
First Molecular Phylogeny of Lumbrineridae (Annelida)

Abstract
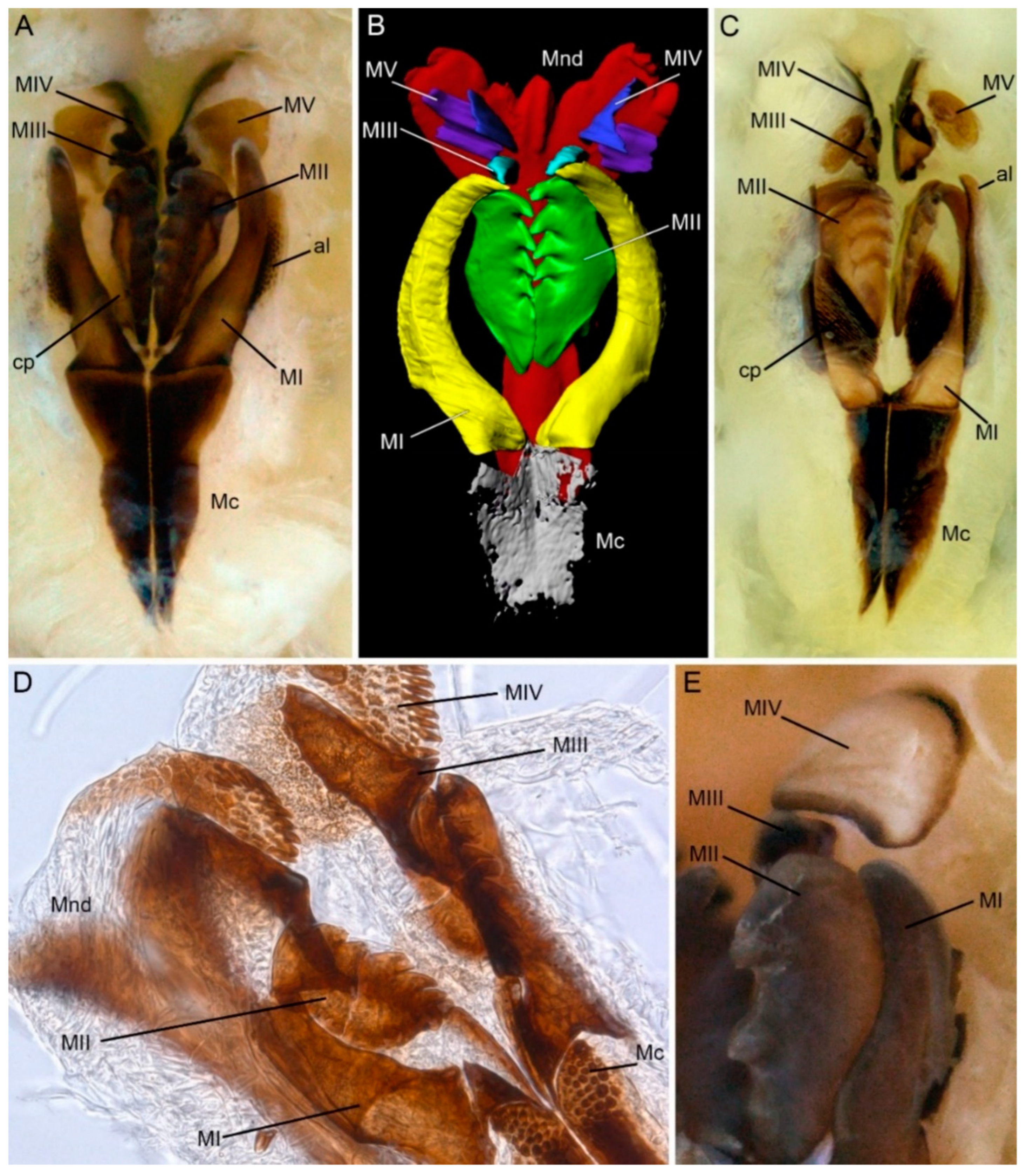
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction, PCR Amplification and DNA Sequencing
2.3. Sequence Alignment
2.4. Phylogenetic Analyses
2.4.1. Bayesian Inference (BI)
2.4.2. Maximum Likelihood (ML)
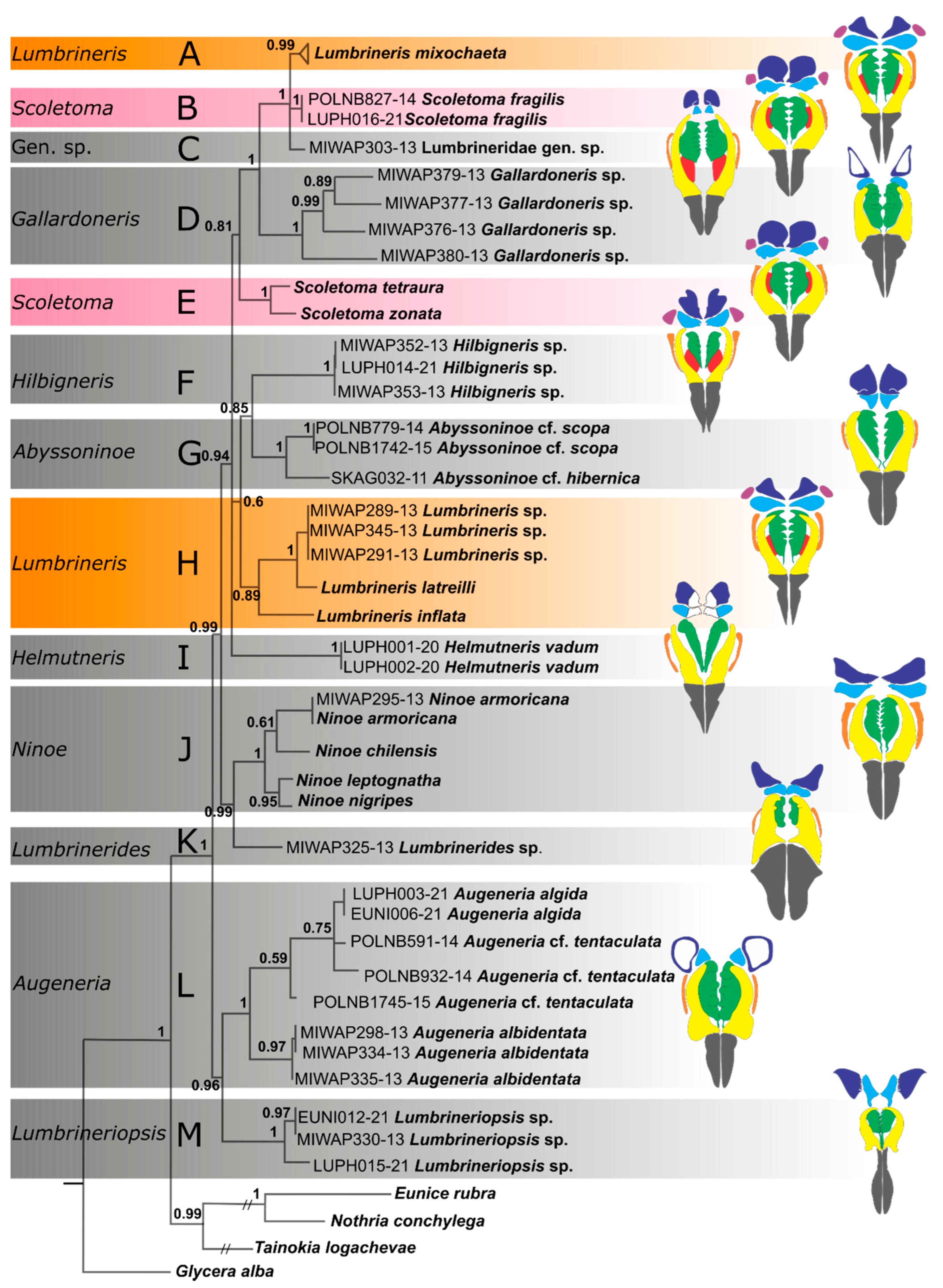
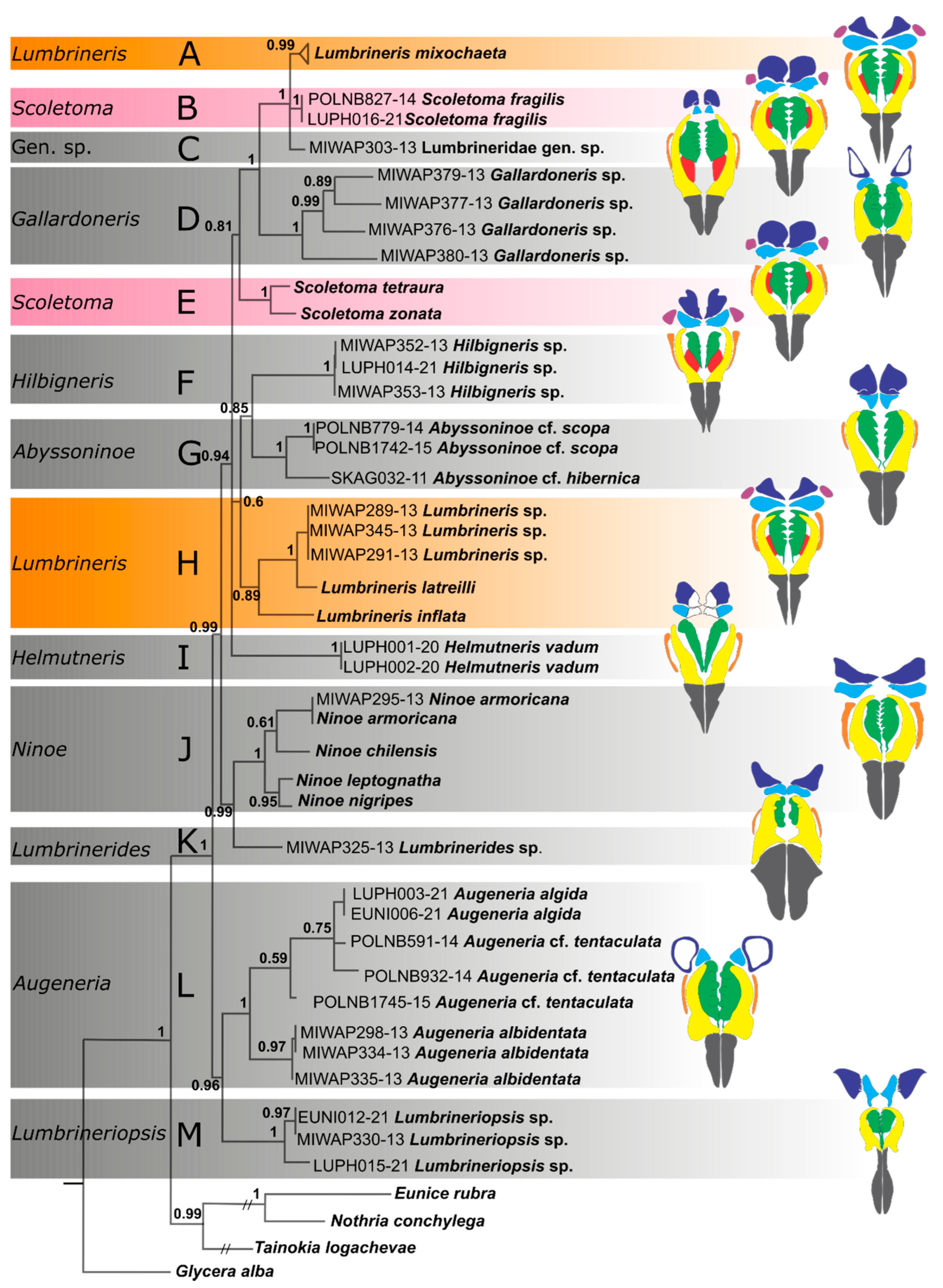
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmarda, L.K. Neue wirbellose thiere beobachtet und gesammelt auf einer reise um die erde 1853 bis 1857. In Turbellarien, Rotatorien und Anneliden; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1861. [Google Scholar]
- Orensanz, J.M. The eunicemorph polychaete annelids from antarctic and subantarctic seas: With addenda to the Eunicemorpha of Argentina, Chile, New Zealand, Australia, and the Southern Indian Ocean. In Biology of the Antarctic Seas XXI; American Geophysical Union (AGU): New Orleans, LA, USA, 1990; pp. 1–183. ISBN 978-1-118-66779-8. [Google Scholar]
- Carrera-Parra, L.F. Revision of Lumbrineris de Blainville, 1828 (Polychaeta: Lumbrineridae) Sars. Zootaxa 2006, 1336, 1–64. [Google Scholar] [CrossRef]
- Paxton, H. Phylogeny of Eunicida (Annelida) Based on Morphology of Jaws. Zoosymposia 2009, 2, 241–264. [Google Scholar] [CrossRef] [Green Version]
- Zanol, J.; Carrera-Parra, L.F.; Steiner, T.M.; Amaral, A.C.Z.; Wiklund, H.; Ravara, A.; Budaeva, N. The Current State of Eunicida (Annelida) Systematics and Biodiversity. Diversity 2021, 13, 74. [Google Scholar] [CrossRef]
- Oug, E.; Budaeva, N.; Borisova, P. Lumbrineridae Schmarda, 1861. In Handbook of Zoology Annelida Volume 4: Pleistoannelida, Errantia II; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany, 2022; pp. 1–35. [Google Scholar]
- Carrera-Parra, L.F. Phylogenetic Analysis of Lumbrineridae Schmarda, 1861 (Annelida: Polychaeta). Zootaxa 2006, 1332, 1–36. [Google Scholar] [CrossRef]
- McIntosh, W.C. Report on the Annelida Polychaeta Collected by HMS ”Challenger” during the Years 1873-76. Reports on the Scientific Results of the Voyage of HMS ”Challenger”. Zoology 1885, 12, 1–554. [Google Scholar]
- Hartmann-Schröder, G. Die polychaeten der tropischen nordwestküste Australiens (zwischen derby im norden und port hedland im süden). teil 2. In Zur Kenntnis Des Eulitorals Der Australischen Küsten Unter Besonder Berücksichtigung Der Polychaeten Und Ostracoden. (Teil 2 Und Teil 3); Hartmann-Schröder, G., Hartmann, G., Eds.; Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut: Hamburg, Germany, 1979; Volume 76, pp. 77–218. [Google Scholar]
