
Epizoic Rotifers and Microcrustaceans on Bivalves of Different Size and Behavior



Abstract
:1. Introduction
2. Materials and Methods
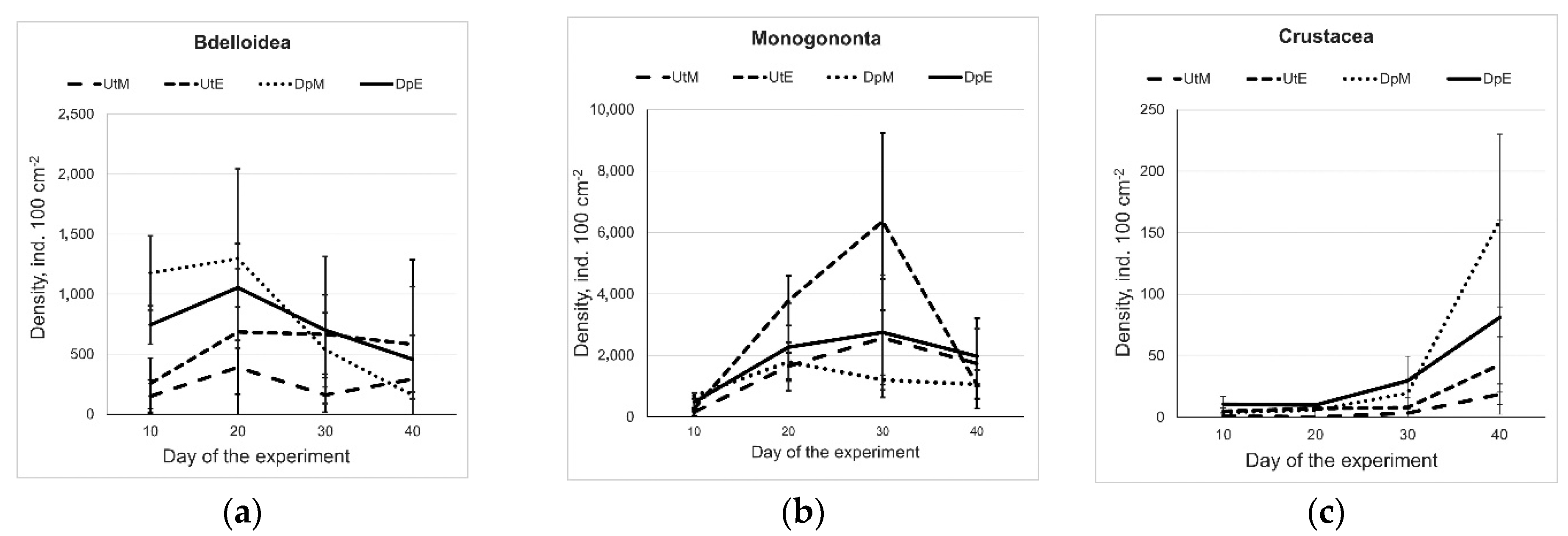
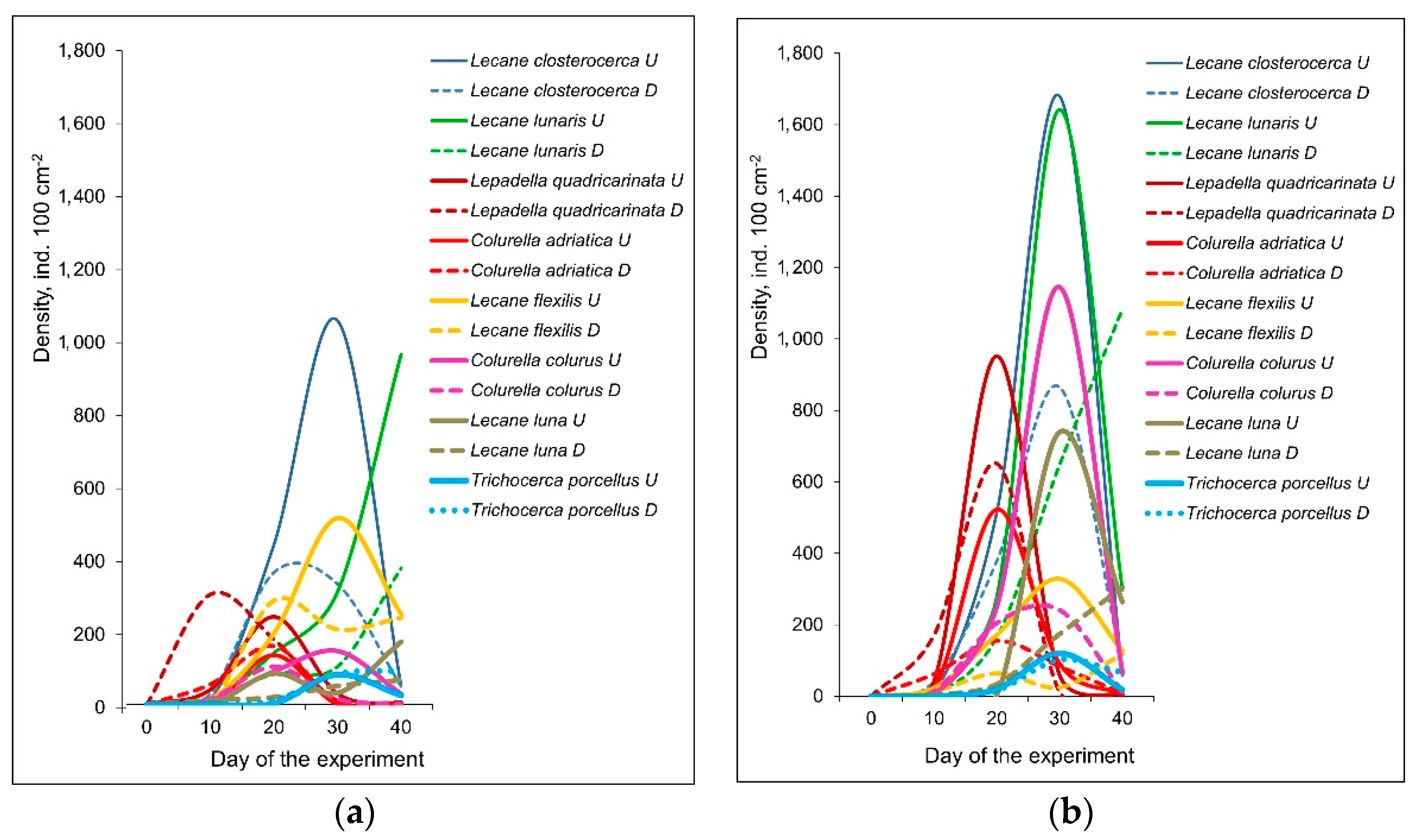
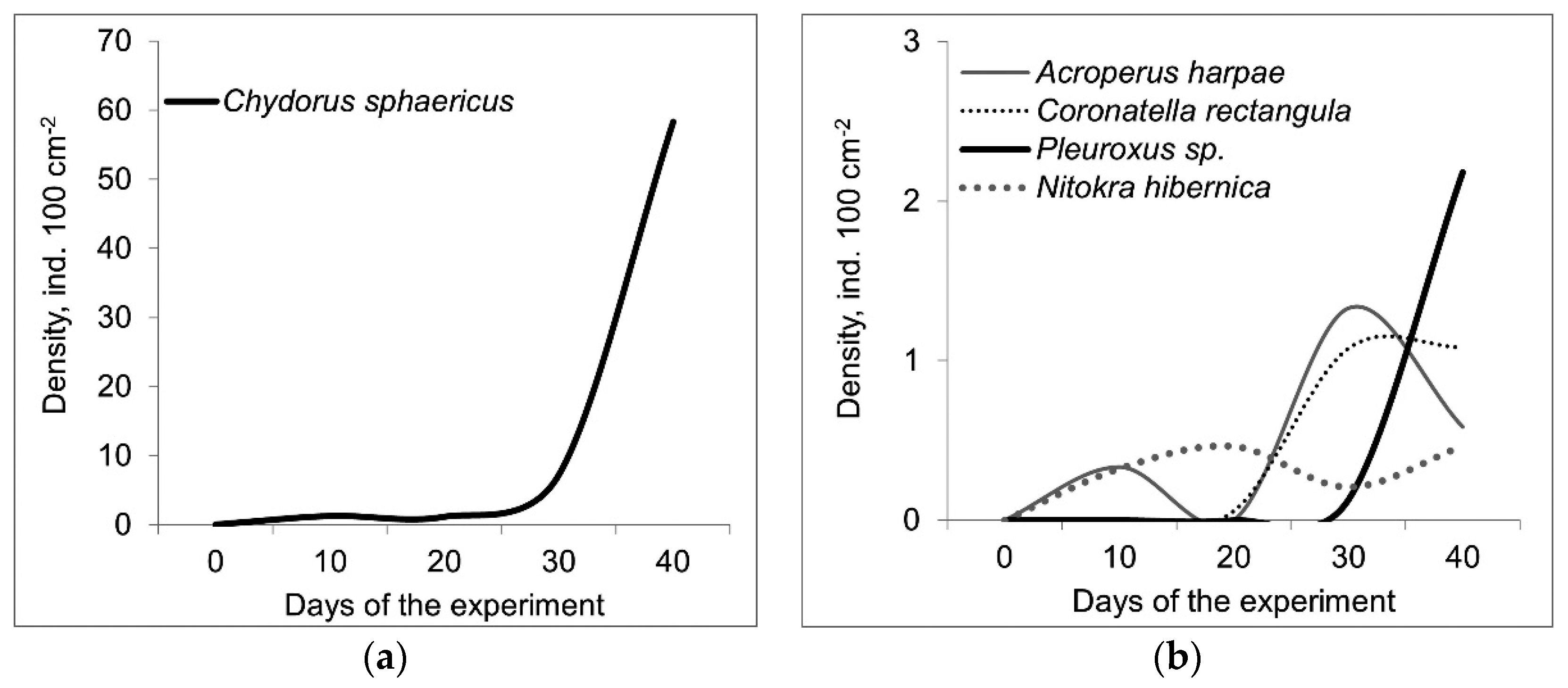
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vaughn, C.C.; Hakenkamp, C.C. The functional role of burrowing bivalves in freshwater ecosystems. Freshw. Biol. 2001, 46, 1431–1446. [Google Scholar] [CrossRef] [Green Version]
- May, L. Epizoic and parasitic rotifers. Hydrobiologia 1989, 186/187, 59–67. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J.; Karpowicz, M. Epizoic rotifers on Dreissena polymorpha in relation to biotic factors. Hydrobiologia 2019, 828, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, T.; Kotov, A.A.; Fernández-Leborans, G. A checklist of epibiotic ciliates (Peritrichia and Suctoria) on the cladoceran crustaceans. Biologia 2013, 68, 439–447. [Google Scholar] [CrossRef]
- Sagrario, G.; de los Angeles, M.; Balseiro, E.; Ituarte, R.; Spivak, E. Macrophytes as refuge or risky area for zooplankton: A balance set by littoral predacious macroinvertebrates. Freshw. Biol. 2009, 54, 1042–1053. [Google Scholar] [CrossRef]
- Špoljar, M.; Dražina, T.; Lajtner, J.; Duić Sertić, M.; Radanović, I.; Wallace, R.; Matulić, D.; Tomljanović, T. Zooplankton assemblage in four temperate shallow waterbodies in association with habitat heterogeneity and alternative states. Limnologica 2018, 71, 51–61. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J. Epizoic crustaceans (Cladocera, Copepoda) on Dreissena polymorpha (Pallas) in relation to biotic factors. Fundam. Appl. Limnol. 2018, 191, 169–174. [Google Scholar] [CrossRef]
- Bołtruszko, J. Epizoic communities of Rotifera on freshwater bivalves. Oceanol. Hydrobiol. St. 2011, 39, 75–82. [Google Scholar] [CrossRef]
