
How Will the Distributions of Native and Invasive Species Be Affected by Climate Change? Insights from Giant South American Land Snails

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Occurrence Datasets
2.2. Environmental Data
2.3. Modeling Procedures
2.4. Bioclimatic Niche Analysis
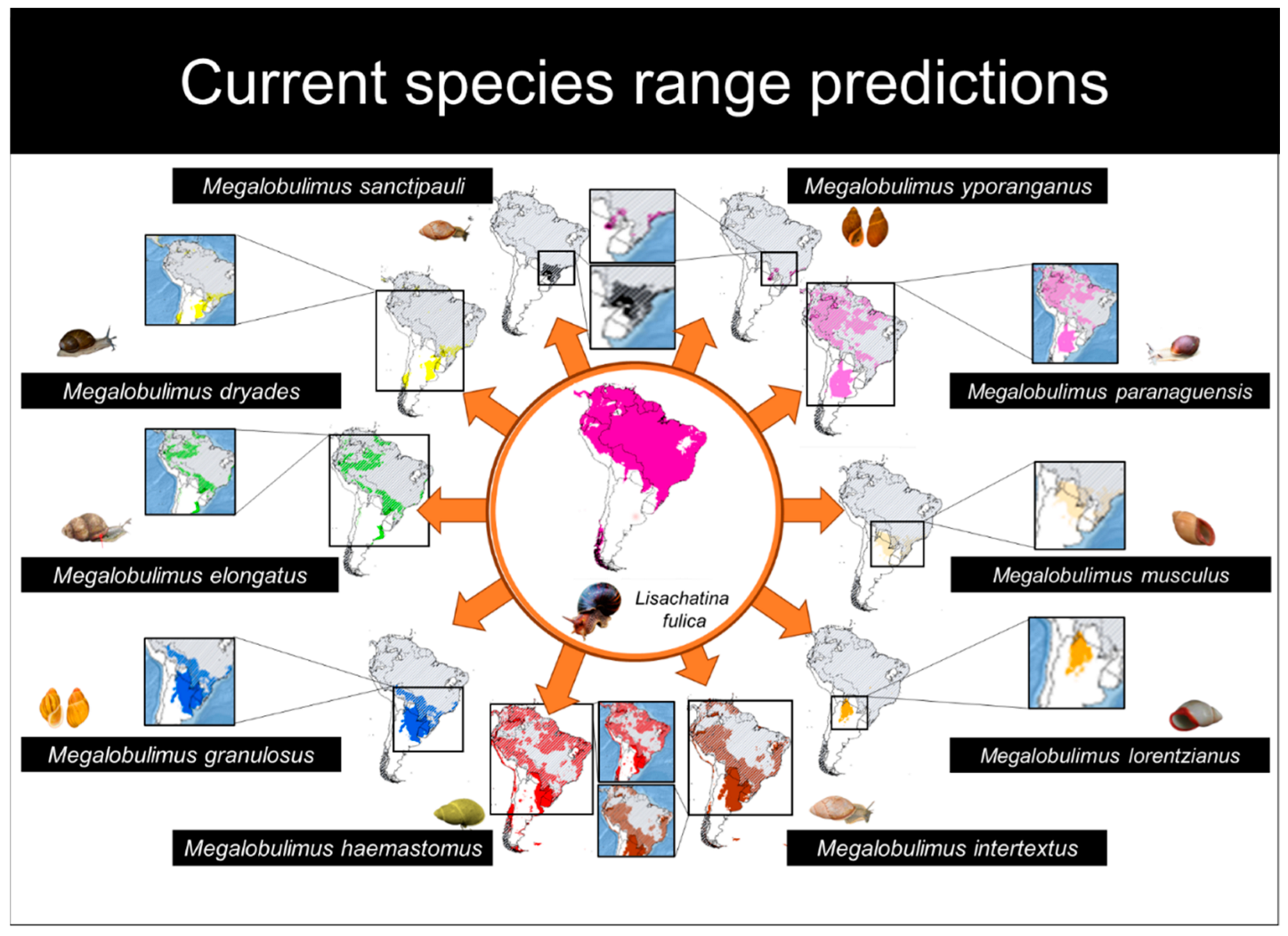
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vale, M.M.; Alves, M.A.S.; Lorini, M.L. Mudanças climáticas: Desafios e oportunidades para a conservação da biodiversidade brasileira. Oecol. Bras. 2009, 13, 518–535. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Evol. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Cordellier, M.; Pfenninger, A.; Streit, B.; Pfenninger, M. Assessing the effects of climate change on the distribution of pulmonate freshwater snail biodiversity. Mar. Biol. 2012, 159, 2519–2531. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; Ferreira-de-Siqueira, M.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef] [PubMed]
- Bøhn, T.; Amundsen, P.A.; Sparrow, A. Competitive exclusion after invasion? Biol. Invasions 2007, 10, 359–368. [Google Scholar] [CrossRef]
- Parker, I.M.; Simberloff, D.; Lonsdale, W.M.; Goodell, K.; Wonham, M.; Kareiva, P.M.; Williamson, M.H.; Holle, B.; Moyle, P.B.; Byers, J.E. Impact: Toward a framework for understanding the ecological effects of invaders. Biol. Invasions 1999, 1, 3–19. [Google Scholar] [CrossRef]
- Mooney, H.A.; Cleland, E.E. The evolutionary impact of invasive species. Proc. Natl. Acad. Sci. USA 2001, 98, 5446–5451. [Google Scholar] [CrossRef]
- Novacek, M.J.; Cleland, E.E. The current biodiversity extinction event: Scenarios for mitigation and recovery. Proc. Natl. Acad. Sci. USA 2001, 98, 5466–5470. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with invasive-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Nurinsiyah, A.S.; Hausdorf, B. Listing, impact assessment and prioritization of introduced land snail and slug species in Indonesia. J. Molluscan Stud. 2019, 85, 92–102. [Google Scholar] [CrossRef]
- Adelino, J.R.P.; Heringer, G.; Diagne, C.; Courchamp, F.; Faria, L.B.; Zenni, R.D. The economic costs of biological invasions in Brazil: A first assessment. NeoBiota 2021, 67, 349–374. [Google Scholar] [CrossRef]
- Bellard, C.; Jeschke, J.M.; Leroy, B.; Mace, G.M. Insights from modeling studies on how climate change affects invasive alien species geography. Ecol. Evol. 2018, 8, 5688–5700. [Google Scholar] [CrossRef] [PubMed]
- Bequaert, J.C. Studies in the Achatininae, a group of African land snails. Bull. Mus. Comp. Zool. 1950, 105, 1–216. [Google Scholar]
