
Shopping for Ecological Indices? On the Use of Incidence-Based Species Compositional Similarity Measures

 ,
,  , , , , , , and
, , , , , , and
Abstract
1. Introduction
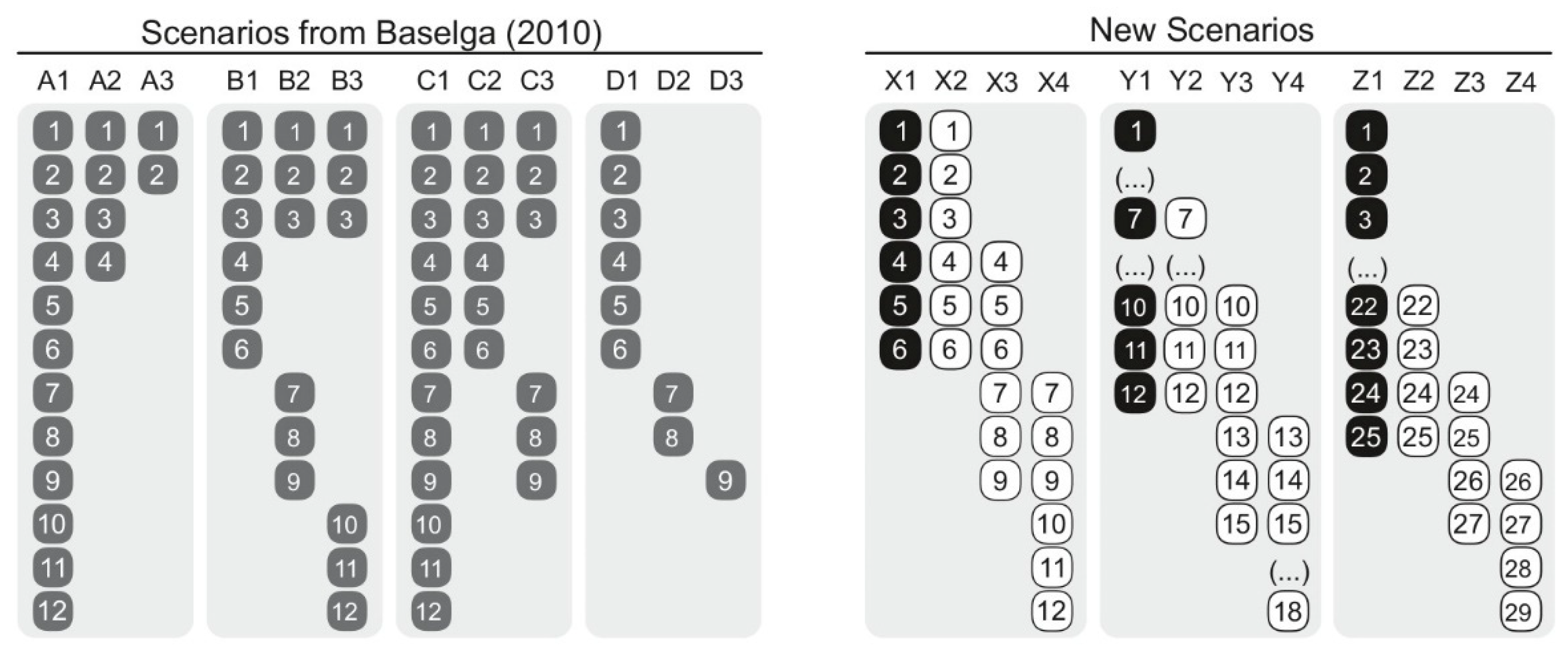
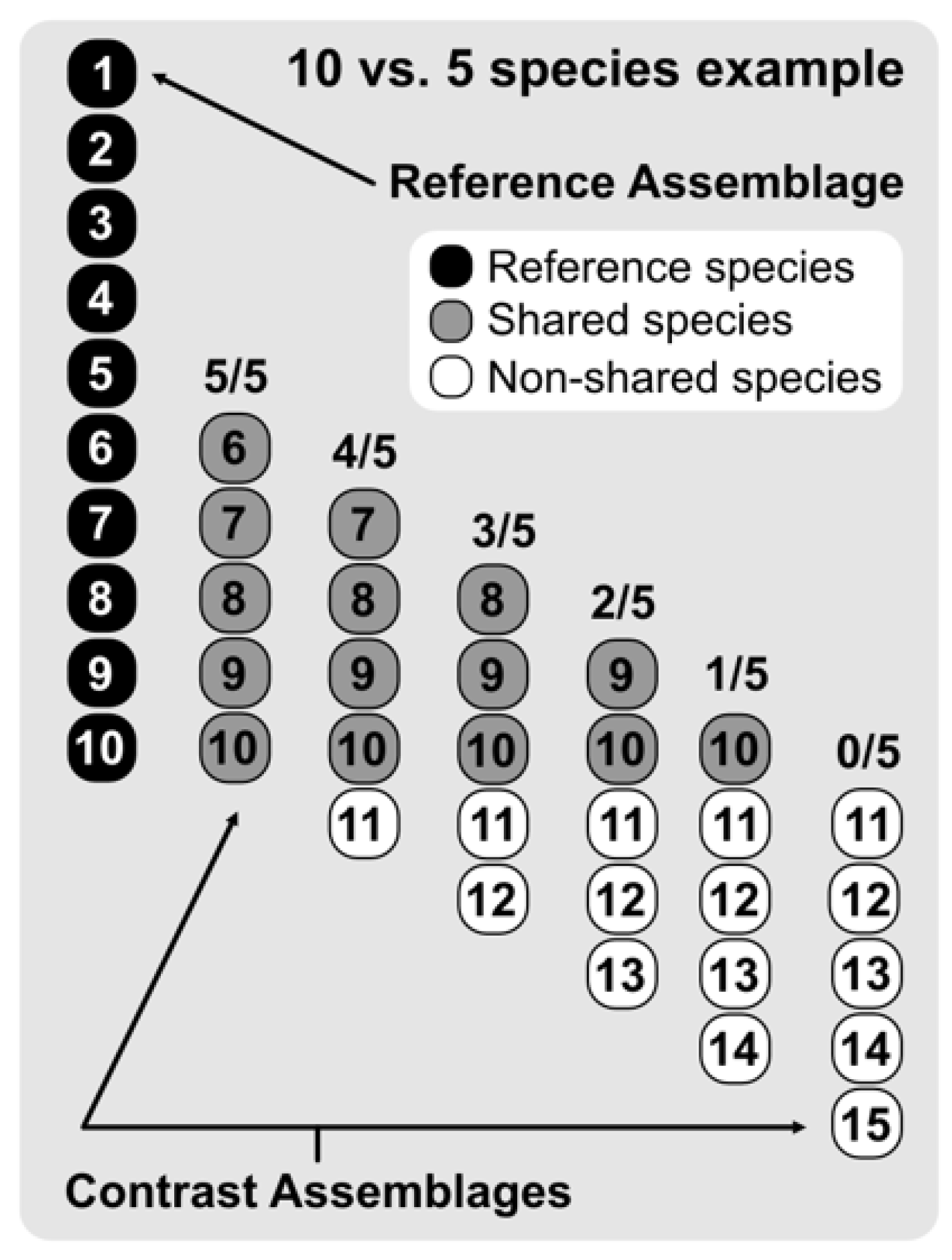
2. Materials and Methods
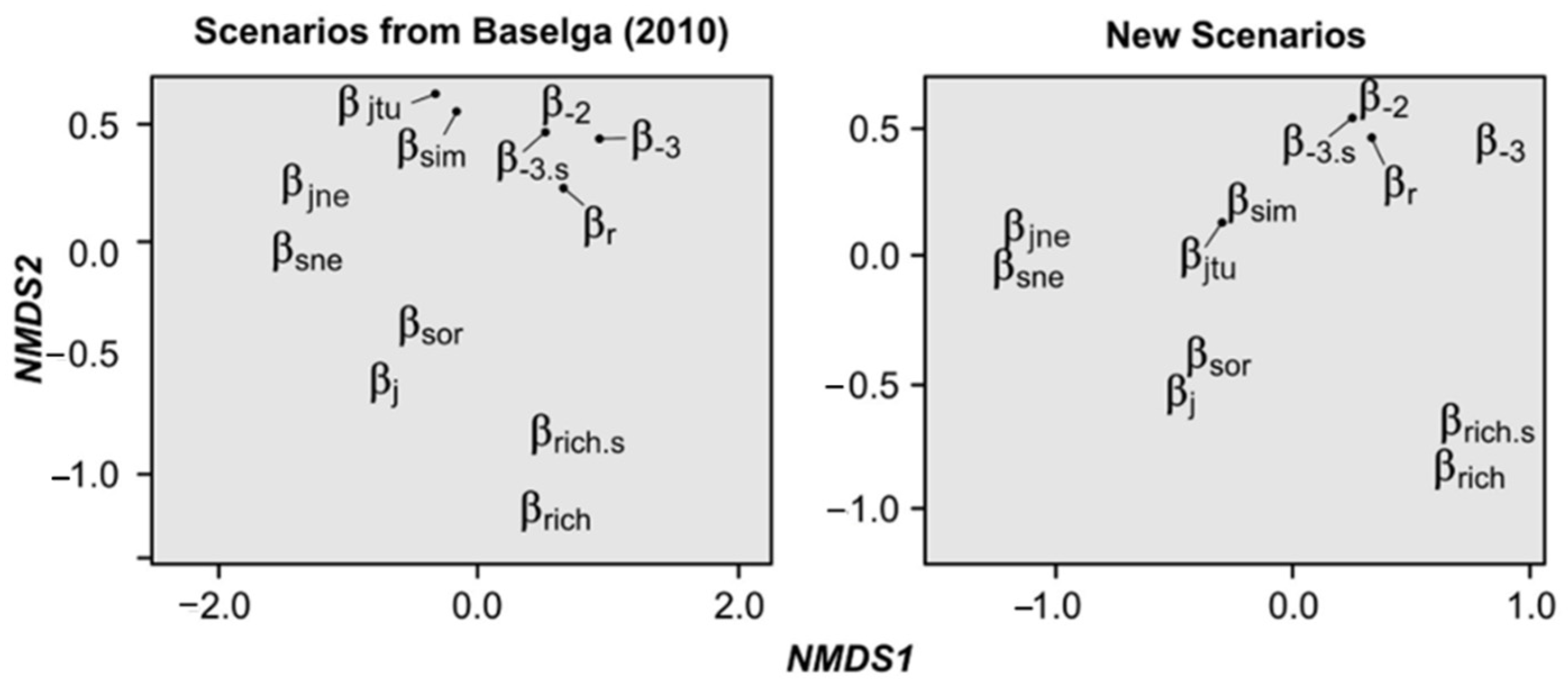
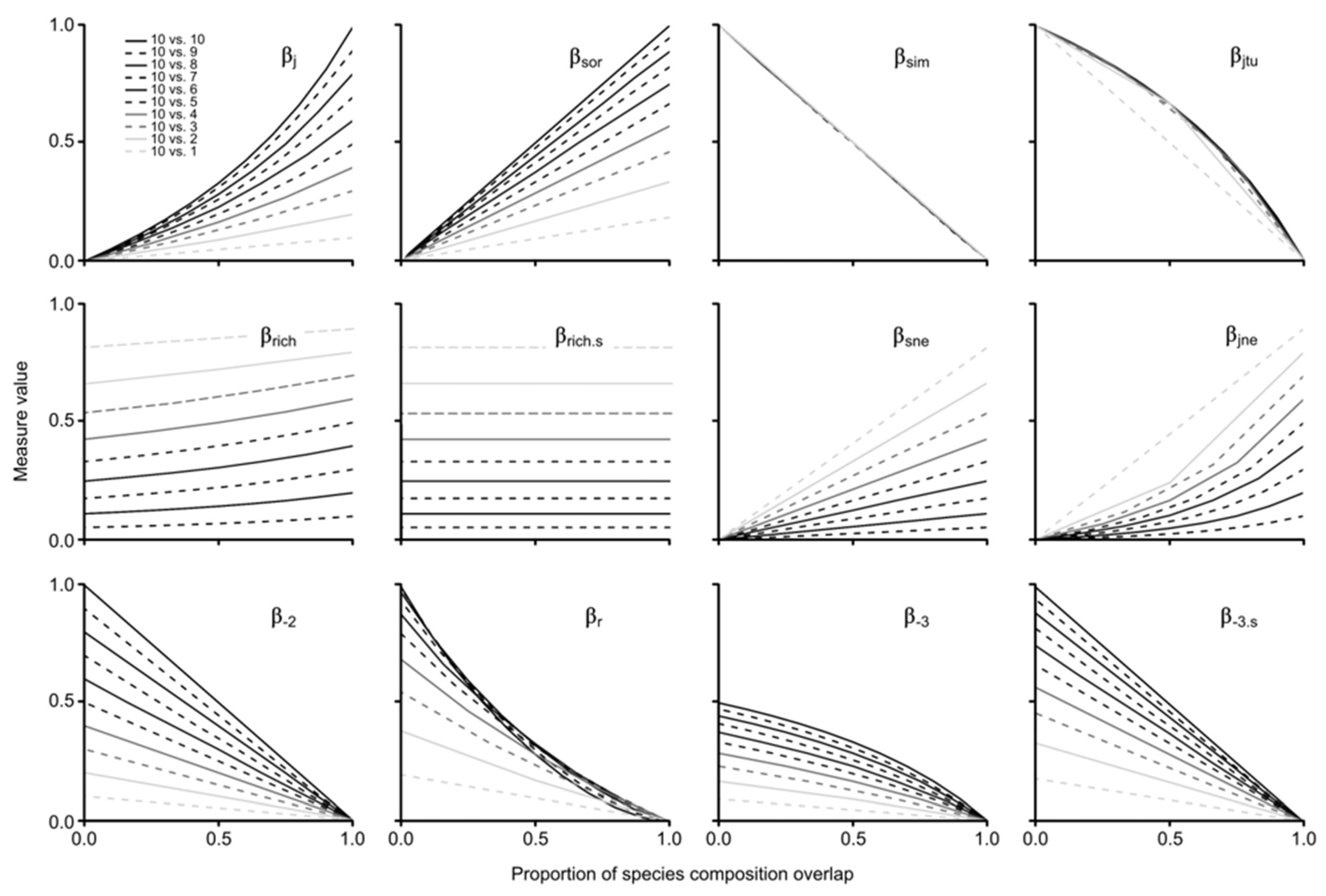
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jaccard, P. The Distribution of the Flora in the Alpine Zone. New Phytol. 1912, 11, 37–50. [Google Scholar] [CrossRef]
- Koch, L. Index of Biota Dispersity. Ecology 1957, 38, 145–148. [Google Scholar] [CrossRef]
- Moreno, C.E.; Rodríguez, P. A Consistent Terminology for Quantifying Species Diversity? Oecologia 2010, 163, 279–282. [Google Scholar] [CrossRef]
- Anderson, M.J.; Crist, T.O.; Chase, J.M.; Vellend, M.; Inouye, B.D.; Freestone, A.L.; Sanders, N.J.; Cornell, H.V.; Comita, L.S.; Davies, K.F.; et al. Navigating the Multiple Meanings of β Diversity: A Roadmap for the Practicing Ecologist. Ecol. Lett. 2011, 14, 19–28. [Google Scholar] [CrossRef]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shen, T.-J. Abundance-Based Similarity Indices and Their Estimation When There Are Unseen Species in Samples. Biometrics 2006, 62, 361–371. [Google Scholar] [CrossRef]
- Chao, A.; Chiu, C.-H. Bridging the Variance and Diversity Decomposition Approaches to Beta Diversity via Similarity and Differentiation Measures. Methods Ecol. Evol. 2016, 7, 919–928. [Google Scholar] [CrossRef]
- Wilson, M.V.; Shmida, A. Measuring Beta Diversity with Presence-Absence Data. J. Ecol. 1984, 72, 1055–1064. [Google Scholar] [CrossRef]
- Calderón-Patrón, J.M.; Moreno, C.E.; Zuria, I. La Diversidad Beta: Medio Siglo de Avances. Rev. Mex. Biodivers. 2012, 83, 879–891. [Google Scholar] [CrossRef]
