
Purely Australian Essential Oils Past and Present: Chemical Diversity, Authenticity, Bioactivity, and Commercial Value


Abstract
:1. Introduction
2. Progress in Research
2.1. Bioactivity of Australian Essential Oils
2.1.1. Insect Repellent or Insecticidal Essential Oils
2.1.2. Antimicrobial Effects of Essential Oils
2.1.3. Toxic Australian Essential Oils
2.2. Chemophenetics of Natural Volatiles and Essential Oils
2.2.1. Examples of Essential Oils in Australian Chemophenetic Studies
“no dataset has primacy in defining segregate taxa, and … a combination of morphological and molecular data was required to determine the taxa within.”
2.2.2. Examples of Solvent Extracted Volatiles in Australian Chemophenetic Studies
2.2.3. Chemophenetics as an Authentication Tool
3. Natural Volatiles in Ethnopharmacology
3.1. The Eucalyptus Paradox
3.2. How Volatile Organic Compounds Were Used in Ancient Australian History
4. Significant Genera and Industrial Progress
4.1. Prostanthera (Lamiaceae)
4.2. Eremophila and Myoporum (Scrophylariaceae)
4.3. Correa (Rutaceae)
4.4. Geiera (Rutaceae)
4.5. Melaleuca (Myrtaceae)
“The essential oil derived from this tree, which is one of a number of trees and shrubs known collectively as Tea Trees, is about to be placed upon the markets of the world.”[96]
4.6. Backhousia (Myrtaceae)
4.7. Leptospermum and Kunzea (Myrtaceae)
4.8. Philotheca and Eriostemon (Rutaceae)
4.9. Syzygium (Myrtaceae)
4.10. Calytrix (Myrtaceae)
4.11. Cassinia (Asteraceae)
4.12. Eucalyptus and Corymbia (Myrtaceae)
4.13. Phebalium and Leionema (Rutaceae)
4.14. Boronia (Rutaceae)
4.15. Callitris (Cupressaceae)
“there is nothing more delightful in the approach, on a winter evening, to a township where Cypress pine is used as a fuel. Its delicious perfume is borne on the air for miles, and is often the first intimation that the weary traveller experiences that he is approaching a human habitation, and that his long journey is drawing to a close.”[119]
4.16. Santalum (Santalaceae)
4.17. Lagarostrobos franklinii (Hook.F.) Quinn (Podocarpaceae)
4.18. Pittosporum (Pittosporaceae)
4.19. Zieria (Rutaceae)
4.20. Citrus (Rutaceae)
4.21. Tasmannia (Winteraceae)
4.22. Agonis fragrans (syn. Taxandria) (Myrtaceae)
5. Miscellaneous GC-MS Characterisations by Joseph Brophy (UNSW)

{kind=link}
| Species | Chemistry Info from Abstract | Family | Year and Ref. |
|---|---|---|---|
| Acmenosperma claviflorum (Roxb.) E. Kausel | Bicyclogermacrene (36%), α-copaene (7%), δ-cadinene (4.9%), globulol (5.2%) | Myrtaceae | 1999, [142] |
| Acradenia euodiiformis (F.Muell.), A. frankliniae Kippist | A. euodiiformis: type 1, α-pinene (16%), limonene (8%), p-cymene (13%), type 2, aromadendrene (11–22%), caryophyllene oxide (4–13%), globulol (11–13%), spathulenol (12–18%). A. frankliniae: 1.2–3.0% yield, type 1, xanthoxylin (90%), type 2, α-pinene (19–21%), camphene (13–14%), β-pinene (24–25%), (E)-β-ocimene (8–10%) | Rutaceae | 2001, [143] |
