Abstract
The Mediterranean Sea diversity is still far from being fully disclosed. Marine Heterobranchia are one of the most paradigmatic species-rich groups, with many recent systematic studies revealing the high density of new, cryptic, and endemic species occurring in the Mediterranean basin. In this study, sea slug Doriopsilla areolata, which was considered until today one of the most widespread nudibranchs worldwide, was investigated using a molecular approach to compare Mediterranean and Atlantic populations for the first time. The molecular analyses involved three different molecular markers, the two mitochondrial COI and 16S, and the nuclear H3 gene. The results revealed a complex of species within D. areolata that indeed consists of three potentially species, two of which are endemic to the Mediterranean Sea: Doriopsilla areolata, which is distributed in the Adriatic Sea (the type locality of the former species), D. rarispinosa, which occurs in the Western Mediterranean basin and along the Tunisian coast, and one additional Atlantic species here provisionally defined as Doriopsilla sp. 1. This study helps to unveil another case of cryptic diversity within Mediterranean Heterobranchia and to increase the knowledge on Doriopsilla genus diversity.
1. Introduction
Molluscs form the second largest phylum after arthropods with approximately 100,000 described species and potentially still 100,000 species to be described [1] with nearly 75% of species belonging to the Class Gastropoda [2]. Nudibranchs are the largest marine suborder within the Heterobranchia clade, and they are characterized by having lost their shell secondarily [3]. Doriopsilla areolata Bergh, 1880, is a radula-less nudibranch belonging to the family Dendrodorididae O’Donoghue, 1924 [4,5,6]. The absence of the radula is a characteristic feature within the family Dendrodorididae, and the reason they were classified in the group Porostomata Bergh, 1878 [5] together with the Phyllidiidae Rafinesque, 1814. However, the validity of Porostomata group has been discussed for several years and still needs to be better clarified [5,7,8,9,10]. Historically, the species within Dendrodorididae and Phyllidiidae are distinguished on the base of morphological diagnostic features [5,10] as the first ones are similar in appearance to other doridoidea, with their gills on the dorsum [5,10], while Phyllidiidae have their secondary gills typically located in a ventral/lateral position [5,10]. Two genera of the family Dendrodorididae are present in the Mediterranean: Doriopsilla Bergh 1880 and Dendrodoris Ehrenberg 1831 [4,10]. Their morphological and molecular separations are currently clearly defined and supported [5,10,11,12,13].
The genus Doriopsilla groups 24 accepted taxa, with D. areolata as the type species [14]. The distribution range of the different species included in the mentioned genus spans almost worldwide [10,15], as they occur in the shallow coastal waters of the Pacific coast of North America, Australia, Indo-Pacific, Persian Gulf, South and West Africa, Mediterranean Sea, European Eastern Atlantic, North American Western Atlantic, and Caribbean Sea [10,15,16,17]. To date, three species are reported in the Mediterranean Sea: Doriopsilla areolata Bergh, 1880 [10,15]; D. evanae Ballesteros and Ortea, 1980 [18]; and D. pelseneeri d’Oliveira, 1895 [11] as Dendrodoris minima [15,19,20,21,22,23,24]. Some authors [15] hypothesized D. evanae as a possible synonym of D. areolata; however, they are currently regarded as distinct valid species. Other synonymized names are known for D. areolata, including D. fedalae Pruvot-Fol, 1953; D. pusilla Pruvot-Fol, 1951; D. rarispinosa Pruvot-Fol, 1951; and Doris reticulata Schulz in Philippi, 1836: The latter is considered as invalid, being the junior homonym of Doris reticulata Quoy & Gaimard, 1832 [14]. A possibility that D. areolata hides a complex of cryptic species has also been suggested [25,26].
Doriopsilla areolata was described for the first time based on two specimens from Hvar Island in the Adriatic Sea (Eastern Mediterranean Sea) [27], but its geographic distribution range is currently deemed from the Eastern Atlantic Ocean (Spain to Angola), the Mediterranean Sea, and the Caribbean Sea [6,15,28]. Doriopsilla areolata can feed on corneous sponges [4,5] by secreting digestive enzymes directly onto the sponge and sucking up predigested organic matter by means of a pore-like transformed mouth and a foregut modified into a suctorial tube [4,5,29,30]. The body color pattern shows a high intraspecific variability varying from yellow in smaller D. areolata individuals to light brown or grayish in larger individuals [15]. Interestingly, different morphotypes can be observed within the species, with individuals showing patterns with small white spots or rings and lines on the dorsum and many tubercles that can be flat or rounded [15]. Doriopsilla areolata was split into three distinct subspecies based on their different geographical distributions [15]: D. areolata areolata Bergh, 1880, in the Mediterranean Sea and the Eastern Atlantic Ocean (from the coasts of Spain to the Cape Verde Islands), D. areolata albolineata Edmunds, 1968, along the southwestern coasts of Africa (from Ghana to Angola and the island of São Tomé) [15,31], and D. areolata nigrolineata Meyer, 1977 along the Caribbean coast of Panama [15,32]. The institution of subspecies was encouraged by the widespread distribution of D. areolata, supported by its planktotrophic developmental strategy that could have led to a higher dispersal potential compared to species with lecithotrophic or direct development [6,33]. However, molecular investigations based on the mitochondrial 16S and the nuclear H3 molecular markers [28,33] put in doubt the validity of the division of D. areolata into different subspecies. To confirm these speculations, the need to repeat the analyses with a larger sample size and additional and more informative genes, such as the barcoder COI, was highlighted [33]. Currently, the most used molecular markers in Heterobranchia systematics, both at species and genus taxonomic levels, are the mitochondrial gene regions of the cytochrome oxidase subunit 1 (COI) and 16S rRNA (16S) and the nuclear H3 DNA region. In fact, previous studies showed that the two mitochondrial markers are well suited to study closely related nudibranch species [34,35,36,37], while nuclear H3 revealed to be useful only at higher taxonomic levels due to its high conservation and slower mutation rate [38].
Hitherto, none of the previous molecular studies on Doriopsilla genus included Mediterranean D. areolata specimens. This is quite surprising being D. areolata the type-taxon of the genus and the Adriatic Sea (Mediterranean Sea) the type locality. This is a significant deficiency not only in terms of principle but also when we consider how many integrative systematic studies have recently shown that Mediterranean populations often are separated from the Atlantic ones with several cases of endemism and cryptic and new species [35,36,38,39,40,41,42,43,44,45,46,47]. Therefore, taking all these factors into consideration, the aims of the present study are as follows: (1) to molecularly compare the Mediterranean and Atlantic D. areolata populations by adding an additional useful molecular marker, COI barcoder, on an extended dataset; (2) to investigate on the possible occurrence of cryptic diversity within D. areolata in the Mediterranean Sea; and (3) to define the range of variability of the color pattern characterizing D. areolata.