- Jurasinski, G.; Retzer, V.; Beierkuhnlein, C. Inventory, Differentiation, and Proportional Diversity: A Consistent Terminology for Quantifying Species Diversity. Oecologia 2009, 159, 15–26. [Google Scholar] [CrossRef]
- Lande, R. Statistics and Partitioning of Species Diversity, and Similarity among Multiple Communities. Oikos 1996, 76, 5–13. [Google Scholar] [CrossRef]
- Jost, L. Partitioning Diversity into Independent Alpha and Beta Components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [PubMed]
- Koleff, P.; Gaston, K.J.; Lennon, J.J. Measuring Beta Diversity for Presence–Absence Data. J. Anim. Ecol. 2003, 72, 367–382. [Google Scholar] [CrossRef]
- Cardoso, P.; Borges, P.A.V.; Veech, J.A. Testing the Performance of Beta Diversity Measures Based on Incidence Data: The Robustness to Undersampling. Divers. Distrib. 2009, 15, 1081–1090. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the Turnover and Nestedness Components of Beta Diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A. The Relationship between Species Replacement, Dissimilarity Derived from Nestedness, and Nestedness. Glob. Ecol. Biogeogr. 2012, 21, 1223–1232. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P.; Borges, P.A.V.; Schmera, D.; Podani, J. Measuring Fractions of Beta Diversity and Their Relationships to Nestedness: A Theoretical and Empirical Comparison of Novel Approaches. Oikos 2013, 122, 825–834. [Google Scholar] [CrossRef]
- Baselga, A.; Leprieur, F. Comparing Methods to Separate Components of Beta Diversity. Methods Ecol. Evol. 2015, 6, 1069–1079. [Google Scholar] [CrossRef]
- Chao, A.; Chiu, C.-H.; Wu, S.-H.; Huang, C.-L.; Lin, Y.-C. Comparing Two Classes of Alpha Diversities and Their Corresponding Beta and (Dis)Similarity Measures, with an Application to the Formosan Sika Deer Cervus Nippon Taiouanus Reintroduction Programme. Methods Ecol. Evol. 2019, 10, 1286–1297. [Google Scholar] [CrossRef]
