
Time, Mediated through Plant Versatility, Is a Better Predictor of Medicinal Status of Alien Plants

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.2.1. South Africa’s Alien Woody Flora and Residence Time
2.2.2. Availability, Versatility, and Medicinal Use Data of Alien Woody Plants
2.3. Data Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harshberger, J.W. The Purposes of Ethno-Botany. Bot. Gaz. 1896, 21, 146–154. [Google Scholar] [CrossRef]
- Alexiades, M.N. Selected Guidelines for Ethnobotanical Research: A Field Manual; The New York Botanical Garden: Bronx, NY, USA, 1996; Volume 10. [Google Scholar]
- Soejarto, D.D.; Fong, H.H.S.; Tan, G.T.; Zhang, H.J.; Ma, C.Y.; Franzblau, S.G.; Gyllenhaal, C.; Riley, M.C.; Kadushin, M.R.; Pezzuto, J.M.; et al. Ethnobotany/ethnopharmacology and mass bioprospecting: Issues on intellectual property and benefit-sharin. J. Ethnopharmacol. 2005, 100, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Balick, M. Transforming ethnobotany for the new millennium. Ann. Mol. Bot. Gard. 1996, 83, 58–66. [Google Scholar] [CrossRef]
- Etkin, N.L. Ethnopharmacology: Biobehavioral approaches in the anthropological study of indigenous medicines. Annu. Rev. Anthropol. 1988, 17, 23–42. [Google Scholar] [CrossRef]
- Hurrell, J.A.; Albuquerque, U.P. Is ethnobotany an ecological science? Steps towards a complex ethnobotany. Ethnobiol. Conserv. 2012, 1, 1–16. [Google Scholar]
- Soldati, G.T.; Albuquerque, U.P. A new application for the optimal foraging theory: The extraction of medicinal plants. Evid.-Based Complementary Altern. Med. 2012, 2012, 364564. [Google Scholar] [CrossRef]
- Saslis-Lagoudakis, C.H.; Hawkins, J.A.; Greenhill, S.J.; Pendry, C.A.; Watson, M.F.; Tuladhar-Douglas, W.; Baral, S.R.; Savolainen, V. The evolution of traditional knowledge: Environment shapes medicinal plant use in Nepal. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132768. [Google Scholar] [CrossRef]
- Yessoufou, K.; Daru, B.H.; Muasya, A.M. Phylogenetic exploration of commonly used medicinal plants in South Africa. Mol. Ecol. Resour. 2015, 15, 405–413. [Google Scholar] [CrossRef]
- Cámara-Leret, R.; Faurby, S.; Macía, M.J.; Balslev, H.; Göldel, B.; Svenning, J.-C.; Kissling, W.D.; Rønsted, N.; Saslis-Lagoudakis, C.H. Fundamental species traits explain provisioning services of tropical American palms. Nat. Plants 2017, 3, 16220. [Google Scholar] [CrossRef]
- Hart, G.; Gaoue, O.G.; de la Torre, L.; Navarrete, H.; Muriel, P.; Macía, M.J.; Balslev, H.; León-Yánez, S.; Jørgensen, P.; Duffy, D.C. Availability, diversification and versatility explain human selection of introduced plants in Ecuadorian traditional medicine. PLoS ONE 2017, 12, e0184369. [Google Scholar] [CrossRef]
- Teixidor-Toneu, I.; Jordan, F.M.; Hawkins, J.A. Comparative phylogenetic methods and the cultural evolution of medicinal plant use. Nat. Plants 2018, 4, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, U.P.; de Medeiros, P.M.; Ferreira Júnior, W.S.; da Silva, T.C.; da Silva, R.R.V.; Gonçalves-Souza, T. Socialecological theory of maximization: Basic concepts and two initial models. Biol. Theory 2019, 14, 73–85. [Google Scholar] [CrossRef]
- Seyler, B.C.; Gaoue, O.G.; Tang, Y.; Duffy, D.C. Understanding knowledge threatened by declining wild orchid populations in an urbanizing China (Sichuan). Environ. Conserv. 2019, 46, 318–325. [Google Scholar] [CrossRef]
- Gaoue, O.G.; Coe, M.A.; Bond, M.; Hart, G.; Seyler, B.C.; McMillen, H. Theories and Major Hypotheses in Ethnobotany. Econ. Bot. 2017, 71, 269–287. [Google Scholar] [CrossRef]
- Gaoue, O.G.; Moutouama, J.K.; Coe, M.A.; Bond, M.O.; Green, E.; Sero, N.B.; Bezeng, B.S.; Yessoufou, K. Methodological advances for hypothesis driven Ethnobiology. Biol. Rev. 2021, 96, 2281–2303. [Google Scholar] [CrossRef]
- Ford, R.I. The Nature and Status of Ethnobotany; University of Michigan: Ann Arbor, MI, USA, 1978. [Google Scholar]