- Monro, C.C.A. Polychaete Worms. Discov. Rep. Camb. 1930, 2, 1–222. [Google Scholar]
- Chamberlin, R.V. The Annelida Polychaeta; Museum of Comparative Zoölogy at Harvard College: Cambridge, MA, USA, 1919; Volume 48. [Google Scholar]
- Kinberg, J.G.H. Annulata Nova, Eunicea; Fversi gt af Kongl Vetenskaps-Akademiens Forhandlingar: Stockholm, Sweden, 1865; Volume 21, pp. 559–574. [Google Scholar]
- Gallardo, V.A. Polychaeta from the Bay of Nha Trang, South Viet Nam. Naga Rep. 1968, 4, 35–279. [Google Scholar]
- Fage, L. Sur l’association d’un Annélide Polychète, Lumbriconereis flabellicola n. sp. et d’un Madrépore Flabellum pavoninum distinctum e. et h. XII Congrs Int. De Zool. Lisb. 1936, 2, 941–945. [Google Scholar]
- Mohammad, M.-B.M. New Species and Records of Polychaete Annelids from Kuwait, Arabian Gulf. Zool. J. Linn. Soc. 1973, 52, 23–44. [Google Scholar] [CrossRef]
- Frame, A.B. The Lumbrinerids (Annelida: Polychaeta) Collected in Two Northwestern Atlantic Surveys with Descriptions of a New Genus and Two New Species. Proc. Biol. Soc. Wash. 1992, 105, 185–218. [Google Scholar]
- Grube, A.E. Untersuchungen Über Die Familie Eunicea. Jahresber. Schlesichen Ges. Fr Vaterlndische Kult. 1878, 55, 79–104. [Google Scholar]
- Orensanz, J.M. Los Anélidos Poliquetos de La Provincia Biogeográfica Argentina. IV. Lumbrineridae. [The Polychaetous Annelids of the Argentine Biogeographic Province. IV. Lumbrineridae]. Physis 1973, 32, 343–393. [Google Scholar]
- Ehlers, E. Die bodensässigen anneliden aus den sammlungen der Deutschen tiefsee-expedition. In Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer ”Valdivia” 1898–1899; Chun, C., Ed.; Verlag von Gustav Fischer: Stuttgart, Germany, 1908; pp. 1–168. [Google Scholar]
- De Blainville, H. Mollusques, vers et zoophytes. In Dictionnaire des Sciences Naturelles; Leuvrant, F.G., Ed.; Dictionnaire des Sciences Naturelles: Strasbourg, France, 1828; Volume 57, p. 628. [Google Scholar]
- Audouin, J.V.; Milne-Edwards, H. Recherches Pour Servir L’histoire Naturelle Du Littoral de La France, Ou Recueil de Mémoires Sur L’anatomie, La Physiologie, La Classification et Les Moeurs Des Animaux de Nos Côtes; Ouvrage Accompagné Des Planches Faites de Après Nature. 2. Annélides; Crochard Libraire: Paris, France, 1834. [Google Scholar]
- Müller, O.F. Zoologiae Danicae Prodromus: Seu Animalium Daniae et Norvegiae Indigenarum Characteres, Nomina, et Synonyma Imprimis Popularium; Typis Havniæ [Copenhagen], Hallageriis: Canberra, Austrilia, 1776. [Google Scholar]