- Almeida-Neto, M.; Frensel, D.M.B.; Ulrich, W. Rethinking the Relationship between Nestedness and Beta Diversity: A Comment on Baselga (2010). Glob. Ecol. Biogeogr. 2012, 21, 772–777. [Google Scholar] [CrossRef]
- Chen, Y.; Schmera, D. Additive Partitioning of a Beta Diversity Index Is Controversial. Proc. Natl. Acad. Sci. USA 2015, 112, E7161. [Google Scholar] [CrossRef]
- Podani, J.; Schmera, D. A New Conceptual and Methodological Framework for Exploring and Explaining Patterns in Presence-Absence Data. Oikos 2011, 120, 1625–1638. [Google Scholar] [CrossRef]
- Podani, J.; Schmera, D. Once Again on the Components of Pairwise Beta Diversity. Ecol. Inform. 2016, 32, 63–68. [Google Scholar] [CrossRef][Green Version]
- Diniz-Filho, J.A.F.; Collevatti, R.G.; Soares, T.N.; de Campos Telles, M.P. Geographical Patterns of Turnover and Nestedness-Resultant Components of Allelic Diversity among Populations. Genetica 2012, 140, 189–195. [Google Scholar] [CrossRef]
- Mouillot, D.; De Bortoli, J.; Leprieur, F.; Parravicini, V.; Kulbicki, M.; Bellwood, D.R. The Challenge of Delineating Biogeographical Regions: Nestedness Matters for Indo-Pacific Coral Reef Fishes. J. Biogeogr. 2013, 40, 2228–2237. [Google Scholar] [CrossRef]
- Norhazrina, N.; Wang, J.; Hagborg, A.; Geffert, J.L.; Mutke, J.; Gradstein, S.R.; Baselga, A.; Vanderpoorten, A.; Patiño, J. Tropical Bryophyte Floras: A Homogeneous Assemblage of Highly Mobile Species? Insights from Their Spatial Patterns of Beta Diversity. Bot. J. Linn. Soc. 2017, 183, 16–24. [Google Scholar] [CrossRef]