- Schwalb, A.N.; Pusch, M.T. Horizontal and vertical movements of unionid mussels in a lowland river. J. N. Am. Benthol. Soc. 2007, 26, 261–272. [Google Scholar] [CrossRef]
- Kutikova, L.A. Kolovratki Fauny SSSR; [Rotifer fauna of USSR] Izd. Nauka: Leningrad, Russian, 1970; p. 742. (In Russian) [Google Scholar]
- Koste, W. Rotatoria, die Rädertiere Mitteleuropas; Uberordnung Monogononta, 1978. Revision after M. Voigt 1956/ 57, 2; Borntraeger: Stuttgart, Germany, 1978. [Google Scholar]
- Nogrady, T.; Pourriot, R.; Segers, H. Rotifera 3: Notommatidae and Scaridiidae. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Dumont, H.J., Eds.; SPB Academic Publishing b.v.: Amsterdam, NY, USA, 1995; Volume 8, p. 248. [Google Scholar]
- Segers, H. Rotifera 2. The Lecanidae (Monogononta). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H.J., Nogrady, T., Eds.; SPB Academic Publishing b.v.: Amsterdam, NY, USA, 1995; Volume 6, p. 226. [Google Scholar]
- De Smet, W.H. Rotifera 4. The Proalidae (Monogononta). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Ed.; SPB Academic Publishing b.v.: Amsterdam, NY, USA, 1996; Volume 9, p. 102. [Google Scholar]
- De Smet, W.H.; Pourriot, R. Rotifera. The Dicranophoridae and the Ituridae(Monogononta). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Ed.; SPB Academic Publishing b.v.: Amsterdam, NY, USA, 1997; Volume 12, p. 344. [Google Scholar]
- Nogrady, T.; Segers, H. Asplanchnidae, Gastropodidae, Lindiidae, Microcodidae, Synchaetidae, Trochosphaeridae and Filinia. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Segers, H., Eds.; SPB Academic Publishing b.v.: Amsterdam, NY, USA, 2002; Volume 18, p. 264. [Google Scholar]
- Bielańska-Grajner, I.; Ejsmont-Karabin, J.; Radwan, S. Rotifers (Rotifera, Monogononta). In Freshwater Fauna of Poland; Wydawnictwo Uniwersytetu Łódzkiego: Łódż, Poland, 2015; Volume 32, p. 577. [Google Scholar]
