
Mechanisms Generating Dichotomies in the Life Strategies of Heterotrophic Marine Prokaryotes


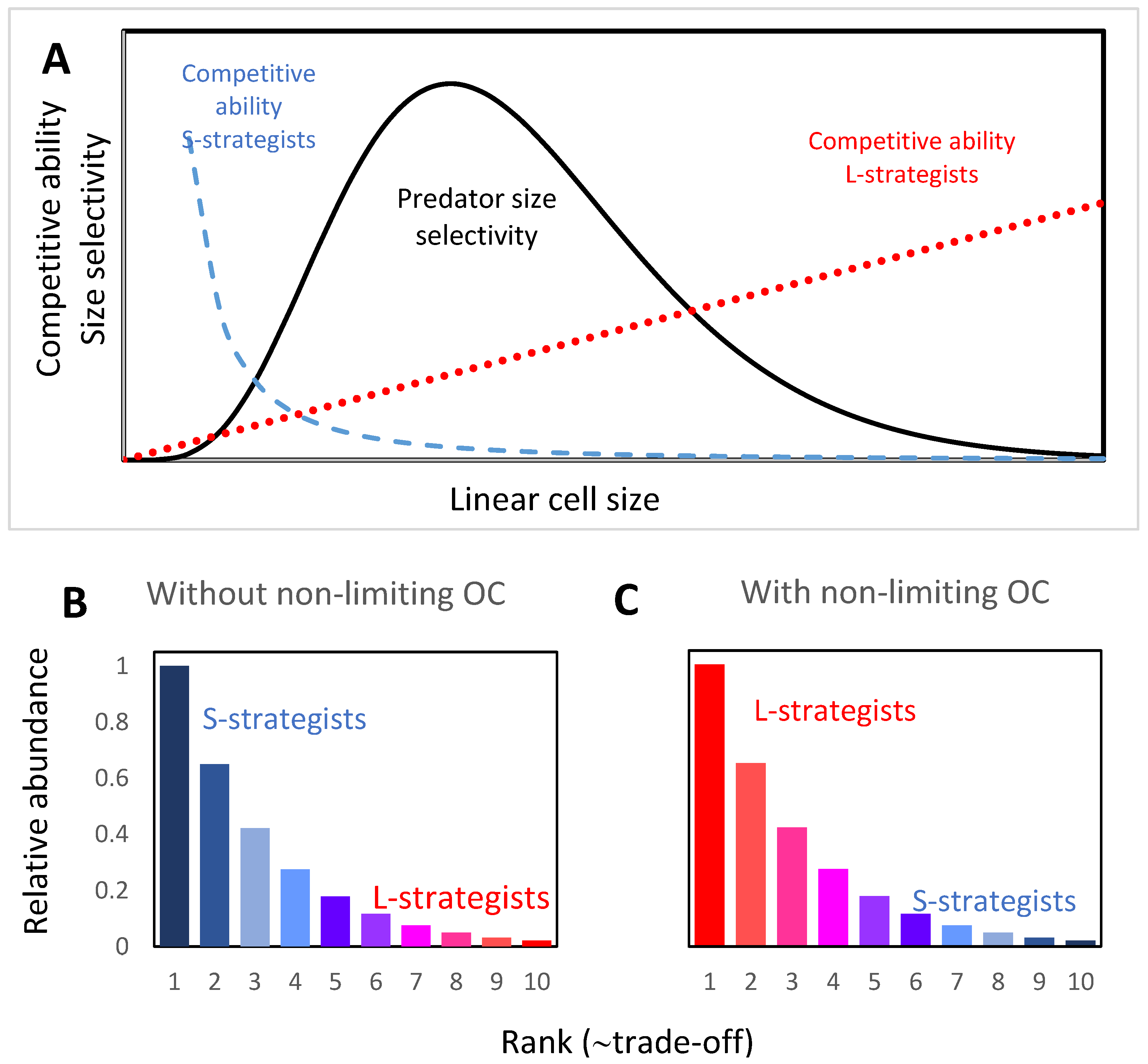{kind=link}
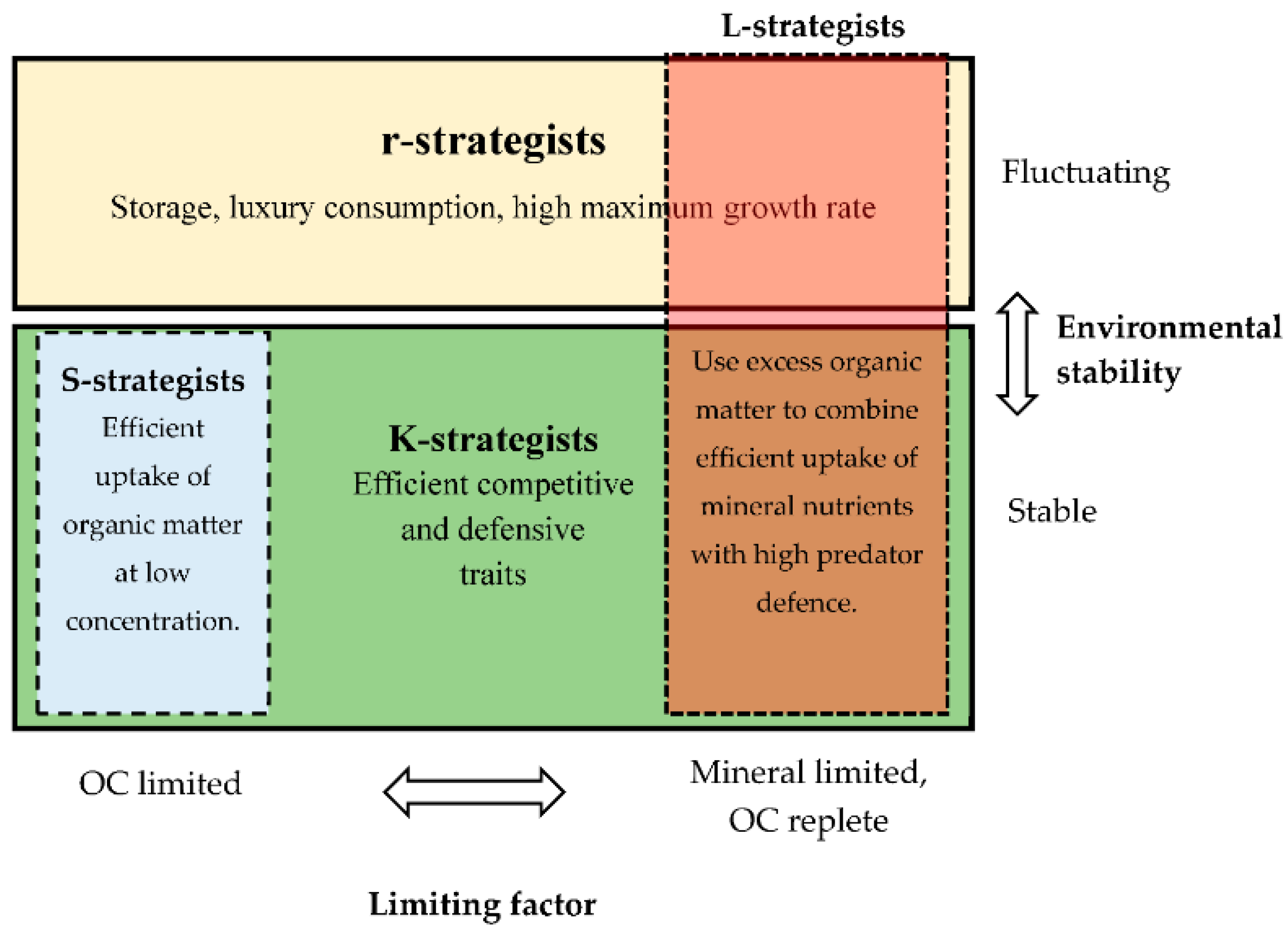{kind=link}
Abstract
:1. Introduction
2. Connections between Non-Limiting Organic C, Predation Resistance, and Pathogenicity
3. Observational Studies of Marine Vibrios
4. Bacterial Community Shifts in Mesocosms Amended with Glucose
5. Viability of Marine Larvae
6. What Drives Natural Environments toward Enrichment in Organic C?
7. Are the Flow-Cytometer Groups of LNA and HNA Populations Reflecting S- and L-Strategists, Respectively?
8. Relating S- vs. L- to r- vs. K-Selection
9. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Structure and function of the global ocean microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguíluz, V.M.; Salazar, G.; Fernández-Gracia, J.; Pearman, J.K.; Gasol, J.M.; Acinas, S.G.; Sunagawa, S.; Irigoien, X.; Duarte, C.M. Scaling of species distribution explains the vast potential marine prokaryote diversity. Sci. Rep. 2019, 9, 18710. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, S.J. SAR11 Bacteria: The Most Abundant Plankton in the Oceans. Annu. Rev. Mar. Sci. 2017, 9, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Yooseph, S.; Nealson, K.H.; Rusch, D.B.; McCrow, J.P.; Dupont, C.L.; Kim, M.; Johnson, J.; Montgomery, R.; Ferriera, S.; Beeson, K.; et al. Genomic and functional adaptation in surface ocean planktonic prokaryotes. Nature 2010, 468, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Tagomori, K.; Iida, T.; Honda, T. Comparison of genome structures of vibrios, bacteria possessing two chromosomes. J. Bacteriol. 2002, 184, 4351–4358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Valera, F.; Martin-Cuadrado, A.-B.; Rodriguez-Brito, B.; Pasic, L.; Thingstad, T.F.; Rohwer, F.; Mira, A. OPINION Explaining microbial population genomics through phage predation. Nat. Rev. Microbiol. 2009, 7, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Thingstad, T.F.; Våge, S.; Storesund, J.E.; Sandaa, R.-A.; Giske, J. A theoretical analysis of how strain-specific viruses can control microbial species diversity. Proc. Natl. Acad. Sci. USA 2014, 111, 7813–7818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannoni, S. Streamlining theory in microbial evolution. FEBS J. 2014, 281, 44. [Google Scholar]
- Morris, R.M.; Rappe, M.S.; Connon, S.A.; Vergin, K.L.; Siebold, W.A.; Carlson, C.A.; Giovannoni, S.J. SAR11 clade dominates ocean surface bacterioplankton communities. Nature 2002, 420, 806–810. [Google Scholar] [CrossRef]