- Ramachandran, V.; Robin, V.V.; Tamma, K.; Ramakrishnan, U. Climatic and Geographic Barriers Drive Distributional Patterns of Bird Phenotypes within Peninsular India. J. Avian Biol. 2017, 48, 620–630. [Google Scholar] [CrossRef]
- Schmera, D.; Podani, J. Comments on Separating Components of Beta Diversity. Community Ecol. 2011, 12, 153–160. [Google Scholar] [CrossRef]
- Legendre, P. Interpreting the Replacement and Richness Difference Components of Beta Diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef]
- Murray, K.A.; Baselga, A. Reply to Chen and Schmera: Partitioning Beta Diversity into Replacement and Nestedness-Resultant Components Is Not Controversial. Proc. Natl. Acad. Sci. USA 2015, 112, E7162. [Google Scholar] [CrossRef]
- Lennon, J.J.; Koleff, P.; Greenwood, J.J.D.; Gaston, K.J. The Geographical Structure of British Bird Distributions: Diversity, Spatial Turnover and Scale. J. Anim. Ecol. 2001, 70, 966–979. [Google Scholar] [CrossRef]
- Georgopoulou, E.; Neubauer, T.A.; Strona, G.; Kroh, A.; Mandic, O.; Harzhauser, M. Beginning of a New Age: How Did Freshwater Gastropods Respond to the Quaternary Climate Change in Europe? Quat. Sci. Rev. 2016, 149, 269–278. [Google Scholar] [CrossRef]
- Koyanagi, T.F.; Furukawa, T.; Osawa, T. Nestedness-Resultant Community Disassembly Process of Extinction Debt in a Highly Fragmented Semi-Natural Grassland. Plant Ecol. 2018, 2019, 1093–1103. [Google Scholar] [CrossRef]
- Andrew, M.E.; Wulder, M.A.; Coops, N.C.; Baillargeon, G. Beta-Diversity Gradients of Butterflies along Productivity Axes. Glob. Ecol. Biogeogr. 2012, 21, 352–364. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, D.; Villeger, S.; De Bortoli, J.; Leprieur, F. Package ‘Betapart’: Partitioning Beta Diversity into Turnover and Nestedness Components. R Package Version 1.4–1. 2017. Available online: https://CRAN.R-project.org/package=betapart (accessed on 22 April 2018).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘Vegan’: Community Ecology Package. R Package Version 2.4–3. 2017. Available online: https://CRAN.R-project.org/package=vegan (accessed on 20 August 2017).