- Teles, H.M.S.; Vaz, J.F.; Fontes, R.L.; Domingos, M.F. Registro de Achatina fulica Bowdich, 1822 (Mollusca, Gastropoda) no Brasil: Caramujo hospedeiro intermediário da angiostrongilíase. Rev. Saúde Pública 1997, 31, 310–312. [Google Scholar] [CrossRef]
- Thiengo, S.; Faraco, F.A.; Salgado, N.C.; Cowie, R.H.; Fernandez, M.A. Rapid spread of an invasive snail in South America: The giant African snail, Achatina fulica, in Brazil. Biol. Invasions 2007, 9, 693–702. [Google Scholar] [CrossRef]
- Thiengo, S.C.; Simões, R.O.; Fernandes, M.A.; Maldonado, A. Angiostrongylus cantonensis and Rat Lungworm disease in Brazil. Hawai’i J. Med. Public Health 2013, 72, 18–22. [Google Scholar]
- Vogler, R.E.; Beltramino, A.A.; Sede, M.M.; Gutiérrez-Gregoric, D.E.; Núñez, V.; Rumi, A. The giant African snail, Achatina fulica (Gastropoda: Achatinidae): Using bioclimatic models to identify South American areas susceptible to invasion. Am. Malac. Bull. 2013, 31, 39–50. [Google Scholar] [CrossRef]
- Caldeira, R.L.; Mendonça, C.L.G.F.; Goveia, C.O.; Lenzi, H.L.; Graeff-Texteira, C.; Lima, W.S.; Mota, E.M.; Pecora, I.L.; Medeiros, A.M.Z.; Carvalho, O.S. First record of molluscs naturally infected with Angiostrongylus cantonensis (Chen, 1935) (Nematoda: Metastrongylidae) in Brazil. Memórias Inst. Oswaldo Cruz 2007, 102, 887–889. [Google Scholar] [CrossRef]
- Morassutti, A.L.; Thiengo, S.C.; Fernandez, M.; Sawanyawisuth, K.; Graeff-Teixeira, C. Eosinophilic meningitis caused by Angiostrongylus cantonensis: An emergent disease in Brazil. Mem. Inst. Oswaldo Cruz 2014, 109, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, F.S.; Peso-Aguiar, M.C.; Assunção-Albuquerque, M.J.T. Distribution, feeding behavior and control strategies of the exotic land snail Achatina fulica (Gastropoda: Pulmonata) in the northeast of Brazil. Braz. J. Biol. 2008, 68, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Rault, S.K.; Baker, G.M. Achatina fulica Bowdich and other Achatinidae as pests in tropical agriculture. In Molluscs as Crop Pests; Baker, G.M., Ed.; CAB International: New York, NY, USA, 2002; pp. 55–114. [Google Scholar]
- Meyer, W.M., III; Hayes, K.A.; Meyer, A.L. Giant African snail, Achatina fulica, as a snail predator. Am. Malacol. Bull. 2008, 24, 117–119. [Google Scholar] [CrossRef]
- Bequaert, J.C. Monograph of the Strophocheilidae, a neotropical family of terrestrial mollusks. Bull. Mus. Comp. Zool. 1948, 100, 1–210. [Google Scholar]
- Simone, L.R.L. Land and Freshwater Molluscs of Brazil; EGB/Fapesp: São Paulo, Brazil, 2006; pp. 1–390. [Google Scholar]