- Phillips, O.; Gentry, A.H. The useful plants of Tambopata, Peru: II. Additional hypothesis testing in quantitative ethnobotany. Econ. Bot. 1993, 47, 33–43. [Google Scholar] [CrossRef]
- Gaoue, O.G.; Yessoufou, K.; Mankga, L.; Vodouhe, F. Phylogeny reveals non-random medicinal plant organs selection by local people in Benin. Plants People Planet 2021, 3, 710–720. [Google Scholar] [CrossRef]
- Krebs, C.J. Hypothesis testing in ecology. In Research Techniques in Animal Ecology; Columbia University Press: New York, NY, USA, 2000; pp. 1–14. [Google Scholar]
- Voeks, R.A. Disturbance pharmacopoeias: Medicine and myth from the humid tropics. Ann. Assoc. Am. Geogr. 2004, 94, 868–888. [Google Scholar]
- Alencar, N.L.; de Sousa Araújo, T.A.; Amorim, E.L.C.; Albuquerque, U.P. The inclusion and selection of medicinal plants in traditional pharmacopoeias—Evidence in support of the diversification hypothesis. Econ. Bot. 2010, 64, 68–79. [Google Scholar] [CrossRef]
- Yessoufou, K.; Ambani, A.E.; Elansary, H.O.; Gaoue, O.G. Alien woody plants are more versatile than native, but both share similar therapeutic redundancy in South Africa. PLoS ONE 2021, 16, e0260390. [Google Scholar] [CrossRef]
- Bennett, B.C.; Prance, G.T. Introduced plants in the indigenous pharmacopoeia of northern South America. Econ. Bot. 2000, 1, 90–102. [Google Scholar] [CrossRef]
- Fabricant, D.S.; Farnsworth, N.R. The Value of Plants Used in Traditional Medicine for Drug Discovery. Environ. Health Perspect. 2001, 109, 69–75. [Google Scholar] [PubMed]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; Garcia, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human-induced Species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.R.; Ivey, P.; Manyama, P.; Nanni, I. A new national unit for invasive species detection, assessment and eradication planning. S. Afr. J. Sci. 2013, 109, 111. [Google Scholar] [CrossRef]
- Poe, M.R.; Norman, K.C.; Levin, P.S. Cultural dimensions of socioecological systems: Key connections and guiding principles for conservation in coastal environments. Conserv. Lett. 2014, 7, 166–175. [Google Scholar] [CrossRef]
- Winter, K.B.; Ticktin TQuazi, S.A. Biocultural restoration in Hawai’I also achieves core conservation goals. Ecol. Soc. 2020, 25, 26. [Google Scholar]
- Bennett, N.J.; Roth, R.; Klain, S.C.; Chan, K.; Christie, P.; Clark, D.A.; Cullman, G.; Curran, D.; Durbin, T.J.; Epstein, G.; et al. Conservation social science: Understanding and integrating human dimensions to improve conservation. Biol. Conserv. 2017, 205, 93–108. [Google Scholar] [CrossRef]
- Germishuizen, G.; Meyer, N.L. Plants of Southern Africa: An Annotated Checklist; National Botanical Institute: Pretoria, South Africa, 2003; Volume 14, pp. 192–196. [Google Scholar]
- Bezeng, S.B.; Morales-Castilla, I.; Van der Bank, M.; Yessoufou KDaru, B.H.; Davies, J. Climate change may reduce the spread of non-native species. Ecosphere 2017, 8, e01694. [Google Scholar] [CrossRef]
- Henderson, L. Alien weeds and invasive plants. In Plant Protection Research Institute Handbook No. 12; Agricultural Research Council: Pretoria, South Africa, 2001. [Google Scholar]
- Henderson, L. Invasive, naturalized and casual alien plants in southern Africa: A summary based on the Southern African Plant Invaders Atlas (SAPIA). Bothalia 2007, 37, 215–248. [Google Scholar] [CrossRef]
- Coates-Palgrave, M.; Coates-Palgrave, K. Trees of Southern Africa, 3rd ed.; Struik Nature: Cape Town, South Africa, 2002. [Google Scholar]
- Yessoufou, K.; Ambani, A.E. Are Introduced Alien Species More Predisposed to Invasion in Recipient Environments If They Provide a Wider Range of Services to Humans? Diversity 2021, 13, 553. [Google Scholar] [CrossRef]
- Global Biodiversity Information Facility. 2017. Available online: https://www.gbif.org/occurrence/search?taxon_key=3880247 (accessed on 25 June 2017).
