar] [CrossRef]
- Archer, S. Harry Stzobbs Memorial Lecture, 1993: Herbivore mediation of grass-woody plants interactions. Trop. Grassl. 1995, 29, 218–235. [Google Scholar]
- Pott, R. Effects of human interference on the landscape with special reference to the role of grazing livestock. In Grazing and Conservation Management; Wallis de Vries, M.F., Bakker, J.P., Van Wieren, S.E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 321–347. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zachar, Z.; Pápay, G.; Csontos, P.; Szabó, G.; Zimmermann, Z.; Saláta, D.; Szentes, S.; Pajor, F.; Fuchs, M.; Penksza, K. The Effects of Different Management Methods on Restored Grasslands in Potential Temperate Forest Zones. Diversity 2022, 14, 551. https://doi.org/10.3390/d14070551
Zachar Z, Pápay G, Csontos P, Szabó G, Zimmermann Z, Saláta D, Szentes S, Pajor F, Fuchs M, Penksza K. The Effects of Different Management Methods on Restored Grasslands in Potential Temperate Forest Zones. Diversity. 2022; 14(7):551. https://doi.org/10.3390/d14070551
Chicago/Turabian StyleZachar, Zalán, Gergely Pápay, Péter Csontos, Gábor Szabó, Zita Zimmermann, Dénes Saláta, Szilárd Szentes, Ferenc Pajor, Márta Fuchs, and Károly Penksza. 2022. "The Effects of Different Management Methods on Restored Grasslands in Potential Temperate Forest Zones" Diversity 14, no. 7: 551. https://doi.org/10.3390/d14070551
APA StyleZachar, Z., Pápay, G., Csontos, P., Szabó, G., Zimmermann, Z., Saláta, D., Szentes, S., Pajor, F., Fuchs, M., & Penksza, K. (2022). The Effects of Different Management Methods on Restored Grasslands in Potential Temperate Forest Zones. Diversity, 14(7), 551. https://doi.org/10.3390/d14070551