- Hartman, O. Polychaetous Annelids, Est. 5, Eunicea. Allan Hancock Pacif. Exped 1944, 10, 1–238. [Google Scholar]
- Fauchald, K. Polychaetous Annelids of the Families Eunicidae, Lumbrineridae, Iphitimidae, Arabellidae, Lysaretidae and Dorvilleidae from Western Mexico; Allan Hancock Monographs in Marine Biology: Los Angeles, CA, USA, 1970; pp. 1–112. [Google Scholar]
- Fauchald, K. The Polychaete Worms. Definitions and Keys to the Orders, Families and Genera. Nat. Hist. Mus. Los Angeles Cty. Sci. Ser. 1977, 28, 1–198. [Google Scholar]
- Struck, T.H.; Purschke, G.; Halanych, K.M. Phylogeny of Eunicida (Annelida) and Exploring Data Congruence Using a Partition Addition Bootstrap Alteration (PABA) Approach. Syst. Biol. 2006, 55, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilic, E.; Bartolomaeus, T.; Rouse, G.W. Chaetal Type Diversity Increases during Evolution of Eunicida (Annelida). Org. Divers. Evol. 2016, 16, 105–119. [Google Scholar] [CrossRef]
- Struck, T.H. Direction of Evolution within Annelida and the Definition of Pleistoannelida. J. Zool. Syst. Evol. Res. 2011, 49, 340–345. [Google Scholar] [CrossRef]
- Struck, T.H.; Golombek, A.; Weigert, A.; Franke, F.A.; Westheide, W.; Purschke, G.; Bleidorn, C.; Halanych, K.M. The Evolution of Annelids Reveals Two Adaptive Routes to the Interstitial Realm. Curr. Biol. 2015, 25, 1993–1999. [Google Scholar] [CrossRef] [Green Version]
- Tilic, E.; Stiller, J.; Campos, E.; Pleijel, F.; Rouse, G.W. Phylogenomics Resolves Ambiguous Relationships within Aciculata (Errantia, Annelida). Mol. Phylogenetics Evol. 2022, 166, 107339. [Google Scholar] [CrossRef]
- Oug, E. A New Small Species of Lumbrineris from Northern Norway and Arctic Waters, with Comments on L. minuta (Théel, 1879) and L. vanhoeefeni (Michaelsen, 1898) (Polychaeta: Lumbrineridae). Ophelia 1998, 49, 147–162. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, Z.; Chen, H.; Sun, R.; Wang, H.; Guo, L.; Pan, H. Integrating a DNA Barcoding Project with an Ecological Survey: A Case Study on Temperate Intertidal Polychaete Communities in Qingdao, China. Chin. J. Oceanol. Limnol. 2010, 28, 899–910. [Google Scholar] [CrossRef]
- McIntosh, W.C. Notes from the Gatty Marine Laboratory, St. Andrews. No. 25. 1. On the Eunicidae Dredged by HMS ‘Porcupine’ in 1869 and 1870. 2. On Canadian Eunicidae Dredged by Dr. Whiteaves, of the Canadian Geological Survey, in 1871–1873. 3. On Norwegian Eunicidae Collected by Canon Norman DCL, FRS. Ann. Mag. Nat. Hist. 1903, 12, 128–165. [Google Scholar]