- Borda, V.; Ramirez, R. Re-characterization of the Red-lip Megalobulimus (Gastropoda: Strophocheilidae) from Peru with description of a new species. Zoologia 2013, 30, 675–691. [Google Scholar] [CrossRef]
- Chase, R.; Robinson, D.G. The uncertain history of land snails on Barbados: Implications for conservation. Malacologia 2001, 43, 33–57. [Google Scholar]
- Rosenberg, G.; Muratov, I. Status Report on the Terrestrial Mollusca of Jamaica. Proc. Acad. Nat. Sci. Phila. 2005, 155, 117–161. [Google Scholar] [CrossRef]
- Scott, M.I.H. Estudio anatómico del Borus Strophocheilus lorentzianus (Doer.) (Mol. Pulm.). Rev. Mus. Plata 1939, 7, 217–278. [Google Scholar]
- Miranda, M.S.; Correia, L.V.B.; Pecora, I.L. Activity and reproduction in Megalobulimus paranaguensis (Gastropoda, Eupulmonata): Implications for conservation in captivity for a South American land snail. J. Nat. Hist. 2020, 54, 435–443. [Google Scholar] [CrossRef]
- Miranda, M.S.; Fontenelle, J.H. Population dynamics of Megalobulimus paranaguensis in the Brazilian southeast coast. Zoologia 2015, 32, 463–468. [Google Scholar] [CrossRef]
- Fontenelle, J.H.; Miranda, M.S. Aspects of biology of Megalobulimus paranaguensis (Gastropoda, Acavoidea) in the coastal plain of the Brazilian southeast. Iheringia Ser. Zool. 2017, 107, e2017004. [Google Scholar] [CrossRef]
- Miranda, M.S.; Fontenelle, J.H.; Pecora, I.L. Population structure of a native and an invasive species of snail in an urban fragment of Atlantic Rainforest. J. Nat. Hist. 2015, 49, 19–35. [Google Scholar] [CrossRef]
- Santos, S.B.; Miyahira, I.C.; Mansur, M.C.D. Freshwater and terrestrial molluscs in Brasil: Current status of knowledge and conservation. Tentacle 2013, 21, 40–42. [Google Scholar]
- Beltramino, A.A.; Vogler, R.E.; Gutiérrez-Gregoric, D.E.; Rumi, A. Impact of climate change on the distribution of a giant land snail from South America: Predicting future trends for setting conservation priorities on native malacofauna. Clim. Change 2015, 131, 621–633. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Araújo, M.B.; Guisan, A. Five (or so) challenges for species distribution modelling. J. Biogeogr. 2006, 33, 1677–1688. [Google Scholar] [CrossRef]
- Devictor, V.; Clavel, J.; Julliard, R.; Lavergne, S.; Mouillot, D.; Thuiller, W.; Venail, P.; Villéger, S.; Mouquet, N. Defining and measuring ecological specialization. J. Appl. Ecol. 2010, 47, 15–25. [Google Scholar] [CrossRef]
- SpeciesLink. Available online: https://splink.cria.org.br/ (accessed on 1 February 2021).