- Van Wyk, B.; van Wyk, P. Field Guide to Trees of Southern Africa, 2nd ed.; Struik Publisher: Cape Town, South Africa, 2013. [Google Scholar]
- Glen, H.; Van Wyk, B. Guide to Trees Introduced into Southern Africa; Penguin Random House South Africa: Cape Town, South Africa, 2016. [Google Scholar]
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Fan, Y.; Chen, J.; Shirkey, G.; John, R.; Wu, S.R.; Park, H.; Shao, C. Applications of structural equation modeling (SEM) in ecological studies: An updated review. Ecol. Processes 2016, 5, 19. [Google Scholar] [CrossRef]
- Schermelleh-Engel, K.; Moosbrugger, H.; Müller, H. Evaluating the Fit of Structural Equation Models: Tests of Significance and Descriptive Goodness-Of-Fit Measures. Methods Psychol. Res. 2003, 8, 23–74. [Google Scholar]
- O’Hara, R.B.; Kotze, D.J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef]
- Albuquerque, U.P. Re-examining hypotheses concerning the use and knowledge of medicinal plants a study in the Caatinga vegetation of NE Brazil. J. Ethnobiol. Ethnomed. 2006, 2, 1–10. [Google Scholar] [CrossRef]
- Medeiros, P.M. Why is change feared? Exotic species in traditional pharmacopoeias. Ethnobiol. Conserv. 2013, 2, 1–5. [Google Scholar]
- Bletter, N.; Satdichan, M.; Sounthala, L.; Satdichan, D.; Sudmoon, R.; Noikotr, K.; Thani, T.; Yongvanit, S.; Chaveerach, A. Plants up for adoption: Why do Southeast Asians so readily accept introduced plants into use, especially those from the Neotropics? In Proceedings of the 2010 Southeast Asian Geography Association, Hanoi, Vietnam, 23–26 November 2010; pp. 1–17. [Google Scholar]