- Jersabek, C.D.; Leitner, M.F. The Rotifer World Catalog. World Wide Web Electronic Publication, 2013. Available online: http://www.rotifera.hausdernatur.at/ (accessed on 8 April 2022).
- Janetzky, W.; Enderle, R.; Noodt, W. Crustacea-1: Copepoda: Gelyelloida und Harpacticoida. In Süßwasserfauna von Mitteleuropa; Schwoerbel, J., Zwick, P., Eds.; Gustav Fischer Verlag: Stuttgart, Germany, 1996; Volume 8, p. 227. [Google Scholar]
- Flößner, D. Die Haplopoda und Cladocera (Ohne Bosminidae) Mitteleuropas; Backhuys: Leiden, The Netherlands, 2000; p. 428. [Google Scholar]
- Błędzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe; Springer: Cham, Switzerland, 2016; p. 918. [Google Scholar]
- Karpowicz, M.; Ejsmont-Karabin, J. Diversity and Structure of Pelagic Zooplankton (Crustacea, Rotifera) in NE Poland. Water 2021, 13, 456. [Google Scholar] [CrossRef]
- Brodzińska, B.; Jańczak, J.; Kowalik, A.; Sziwa, R. Atlas Jezior Polski, Tom III [The Atlas of Polish Lakes, Volume 3]; Wydawnictwo Naukowe: Poznań, Poland, 1999; p. 240. (In Polish) [Google Scholar]
- Ejsmont-Karabin, J.; Karpowicz, M. Rotifera in lake subhabitats. Aquat. Ecol. 2021, 55, 1285–1296. [Google Scholar] [CrossRef]
- Jonsson, A.; Bertilsson, A.; Rydgård, M. Spatial distribution and age structure of the freshwater unionid mussels Anodonta anatina and Unio tumidus: Implications for environmental monitoring. Hydrobiologia 2013, 711, 61–70. [Google Scholar] [CrossRef]
- Covich, A.P.; Palmer, M.A.; Crowl, T.A. The Role of Benthic Invertebrate Species in Freshwater Ecosystems. Zoobenthic species influence energy flows and nutrient cycling. BioScience 1999, 49, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Zaiko, A.; Daunys, D.; Olenin, S. Habitat engineering by the invasive zebra mussel Dreissena polymorpha (Pallas) in a boreal coastal lagoon: Impact on biodiversity. Helgol. Mar. Res. 2008, 63, 85–94. [Google Scholar] [CrossRef]