- Giovannoni, S.J.; Tripp, H.J.; Givan, S.; Podar, M.; Vergin, K.L.; Baptista, D.; Bibbs, L.; Eads, J.; Richardson, T.H.; Noordewier, M.; et al. Genome Streamlining in a Cosmopolitan Oceanic Bacterium. Science 2005, 309, 1242–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.L.; Temperton, B.; Thrash, J.C.; Schwalbach, M.S.; Vergin, K.L.; Landry, Z.C.; Ellisman, M.; Deerinck, T.; Sullivan, M.B.; Giovannoni, S.J. Abundant SAR11 viruses in the ocean. Nature 2013, 494, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pedrós-Alió, C. The rare bacterial biosphere. Ann. Rev. Mar. Sci. 2012, 4, 449–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, J.H.; Harris, R.F. R-Selection and K-Selection and Microbial Ecology. Adv. Microb. Ecol. 1986, 9, 99–147. [Google Scholar]
- Vezzulli, L.; Pezzati, E.; Stauder, M.; Stagnaro, L.; Venier, P.; Pruzzo, C. Aquatic Ecology of the Oyster Pathogens Vibrio splendidus and Vibrio aestuarianus. Environ. Microbiol. 2014, 17, 1065–1080. [Google Scholar] [CrossRef] [PubMed]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio anguillarum as a fish pathogen: Virulence factors, diagnosis and prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.R. Global climate and infectious disease: The cholera paradigm. Science 1996, 274, 2025–2031. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.K.; Oliver, J.D. Vibrio vulnificus: Disease and Pathogenesis. Infect. Immun. 2009, 77, 1723–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.-C.; Liu, C. Vibrio parahaemolyticus: A concern of seafood safety. Food Microbiol. 2007, 24, 549–558. [Google Scholar] [CrossRef]
- Bourassa, L.; Camilli, A. Glycogen contributes to the environmental persistence and transmission of Vibrio cholerae. Mol. Microbiol. 2009, 72, 124–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adiba, S.; Nizak, C.; van Baalen, M.; Denamur, E.; Depaulis, F. From grazing resistance to pathogenesis: The coincidental evolution of virulence factors. PLoS ONE 2010, 5, e11882. [Google Scholar] [CrossRef]
- Hilbi, H.; Weber, S.S.; Ragaz, C.; Nyfeler, Y.; Urwyler, S. Environmental predators as models for bacterial pathogenesis. Environ. Microbiol. 2007, 9, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Erken, M.; Lutz, C.; McDougald, D. The Rise of Pathogens: Predation as a Factor Driving the Evolution of Human Pathogens in the Environment. Microb. Ecol. 2013, 65, 860–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, R.; Kjelleberg, S.; McDougald, D.; Jurgens, K. Species-specific patterns in the vulnerability of carbon-starved bacteria to protist grazing. Aquat. Microb. Ecol. 2011, 64, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Matz, C.; Jürgens, K. Interaction of nutrient limitation and protozoan grazing determines the phenotypic structure of a bacterial community. Microb. Ecol. 2003, 45, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Jürgens, K.; Matz, C. Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Antonie Van Leeuwenhoek 2002, 81, 413–434. [Google Scholar] [CrossRef]
- Matz, C.; Boenigk, J.; Arndt, H.; Jürgens, K. Role of bacterial phenotypic traits in selective feeding of the heterotrophic nanoflagellate Spumella sp. Aquat. Microb. Ecol. 2002, 27, 137–148. [Google Scholar] [CrossRef]
- Jürgens, K.; Pernthaler, J.; Schalla, S.; Amann, R. Morphological and compositional changes in a planktonic bacterial community in response to enhanced protozoan grazing. Appl. Environ. Microbiol. 1999, 65, 1241–1250. [Google Scholar] [CrossRef] [Green Version]
- Andersson, A.; Ahlinder, J.; Mathisen, P.; Hagglund, M.; Backman, S.; Nilsson, E.; Sjodin, A.; Thelaus, J. Predators and nutrient availability favor protozoa-resisting bacteria in aquatic systems. Sci. Rep. 2018, 8, 8415. [Google Scholar] [CrossRef] [Green Version]
- Preiss, J.; Romeo, T. Physiology, biochemistry and genetics of bacterial glycogen synthesis. Adv. Microb. Physiol. 1989, 30, 183–238. [Google Scholar]
- Wilkinson, J.F. Carbon and Energy Storage in Bacteria. J. Gen. Microbiol. 1963, 32, 171–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, B.W.; Mauchline, W.S.; Dennis, P.J.; Keevil, C.W.; Wait, R. Poly-3-Hydroxybutyrate in Legionella pneumophila, an Energy Source for Survival in Low-Nutrient Environments. Appl. Environ. Microbiol. 1999, 65, 822–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thingstad, T.F.; Øvreås, L.; Egge, J.K.; Løvdal, T.; Heldal, M. Use of non-limiting substrates to increase size; a generic strategy to simultaneously optimize uptake and minimize predation in pelagic osmotrophs? Ecol. Lett. 2005, 8, 675–682. [Google Scholar] [CrossRef]
- Jumars, P.; Deming, J.; Hill, P.; Karp-Boss, L.; Dade, W. Physical constraints on marine osmotrophy in an optimal foraging context. Mar. Microb. Food Webs 1993, 7, 121–161. [Google Scholar]
- Shannon, S.P.; Chrzanowski, T.H.; Grover, J.P. Prey Food Quality Affects Flagellate Ingestion Rates. Microb. Ecol. 2007, 53, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Olsen, Y.; Andersen, T.; Gismervik, I.; Vadstein, O. Marine heterotrophic bacteria, protozoan and metazoan zooplankton may experience protein N or mineral P limitation in coastal waters. Mar. Ecol. Prog. Ser. 2011, 436, 81–100. [Google Scholar] [CrossRef]