- Portal da Biodiversidade Reflora. Available online: https://portaldabiodiversidade.icmbio.gov.br/portal/ (accessed on 1 February 2021).
- Global Biodiversity Information Facility (GBIF). Available online: https://www.gbif.org (accessed on 1 February 2021).
- iNaturalist. Available online: https://www.inaturalist.org/ (accessed on 1 February 2021).
- Web of Science. Available online: www.webofknowledge.com (accessed on 1 February 2021).
- Miranda, M.S.; (University of Campinas, Campinas, SP, Brazil). Personal communication, 2021.
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- WorldClim 2.0. Available online: http://www.worldclim.org (accessed on 1 February 2021).
- Meinshausen, M.; Nicholls, Z.R.J.; Lewis, J.; Gidden, M.J.; Vogel, E.; Freund, M.; Beyerle, U.; Gessner, C.; Nauels, A.; Bauer, N.; et al. The shared socio-economic pathway (SSP) greenhouse gas concentrations and their extensions to 2500. Geosci. Model Dev. 2020, 13, 3571–3605. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- De Marco, P.; Nóbrega, C.C. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PLoS ONE 2018, 13, e0202403. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Leroy, B.; Delsol, R.; Hugueny, B.; Meynard, C.N.; Barhoumi, C.; Barbet-Massin, M.; Bellard, C. Without quality presence–absence data, discrimination metrics such as TSS can be misleading measures of model performance. J. Biogeogr. 2018, 45, 1994–2002. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 3.6.6.; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 February 2020).
- Andrade, A.F.A.; Velazco, S.J.E.; Júnior, P.M. ENMTML: An R package for a straightforward construction of complex ecological niche models. Environ. Model. Softw. 2020, 125, 104615. [Google Scholar] [CrossRef]
- Schoener, T.W. Nonsynchronous spatial overlap of lizards in patchy habitats. Ecology 1970, 51, 408–418. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef]
- Albuquerque, F.S.; Peso-Aguiar, M.C.; Assunção-Albuquerque, M.J.T.; Gálvez, L. Do climate variables and human density affect Achatina fulica (Bowditch) (Gastropoda: Pulmonata) shell length, total weight and condition factor? Braz. J. Biol. 2009, 69, 879–885. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Patiño-Montoya, A.; Giraldo, A.; Tidon, R. Variation in the population density of the giant African snail (Lissachatina fulica) in the Neotropical region. Caldasia 2022, 44, 1–21. [Google Scholar] [CrossRef]
- Fontanilla, I.K.C.; Maria, I.M.P.S.; Garcia, J.R.M.; Ghate, H.; Naggs, F.; Wade, C. Restricted Genetic Variation in Populations of Achatina (Lissachatina) fulica outside of East Africa and the Indian Ocean Islands Points to the Indian Ocean Islands as the Earliest Known Common Source. PLoS ONE 2014, 9, e105151. [Google Scholar] [CrossRef] [PubMed]
- Beltramino, A.A. Distribution of Megalobulimus sanctipauli (Ihering & Pilsbry, 1900) (Gastropoda: Megalobulimidae) in South America. CheckList 2013, 9, 469–471. [Google Scholar] [CrossRef]
- Beltramino, A.A. Distribución histórica y área de distribución potencial del megamolusco terrestre Megalobulimus lorentzianus (Doering, 1876) (Gastropoda: Pulmonata) em América del Sur. Boletín Asoc. Argent. Malacol. 2014, 4, 10–13. [Google Scholar]