- Paul, C.; Halanych, K.M.; Tiedemann, R.; Bleidorn, C. Molecules Reject an Opheliid Affinity for Travisia (Annelida). Syst. Biodivers. 2010, 8, 507–512. [Google Scholar] [CrossRef]
- Johnson, H.P. The Polychaeta of the Puget Sound Region. Proc. Boston Soc. Nat. Hist. 1901, 29, 381–437. [Google Scholar]
- Fauchald, K. Deep-Water Errant Polychaetes from Hardangerfjorden, Western Norway. Sarsia 1974, 57, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.P. The Polychaetous Annelids Dredged by the U.S.S. “Albatross” off the Coast of Southern California in 1904. III. Euphrosynidae to Goniadidae. Proc. Acad. Nat. Sci. Phila. 1911, 63, 234–318. [Google Scholar]
- Struck, T.H.; Purschke, G. The Sister Group Relationship of Aeolosomatidae and Potamodrilidae (Annelida: “Polychaeta”)—A Molecular Phylogenetic Approach Based on 18S RDNA and Cytochrome Oxidase I. Zool. Anz. A J. Comp. Zool. 2005, 243, 281–293. [Google Scholar] [CrossRef]
- Borisova, P.; Budaeva, N. Helmutneris vadum, a New Species of Lumbrineridae (Annelida) from Lizard Island, Great Barrier Reef, Australia. Zootaxa 2020, 4877, 413–421. [Google Scholar] [CrossRef]
- Glémarec, M. Ninoe armoricana n. sp., Polychète Lumbrineridae de La «Grande Vasière»(Golfe de Gascogne); Vie Et Milieu: Banyuls-sur-Mer, France, 1968; pp. 315–322. [Google Scholar]
- Aylagas, E.; Borja, Á.; Irigoien, X.; Rodríguez-Ezpeleta, N. Benchmarking DNA Metabarcoding for Biodiversity-Based Monitoring and Assessment. Front. Mar. Sci. 2016, 3, 96. [Google Scholar] [CrossRef] [Green Version]
- Maturana, C.S.; Moreno, R.A.; Labra, F.A.; González-Wevar, C.A.; Rozbaczylo, N.; Carrasco, F.D.; Poulin, E. DNA Barcoding of Marine Polychaetes Species of Southern Patagonian Fjords. Rev. Biol. Mar. Oceanogr. 2011, 46, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, E. Magellanischen Anneliden Gesammelt Während Der Schwedischen Expedition Nach Den Magellansländern. Nachr. Königliche Ges. Wiss. Göttingen Math. Phys. Kl. 1900, 1900, 206–223. [Google Scholar]
- Verrill, A.E. XVIII. Report upon the Invertebrate Animals of Vineyard Sound and the Adjacent Waters, with an Account of the Physical Characters of the Region; Report on the Condition of the Sea Fisheries of the South Coast of New England; Government Printing Office: Washington, DC, USA, 1873. [Google Scholar]
- Lobo, J.; Costa, P.M.; Teixeira, M.A.L.; Ferreira, M.S.G.; Costa, M.H.; Costa, F.O. Enhanced Primers for Amplification of DNA Barcodes from a Broad Range of Marine Metazoans. BMC Ecol. 2013, 13, 34. [Google Scholar] [CrossRef] [Green Version]