- Lapota, D.; Galt, C.; Losee, J.R.; Huddell, H.D.; Orzech, J.K.; Nealson, K.H. Observations and measurements of planktonic bioluminescence in and around a milky sea. J. Exp. Mar. Biol. Ecol. 1988, 119, 55–81. [Google Scholar] [CrossRef]
- Larsen, J.L. Vibrio anguillarum: Prevalence in three carbohydrate loaded marine recipients and a control. Zentralblatt für Bakteriologie Mikrobiologie und Hygiene: I. Abt. Originale C: Allgemeine, angewandte und ökologische Mikrobiologie 1982, 3, 519–530. [Google Scholar] [CrossRef]
- Westrich, J.R.; Ebling, A.M.; Landing, W.M.; Joyner, J.L.; Kemp, K.M.; Griffin, D.W.; Lipp, E.K. Saharan dust nutrients promote Vibrio bloom formation in marine surface waters. Proc. Natl. Acad. Sci. USA 2016, 113, 5964–5969. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.F.; Kelly, R.L.; Kauffman, K.M.; Reid, A.K.; Lauderdale, J.M.; Follows, M.J.; John, S.G. Growth of marine Vibrio in oligotrophic environments is not stimulated by the addition of inorganic iron. Earth Planet. Sci. Lett. 2019, 516, 148–155. [Google Scholar] [CrossRef]
- Djaoudi, K.; Wambeke, F.; Barani, A.; Bhairy, N.; Chevaillier, S.; Desboeufs, K.; Nunige, S.; Labiadh, M.; Tureaux, T.; Lefèvre, D.; et al. Potential bioavailability of organic matter from atmospheric particles to marine heterotrophic bacteria. Biogeosciences 2020, 17, 6271–6285. [Google Scholar] [CrossRef]
- Herut, B.; Rahav, E.; Tsagaraki, T.M.; Giannakourou, A.; Tsiola, A.; Psarra, S.; Lagaria, A.; Papageorgiou, N.; Mihalopoulos, N.; Theodosi, C.N.; et al. The Potential Impact of Saharan Dust and Polluted Aerosols on Microbial Populations in the East Mediterranean Sea, an Overview of a Mesocosm Experimental Approach. Front. Mar. Sci. 2016, 3, 226. [Google Scholar] [CrossRef] [Green Version]
- Neogi, S.B.; Lara, R.; Alam, M.; Harder, J.; Yamasaki, S.; Colwell, R.R. Environmental and hydroclimatic factors influencing Vibrio populations in the estuarine zone of the Bengal delta. Environ. Monit. Assess. 2018, 190, 565. [Google Scholar] [CrossRef] [PubMed]
- Stachowitsch, M.; Fanuko, N.; Richter, M. Mucus Aggregates in the Adriatic Sea: An Overview of Stages and Occurrences. Mar. Ecol. 1990, 11, 327–350. [Google Scholar] [CrossRef]