2. Materials and Methods
Doriopsilla areolata individuals included in the present study were hand collected in different regions by SCUBA diving between 2012 and 2021 along the coasts of the Mediterranean Sea and the Eastern Atlantic Ocean (Table 1). In particular, 16 samples were collected in the Mediterranean Sea: nine of them were from Italian coastal areas, three were sampled from Spain, one sample was from Croatia, one from Tunisia, and the remaining three samples came from France. Two samples were collected from the coast on Spain in the East Atlantic Ocean. Sequences from other extra Mediterranean specimens were obtained from GenBank. Collected samples were photographed in situ and in laboratory, preserved in 95% ethanol (EtOH) for upcoming molecular analyses, and deposited in the Department of Science of the Roma Tre University collection (Vouchers RM3_ID number).

Table 1.
Species name, voucher code, sampling locality, and GenBank accession numbers of all the analyzed specimens are listed, together with the outgroups. In bold are the specimens examined in this study.
Molecular Analyses
DNA was extracted from the body tissues using the ‘salting out’ procedure [48]. First, a small piece of tissue was cut from the tail and placed in a tube where it was heated for one hour at 40 °C. In the following step, 430 μL of Cell Lysis Buffer and 20 μL of Proteinase K were added to the dried tissue. The samples were then incubated in a thermoblock overnight at a 56 °C. Next, the samples were vortexed and centrifuged at 13,200 rpm for 10 min. After this first centrifugation, the liquid supernatant was carefully pipetted into new tubes. Afterwards, 160 μL of NaCl 5 M was added to the samples, and these were gently vortexed and centrifuged for 10 min at 13,200 rpm. The supernatant was carefully taken and placed into the final tubes, and 500 μL of isopropanol was added. Next, the samples were gently vortexed and centrifuged under the same conditions used in the previous steps and finally the supernatant was discarded, leaving the DNA pellet adhering to the wall of the tubes. One ml of 80% EtOH was added, and the tubes were centrifuged for the last time for 10 min at 13,200 rpm. The supernatant was carefully discarded again, and the samples were left to dry for 1–2 h at room temperature. Finally, dried samples were diluted with the 60–100 μL of double distilled H2O. Two different mitochondrial gene regions, COI and 16S, and the nuclear H3 were amplified. The universal primers LCO1490 and HCO2198 [49] and 16Sar-L and 16Sbr-H [50] were used for the COI and 16S mitochondrial markers, respectively, while H3AD-F and H3BD-R universal primers [51] were used for nuclear H3. The temperature profile for the PCR reactions was the same for the three molecular markers at the beginning of an initial denaturation step at 94 °C, which lasted 5 min. This step was followed by 35 cycles consisting of 30 s. at 94 °C for the denaturation step, 60 s. at an annealing temperature of 46–50 °C and 60 s. at an elongation temperature of 72 °C. After this cycle, the temperature was held for another 7 min. at 72 °C. Once all these steps were completed, the entire reaction was cooled down to a temperature of 10 °C. The PCR reaction mix has a final volume of 20 μL and consisted of 14.6 μL of dH2O, 4.0 μL of 5x FIREPol Mastermix (5x Reaction buffer (0.4 M Tris-HCl, 0.1 M (NH4)2SO4, 0.1% w/v Tween-20], 12.5 mM MgCl2, 1 mM dNTP), 0.2 μL of each forward and reverse primers (20 μM), and 1.0 μL DNA. The quality of all obtained PCR products was controlled on 1.2% agarose gels. Samples were sequenced by Macrogen Europe (1105 Amsterdam, The Netherlands). Before the sequences were used for the alignment, they were controlled with the Basic Local Alignment Search Tool (BLAST) to exclude possible contamination. Sequences were aligned together with GenBank sequences using the Muscle algorithm implemented in MEGA 6.0 [52]. Four different alignments were generated, and three single-gene dataset (COI, 16S, and H3) and one with the three genes were concatenated and partitioned (ConcDNA). Primer regions were always removed from the final alignments. In the case of the 16S alignment, it was proofread with Gblocks 0.91b [53,54], allowing less strict flanking positions as the less stringent setting selection, to remove the hyper-divergent regions. The best-fitting evolutionary model for each of the four datasets (three single genes and one concatenated and partitioned) was determined by using JModelTest version 2.1.10 under the BIC model [55]. To generate the concatenated and partitioned dataset, the program DnaSP 6.12.03 [56] was used. The mean p-distances between groups were calculated using MEGA 6.0 [52].
Different types of species delimitation analyses were carried out. In particular, we used ASAP (available at http://wwwabi.snv.jussieu.fr/public/abgd/) (accessed on 6 February 2022) to detect the barcode gap in the distribution of pairwise distances calculated on the COI sequence alignment [57,58]. ASAP analysis was performed on the in-group dataset using the Kimura Two Parameter (K2P) genetic distance and the default settings parameters. The Species Identifier program [59] was used to calculate maximum intraspecific and minimum interspecific distances (p-distance) and for clustering sequences based on pairwise distances. To assess the number of putative species in our COI DNA dataset, we used the Poisson Tree Processes model as implemented in the PTP web server [60] applied on the Bayesian tree. This species delimitation method outperforms other methods based on single-locus molecular phylogenies [60].
Two different phylogenetic analyses were carried out. Bayesian inference analysis (BI) was performed using the program MrBayes (v. 3.2.6) [61] by applying a Bayesian posterior likelihood methodology. Each of the four runs were conducted with four MCMCs (Markov Chain–Monte Carlo) for five million generations, a sample frequency of one tree per 1000 generations, and a burn-in of 25%. The maximum likelihood analysis was performed in raxmlGUI 1.5b2 [62], a graphical front-end for RAxML 8.2.1 [63], with 100 independent ML searches and 1000 bootstrap replicates. Felimare tricolor Cantraine, 1835 species was used as the outgroup for this analysis.
3. Results
A total of 53 Doriopsilla sequences were obtained from 18 specimens (Table 1), with 47 sequences derived from 16 individuals sampled in the Mediterranean Sea and six from two Eastern Atlantic Ocean individuals. Furthermore, 157 sequences from GenBank, including all available Doriopsilla species, were added to the final dataset, leading to a total of 209 sequences (Table 1).
Results from the mean p-distances (COI) calculated between the groups here investigated are reported in Table 2.
Table 2.