| Acronychia aberrans, A. acuminata, A. acidula, A. acronychioides, A. baeuerlenii, A. chooreechillum, A. crassipetala, A. eungellensis, A. imperforata, A. laevis, A. wilcoxiana, A. littoralis, A. octandra, A. oblongifolia, A. parviflora, A. pubescens, A. sp. (Batavia Downs, J.R.Clarkson + 8511), A. suberosa, A. vestita | A. aberrans: (Z)-ocimenone (40–55%), (E)-ocimenone (23–28%). A. acuminata: α-pinene (33–64%). A. acidula: δ-3-carene (32–40%), terpinolene (13–46%) α-santalene (2–15%), aromadendrene (2–8%), germacrene B (0.6–18%). A. acronychioides: spathulenol (37–52%). A. baeuerlenii: α-pinene (65%). A. chooreechillum: α-pinene (45–66%). A. crassipetala: 40% monoterpenes. A. eungellensis: α-pinene (21–26%). A. imperforata: β-caryophyllene (13–20%), bicyclogermacrene (21–26%). A. laevis: elemol, α-pinene, bicyclogermacrene. A. wilcoxiana and A. littoralis: β-caryophyllene, pregeijerene, geijerene. A. octandra: (Z)-β-ocimene (15–23%), (E)-β-ocimene (15–23%), limonene (4–21%). A. oblongifolia: α-pinene (34–87%), limonene (0.1–26%), terpinolene (0.1–29%). A. parviflora: β-caryophyllene (2–34%), allo-aromadendrene (0.3–20%). A. pauciflora: α-pinene (14–45%), β-caryophyllene (10–24%). A. pubescens: β-caryophyllene (15–39%). A. sp. (Batavia Downs): (3–21%). A. suberosa: (Z)-ocimenone (20–23%), (E)-ocimenone (9–13%), β-caryophyllene (4–10%). A. vestita: α-pinene (40%), β-caryophyllene (23.5%), limonene (67–80%) | Rutaceae | 2004, [144] |
| Actinodium cunninghamii | 90% α-Pinene | Myrtaceae | 1994, [145] |
| Actinostrobus pyramidalis, A. arenarius, A. acuminatus | A. pyramidalis: α-pinene (60–78%), limonene (1–17%), citronellal, citronellyl acetate, citronellol. A. arenarius: α-pinene (40–76%), limonene (1–28%), citronellal, citronellyl acetate, citronellol. A. acuminatus: α-pinene (17–79%), limonene (1–39%), spathulenol (6–17%) | Cupressaceae | 2004, [146] |
| Agathis atropurpurea, A. microstachya, A. robusta, A. australis, A. macrophylla, A. moorei, A. ovata | A. atropurpurea: phyllocladene (13%), 16-kaurene (19%), α-pinene (8%), δ-cadinene (9%). A. microstachya: α-pinene (18%) myrcene (7%), bicyclogermacrene (6%), δ-cadinene (6%). A. robusta: spathulenol (37%), rimuene (6%). A. australis: 16-kaurene (37%), sclarene (5%) germacrene-D (9%). A. macrophylla: 5,15-rosadiene (60%), 16-kaurene (7%). A. moorei: allo-aromadendrene (6%), germacrene-D, δ-cadinene (10%), 16-kaurene (6%). A. ovata: caryophyllene oxide (15%), phyllocladene (39%) | Araucariaceae | 2000, [147] |
| Agonis obtusissima F.Muell., (syn. Agonis baxteri (Benth.) J.R.Wheeler & N.G.Marchant) | α-Pinene (12%), trans-β-ocimene (16%), globulol (39%) | Myrtaceae | 2004, [148] |
| Allosyncarpia ternata S.T.Blake | β- and α-Pinene, limonene, β-caryophyllene, globulol, spathulenol | Myrtaceae | 1992, [149] |
| Angasomyrtus salina | α-Pinene (77–83%), campholenic aldehyde (1%), 1,8-cineole (0.1–1.0%), β-pinene (1–2%), bicyclogermacrene (1–3%) | Myrtaceae | 1994, [150] |