- Kalinowska, K.; Ejsmont-Karabin, J.; Rybak, J.I. The role of lake shore sand deposits as bank of ciliate, rotifer and crustacean resting forms: Experimental approach. Pol. J. Ecol. 2010, 58, 323–332. [Google Scholar]
- Fryer, G. Evolution and adaptive radiation in the Chydoridae (Crustacea: Cladocera): A study in comparative functional morphology and ecology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1968, 254, 221–385. [Google Scholar] [CrossRef]
- Williams, J.B. Temporal and spatial patterns of abundance of the Chydoridae (Cladocera) in Lake Itasca, Minnesota. Ecology 1982, 63, 345–353. [Google Scholar] [CrossRef]
- Vijverberg, J.; Boersma, M. Long-term dynamics of small-bodied and large-bodied cladocerans during the eutrophication of a shallow reservoir, with special attention for Chydorus sphaericus. Hydrobiologia 1997, 360, 233–242. [Google Scholar] [CrossRef]
- Górniak, A.; Karpowicz, M. Development of crustacean plankton in a shallow, polyhumic reservoir in the first 20 years after impoundment (northeast Poland). Inland Waters 2014, 4, 311–318. [Google Scholar] [CrossRef]
- Makhutova, O.N.; Protasov, A.A.; Gladyshev, M.I.; Sylaieva, A.A.; Sushchik, N.N.; Morozovskaya, I.A.; Kalachova, G.S. Feeding spectra of bivalve mollusks Unio and Dreissena from Kanevskoe Reservoir, Ukraine: Are they food competitors or not? Zool. Stud. 2013, 52, 56. Available online: http://www.zoologicalstudies.com/content/52/1/56 (accessed on 23 March 2022). [CrossRef] [Green Version]
- Nichols, S.J.; Garling, D. Food-web dynamics and trophic-level interactions in a multispecies community of freshwater unionids. Can. J. Zool. 2000, 78, 871–882. [Google Scholar] [CrossRef]