- Rousset, V.; Pleijel, F.; Rouse, G.W.; Erséus, C.; Siddall, M.E. A Molecular Phylogeny of Annelids. Cladistics 2007, 23, 41–63. [Google Scholar] [CrossRef]
- Aylagas, E.; Borja, A.; Rodríguez-Ezpeleta, N. Environmental Status Assessment Using DNA Metabarcoding: Towards a Genetics Based Marine Biotic Index (GAMBI). PLoS ONE 2014, 9, e90529. [Google Scholar] [CrossRef] [Green Version]
- Zanol, J.; Halanych, K.M.; Struck, T.H.; Fauchald, K. Phylogeny of the Bristle Worm Family Eunicidae (Eunicida, Annelida) and the Phylogenetic Utility of Noncongruent 16S, COI and 18S in Combined Analyses. Mol. Phylogenet. Evol. 2010, 55, 660–676. [Google Scholar] [CrossRef]
- Sars, M. Beskrivelser og Iagttagelser Over Nogle Mærkelige Eller Nye i Havet Ved Den Bergenske Kyst Levende Dyr af Polypernes, Acalephernes, Radiaternes, Annelidernes, og Molluskernes Classer: Med en Kort Oversigt Over de Hidtil af Forfatteren Sammesteds Fundne Arter og Deres Forekommen; Thorstein Hallagers Forlag hos Chr. Dahl, R.S.: Bergen, Norway, 1835. [Google Scholar]
- Carr, C.M.; Hardy, S.M.; Brown, T.M.; Macdonald, T.A.; Hebert, P.D.N. A Tri-Oceanic Perspective: DNA Barcoding Reveals Geographic Structure and Cryptic Diversity in Canadian Polychaetes. PLoS ONE 2011, 6, e22232. [Google Scholar] [CrossRef] [Green Version]
- Budaeva, N.; Schepetov, D.; Zanol, J.; Neretina, T.; Willassen, E. When Molecules Support Morphology: Phylogenetic Reconstruction of the Family Onuphidae (Eunicida, Annelida) Based on 16S RDNA and 18S RDNA. Mol. Phylogenet. Evol. 2016, 94, 791–801. [Google Scholar] [CrossRef]
- Ravara, A.; Cunha, M.R. An Unusual New Species of Oenonidae (Polychaeta, Eunicida) from the North Atlantic Ocean. Mar. Biodivers. 2017, 48, 1037–1043. [Google Scholar] [CrossRef]
- Palumbi, S. Simple Fool’s Guide to PCR; Department of Zoology and Kewalo Marine Laboratory, University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Martins, R.; Carrera-Parra, L.F.; Quintino, V.; Rodrigues, A.M. Lumbrineridae (Polychaeta) from the Portuguese Continental Shelf (NE Atlantic) with the Description of Four New Species. Zootaxa 2012, 3416, 1–21. [Google Scholar] [CrossRef] [Green Version]
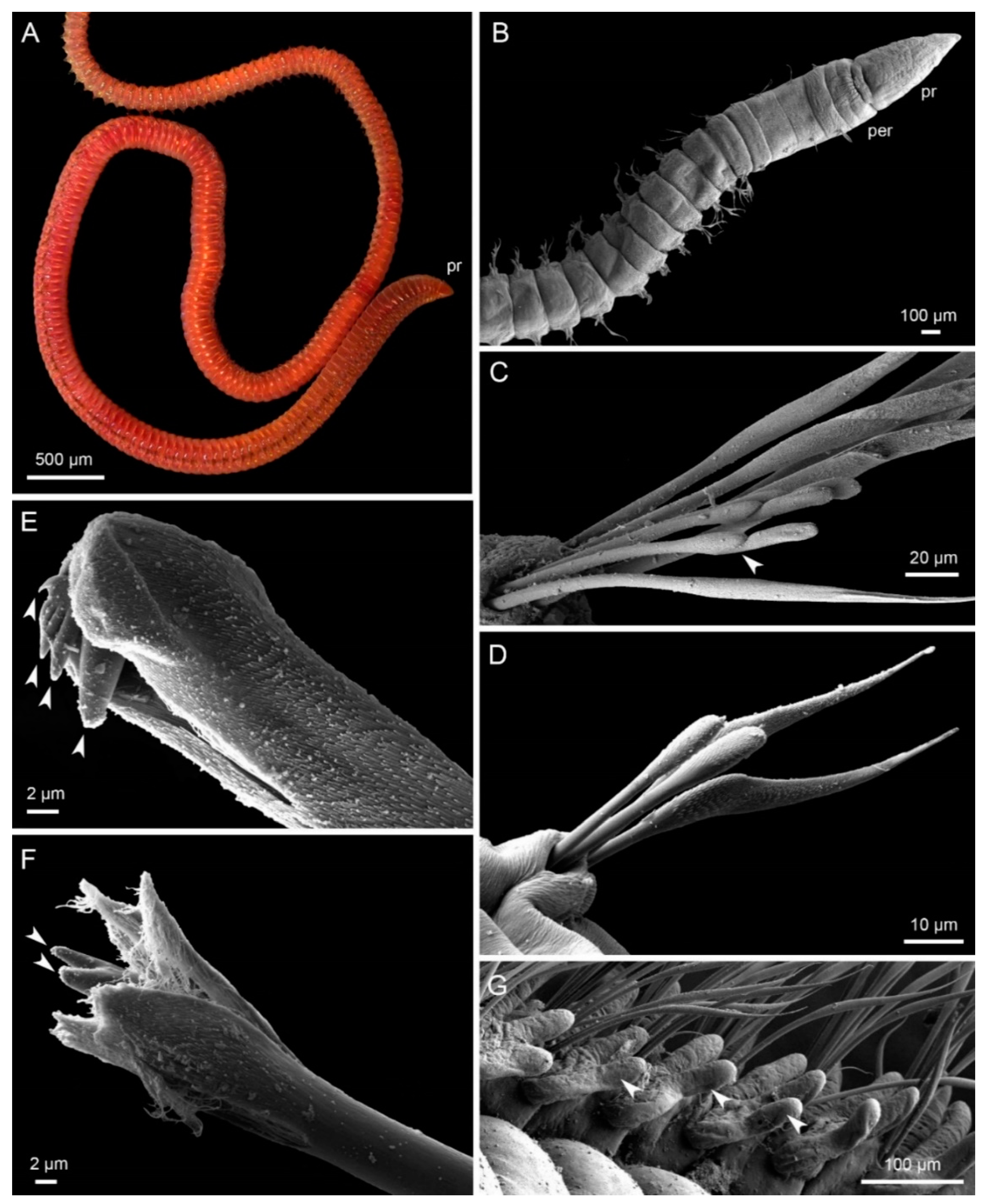
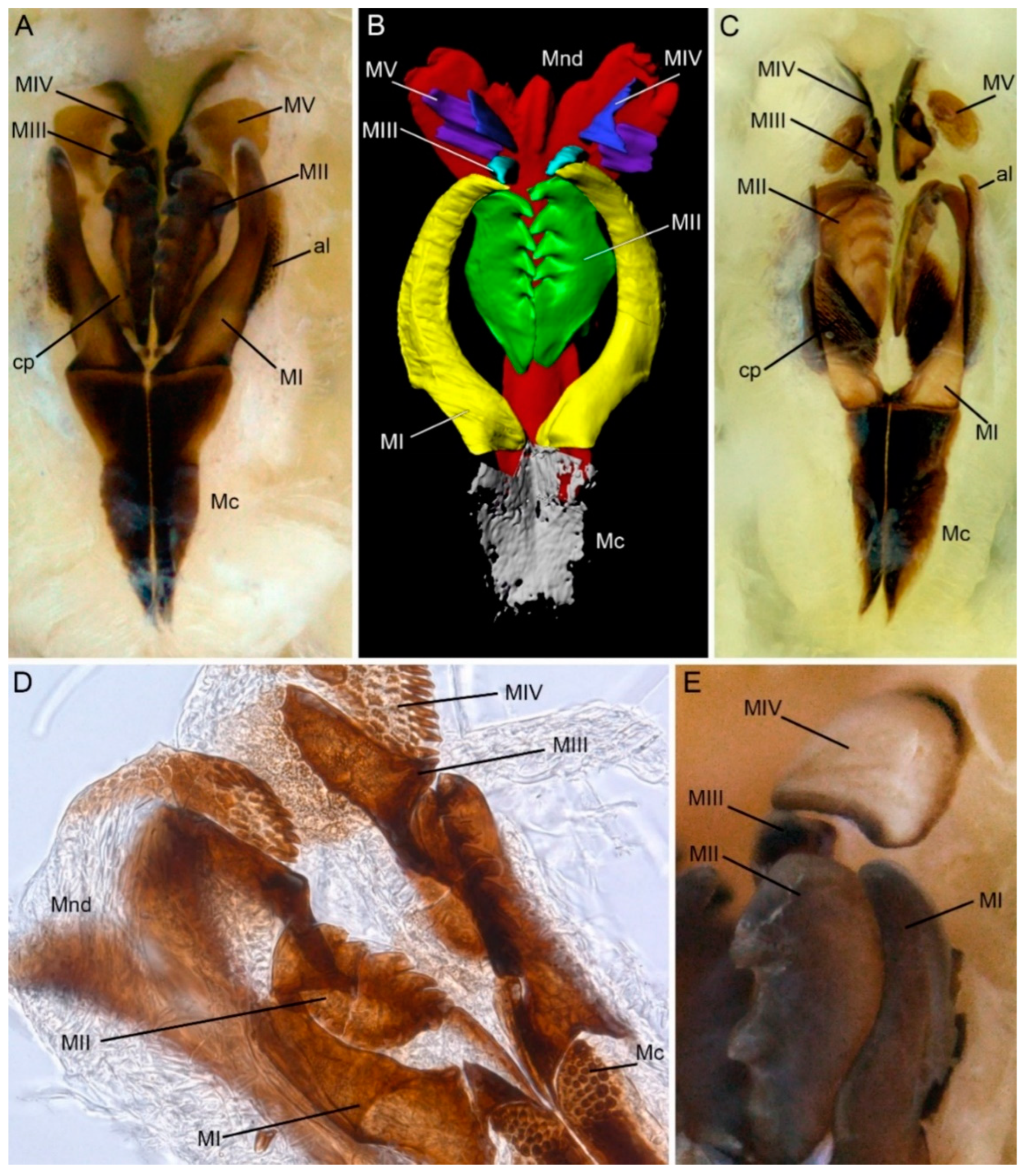

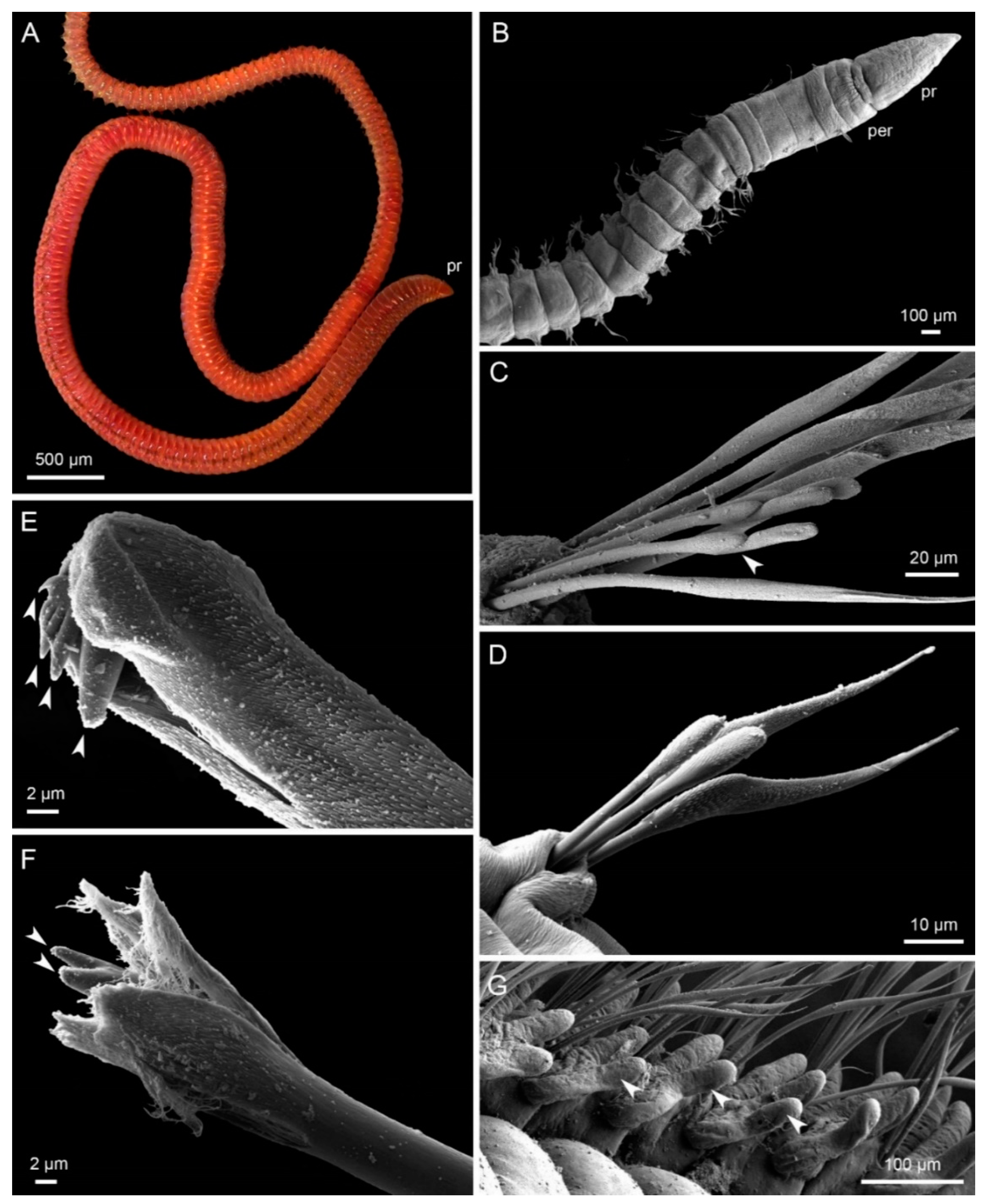{kind=link}
{kind=link}
{kind=link}
| Genus | Author | Type Species | Number of Species | Number of Species Used in This Study |
|---|---|---|---|---|
| Abyssoninoe | Orensanz, 1990 [2] | Lumbriconereis abyssorum McIntosh, 1885 [8] | 8 | 2 |