- Huq, A.; Small, E.B.; West, P.A.; Huq, M.I.; Rahman, R.; Colwell, R.R. Ecological relationships between Vibrio cholerae and planktonic crustacean copepods. Appl. Environ. Microbiol. 1983, 45, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzulli, L.; Brettar, I.; Pezzati, E.; Reid, P.C.; Colwell, R.R.; Höfle, M.G.; Pruzzo, C. Long-term effects of ocean warming on the prokaryotic community: Evidence from the vibrios. ISME J. 2012, 6, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raszl, S.M.; Froelich, B.A.; Vieira, C.R.; Blackwood, A.D.; Noble, R.T. Vibrio parahaemolyticus and Vibrio vulnificus in South America: Water, seafood and human infections. J. Appl. Microbiol. 2016, 121, 1201–1222. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [Green Version]
- Baker-Austin, C.; Trinanes, J.A.; Taylor, N.G.H.; Hartnell, R.; Siitonen, A.; Martinez-Urtaza, J. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Chang. 2013, 3, 73–77. [Google Scholar] [CrossRef]
- Takemura, A.; Chien, D.; Polz, M. Associations and dynamics of Vibrionaceae in the environment, from the genus to the population level. Front. Microbiol. 2014, 5, 38. [Google Scholar] [CrossRef] [Green Version]
- Montánchez, I.; Ogayar, E.; Plágaro, A.H.; Esteve-Codina, A.; Gómez-Garrido, J.; Orruño, M.; Arana, I.; Kaberdin, V.R. Analysis of Vibrio harveyi adaptation in sea water microcosms at elevated temperature provides insights into the putative mechanisms of its persistence and spread in the time of global warming. Sci. Rep. 2019, 9, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joint, I.; Henriksen, P.; Fonnes, G.A.; Bourne, D.; Thingstad, T.F.; Riemann, B. Competition for inorganic nutrients between phytoplankton and bacterioplankton in nutrient manipulated mesocosms. Aquat. Microb. Ecol. 2002, 29, 145–159. [Google Scholar] [CrossRef]
- Øvreås, L.; Bourne, D.; Sandaa, R.A.; Casamayor, E.O.; Benlloch, S.; Goddard, V.; Smerdon, G.; Heldal, M.; Thingstad, T.F. Response of bacterial and viral communities to nutrient manipulations in seawater mesocosms. Aquat. Microb. Ecol. 2003, 31, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Tsagaraki, T.M.; Pree, B.; Leiknes, O.; Larsen, A.; Bratbak, G.; Øvreås, L.; Egge, J.K.; Spanek, R.; Paulsen, M.L.; Olsen, Y.; et al. Bacterial community composition responds to changes in copepod abundance and alters ecosystem function in an Arctic mesocosm study. ISME J. 2018, 12, 2694–2705. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Nogi, Y. The Family Psychromonadaceae. In The Prokaryotes: Gammaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 583–590. [Google Scholar] [CrossRef]
- Hoffmann, K.; Hassenruck, C.; Salman-Carvalho, V.; Holtappels, M.; Bienhold, C. Response of Bacterial Communities to Different Detritus Compositions in Arctic Deep-Sea Sediments. Front. Microbiol. 2017, 8, 266. [Google Scholar] [CrossRef] [Green Version]