- Waters, C.N.; Zalasiewicz, J.; Summerhayes, C.; Barnosky, A.D.; Poirier, C.; Gałuszka, A.; Cearreta, A.; Edgeworth, M.; Ellis, E.C.; Ellis, M.; et al. The Anthropocene Is Functionally and Stratigraphically Distinct from the Holocene. Science 2016, 351, aad2622. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Forcino, F.L.; Leighton, L.R.; Twerdy, P.; Cahill, J.F. Reexamining Sample Size Requirements for Multivariate, Abundance-Based Community Research: When Resources Are Limited, the Research Does Not Have to Be. PLoS ONE 2015, 10, e0128379. [Google Scholar] [CrossRef]
- Cardoso, P.; Rigal, F.; Carvalho, J.C. BAT—Biodiversity Assessment Tools, an R Package for the Measurement and Estimation of Alpha and Beta Taxon, Phylogenetic and Functional Diversity. Methods Ecol. Evol. 2015, 6, 232–236. [Google Scholar] [CrossRef]
- Crawley, M.J. The R Book, 2nd ed.; Wiley: Chichester, UK, 2013; ISBN 978-0-470-97392-9. [Google Scholar]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Harrison, S.; Ross, S.J.; Lawton, J.H. Beta Diversity on Geographic Gradients in Britain. J. Anim. Ecol. 1992, 61, 151–158. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Malden, MA, USA, 2004. [Google Scholar]
- Holz, I.; Gradstein, S.R.; Heinrichs, J.; Kappelle, M. Bryophyte Diversity, Microhabitat Differentiation, and Distribution of Life Forms in Costa Rican Upper Montane Quercus Forest. Bryologist 2002, 105, 334–348. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E. The Role of Exotic Species in Homogenizing the North American Flora. Ecol. Lett. 2006, 9, 1293–1298. [Google Scholar] [CrossRef]