| Arabelloneris | Hartmann-Schröder, 1979 [9] | Arabelloneris broomensis Hartmann-Schröder 1979 [9] | 2 | 0 |
| Augeneria | Monro, 1930 [10] | Augeneria tentaculata Monro, 1930 [10] | 9 | 3 |
| Cenogenus | Chamberlin, 1919 [11] | Cenogenus descendens Chamberlin, 1919 [11] | 12 | 0 |
| Eranno | Kinberg, 1865 [12] | Eranno bifrons Kinberg, 1865 [12] | 11 | 0 |
| Gallardoneris | Carrera-Parra, 2006 [7] | Lumbrineris shiinoi Gallardo, 1968 [13] | 3 | 4 |
| Helmutneris | Carrera-Parra, 2006 [7] | Lumbriconereis flabellicola Fage, 1936 [14] | 3 | 1 |
| Hilbigneris | Carrera-Parra, 2006 [7] | Hilbigneris pleijeli Carrera-Parra, 2006 [7] | 3 | 1 |
| Kuwaita | Mohammad, 1973 [15] | Kuwaita magna Mohammad, 1973 [15] | 5 | 0 |
| Loboneris | Carrera-Parra, 2006 [7] | Lumbrineris pterignatha Gallardo, 1968 [13] | 1 | 0 |
| Lumbricalus | Frame, 1992 [16] | Lumbriconereis januarii Grube, 1878 [17] | 9 | 0 |
| Lumbrinerides | Orensanz, 1973 [18] | Lumbrinerides gesae Orensanz, 1973 [18] | 16 | 1 |
| Lumbrineriopsis | Orensanz, 1973 [18] | Lumbriconereis mucronata Ehlers, 1908 [19] | 5 | 2 |
| Lumbrineris | Blainville, 1828 [20] | Lumbrineris latreilli Audouin & Milne-Edwards, 1833 [21] | ~ 50 | 4 |
| Lysarete | Kinberg, 1865 [12] | Lysarete brasiliensis Kinberg, 1865 [12] | 3 | 0 |
| Ninoe | Kinberg, 1865 [12] | Ninoe chilensis Kinberg, 1865 [12] | 33 | 4 |
| Scoletoma | Blainville, 1828 [20] | Lumbricus fragilis O.F. Müller, 1776 [22] | 25 | 3 |
| Sergioneris | Carrera-Parra, 2006 [7] | Lumbrineris nagae Gallardo, 1968 [13] | 1 | 0 |
| Lumbrineridae gen. sp. | - | 1 | ||
| 199 | 26 |
| Clade | Taxon | Author | BOLD Process ID | GenBank Accession Number | Source | ||
|---|---|---|---|---|---|---|---|
| COI | 16S | 18S | |||||
| A | Lumbrineris mixochaeta | Oug, 1998 [31] | POLNB1550-15 | OM237808 | OM237844 | OM312047 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | POLNB1560-15 | OM237807 | OM237843 | OM312046 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | POLNB1562-15 | OM237806 | OM237842 | OM312045 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | EUNI007-21 | OM237805 | OM237841 | OM312044 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH004-21 | OM237804 | OM237840 | OM312043 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH005-21 | - | OM237831 | OM312034 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH006-21 | OM237803 | OM237839 | OM312042 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH007-21 | OM237802 | OM237838 | OM312041 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH008-21 | OM237801 | OM237837 | OM312040 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH009-21 | OM237800 | OM237836 | OM312039 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH010-21 | OM237799 | OM237835 | OM312038 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH011-21 | OM237798 | OM237834 | OM312037 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH012-21 | - | OM237833 | OM312036 | This study |
| A | Lumbrineris mixochaeta | Oug, 1998 | LUPH013-21 | - | OM237832 | OM312035 | This study |
| B | Scoletoma fragilis | (O.F. Müller, 1776) [22] | LUPH016-21 | OM237813 | - | OM312052 | This study |
| B | Scoletoma fragilis | (O.F. Müller, 1776) | POLNB827-14 | OM237814 | - | - | This study |
| C | Lumbrineridae gen. sp. | MIWAP303-13 | OM237794 | OM237827 | OM312031 | This study | |
| D | Gallardoneris sp. | MIWAP377-13 | OM237790 | OM237824 | - | This study | |
| D | Gallardoneris sp. | MIWAP379-13 | OM237788 | OM237822 | OM312027 | This study | |
| D | Gallardoneris sp. | MIWAP380-13 | OM237789 | OM237823 | OM312028 | This study | |
| D | Gallardoneris sp. | MIWAP376-13 | OM237789 | OM237823 | OM312026 | ||
| E | Scoletoma tetraura | (Schmarda, 1861) [1] | GU362689 | GU362682 | - | [32] | |
| E | Scoletoma zonata | (Johnson, 1901) [33] | - | HM746713 | HM746727 | [34] | |
| F | Hilbigneris sp. | MIWAP353-13 | OM237793 | OM237826 | - | This study | |
| F | Hilbigneris sp. | MIWAP352-13 | OM237792 | OM237825 | OM312030 | This study | |
| F | Hilbigneris sp. | LUPH014-21 | OM237791 | - | OM312029 | This study | |
| G | Abyssoninoe cf. hibernica | (McIntosh, 1903) [35] | SKAG032-11 | OM237776 | - | - | This study |
| G | Abyssoninoe cf. scopa | (Fauchald, 1974) [36] | POLNB779-14 | OM237778 | - | - | This study |
| G | Abyssoninoe cf. scopa | (Fauchald, 1974) | POLNB1742-15 | OM237777 | OM237815 | OM312019 | This study |
| H | Lumbrineris inflata | Moore, 1911 [37] | BOLD Process ID HZPLY357 | AY838832 | AY525622 | [26,38] | |
| H | Lumbrineris latreilli | Audouin and Milne-Edwards, 1834 [21] | KR916859 | AY838833 | - | [27,29] | |
| H | Lumbrineris sp. | MIWAP289-13 | OM237809 | OM237845 | OM312048 | This study | |
| H | Lumbrineris sp. | MIWAP291-13 | OM237811 | OM237846 | OM312050 | This study | |
| H | Lumbrineris sp. | MIWAP345-13 | OM237810 | - | OM312049 | This study | |
| I | Helmutneris vadum | Borisova & Budaeva, 2020 [39] | LUPH001-20 | MT763201 | MT763203 | - | |
| I | Helmutneris vadum | Borisova & Budaeva, 2020 | LUPH002-20 | MT763200 | MT763202 | - | |