Mean p-distances of the COI mitochondrial marker between Doriopsilla species and other related genera analyzed in the present study.
All resulting trees were congruent with each other, differing only in the ability to resolve phylogenetic relationships at different taxonomic levels.
Bayesian inference (BI) and maximum likelihood (ML) analyses were generated from the single COI gene dataset consisting of 68 sequences with a length of 633 bp from 64 Doriopsilla and three Phyllidia specimens (Figure 1).
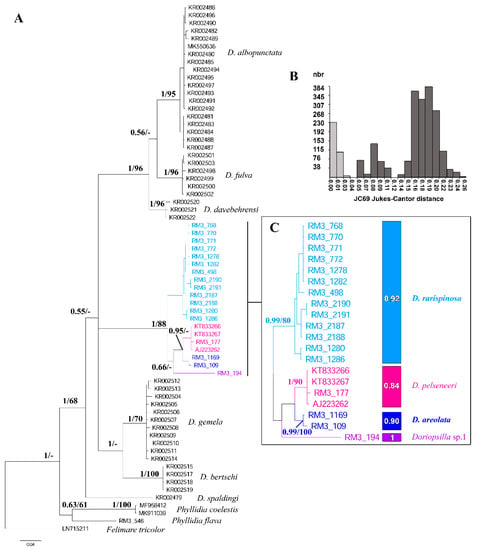
Figure 1.
Bayesian inference tree based on the COI sequence dataset along with results from species-delimitation analyses. (A) Bayesian COI tree. Numbers at nodes indicate Bayesian Posterior probability (BPP; left) and bootstrap support from the maximum likelihood analysis (BP; right). BPP < 0.50 and BP < 50% are not reported. (B) The histogram shows the distribution of the pairwise genetic distances (JC69) in intraspecific (left, light grey) and interspecific (right, dark grey) comparisons. (C) Species delimitation analyses on the Mediterranean Doriopsilla spp. The colored rectangles show the results from the ASAP analysis with the Bayesian support values from the PTP reported inside each rectangle. The ‘-’ symbol indicates unsupported values.
TPM1uf + I + G resulted to be the best evolutionary model for this single gene dataset. A clade formed by Doriopsilla and Phyllidia specimens was strongly supported by BI analysis and was clearly separated from the basal outgroup (Bayesian posterior probability (BPP) = 1, Bootstrap (BP) = <50). The monophyly of the genus Doriopsilla was strongly supported only for the Bayesian analysis (BPP = 1, BP = 68). Inside the Doriopsilla clade, the D. spaldingi Valdés and Behrens, 1998, specimen obtained from GenBank had no statistical support. Within the rest Doriopsilla clade, D. albopunctata Cooper, 1863, D. fulva MacFarland, 1905, and D. davebehrensi Hoover et al., 2015, were grouped in a well-supported monophyletic group (BPP = 1, BP = 96). Within this group, the monophyly of these three species was well supported with BPP = 1 and BP = 96 for D. davebehrensi and D. fulva and with BPP = 1 and BP = 95 for D. albopunctata, respectively. Another clade strongly supported by BI includes D. gemela Gosliner et al., 1999, and D. bertschi Hoover et al., 2015 (BPP = 1, BP = <50). Within this group, D. bertschi and D. gemela are each monophyletic with strong statistical support at BI and moderately supported for the ML of the latter species (BPP = 1 and BP = 100; and BPP = 1 and BP = 70, respectively). Considering the Mediterranean/Atlantic species group, they form a well-supported clade (BPP = 1, BP = 88), which are in turn divided into two groups: one grouping all the Mediterranean specimens previously identified as D. areolata and called from now on as D. rarispinosa (BPP = 1, BP = 95), and the second consisting in D. pelseneeri, D. areolata, and Doriopsilla sp. 1. Doriopsilla areolata (BPP = 0.99, BP = 100) and D. pelseneeri (BPP = 1, BP = 90) were grouped in a monophyletic clade (BPP = 0.95, BP = <50) that is a sister to Doriopsilla sp.1 with low support (BPP = 0.66, BP = <50).
The concatenated and partitioned (ConcDNA) dataset consisted of 79 sequences with a bp length of 1315 obtained from 74 specimens belonging to Doriopsilla, three belonging to Phyllidia, and the outgroup Felimare tricolor (Figure 2).

Figure 2.
Bayesian phylogenetic tree based on the ConcDNA dataset (H3, 16S, COI). Bayesian Posterior probability (BPP; left) and bootstrap support from the maximum likelihood analysis (BP; right) are indicated at each node. The ‘-’ symbol indicates unsupported values.
The evolutionary model used for COI and H3 calculations was HKY + G, while the TPM3uf + G was the one selected for the 16S. Doriopsilla and Phyllidia formed a clade that is well supported and separated from the basal outgroup (BPP = 1, BP = 100). All Doriopsilla species formed a strongly supported monophyletic clade (BPP = 1, BP = 100), with D. spaldingi as the basal species, which is a sister to all the other Doriopsilla spp. The remaining eight Doriopsilla were grouped in a monophyletic clade (BPP = 1, BP = 76) and, in turn, divided in two monophyletic clades. Doriopsilla bertschi (BPP = 1, BP = 100) and D. gemela (BPP = 1, BP = 99) formed one of these clades (BPP = 1, BP = 100). The second one is composed by the remaining Doriopsilla species (BPP = 1, BP =70). This second monophyletic clade was divided into two big clades. One monophyletic well-supported group (BPP = 1, BP = 100) comprises the D. janaina sister to a clade with D. fulva (BPP = 1, BP = 96), D. davebehrensi (BPP = 1, BP = 98) and D. albopunctata (BPP = 1, BP = 100). Doriopsilla fulva is the sister (BPP = 1, BP = 78) to the two latter species. The second big group (BPP = 1, BP = 97) is composed by the Mediterranean/Atlantic Doriopsilla spp. with D. miniata (BPP = 1, BP = 96) as the sister species to all the remaining ones. A group of non-supported sequences named as ‘D. areolata’ is the sister to a monophyletic clade (BPP = 0.99, BP = 50), which grouped D. areolata (BPP = 0.92, BP = 95), D. pelseneeri (BPP = 1, BP = 72), D. rarispinosa (BPP = 0.96, BP =87), and Doriopsilla sp.1.