- Lozano, A.; Araújo, E.L.; Medeiros, M.F.T.; Albuquerque, U.P. The apparency hypothesis applied to a local pharmacopoeia in the Brazilian northeast. J. Ethnobiol. Ethnomed 2014, 10, 2. [Google Scholar] [CrossRef]
- Phillips, O.; Gentry, A.H. The useful plants of Tambopata, Peru: I. Statistical hypotheses tests with a new quantitative technique. Econ. Bot. 1993, 47, 15–32. [Google Scholar] [CrossRef]
- Albuquerque, U.P. Quantitative ethnobotany or quantification in ethnobotany? Ethnobot. Res. Appl. 2009, 7, 1–3. [Google Scholar] [CrossRef]
- Hoffman, B.; Gallaher, T. Importance indices in ethnobotany. Ethnobot. Res. Appl. 2007, 5, 201–218. [Google Scholar] [CrossRef]
- Bennett, B.C. Ethnobotany education, opportunities, and needs in the US. Ethnobot. Res. Appl. 2005, 3, 113–122. [Google Scholar] [CrossRef][Green Version]
- Martin, G.J. Ethnobotany: A Methods Manual; Earthscan: London, UK, 2007. [Google Scholar]
- Albuquerque, U.P.; Hanazaki, N. Commentary: Five problems in current ethnobotanical research and some suggestions for strengthening them. Hum. Ecol. 2009, 37, 653–661. [Google Scholar] [CrossRef]
- Weiner, J. On the practice of ecology. J. Ecol. 1995, 83, 153–158. [Google Scholar] [CrossRef]
- Aarssen, L.W. On the progress of ecology. Oikos 1997, 80, 177–178. [Google Scholar] [CrossRef]
- Lawton, J.H. Are there general laws in ecology? Oikos 1999, 84, 177–192. [Google Scholar] [CrossRef]
- Marquet, P.A.; Allen, A.P.; Brown, J.H.; Dunne, J.A.; Enquist, B.J.; Gillooly, J.F.; Gowaty, P.A.; Green, J.L.; Harte, J.; Hubbell, S.P.; et al. On Theory in Ecology. BioScience 2014, 64, 701–710. [Google Scholar] [CrossRef]