- Cole, J.J.; Solomon, C.T. Terrestrial support of zebra mussels and Hudson River food web: A multi-isotope, Bayesian analysis. Limnol. Oceanogr. 2012, 57, 1802–1815. [Google Scholar] [CrossRef] [Green Version]
- Kuczyńska-Kippen, N.; Nagengast, B. The influence of the spatial structure of hydromacrophytes and differentiating habitat on the structure of the rotifer and cladoceran communities. Hydrobiologia 2006, 559, 203–212. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J.; Strzałek, M. Biodiversity of zooplankton (Rotifera and Crustacea) in water soldier (Stratiotes aloides) habitats. Biologia 2016, 71, 563–573. [Google Scholar] [CrossRef]
- Lawton, J.H.; Brown, V.K. Redundancy in ecosystems. In Biodiversity and Ecosystem Function; Schulze, E.-D., Mooney, H.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; pp. 255–270. [Google Scholar]
- Loreau, M. Does functional redundancy exist? Oikos 2004, 104, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Levin, S. Multiple Scales and the Maintenance of Biodiversity. Ecosystems 2000, 3, 498–506. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Sousa, R.; Geist, J.; Aldridge, D.C.; Araujo, R.; Bergengren, J.; Bespalaya, Y.; Bódis, E.; Burlakova, L.; van Damme, D.; et al. Conservation status of freshwater mussels in Europe: State of the art and future challenges. Biol. Rev. 2017, 92, 572–607. [Google Scholar] [CrossRef]
- Aldridge, D.C.; Fayle, T.M.; Jackson, N. Freshwater mussel abundance predicts biodiversity in UK lowland rivers. Aquat. Conserv. 2007, 17, 554–564. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Rotifer Species Richness | Microcrustacean Species Richness | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Day 10 | Day 20 | Day 30 | Day 40 | Day 10 | Day 20 | Day 30 | Day 40 | ||
| UtM | Total | 19 | 43 | 36 | 25 | 1 | 0 | 3 | 5 |