- Larsen, A.; Egge, J.K.; Nejstgaard, J.C.; Di Capua, I.; Thyrhaug, R.; Bratbak, G.; Thingstad, T.F. Contrasting response to nutrient manipulation in Arctic mesocosms are reproduced by a minimum microbial food web model. Limnol. Oceanogr. 2015, 60, 360–374. [Google Scholar] [CrossRef] [Green Version]
- Vadstein, O.; Øie, G.; Olsen, Y.; Salvesen, I.; Skjermo, J.; Skjåk-Bræk, G. A strategy to obtain microbial control during larval development of marine fish. In Fish Farming Technology; Reinertsen, H., Dahle, L.A., Jørgensen, L., Tvinnereim, K., Eds.; A.A. Balkema Publishers: Amsterdam, The Netherlands, 1993; pp. 69–75. [Google Scholar]
- Vadstein, O.; Attramadal, K.J.K.; Bakke, I.; Forberg, T.; Olsen, Y.; Verdegem, M.; Giatsis, C.; Skjermo, J.; Aasen, I.M.; Gatesoupe, F.J.; et al. Managing the Microbial Community of Marine Fish Larvae: A Holistic Perspective for Larviculture. Front. Microbiol. 2018, 9, 1820. [Google Scholar] [CrossRef]
- Vadstein, O.; Attramadal, K.J.K.; Bakke, I.; Olsen, Y. K-Selection as Microbial Community Management Strategy: A Method for Improved Viability of Larvae in Aquaculture. Front. Microbiol. 2018, 9, 2730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Schryver, P.; Defoirdt, T.; Sorgeloos, P. Early Mortality Syndrome Outbreaks: A Microbial Management Issue in Shrimp Farming? PLoS Path. 2014, 10, e1003919. [Google Scholar] [CrossRef]
- Kumar, T.S.; Vidya, R.; Kumar, S.; Alavandi, S.V.; Vijayan, K.K. Zoea-2 syndrome of Penaeus vannamei in shrimp hatcheries. Aquaculture 2017, 479, 759–767. [Google Scholar] [CrossRef]
- Matz, C.; Webb, J.S.; Schupp, P.J.; Phang, S.Y.; Penesyan, A.; Egan, S.; Steinberg, P.; Kjelleberg, S. Marine Biofilm Bacteria Evade Eukaryotic Predation by Targeted Chemical Defense. PLoS ONE 2008, 3, e2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olafsen, J. Interaction between fish larvae and bacteria in marine aquaculture. Aquaculture 2001, 200, 223–247. [Google Scholar] [CrossRef]
- Bergh, O. The dual myths of the healthy wild fish and the unhealthy farmed fish. Dis. Aquat. Org. 2007, 75, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Moland, E.; Synnes, A.-E.; Naustvoll, L.-J.; Brandt, C.; Norderhaug, K.; Thormar, J.; Biuw, M.; Jorde, P.; Knutsen, H.; Dahle, G.; et al. Krafttak for Kysttorsken_ Kunnskap for Stedstilpasset Gjenoppbygging av Bestander, Naturtyper og økosystem i Færder- og Ytre Hvaler Nasjonalparker; Institute of Marine Research: Bergen, Norway, 2021. (In Norwegian) [Google Scholar]
- Norwegian Institute of Public Health. Bakterier i Sjøvann kan gi Infeksjoner. Available online: https://www.fhi.no/ml/badevann/bakterier-i-sjovann-kan-gi-infeksjoner/ (accessed on 9 March 2022).