The results of the phylogenetic analyses showed that the genus Doriopsilla formed a monophyletic clade separated from the outgroup. Furthermore, COI as well as ConcDNA dataset revealed D. areolata as a complex of at least three different species (Figure 1 and Figure 2). Based on this species division, the Adriatic population is, henceforth, listed as the bona fide D. areolata, since Bergh [27] used this population for the original description. The Western-central Mediterranean population formed a monophyletic clade that is from now on referred to as D. rarispinosa. The reassumption of this name for this clade is supported by the following criteria: (I) The original description and the subsequent redescription (Perrone 1989) show a high morphological correspondence with the specimens here examined. (II) D. reticulata (the first species whose description corresponds to our specimens and subsequently considered synonym of D. areolata) cannot be used, as already specified, as a junior homonymous of D. reticulata Quoy & Gaimard, 1832 because it is invalid. (III) D. pusilla described in the same year as D. rarispinosa was excluded considering that Pruvot-Fol herself assessed it to be of an uncertain genus, that it lacks a description allowing its definition, and probably the specimen, which is no longer available, was in the juvenile stage (3 mm). (IV) The third available name, D. fedalae, has been excluded, being an Atlantic species described in 1953, after D. rarispinosa, and it is quite different in external morphology. It is possible that this taxon may be used to name the Atlantic Doriopsilla species, which is up until now erroneously named as D. areolata. However, Atlantic populations need an in-depth systematic study that proceeds beyond the aims of the present work. Finally, an additional Doriopsilla species was revealed in the Atlantic Ocean; based on the results of COI and ConcDNA analyses, this specimen should be regarded as a separate species and is here provisionally reported as Doriopsilla sp.1.
4. Discussion
The range-wide sampling along the Mediterranean and Atlantic areas allowed revealing cryptic diversity within D. areolata species (Figure 3).
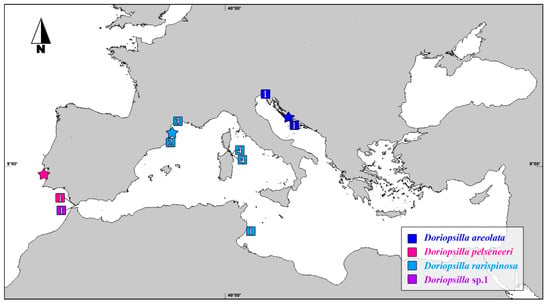
Figure 3.
Map showing the type localities (stars) and the provenance of the Atlantic/Mediterranean Doriopsilla specimens included in the present study (squares). The numbers within the squares refer to the number of individuals collected at each site.
In fact, even if the diversity of the Mediterranean nudibranchs is still far from being comprehensively unveiled, several steps forward have been made thanks to molecular methods [38]. The morphological and anatomical identification of nudibranch species is based on characteristics that are quite variable, difficult to compare, and not always present [38]. The radula, the hard structure part of the buccal apparatus typically used in nudibranchs for feeding, is lacking in the Doriopsilla species, failing one of the most important and diagnostic morphological characters. Therefore, particularly in the case of the Dendrodorididae, morphological investigations could leave room for errors and confusion in species identification. On the contrary, molecular identification methods seem to be the most powerful tool to reveal taxonomic misinterpretations made in the past and to highlight the presence of cryptic species across different marine animal groups, especially in the Mediterranean Sea where several cases of cryptic species and cases of endemism are continuously reported [38,43,44,45,46,64].
In the present work, molecular investigations implemented with the COI barcoder marker above the already used 16S and H3 markers and carried out on an extended dataset helped to unveil, as hypothesized, the presence of at least three different species under the D. areolata complex, two of which are D. areolata and D. rarispinosa, which are endemic to the Mediterranean Sea (Figure 1, Figure 2 and Figure 3).
Interestingly, no attention has been paid before to the possible separation between Mediterranean and Atlantic populations of D. areolata (e.g., in [5,15,28,33]). In fact, until now, only the populations from the Eastern Atlantic and Caribbean Sea have been studied but without including specimens from the type localities (i.e., the Adriatic Sea), thus ignoring an important and reference point for this species (e.g., [15,28]). Therefore, by filling the gap, this study shed some light on the Mediterranean population of D. areolata, revealing a new case of cryptic diversity. Results from phylogenetic analyses (considering single gene datasets and concatenated and partitioned data) were congruent with each other considering D. areolata, as previously conceived, as a complex of at least three distinct phylogenetic lineages.
Doriopsilla areolata specimens from the Adriatic Sea (RM3_109 and RM3_1169) formed one of these well-separated and monophyletic lineages and appeared to be more related to Atlantic D. pelseneeri than to the other Mediterranean species (Figure 1 and Figure 2). Doriopsilla areolata was described by Bergh in 1880 based on two specimens from Hvar Island in the Adriatic Sea. He described the ground coloration as light yellow–gray, with a reddish–brownish spot on the back (due to the peritoneum and viscera). Furthermore, he described that all over the mantle and up to the edge, there were thin whitish lines that branched and anastomosed to form an irregular grid with large and small meshes of different shapes. Bergh’s description is consistent with the morphology of the specimens sampled from the Adriatic Sea and here reported (Figure 4A,B).

Figure 4.
Mediterranean Doriopsilla species. (A,B) Doriopsilla areolata from the Adriatic Sea (in B, specimen with Voucher RM3_1169). Doriopsilla rarispinosa individuals from (C,D) Sardinia (Italy) (Voucher RM3_1278), (E) Catalonia (Spain) (Voucher RM3_2190), (F) Occitanie, France (Voucher RM3_2189), and (G) Tunisia (Voucher RM3_498).
The ground coloration was very light yellowish almost grayish transparent, and small white dots could be observed all over the body. On the dorsum, reddish-dark spots were present which originate from the organs lying underneath the epithelium. The dorsum was covered with small transparent tubercles, which could reach the mantle. An irregular grid pattern formed of prominent thin white lines could be observed on the dorsum and mantle. Since molecular evidence presented in this study indicates that D. areolata from the Adriatic Sea forms a separate species and considering that the first description of Bergh [27] was based on individuals from the same geographic area, it could be concluded that the species from the Adriatic Sea (the type locality of the species) must be considered, from now on, as the bona fide D. areolata. Interestingly D. areolata species seems to be geographically restricted to the Adriatic Sea since the only Doriopsilla reported to date from the nearby Ionian Sea is D. rarispinosa [65,66].