- Miranda, M.S.; Correia, L.V.B. Food choice in Megalobulimus paranaguensis (Gastropoda, Eupulmonata). Stud. Neotrop. Fauna Environ. 2021, 56, 108–111. [Google Scholar] [CrossRef]
- Borrero, F.J.; Breure, A.S.H.; Christensen, C.; Correoso, M.; Mogollón-Ávila, V. Into the Andes: Three new introductions of Lissachatina fulica (Gastropoda, Achatinidae) and its potential distribution in South America. Tentacle 2009, 17, 6–8. [Google Scholar]
- Correoso, M.; Coello, M. Modelación y distribución de Lissachatina fulica (Gastropoda: Achatinidae) en Ecuador. Potenciales impactos ambientales y sanitarios. Rev. Geoespacial 2009, 6, 79–90. [Google Scholar]
- Lydeard, C.; Cowie, R.H.; Ponder, W.F.; Bogan, A.E.; Bouchet, P.; Clark, S.A.; Cummings, K.S.; Frest, T.J.; Gargominy, O.; Herbert, D.G.; et al. The global decline of nonmarine mollusks. Bioscience 2004, 54, 321–330. [Google Scholar] [CrossRef]
- Régnier, C.; Fontaine, B.; Bouchet, P. Not knowing, not recording, not listing: Numerous unnoticed mollusk extinctions. Conserv. Biol. 2009, 23, 1214–1221. [Google Scholar] [CrossRef]
- Ramírez, R.; Paredes, C.; Arenas, J. Moluscos del Perú. Rev. Biol. Trop. 2003, 51, 225–284. [Google Scholar]
- Gutiérrez-Gregoric, D.E.; Núñez, V.; Vogler, R.E.; Beltramino, A.A.; Rumi, A. Gasterópodos terrestres de la provincia de Misiones, Argentina. Rev. Biol. Trop. 2013, 61, 1759–1768. [Google Scholar] [CrossRef]
- Simone, L.R.L. Mollusca terrestres. In Biodiversidade do Estado de São Paulo: Uma Síntese do Conhecimento ao Final do Século XX. Invertebrados Terrestres; Brandão, C.R.F., Cancello, E.M., Eds.; FAPESP: São Paulo, Brazil, 1999; Volume 5, pp. 3–8. [Google Scholar]
- Salvador, R.B. Land snail diversity in Brazil. Strombus 2019, 25, 10–20. [Google Scholar]
- Foden, W.; Mace, G.; Vié, J.C.; Ângulo, A.; Butchart, S.; DeVantier, L.; Dublin, H.; Gutsche, A.; Stuart, S.; Turak, E. Species susceptibility to climate change impacts. In The 2008 Review of the IUCN Red List of Threatened Species; Vié, J.C., Hilton-Taylor, C., Stuart, S.N., Eds.; IUCN: Gland, Switzerland, 2008; pp. 1–11. [Google Scholar]
- Johnson, D.M.; Büntgen, U.; Frank, D.C.; Kausrud, K.; Haynes, K.J.; Liebhold, A.M.; Esper, J.; Stenseth, N.C. Climatic warming disrupts recurrent Alpine insect outbreaks. Proc. Natl. Acad. Sci. USA 2010, 107, 20576–20581. [Google Scholar] [CrossRef]
- Barker, G.M. The Biology of Terrestrial Molluscs; CABI: Wallingford, New Zealand, 2001; pp. 1–560. [Google Scholar]
- Fischer, M.L.; Colley, E. Espécie invasora em reservas naturais: Caracterização da população de Achatina fulica Bowdich, 1822 (Mollusca—Achatinidae) na Ilha Rasa, Guaraqueçaba, Paraná, Brasil. Biota Neotrop. 2005, 5, 127–144. [Google Scholar] [CrossRef]
- Fischer, M.L.; Simião, M.; Colley, E.; Zenni, R.D.; Silva, D.A.T.; Latoski, N. O caramujo exótico invasor na vegetação nativa em Morretes, PR: Diagnóstico da população de Achatina fulica Bowdich, 1822 em um fragmento de Floresta Ombrófila Densa aluvial. Biota Neotrop. 2006, 6, 1–5. [Google Scholar] [CrossRef]
- Eston, M.R.; Menezes, G.V.; Antunes, A.Z.; Santos, A.S.R.; Santos, A.M.R. Espécie invasora em unidade de conservação: Achatina fulica (Bowdich, 1822) no Parque Estadual Carlos Botelho, Sete Barras, SP, Brasil. Rev. Inst. Florest. 2006, 18, 173–179. [Google Scholar]