- Olsen, L.M.; Reinertsen, H.; Vadstein, O. Can phosphorus limitation inhibit dissolved organic carbon consumption in aquatic microbial food webs? A study of three food web structures in microcosms. Microb. Ecol. 2002, 43, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.; Ducklow, H. Growth of bacterioplankton and consumption of dissolved organic carbon in the Sargasso Sea. Aquat. Microb. Ecol. 1996, 10, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Thingstad, T.F.; Hagstrom, A.; Rassoulzadegan, F. Accumulation of degradable DOC in surface waters: Is it caused by a malfunctioning microbial loop? Limnol. Oceanogr. 1997, 42, 398–404. [Google Scholar] [CrossRef]
- Pengerud, B.; Skjoldal, E.F.; Thingstad, T.F. The reciprocal interaction between degradation of glucose and ecosystem—Studies in mixed chemostat cultures of marine-bacteria, algae, and bacterivorous nanoflagellates. Mar. Ecol. Prog. Ser. 1987, 35, 111–117. [Google Scholar] [CrossRef]
- Vadstein, O.; Jensen, A.; Olsen, Y.; Reinertsen, H. Growth and Phosphorus Status of Limnetic Phytoplankton and Bacteria. Limnol. Oceanogr. 1988, 33, 489–503. [Google Scholar] [CrossRef]
- Vadstein, O. Heterotrophic, planktonic bacteria and cycling of phosphorus—Phosphorus requirements, competitive ability, and food web interactions. Adv. Microb. Ecol. 2000, 16, 115–167. [Google Scholar]
- Thingstad, T.F.; Skjoldal, E.F.; Bohne, R.A. Phosphorus cycling and algal-bacterial competition in Sandsfjord, western Norway. Mar. Ecol. Prog. Ser. 1993, 99, 239–259. [Google Scholar] [CrossRef]
- Rivkin, R.B.; Anderson, M.R. Inorganic nutrient limitation of oceanic bacterioplankton. Limnol. Oceanogr. 1997, 42, 730–740. [Google Scholar] [CrossRef] [Green Version]
- Zohary, T.; Robarts, R.D. Experimental study of microbial P limitation in the eastern Mediterranean. Limnol. Oceanogr. 1998, 43, 387–395. [Google Scholar] [CrossRef]
- Lancelot, C.; Billen, G.; Sournia, A.; Weisse, T.; Coljin, F.; Veldhuis, M.; Davies, A.; Wassman, P. Phaeocystis blooms and nutrient enrichment in the continental coastal zones of the North Sea. Ambio 1987, 16, 38–46. [Google Scholar]
- Thingstad, T.F.; Bellerby, R.G.J.; Bratbak, G.; Borsheim, K.Y.; Egge, J.K.; Heldal, M.; Larsen, A.; Neill, C.; Nejstgaard, J.; Norland, S.; et al. Counterintuitive carbon-to-nutrient coupling in an Arctic pelagic ecosystem. Nature 2008, 455, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Thingstad, T.F.; Havskum, H.; Zweifel, U.L.; Berdalet, E.; Sala, M.M.; Peters, F.; Alcaraz, M.; Scharek, R.; Perez, M.; Jacquet, S.; et al. Ability of a “minimum” microbial food web model to reproduce response patterns observed in mesocosms manipulated with N and P, glucose, and Si. J. Mar. Syst. 2007, 64, 15–34. [Google Scholar] [CrossRef] [Green Version]
- Avcı, B.; Krüger, K.; Fuchs, B.M.; Teeling, H.; Amann, R.I. Polysaccharide niche partitioning of distinct Polaribacter clades during North Sea spring algal blooms. ISME J. 2020, 14, 1369–1383. [Google Scholar] [CrossRef]
- Delpech, L.M.; Vonnahme, T.R.; McGovern, M.; Gradinger, R.; Praebel, K.; Poste, A.E. Terrestrial Inputs Shape Coastal Bacterial and Archaeal Communities in a High Arctic Fjord (Isfjorden, Svalbard). Front. Microbiol. 2021, 12, 614634. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, M.L.; Nielsen, S.E.B.; Müller, O.; Møller, E.F.; Stedmon, C.A.; Juul-Pedersen, T.; Markager, S.; Sejr, M.K.; Delgado Huertas, A.; Larsen, A.; et al. Carbon Bioavailability in a High Arctic Fjord Influenced by Glacial Meltwater, NE Greenland. Front. Mar. Sci. 2017, 4, 176. [Google Scholar] [CrossRef] [Green Version]