- Bond, M.O.; Gaoue, O.G. Prestige and homophily predict network structure for social learning of medicinal plant knowledge. PLoS ONE 2020, 15, e0239345. [Google Scholar] [CrossRef]
- Coe, M.A.; Gaoue, O.G. Most cultural importance indices do not predict species’ cultural keystone status. Hum. Ecol. 2020, 48, 721–732. [Google Scholar] [CrossRef]
- Muleba, I.; Yessoufou, K.; Rampedi, I.T. Testing the non-random hypothesis of medicinal plant selection using the woody flora of the Mpumalanga Province, South Africa. Environ. Dev. Sustain. 2021, 23, 4162–4173. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Responses | Predictors | Estimate | Std. Error | DF | Crit. Value | p-Value |
|---|---|---|---|---|---|---|
| Availability | Residence time | 0.0009 | 0.0010 | 108 | 0.8538 | 0.3932 |
| Number of medicinal recipes | Availability | 0.0018 | 0.1002 | 101 | 0.0181 | 0.9856 |
| Number of medicinal recipes | Residence time | −0.0012 | 0.0024 | 101 | −0.4986 | 0.6181 |
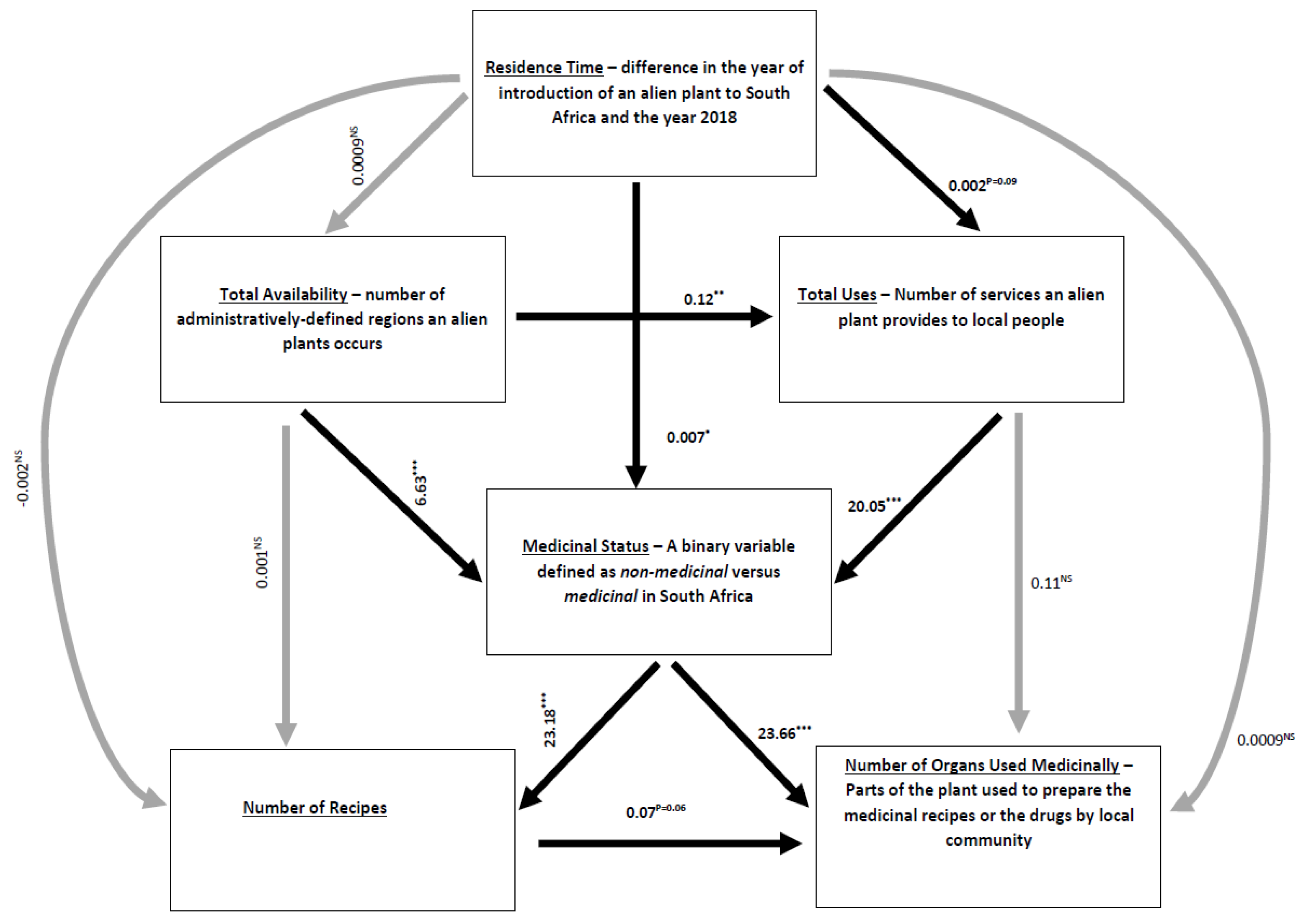
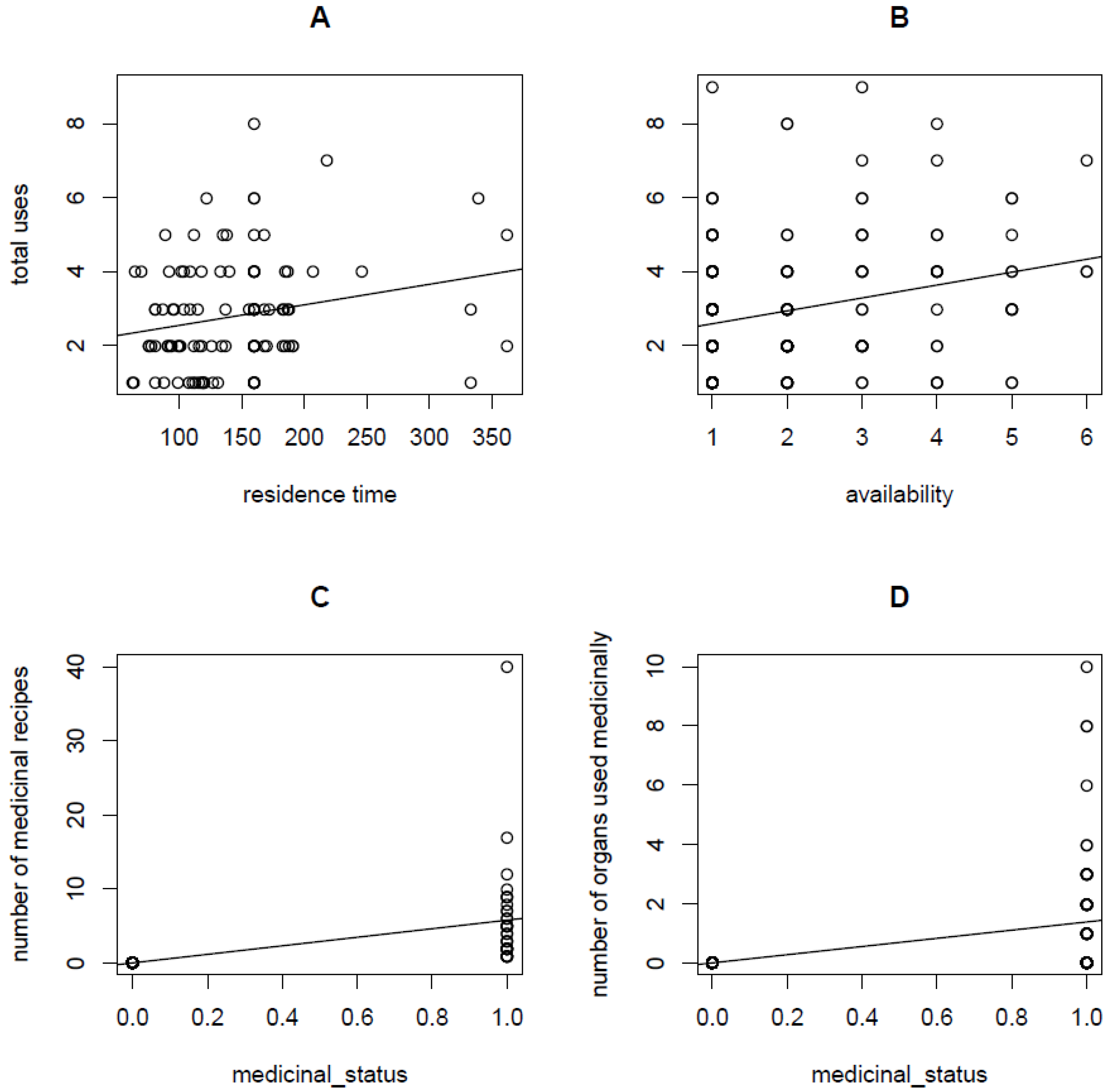
| Number of medicinal recipes | Medicinal status | 23.1849 | 2.0860 | 101 | 11.1144 | 0.0000 *** |
| Medicinal status | Residence time | 0.0068 | 0.0028 | 108 | 2.4456 | 0.0145 * |
| Medicinal status | Availability | 6.6288 | 0.0020 | 108 | 3383.6146 | 0.0000 *** |
| Medicinal status | Total uses | 20.0485 | 0.0014 | 108 | 14,009.9666 | 0.0000 *** |
| Total uses | Residence time | 0.0016 | 0.0009 | 108 | 1.6691 | 0.0951 ms |