- Lake, P.S.; O’Dowd, D.J. Red crabs in rain forest, Christmas Island: Biotic resistance to invasion by an exotic snail. Oikos 1991, 62, 25–29. [Google Scholar] [CrossRef]
- Nurinsiyah, A.S.; Fauzia, H.; Hennig, C.; Hausdorf, B. Native and introduced land snail species as ecological indicators in different land use types in Java. Ecol. Indic. 2016, 70, 557–565. [Google Scholar] [CrossRef]
- Cowie, R.H. Decline and homogenization of Pacific faunas: The land snails of American Samoa. Biol. Conserv. 2001, 99, 207–222. [Google Scholar] [CrossRef]
- Cowie, R.H. Invasive non-marine molluscs in the islands of the tropical and subtropical Pacific: A review. Am. Malacol. Bull. 2005, 20, 95–103. [Google Scholar]
- Colley, E.; Fischer, M.L. Avaliação dos problemas enfrentados no manejo do caramujo gigante africano Achatina fulica (Gastropoda: Pulmonata) no Brasil. Zoologia 2009, 26, 674–683. [Google Scholar] [CrossRef]
- Miranda, M.S.; Pecora, I.L. Conservation implications of behavioural interactions between the Giant African Snail and a Native Brazilian species. Ethol. Ecol. Evol. 2017, 29, 209–217. [Google Scholar] [CrossRef]
- Barker, G.M.; Mayhill, P.C. Patterns of diversity and habitat relationships in terrestrial mollusc communities of the Pukeamaru Ecological District, northeastern New Zealand. J. Biogeogr. 1999, 26, 215–238. [Google Scholar] [CrossRef]
- Chiba, S.; Cowie, R.H. Evolution and extinction of land snails on oceanic islands. Ann. Rev. Ecol. Evol. Syst. 2016, 47, 123–141. [Google Scholar] [CrossRef]
- O’Loughlin, L.S.; Green, P.T. The secondary invasion of giant African land snail has little impact on litter or seedling dynamics in rainforest. Austral Ecol. 2017, 42, 819–830. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 |
|---|---|---|---|---|---|---|
| Annual mean temperature (bio1) | 0.272 | −0.223 | −0.132 | −0.050 | 0.060 | −0.001 |
| Mean diurnal range (bio2) | −0.173 | −0.242 | −0.128 | 0.495 | −0.122 | −0.524 |
| Isothermality (bio3) | 0.239 | −0.017 | 0.312 | 0.105 | 0.225 | −0.529 |
| Temperature seasonality (bio4) | −0.250 | 0.008 | −0.386 | −0.063 | −0.228 | 0.101 |
| Maximum temperature of warmest period (bio5) | 0.193 | −0.316 | −0.343 | −0.036 | −0.134 | −0.065 |
| Minimum temperature of warmest period (bio6) | 0.295 | −0.118 | 0.005 | −0.160 | 0.085 | 0.017 |
| Annual temperature range (bio7) | −0.249 | −0.125 | −0.329 | 0.200 | −0.250 | −0.086 |
| Mean temperature of wettest quarter (bio8) | 0.235 | −0.251 | −0.235 | 0.056 | 0.116 | 0.010 |
| Mean temperature of driest quarter (bio9) | 0.277 | −0.154 | 0.016 | −0.172 | −0.001 | −0.029 |
| Mean temperature of warmest quarter (bio10) | 0.231 | −0.260 | −0.306 | −0.103 | −0.034 | 0.031 |
| Mean temperature of coldest quarter (bio11) | 0.287 | −0.181 | −0.005 | −0.033 | 0.101 | −0.029 |
| Annual precipitation (bio12) | 0.262 | 0.224 | −0.040 | 0.187 | −0.193 | 0.065 |
| Precipitation of wettest period (bio13) | 0.270 | 0.081 | 0.114 | 0.242 | −0.324 | 0.215 |
| Precipitation of driest period (bio14) | 0.141 | 0.397 | −0.241 | 0.029 | 0.113 | −0.296 |
| Precipitation seasonality (bio15) | −0.029 | −0.338 | 0.401 | 0.340 | −0.134 | 0.015 |
| Precipitation of wettest quarter (bio16) | 0.270 | 0.091 | 0.102 | 0.247 | −0.321 | 0.212 |
| Precipitation of driest quarter (bio17) | 0.152 | 0.396 | −0.232 | 0.031 | 0.084 | −0.269 |