The second monophyletic clade resulting from the molecular investigation reported here corresponds to D. rarispinosa Pruvot-Fol, 1951, which in fact could not be considered anymore as the synonym of D. areolata and was reinstated here as a valid species name according to the principle of priority of the International Code of Zoological Nomenclature (ICZN Art. 23.3.5). This species was originally described from Banyuls-sur-Mer, France, Mediterranean Sea [67], and this locality is in line with the Western-central Mediterranean distribution we observed (Figure 3). This species shows a variable body color pattern from specimens displaying a yellow–orange background (Figure 4C–E) to others characterized by a strong dominance of the white color (Figure 4F). The Western Mediterranean populations correspond well with Pruvot-Fol’s description, the redescription by Perrone (1986) [68] and the image reported by Cattaneo-Vietti [69] (pag. 229 Figure 8). This correspondence with our samples is well represented in Figure 4D. It is noticeable that the coloration intensity of tubercles and lines and their texture do not change along a geographic gradient. In the middle of the dorsum, the underlying organs are visible as a dark grayish spot. It should also be noted that D. rarispinosa is the first available name, as Doris reticulata Schultz in Philippi, 1836 (despite its original description from a specimen collected in Palermo corresponds with our sample in Figure 4D), cannot be used, being an invalid junior synonym of Doris reticulata (Quoy & Gaimard, 1832). The Tunisian morphotype (Voucher RM3_498) (Figure 4G) shows a yellowish–white ground coloration of the dorsum. The mantle is whitish transparent, and in the dorsal area from the rhinophores to the gills, it assumes a yellowish color in correspondence with the internal organs. Flat tubercles, which are white bordered and transparent in the middle, are visible from the dorsum to the mantle edge. The most evident peculiarity of this morphotype is an irregular and distinctive web of fine white lines connected with the white spots on the dorsum and mantle, which are very different from the typical pattern of lines of D. areolata. Compared to the other D. rarispinosa morphotypes, the swarthy yellow–orange coloration of the rhinophores and their clearly rounded shape are outstanding and deserves further in-depth studies.
The third phylogenetic lineage reported here includes the Atlantic specimen (Voucher RM3_194) from Tarifa (Spain) that formed a well-supported clade in both COI and ConcDNA analyses (Figure 1 and Figure 2), suggesting that it could be ascribed to another species. The color of the specimen was orange–yellowish, and different larger reddish–brown areas were visible on the dorsum because of the organs lying under the epithelium. Moreover, in this case, thin white lines forming an irregular grid were observed. Since this study mainly focused on the Mediterranean D. areolata species complex, we provisionally referred to it as Doriopsilla sp. 1 (Figure 5), but an in depth-study of this possible additional Atlantic/Mediterranean species is advisable. However, the results from the 16S single gene dataset analysis (not shown) revealed a sister relationship with an individual from Cape Verde (E-ATL) reported in GenBank as ‘D. areolata’ (Voucher MNCN:15.05/23781 and 16S accession number KC171027), which deserves further investigations.
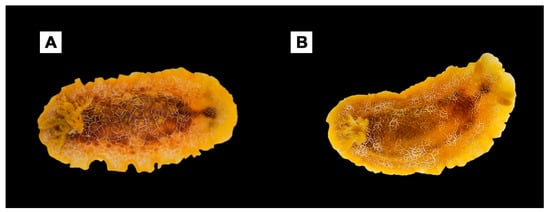
Figure 5.
Atlantic Doriopsilla species analyzed here. (A) Doriopsilla pelseneeri (Voucher RM3_177) from Tarifa, Andalusia, Spain (E-ATL). (B) Doriopsilla sp.1 (Voucher RM3_194) from Tarifa, Andalusia, Spain (E-ATL).
The closer relation between D. areolata from the Adriatic Sea and the Atlantic species, in contrast to D. rarispinosa from the Western Mediterranean basin, was unexpected. As a possible explanation, the propagules of the Atlantic Doriopsilla could have entered and colonized the Adriatic Sea. A possible pathway for larval dispersion could be the unidirectional surface current called ‘the Algerian current’, which is known to trap and transport larvae [70,71]. The current starts in the Atlantic Ocean and moves eastwards through the Mediterranean along the Northern African coast, bypassing the entire Western-central Mediterranean basin until it reaches the northern coast of Tunisia near Kelibia [72]. At this point, the current splits into two main branches: The first reaches Lampedusa, and the second follows the Sicilian coast towards the Ionian Sea [73]. From there, a northward cyclonic flow could have allowed the entry of larvae through the Otranto Channel, which are then further distributed along the Balkan coasts towards the Northern Adriatic [74]. The relationships between Mediterranean Doriopsilla species could be also related to the post-Messinian flood, which ended the Messinian Salinity Crisis (6.0–5.3 Ma) [75,76]. During this event, the Mediterranean Sea was filled with Atlantic Ocean Sea water, and consequently, Atlantic species invaded the Mediterranean basin. This flood could have also led to the introduction of a possible Doriopsilla ancestor. This introduction could have caused the formation of two different phyletic lineages: the Adriatic lineage, which is more related to the ancestor, and another one, which is endemic to the Mediterranean. The close relation of the Adriatic lineage to Atlantic Doriopsilla species would be likely since the Adriatic Sea is cooler and less salty than the rest of the Mediterranean Sea, creating conditions more similar to those of the Atlantic Ocean [77]. The abiotic isolation and the fact that the Adriatic Sea is semi-enclosed showed, in different studies [78,79], that genetic isolation can occur and lead to speciation processes [78,79]. In order to effectively address these possible scenarios, an extended investigation on other possible Mediterranean cryptic species with the possibility to calibrate the analyses to investigate the ancestral areas, is strongly recommended.
Finally, the separation during the evolution of the group from the common ancestor occurred recently, and this could explain the lack of lineage sorting observed in the H3 nuclear marker. Therefore, for future perspectives, it could be considered to explore alternative and fast-evolving markers as, for example, nuclear ITS2 instead of H3 to obtain additional useful information at a lower taxonomic scale [38]. Cryptic diversity is an intriguing challenge, especially regarding the Mediterranean Heterobranchia fauna that deserves an integrative systematic approach due to the recently separated species and the close evolutionary history with relative Atlantic fauna.
5. Conclusions
The results of this study revealed that D. areolata, as previously conceived, is a complex of cryptic species that includes at least two different species, D. areolata and D. rarispinosa, endemic to the Mediterranean Sea. In fact, this latter species is valid, and D. rarispinosa species name is here reinstated. Further phylogenetic analyses are still needed to investigate the possible additional species, provisionally reported here as Doriopsilla sp.1, whose occurrence may be searched both in the Eastern Atlantic Ocean, along the Spanish and Portuguese coasts, and in the Southern Mediterranean coasts of the Alboran Sea. Further research is also needed to solve the taxonomic problem of the Eastern Atlantic specimens, which is currently erroneously classified as Doriopsilla areolata.