| Total uses | Availability | 0.1155 | 0.0392 | 108 | 2.9500 | 0.0032 ** |
| Number of organs used medicinally | Number of medicinal recipes | 0.0734 | 0.0404 | 101 | 1.8162 | 0.0693 ms |
| Number of organs used medicinally | Medicinal status | 23.6634 | 2.9541 | 101 | 8.0103 | 0.0000 *** |
| Number of organs used medicinally | Residence time | 0.0009 | 0.0024 | 101 | 0.3566 | 0.7214 |
| Number of organs used medicinally | Total uses | 0.1134 | 0.1234 | 101 | 0.9187 | 0.3582 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yessoufou, K.; Ambani, A.E.; Elansary, H.O.; El-Sabrout, A.M.; Shokralla, S. Time, Mediated through Plant Versatility, Is a Better Predictor of Medicinal Status of Alien Plants. Diversity 2022, 14, 286. https://doi.org/10.3390/d14040286
Yessoufou K, Ambani AE, Elansary HO, El-Sabrout AM, Shokralla S. Time, Mediated through Plant Versatility, Is a Better Predictor of Medicinal Status of Alien Plants. Diversity. 2022; 14(4):286. https://doi.org/10.3390/d14040286
Chicago/Turabian StyleYessoufou, Kowiyou, Annie Estelle Ambani, Hosam O. Elansary, Ahmed M. El-Sabrout, and Shadi Shokralla. 2022. "Time, Mediated through Plant Versatility, Is a Better Predictor of Medicinal Status of Alien Plants" Diversity 14, no. 4: 286. https://doi.org/10.3390/d14040286
APA StyleYessoufou, K., Ambani, A. E., Elansary, H. O., El-Sabrout, A. M., & Shokralla, S. (2022). Time, Mediated through Plant Versatility, Is a Better Predictor of Medicinal Status of Alien Plants. Diversity, 14(4), 286. https://doi.org/10.3390/d14040286