- Lasram, F.B.R.; Hattab, T.; Halouani, G.; Romdhane, M.S.; Le Loc’h, F. Modeling of Beta Diversity in Tunisian Waters: Predictions Using Generalized Dissimilarity Modeling and Bioregionalisation Using Fuzzy Clustering. PLoS ONE 2015, 10, e0131728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lei, M.; Li, Y.; Wang, P.; Wang, C.; Gao, Y.; Wu, H.; Xu, C.; Niu, L.; Wang, L.; et al. Determination of Vertical and Horizontal Assemblage Drivers of Bacterial Community in a Heavily Polluted Urban River. Water Res. 2019, 161, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Gotelli, N.J.; Colwell, R.K. Quantifying Biodiversity: Procedures and Pitfalls in the Measurement and Comparison of Species Richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Magurran, A.E.; McGill, B.J. (Eds.) Biological Diversity: Frontiers in Measurement and Assessment; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Keil, P. Z-Scores Unite Pairwise Indices of Ecological Similarity and Association for Binary Data. Ecosphere 2019, 10, e02933. [Google Scholar] [CrossRef]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Notation | Formula | Interpretation | References |
|---|---|---|---|
| βj | a/(a + b + c) | Jaccard dissimilarity | [34] |
| βsor | 2a/(2a + b + c) | Sørensen dissimilarity | [34] |
| βsim | min(b,c)/(min(b,c) + a) | Simpson (turnover component of βsor) | [34] |
| βr | 2bc/[(a + b + c)2 − 2bc] | Routledge degree of species overlap | [34] |
| β-2 | min(b,c)/(max(b,c) + a) | Harrison dissimilarity | [34] |
| β-3 | min(b,c)/(a + b + c) | Williams replacement index | [34] |
| βsne | [max(b,c) − min(b,c)/(2a + b + c)] × [a/(min(b,c) + a)] | Nestedness-resultant component of βsor | [35] |
| βrich | |b − c|/(a + b + c) | Richness difference component of βj | [35] |