Author Contributions
Conceptualization, G.F., E.T. and P.M.; methodology, G.F.; formal analysis, G.F., C.S. and E.T.; investigation, E.T., M.P., E.M. and P.G.; resources, E.T., M.P., E.M. and P.G.; data curation, G.F., C.S. and E.T.; writing—original draft preparation, G.F. and C.S.; writing—review and editing, G.F., E.T., M.P. and P.M.; supervision, P.M.; funding acquisition, G.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Italian Ministry of Education, University and Research MIUR, PON 2014–2020, grant number AIM 1848751-2, Linea 2, and the APC was funded by MIUR, PON 2014–2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This paper is dedicated to the late Barbara Camassa (Sistiana, Trieste), who collected samples from the Gulf of Trieste (Adriatic Sea) and whose contribution to the deepening of Mediterranean nudibranchs has yet to be fully developed. The authors wish to thank Stefano Piraino (Lecce, Italy) who helped improving the quality of this manuscript. Thanks is given to Lucas Cervera Currado (Cadiz, Spain) for his help in sampling the specimens from Tarifa. The collection of samples in the Marine Protected Area ‘Tavolara-Punta Coda Cavallo’ (Olbia, Italy) was authorized by the managing body, which is thanked here. GF wishes to thank the Scubalandia Team for technical underwater support. GF is supported by funds from the Italian Ministry of Education, University and Research (MIUR, PON 2014–2020, grant AIM 1848751-2, Linea 2).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Strong, E.E.; Gargominy, O.; Ponder, W.F.; Bouchet, P. Global Diversity of Gastropods (Gastropoda; Mollusca) in Freshwater. Hydrobiologia 2008, 595, 149–166. [Google Scholar] [CrossRef]
- Brown, K.M. 10—Mollusca: Gastropoda. In Ecology and Classification of North American Freshwater Invertebrates, 2nd ed.; Thorp, J.H., Covich, A.P., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 297–329. ISBN 978-0-12-690647-9. [Google Scholar]
- Wägele, H.; Willan, R.C. Phylogeny of the Nudibranchia. Zool. J. Linn. Soc. 2000, 130, 83–181. [Google Scholar] [CrossRef]
- Gavagnin, M.; Mollo, E.; Castelluccio, F.; Ghiselin, M.T.; Calado, G.; Cimino, G. Can Molluscs Biosynthesize Typical Sponge Metabolites? The Case of the Nudibranch Doriopsilla areolata. Tetrahedron 2001, 57, 8913–8916. [Google Scholar] [CrossRef]
- Valdés, A. Preliminary Molecular Phylogeny of the Radula-Less Dorids (Gastropoda: Opisthobranchia), Based on 16S MtDNA Sequence Data. J. Molluscan Stud. 2003, 69, 75–80. [Google Scholar] [CrossRef]
- Soares, C.; Calado, G. Spawn and Early Development of Two Sympatric Species of the Genus Doriopsilla (Opisthobranchia: Nudibranchia) with Contrasting Development Strategies. Rec. Aust. Mus. 2006, 69, 103–109. [Google Scholar] [CrossRef][Green Version]
- Healy, J.M.; Willan, R.C. Nudibranch Spermatozoa: Comparative Ultrastructure and Systematic Importance. Veliger 1991, 34, 134–165. [Google Scholar]
- Brunckhorst, D.J. The Systematics and Phylogeny of Phyllidiid Nudibranchs (Doridoidea). Rec. Aust. Mus. 1993, 16 (Suppl. 16), 1–107. [Google Scholar] [CrossRef]
- Valdés, A. Revisión de La Superfamilia Porodoridoidea Odhner En Franc, 1968 (Mollusca: Nudibranchia) En El Océano Atlántico. Ph.D. Thesis, Universidad de Oviedo, Oviedo, Spain, 1996. [Google Scholar]
- Valdés, Á.; Gosliner, T.M. Phylogeny of the Radula-Less Dorids (Mollusca, Nudibranchia), with the Description of a New Genus and a New Family. Zool. Scr. 1999, 28, 315–360. [Google Scholar] [CrossRef]
- Pruvot-Fol, A. Mollusques Opisthobranches. In Faune de France; Paul Lechevalier: Paris, France, 1954; Volume 58. [Google Scholar]
- Burn, R.F. Notes on a Collection of Nudibranchia (Gastropoda: Dorididae and Dendrodorididae) from South Australia with Remarks on the Species of Basedow and Hedley, 1905. Mem. Natl. Mus. Vic. 1962, 25, 149–171. [Google Scholar] [CrossRef]
- Valdés, A.; Ortea, J.; Ávila, C.; Ballesteros, M. Review of the Genus Dendrodoris (Ehrenberg, 1831) (Gastropoda: Nudibranchia) in the Atlantic Ocean. J. Molluscan Stud. 1996, 62, 1–31. [Google Scholar] [CrossRef]
- World Register of Marine Species. Available online: https://www.marinespecies.org (accessed on 30 March 2022).
- Valdés, A.; Ortea, J. Review of the Genus Doriopsilla Bergh, 1880 (Gastropoda: Nudibranchia) in the Atlantic Ocean. Veliger 1997, 40, 240–254. [Google Scholar]
- Yonow, N. Opisthobranchs from the Western Indian Ocean, with Descriptions of Two New Species and Ten New Records (Mollusca, Gastropoda). ZooKeys 2012, 197, 1–130. [Google Scholar] [CrossRef] [PubMed]
- Hoover, C.; Lindsay, T.; Goddard, J.H.R.; Valdés, Á. Seeing Double: Pseudocryptic Diversity in the Doriopsilla albopunctata–Doriopsilla gemela Species Complex of the North-Eastern Pacific. Zool. Scr. 2015, 44, 612–631. [Google Scholar] [CrossRef]
- Ballesteros, M.; Ortea, J. Contribución al Conocimiento de Los Dendrodorididae (Moluscos: Opistobranquios: Doridáceos) Del Litoral Ibérico; Publicaciones del Departamento de Zoología, Universidad de Barcelona, Facultad de Biología: Barcelona, Spain, 1980; Volume 5, pp. 25–37. [Google Scholar]
- d’Oliveira, M.P. Opisthobranches Du Portugal de La Collection de M. Paulino d’Oliveira. O Instituto Rev. Sci. E Litt. 1895, 42, 574–592. [Google Scholar]
- Sea Slug Forum. Available online: http://seaslugforum.net/showall/doripels (accessed on 30 March 2022).