- Schattenhofer, M.; Wulf, J.; Kostadinov, I.; Glöckner, F.O.; Zubkov, M.V.; Fuchs, B.M. Phylogenetic characterisation of picoplanktonic populations with high and low nucleic acid content in the North Atlantic Ocean. Syst. Appl. Microbiol. 2011, 34, 470–475. [Google Scholar] [CrossRef]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [Green Version]
- Mojica, K.D.A.; Carlson, C.A.; Behrenfeld, M.J. Regulation of Low and High Nucleic Acid Fluorescent Heterotrophic Prokaryote Subpopulations and Links to Viral-Induced Mortality Within Natural Prokaryote-Virus Communities. Microb. Ecol. 2020, 79, 213–230. [Google Scholar] [CrossRef]
- Van Wambeke, F.; Catala, P.; Pujo-Pay, M.; Lebaron, P. Vertical and longitudinal gradients in HNA-LNA cell abundances and cytometric characteristics in the Mediterranean Sea. Biogeosciences 2011, 8, 1853–1863. [Google Scholar] [CrossRef] [Green Version]
- Eagon, R.G. Pseudomonas natriegens, a marine bacterium with a generation time of less than 10 minutes. J. Bacteriol. 1962, 83, 736–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, Y.Y.; Lee, C.W.; Bong, C.W.; Lim, J.H.; Narayanan, K.; Sim, E.U.H. Environmental control of Vibrio spp. abundance and community structure in tropical waters. FEMS Microbiol. Ecol. 2019, 95, fiz176. [Google Scholar] [CrossRef]
- Lauro, F.M.; McDougald, D.; Thomas, T.; Williams, T.J.; Egan, S.; Rice, S.; DeMaere, M.Z.; Ting, L.; Ertan, H.; Johnson, J.; et al. The genomic basis of trophic strategy in marine bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 15527–15533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westoby, M.; Nielsen, D.A.; Gillings, M.R.; Litchman, E.; Madin, J.S.; Paulsen, I.T.; Tetu, S.G. Cell size, genome size, and maximum growth rate are near-independent dimensions of ecological variation across bacteria and archaea. Ecol. Evol. 2021, 11, 3956–3976. [Google Scholar] [CrossRef]
- Lopez-Perez, M.; Haro-Moreno, J.M.; Coutinho, F.H.; Martinez-Garcia, M.; Rodriguez-Valera, F. The Evolutionary Success of the Marine Bacterium SAR11 Analyzed through a Metagenomic Perspective. Msystems 2020, 5, e00605-20. [Google Scholar] [CrossRef]
- Polovina, J.J.; Howell, E.A.; Abecassis, M. Ocean’s least productive waters are expanding. Geophys. Res. Lett. 2008, 35, L031745. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thingstad, T.F.; Øvreås, L.; Vadstein, O. Mechanisms Generating Dichotomies in the Life Strategies of Heterotrophic Marine Prokaryotes. Diversity 2022, 14, 217. https://doi.org/10.3390/d14030217
Thingstad TF, Øvreås L, Vadstein O. Mechanisms Generating Dichotomies in the Life Strategies of Heterotrophic Marine Prokaryotes. Diversity. 2022; 14(3):217. https://doi.org/10.3390/d14030217
Chicago/Turabian StyleThingstad, Tron Frede, Lise Øvreås, and Olav Vadstein. 2022. "Mechanisms Generating Dichotomies in the Life Strategies of Heterotrophic Marine Prokaryotes" Diversity 14, no. 3: 217. https://doi.org/10.3390/d14030217
APA StyleThingstad, T. F., Øvreås, L., & Vadstein, O. (2022). Mechanisms Generating Dichotomies in the Life Strategies of Heterotrophic Marine Prokaryotes. Diversity, 14(3), 217. https://doi.org/10.3390/d14030217