| βjne | [max(b,c) − min(b,c)/(a + b + c)] × [a/(2min(b,c) + a)] | Nestedness-resultant component of βj | [35] |
| βjtu | 2min(b,c)/2min(b,c) + a | Turnover component of βj | [35] |
| βrich.s | |b − c|/(2a + b + c) | Legendre richness difference index | [35] |
| β-3.s | 2min(b,c)/(2a + b + c) | Legendre replacement index | [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MacGregor-Fors, I.; Escobar, F.; Escobar-Ibáñez, J.F.; Mesa-Sierra, N.; Alvarado, F.; Rueda-Hernández, R.; Moreno, C.E.; Falfán, I.; Corro, E.J.; Pineda, E.; et al. Shopping for Ecological Indices? On the Use of Incidence-Based Species Compositional Similarity Measures. Diversity 2022, 14, 384. https://doi.org/10.3390/d14050384
MacGregor-Fors I, Escobar F, Escobar-Ibáñez JF, Mesa-Sierra N, Alvarado F, Rueda-Hernández R, Moreno CE, Falfán I, Corro EJ, Pineda E, et al. Shopping for Ecological Indices? On the Use of Incidence-Based Species Compositional Similarity Measures. Diversity. 2022; 14(5):384. https://doi.org/10.3390/d14050384
Chicago/Turabian StyleMacGregor-Fors, Ian, Federico Escobar, Juan F. Escobar-Ibáñez, Natalia Mesa-Sierra, Fredy Alvarado, Rafael Rueda-Hernández, Claudia E. Moreno, Ina Falfán, Erick J. Corro, Eduardo Pineda, and et al. 2022. "Shopping for Ecological Indices? On the Use of Incidence-Based Species Compositional Similarity Measures" Diversity 14, no. 5: 384. https://doi.org/10.3390/d14050384
APA StyleMacGregor-Fors, I., Escobar, F., Escobar-Ibáñez, J. F., Mesa-Sierra, N., Alvarado, F., Rueda-Hernández, R., Moreno, C. E., Falfán, I., Corro, E. J., Pineda, E., Bourg, A., Aguilar-López, J. L., & Dáttilo, W. (2022). Shopping for Ecological Indices? On the Use of Incidence-Based Species Compositional Similarity Measures. Diversity, 14(5), 384. https://doi.org/10.3390/d14050384