- García-Gómez, J.C.; Cervera, J.L.; García, F.J.; López de la Cuadra, C.M. Resultados de La Campaña Internacional de Biología Marina “Ceuta-86”: Moluscos Opistobranquios. Boll. Malac. 1989, 25, 223–232. [Google Scholar]
- Domènech, A.; Ávila, C.; Ballesteros, M. Opisthobranch Molluscs from the Subtidal Trawling Grounds off Blanes (Girona, North-East Spain). J. Mar. Biol. Assoc. U. K. 2006, 86, 383–389. [Google Scholar] [CrossRef]
- Templado, J.; Guerra, A.; Bedoya, J.; Moreno, D.; Remón, J.M.; Maldonado, M.; Ramos, M.A. Fauna Marina Circalitoral Del Sur de La Península Ibérica: Resultados de La Campaña Oceanográfica “Fauna I”; Museo Nacional de Ciencias Naturales (CSIC): Madrid, Spain, 1993; ISBN 978-84-00-07380-0. [Google Scholar]
- Schick, K.L. Atlas Submarino de La Costa Del Sol; Marbella: Málaga, Spain, 1998; ISBN 84-605-7552-7. [Google Scholar]
- Trainito, E.; Doneddu, M. Nudibranchi del Mediterraneo, 2nd ed.; Il Castello: Milan, Italy, 2014; ISBN 978-88-6520-480-1. [Google Scholar]
- DORIS. Available online: https://doris.ffessm.fr/ref/specie/1320 (accessed on 30 March 2022).
- Bergh, L.S.R. Die Doriopsen Des Mittelmeeres. Jahrb. Dtsch. Malakozool. Ges. 1880, 7, 297–328. [Google Scholar]
- Valdés, Á.; Hamann, J. Two New Species of Doriopsilla from the Tropical Western Atlantic with Remarks on Cariopsillidae Ortea & Espinosa, 2005. Veliger 2008, 50, 210–218. [Google Scholar]
- Bergh, L.S.R. Die Nudibranchiata Holohepatica Porostomata. Verh. K. Kais. Zool. Bot. Ges. 1892, 42, 1–16. [Google Scholar]
- Young, D.K. The Functional Morphology of the Feeding Apparatus of Some Indo-West-Pacific Dorid Nudibranchs. Malacologia 1969, 9, 421–446. [Google Scholar]
- Edmunds, M. Opisthobranchiate Mollusca from Ghana. J. Moll. Stud. 1968, 38, 83–100. [Google Scholar] [CrossRef]
- Meyer, K.B. Dorid Nudibranchs of the Caribbean Coast of the Panama Canal Zone. Bull. Mar. Sci. 1977, 27, 299–307. [Google Scholar]
- Goodheart, J.; Valdés, Á. Re-Evaluation of the Doriopsilla areolata Bergh, 1880 (Mollusca: Opisthobranchia) Subspecies Complex in the Eastern Atlantic Ocean and Its Relationship to South African Doriopsilla miniata (Alder & Hancock, 1864) Based on Molecular Data. Mar. Biodivers. 2013, 43, 113–120. [Google Scholar] [CrossRef]
- Furfaro, G.; Modica, M.V.; Oliverio, M.; Mariottini, P. A DNA-Barcoding Approach to the Phenotypic Diversity of Mediterranean Species of Felimare Ev. Marcus & Er. Marcus, 1967 (Mollusca: Gastropoda), with a Preliminary Phylogenetic Analysis. Ital. J. Zool. 2016, 83, 195–207. [Google Scholar] [CrossRef]
- Furfaro, G.; Picton, B.E.; Martynov, A.V.; Mariottini, P. Diaphorodoris alba Portmann & Sandmeier, 1960 Is a Valid Species: Molecular and Morphological Comparison with D. luteocincta (M. Sars, 1870) (Gastropoda: Nudibranchia). Zootaxa 2016, 4193, 304–316. [Google Scholar] [CrossRef]
- Furfaro, G.; Mariottini, P.; Modica, M.V.; Trainito, E.; Doneddu, M.; Oliverio, M. Sympatric Sibling Species: The Case of Caloria elegans and Facelina quatrefagesi (Gastropoda: Nudibranchia). Sci. Mar. 2016, 80, 511–520. [Google Scholar] [CrossRef]
- Furfaro, G.; Salvi, D.; Mancini, E.; Mariottini, P. A Multilocus View on Mediterranean Aeolid Nudibranchs (Mollusca): Systematics and Cryptic Diversity of Flabellinidae and Piseinotecidae. Mol. Phylogenet. Evol. 2018, 118, 13–22. [Google Scholar] [CrossRef]
- Furfaro, G.; Salvi, D.; Trainito, E.; Vitale, F.; Mariottini, P. When Morphology Does Not Match Phylogeny: The Puzzling Case of Two Sibling Nudibranchs (Gastropoda). Zool. Scr. 2021, 50, 439–454. [Google Scholar] [CrossRef]
- Reece, J.S.; Castoe, T.A.; Parkinson, C.L. Historical Perspectives on Population Genetics and Conservation of Three Marine Turtle Species. Conserv. Genet. 2005, 6, 235–251. [Google Scholar] [CrossRef]
- Luttikhuizen, P.C.; Campos, J.; van Bleijswijk, J.; Peijnenburg, K.T.C.A.; van der Veer, H.W. Phylogeography of the Common Shrimp, Crangon crangon (L.) across Its Distribution Range. Mol. Phylogenet. Evol. 2008, 46, 1015–1030. [Google Scholar] [CrossRef]
- Triantaphyllidis, A.; Apostolidis, A.P.; Katsares, V.; Kelly, E.; Mercer, J.; Hughes, M.; Jørstad, K.E.; Tsolou, A.; Hynes, R.; Triantaphyllidis, C. Mitochondrial DNA Variation in the European Lobster (Homarus gammarus) throughout the Range. Mar. Biol. 2005, 146, 223–235. [Google Scholar] [CrossRef]
- Galarza, J.A.; Carreras-Carbonell, J.; Macpherson, E.; Pascual, M.; Roques, S.; Turner, G.F.; Rico, C. The Influence of Oceanographic Fronts and Early-Life-History Traits on Connectivity among Littoral Fish Species. Proc. Natl. Acad. Sci. USA 2009, 106, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Boissin, E.; Stöhr, S.; Chenuil, A. Did Vicariance and Adaptation Drive Cryptic Speciation and Evolution of Brooding in Ophioderma longicauda (Echinodermata: Ophiuroidea), a Common Atlanto-Mediterranean Ophiuroid? Mol. Ecol. 2011, 20, 4737–4755. [Google Scholar] [CrossRef] [PubMed]
- Claremont, M.; Reid, D.G.; Williams, S.T. Evolution of Corallivory in the Gastropod Genus Drupella. Coral Reefs 2011, 30, 977–990. [Google Scholar] [CrossRef]
- Barco, A.; Houart, R.; Bonomolo, G.; Crocetta, F.; Oliverio, M. Molecular Data Reveal Cryptic Lineages within the Northeastern Atlantic and Mediterranean Small Mussel Drills of the Ocinebrina edwardsii Complex (Mollusca: Gastropoda: Muricidae). Zool. J. Linn. Soc-Lond. 2013, 169, 389–407. [Google Scholar] [CrossRef]