| Precipitation of warmest quarter (bio18) | 0.160 | 0.193 | −0.205 | 0.562 | 0.342 | 0.328 |
| Precipitation of coldest quarter (bio19) | 0.198 | 0.209 | 0.073 | −0.195 | −0.605 | −0.241 |
| Individual proportion | 0.553 | 0.198 | 0.093 | 0.056 | 0.042 | 0.027 |
| Cumulative proportion | 0.553 | 0.751 | 0.843 | 0.899 | 0.941 | 0.968 |
| Species | Jaccard | Size at Present (n. Cells) | Size at Future (n. Cells) | Percentage of Change |
|---|---|---|---|---|
| M. dryades | 0.741 | 17,904 | 15,599 | −12.874 |
| M. elongatus | 0.444 | 36,317 | 47,879 | +31.836 |
| M. granulosus | 0.881 | 36,579 | 41,984 | +14,776 |
| M. haemastomus | 0.666 | 114,933 | 104,166 | −9680 |
| M. lorentzianus | 0.757 | 5407 | 7983 | +47.641 |
| M. musculus | 0.483 | 17,804 | 18,597 | +4.454 |
| M. paranaguensis | 0.729 | 85,605 | 78,483 | −8319 |
| M. sanctipauli | 0.805 | 7986 | 6394 | −19.934 |
| M. yporanganus | 0.818 | 2444 | 1924 | −21.276 |
| M. intertextus | 0.933 | 99,410 | 96,305 | −3.123 |
| L. fulica | 0.755 | 156,294 | 151,516 | −3.057 |
| Species | Similarity | Unfilling | Stability | ||||
|---|---|---|---|---|---|---|---|
| D | 1→2 | 1←2 | Megalobulimus | L. fulica | Megalobulimus | L. fulica | |
| M. dryades | 0.039 | 0.010 | 0.010 | 0.728 | 0.000 | 0.272 | 1.000 |
| M. elongatus | 0.021 | 0.020 | 0.030 | 0.801 | 0.000 | 0.199 | 1.000 |
| M. granulosus | 0.033 | 0.010 | 0.010 | 0.783 | 0.019 | 0.217 | 0.981 |
| M. haemastomus | 0.033 | 0.040 | 0.040 | 0.601 | 0.042 | 0.399 | 0.958 |
| M. lorentzianus | 0.002 | 0.188 | 0.218 | 0.983 | 0.558 | 0.017 | 0.442 |
| M. musculus | 0.035 | 0.020 | 0.010 | 0.646 | 0.046 | 0.354 | 0.954 |
| M. paranaguensis | 0.046 | 0.010 | 0.010 | 0.736 | 0.000 | 0.264 | 1.000 |
| M. sanctipauli | 0.024 | 0.040 | 0.020 | 0.790 | 0.013 | 0.210 | 0.987 |
| M. yporanganus | 0.024 | 0.010 | 0.020 | 0.772 | 0.000 | 0.228 | 1.000 |
| M. intertextus | 0.014 | 0.040 | 0.059 | 0.564 | 0.033 | 0.436 | 0.967 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teles, W.S.; Silva, D.d.P.; Vilela, B.; Lima-Junior, D.P.; Pires-Oliveira, J.C.; Miranda, M.S. How Will the Distributions of Native and Invasive Species Be Affected by Climate Change? Insights from Giant South American Land Snails. Diversity 2022, 14, 467. https://doi.org/10.3390/d14060467
Teles WS, Silva DdP, Vilela B, Lima-Junior DP, Pires-Oliveira JC, Miranda MS. How Will the Distributions of Native and Invasive Species Be Affected by Climate Change? Insights from Giant South American Land Snails. Diversity. 2022; 14(6):467. https://doi.org/10.3390/d14060467
Chicago/Turabian StyleTeles, Wanderson Siqueira, Daniel de Paiva Silva, Bruno Vilela, Dilermando Pereira Lima-Junior, João Carlos Pires-Oliveira, and Marcel Sabino Miranda. 2022. "How Will the Distributions of Native and Invasive Species Be Affected by Climate Change? Insights from Giant South American Land Snails" Diversity 14, no. 6: 467. https://doi.org/10.3390/d14060467
APA StyleTeles, W. S., Silva, D. d. P., Vilela, B., Lima-Junior, D. P., Pires-Oliveira, J. C., & Miranda, M. S. (2022). How Will the Distributions of Native and Invasive Species Be Affected by Climate Change? Insights from Giant South American Land Snails. Diversity, 14(6), 467. https://doi.org/10.3390/d14060467