| Mean | 10 ± 5 | 25 ± 8 | 25 ± 4 | 14 ± 1 | 0.3 ± 0.6 | 0 | 1.0 ± 1.0 | 2.3 ± 0.6 | |
| UtE | Total | 24 | 55 | 44 | 26 | 4 | 4 | 5 | 6 |
| Mean | 13 ± 3 | 33 ± 7 | 27 ± 2 | 15 ± 4 | 1.3 ± 0.6 | 1.3 ± 0.6 | 1.7 ± 1.5 | 2.7 ± 0.6 | |
| DpM | Total | 25 | 42 | 41 | 35 | 2 | 2 | 4 | 7 |
| Mean | 18 ± 6 | 27 ± 4 | 28 ± 2 | 22 ± 4 | 1.0 ± 1.0 | 1.0 ± 1.0 | 2.0 ± 1.0 | 3.7 ± 0.6 | |
| DpE | Total | 25 | 44 | 43 | 37 | 5 | 5 | 7 | 9 |
| Mean | 14 ± 3 | 27 ± 10 | 26 ± 9 | 18 ± 4 | 1.3 ± 0.6 | 2.0 ± 0.7 | 3.7 ± 1.5 | 3.7 ± 0.6 | |
| Day 10 | Day 20 | Day 30 | Day 40 | ||
|---|---|---|---|---|---|
| UtM | Lecane pumila | - | 20 | 57 | - |
| Lophocharis naias | 3 | - | - | - | |
| Wulfertia sp. | - | - | 140 | - | |
| UtE | Lecane pumila | 64 | 28 | 130 | - |
| Lophocharis naias | - | 7 | - | - | |
| DpM | Lecane pumila | 28 | 4 | - | 4 |
| Lophocharis naias | 165 | 25 | 15 | 31 | |
| DpE | Lecane pumila | - | - | 9 | 6 |
| Lophocharis naias | 117 | 61 | 14 | 6 | |
| Wulfertia sp. | - | - | 4 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ejsmont-Karabin, J.; Karpowicz, M.; Feniova, I. Epizoic Rotifers and Microcrustaceans on Bivalves of Different Size and Behavior. Diversity 2022, 14, 293. https://doi.org/10.3390/d14040293
Ejsmont-Karabin J, Karpowicz M, Feniova I. Epizoic Rotifers and Microcrustaceans on Bivalves of Different Size and Behavior. Diversity. 2022; 14(4):293. https://doi.org/10.3390/d14040293
Chicago/Turabian StyleEjsmont-Karabin, Jolanta, Maciej Karpowicz, and Irina Feniova. 2022. "Epizoic Rotifers and Microcrustaceans on Bivalves of Different Size and Behavior" Diversity 14, no. 4: 293. https://doi.org/10.3390/d14040293
APA StyleEjsmont-Karabin, J., Karpowicz, M., & Feniova, I. (2022). Epizoic Rotifers and Microcrustaceans on Bivalves of Different Size and Behavior. Diversity, 14(4), 293. https://doi.org/10.3390/d14040293