- Lindsay, T.; Valdés, Á. The Model Organism Hermissenda crassicornis (Gastropoda: Heterobranchia) Is a Species Complex. PLoS ONE 2016, 11, e0154265. [Google Scholar] [CrossRef] [PubMed]
- Furfaro, G.; Mariottini, P. A New Dondice Marcus Er. 1958 (Gastropoda: Nudibranchia) from the Mediterranean Sea Reveals Interesting Insights into the Phylogenetic History of a Group of Facelinidae Taxa. Zootaxa 2020, 4731, 1–22. [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and Rapid Salt-Extraction of High Quality Genomic DNA for PCR-Based Techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Palumbi, S.R.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; Deptartment of Zoology and Kewalo Marine Laboratory, University of Hawaii: Honolulu, HI, USA, 2002. [Google Scholar]
- Colgan, D.J.; McLauchlan, A.; Wilson, G.D.F.; Livingston, S.P.; Edgecombe, G.D.; Macaranas, J.; Cassis, G.; Gray, M.R. Histone H3 and U2 SnRNA DNA Sequences and Arthropod Molecular Evolution. Aust. J. Zool. 1999, 46, 419–437. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of Phylogenies after Removing Divergent and Ambiguously Aligned Blocks from Protein Sequence Alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Posada, D. JModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Puillandre, N.; Modica, M.V.; Zhang, Y.; Sirovich, L.; BOISSELIER, M.-C.; Cruaud, C.; Holford, M.; Samadi, S. Large-Scale Species Delimitation Method for Hyperdiverse Groups. Mol. Ecol. 2012, 21, 2671–2691. [Google Scholar] [CrossRef]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA Barcoding and Taxonomy in Diptera: A Tale of High Intraspecific Variability and Low Identification Success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A General Species Delimitation Method with Applications to Phylogenetic Placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. RaxmlGUI: A Graphical Front-End for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.; Templado, J.; Oliverio, M.; Machordom, A. Hidden Mediterranean Biodiversity: Molecular Evidence for a Cryptic Species Complex within the Reef Building Vermetid Gastropod Dendropoma petraeum (Mollusca: Caenogastropoda). Biol. J. Linn. Soc. 2009, 96, 898–912. [Google Scholar] [CrossRef][Green Version]
- Furfaro, G.; Vitale, F.; Licchelli, C.; Mariottini, P. Two Seas for One Great Diversity: Checklist of the Marine Heterobranchia (Mollusca; Gastropoda) from the Salento Peninsula (South-East Italy). Diversity 2020, 12, 171. [Google Scholar] [CrossRef]
- Salento Sommerso. Available online: https://www.salentosommerso.it/pp_nudibranchia/Doriopsilla areolata/pp_Doriopsilla areolata.php (accessed on 8 March 2022).
- Pruvot-Fol, A. Études Des Nudibranches de La Méditerranée. Arch. Zool. Exp. Gén. 1951, 88, 1–80. [Google Scholar]
- Perrone, A.S. Il Genere Doriopsilla Bergh, 1880 in Mediterraneo: Descrizione Di Doriopsilla Rarispinosa Pruvot-Fol, 1951 (Opisthobranchia: Nudibranchia). Boll. Malac. 1986, 22, 97–112. [Google Scholar]
- Cattaneo-Vietti, R.; Chemello, R.; Giannuzzi-Savelli, R. Atlas of Mediterranean Nudibranchs; La Conchiglia: Rome, Italy, 1990. [Google Scholar]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the Open Mediterranean Sea: A Review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef]
- Elhmaidi, D.; Nefzi, H.; Carton, X.; Lili, T. Particle Dispersion in the Western Mediterranean Basin. Open Oceanogr. J. 2015, 4, 137–143. [Google Scholar] [CrossRef][Green Version]
- Pinardi, N.; Masetti, E. Variability of the Large Scale General Circulation of the Mediterranean Sea from Observations and Modelling: A Review. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 158, 153–173. [Google Scholar] [CrossRef]
- Poulain, P.-M.; Menna, M.; Mauri, E. Surface Geostrophic Circulation of the Mediterranean Sea Derived from Drifter and Satellite Altimeter Data. J. Phys. Oceanogr. 2012, 42, 973–990. [Google Scholar] [CrossRef]
- Valdés, Á.; Alexander, J.; Crocetta, F.; Yokeş, M.B.; Giacobbe, S.; Poursanidis, D.; Zenetos, A.; Lucas Cervera, J.; Caballer, M.; Galil, B.S.; et al. The Origin and Dispersal Pathway of the Spotted Sea Hare Aplysia dactylomela (Mollusca: Opisthobranchia) in the Mediterranean Sea. Aquat. Invasions 2013, 8, 427–436. [Google Scholar] [CrossRef]
- Kyle, C.J.; Boulding, E.G. Comparative Population Genetic Structure of Marine Gastropods (Littorina Spp.) with and without Pelagic Larval Dispersal. Mar. Biol. 2000, 137, 835–845. [Google Scholar] [CrossRef]
- Blanc, P.-L. The Opening of the Plio-Quaternary Gibraltar Strait: Assessing the Size of a Cataclysm. Geodin. Acta 2002, 15, 303–317. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Velarque, M.; Çinar, M.E.; García Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien Species in the Mediterranean Sea by 2010. A Contribution to the Application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial Distribution. Mediterr. Mar. Sci. 2010, 11, 381. [Google Scholar] [CrossRef]
- Maggio, T.; Lo Brutto, S.; Garoia, F.; Tinti, F.; Arculeo, M. Microsatellite Analysis of Red Mullet Mullus barbatus (Perciformes, Mullidae) Reveals the Isolation of the Adriatic Basin in the Mediterranean Sea. ICES J. Mar. Sci. 2009, 66, 1883–1891. [Google Scholar] [CrossRef]
- Ragionieri, L.; Schubart, C.D. Population Genetics, Gene Flow, and Biogeographical Boundaries of Carcinus estuarii (Crustacea: Brachyura: Carcinidae) along the European Mediterranean Coast. Biol. J. Linn. Soc. 2013, 109, 771–790. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).