| J | Ninoe armoricana | Glémarec, 1968 [40] | MIWAP295-13 | OM237812 | OM237847 | OM312051 | This study |
| J | Ninoe armoricana | Glémarec, 1968 | KT307669 | - | - | [41] | |
| J | Ninoe chilensis | Kinberg, 1865 [12] | JF731019 | - | - | [42] | |
| J | Ninoe leptognatha | Ehlers, 1900 [43] | JF731020 | - | - | [42] | |
| Ninoe nigripes | Verrill, 1873 [44] | AY838869 | AY838837 | AY838852 | [26,38] | ||
| K | Lumbrinerides sp. | MIWAP325-13 | OM237795 | OM237828 | OM312032 | This study | |
| L | Augeneria albidentata | (Ehlers, 1908) [19] | MIWAP298-13 | OM237780 | - | OM312020 | This study |
| L | Augeneria albidentata | (Ehlers, 1908) | MIWAP334-13 | OM237779 | OM237816 | - | This study |
| L | Augeneria albidentata | (Ehlers, 1908) | MIWAP335-13 | OM237781 | OM237817 | OM312021 | This study |
| L | Augeneria algida | (Wirén, 1901) | EUNI006-21 | OM237783 | OM237819 | OM312023 | This study |
| L | Augeneria algida | (Wirén, 1901) | LUPH003-21 | OM237782 | OM237818 | OM312022 | This study |
| L | Augeneria cf. tentaculata | Monro, 1930 | POLNB591-14 | OM237786 | - | - | This study |
| L | Augeneria cf. tentaculata | Monro, 1930 | POLNB932-14 | OM237784 | OM237820 | OM312024 | This study |
| L | Augeneria cf. tentaculata | Monro, 1930 | POLNB1745-15 | OM237785 | - | OM312025 | This study |
| M | Lumbrineriopsis sp. | EUNI012-21 | OM237796 | OM237829 | OM312033 | This study | |
| M | Lumbrineriopsis sp. | LUPH015-21 | - | OM237830 | - | This study | |
| M | Lumbrineriopsis sp. | MIWAP330-13 | OM237797 | - | - | This study | |
| Glycera alba | (O.F. Müller, 1776) [22] | KF369131 | DQ779615 | DQ779651 | [45,46] | ||
| Eunice rubra | Grube, 1856 [12] | KF808171 | GQ478132 | GQ497478 | [47,48] | ||
| Nothria conchylega | (Sars, 1835) [49] | HQ023895 | KJ027342 | KJ027382 | [50,51] | ||
| Tainokia logachevae | Ravara & Cunha, 2017 [52] | MF795582 | MF795579 | MF795580 | [52] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borisova, P.; Budaeva, N. First Molecular Phylogeny of Lumbrineridae (Annelida). Diversity 2022, 14, 83. https://doi.org/10.3390/d14020083
Borisova P, Budaeva N. First Molecular Phylogeny of Lumbrineridae (Annelida). Diversity. 2022; 14(2):83. https://doi.org/10.3390/d14020083
Chicago/Turabian StyleBorisova, Polina, and Nataliya Budaeva. 2022. "First Molecular Phylogeny of Lumbrineridae (Annelida)" Diversity 14, no. 2: 83. https://doi.org/10.3390/d14020083
APA StyleBorisova, P., & Budaeva, N. (2022). First Molecular Phylogeny of Lumbrineridae (Annelida). Diversity, 14(2), 83. https://doi.org/10.3390/d14020083