| Angophora spp. | Essential oils overlap with Eucalyptus | Myrtaceae | 1999, [151] |
| Araucaria angustifolia, A. bidwillii, A. columnaris, A. cunninghamii, A. heterophylla, A. hunsteinii, A. luxurians, A. montana, A. muelleri, A. scopulorum, | A. angustifolia: germacrene-D (9%), hibaene (30%), phyllocladene (20). A. bidwillii: hibaene (76%). A. columnaris: hibaene (9%), sclarene (6%), luxuriadiene (13-epi-dolabradiene) (23%). A. cunninghamii: 16-kaurene (53%), hibaene (29%). A. heterophylla: α-pinene (52%), phyllocladene (32%). A. hunsteinii: α-Pinene (18%), sclarene (11%), germacrene-D (5%). A. luxurians: 5,15-rosadiene (20%), luxuriadiene (13-epi-dolabradiene) (66%). A. montana: phyllocladene (61%), 16-kaurene (23%). A. muelleri: sclarene (20%), luxuriadiene (19%). A. scopulorum: 16-α-phyllocladanol (41%), luxuridiene (10%), δ-cadinene, α-copaene | Araucariaceae | 2000, [147] |
| Archirhodomyrtus beckleri (F. Muell.) A.J. Scott | Type 1, (E)-β-ocimene (69–87%), type 2, α-pinene, β-caryophyllene, α-terpineol, (E)-nerolidol and α-, β, and γ-eudesmol | Myrtaceae | 1996, [152] |
| Arillastrum gummiferum (Brongriart & Gris) Pancher ex Baillon | 80% (−)-limonene, α- and β-pinene, caryophyllene (3–7%) | Myrtaceae | 1994, [153] |
| Artabotrys sp. (Claudie River B.Gray 3240) | Oxygenated sesquiterpenes | Annonaceae | 2004, [154] |
| Asteromyrtus angustifolia, A. arnhemica, A. brassii, A. lysicephala, A. symphyocarpa | A. angustifolia: α-pinene (10%), 1,8-cineole (31%), β-caryophyllene (23%). A. arnhemica: α-pinene (92%). A. brassii: α-pinene (6%), 1,8-cineole (24%), γ-terpinene (21%). A. lysicephala: α-pinene (11.2%), 1,8-cineole (49%), α-terpinyl acetate (4%). A. magnifica: α-pinene (14%), β-pinene (20%), 1,8-cineole (36%). A. symphyocarpa: α-pinene (16%), 1,8-cineole (40%) | Myrtaceae | 1994, [155] |
| Austrobaileya scandens | β-Pinene (1.3–44.2%), α-pinene (3.1–30.3%), β-caryophyllene (2.3–13.0%), δ-cadinene (2.8–9.0%), spathulenol (0.9–11.9%) | Austrobaileyaceae | 1994, [156] |
| Austromatthea elegans L. S. Smith | Type 1, benzyl benzoate (96.25%), type 2, benzyl salicylate, benzyl benzoate | Austrobaileyaceae | 1995, [157] |
| Austromyrtus sp. nov. | (E)-β-Ocimene (83%), myrcene | Austrobaileyaceae | 1995, [158] |
| Austromyrtus dulcis (C. T. White) L. S. Smith, A. tenuifolia (Sm.) Burret., | A. dulcis: type 1, isobaeckeol (80%), type 2, β-pinene (34–45%), 1,8-cineole (24–35%). A. tenuifolia: 1.8–3.0% yield of isobaeckeol (97–98%) | Austrobaileyaceae | 1995, [159] |
| Austromyrtus gonoclada, A. floribunda, A. hillii, A. inophloia, A. minutiflora, A. pubiflora, A. shepherdii, A. sp. (Bamaga, B. P. Hyland 10235), A. sp. (Brookfield, L. W. Jessup 155), A. sp. (McIlwraith Range, B. P. Hyland 11148), A. sp. (Pinnacle Track, P.I. Forster PIF15535), A. acmenoides, A. bidwillii, A. dallachiana, A. fragrantissima, A. hillii type 2, A. racemulosa, A. sp. (Byerstown Range, G. P. Guymer 2037), A. sp. (Forty Mile Scrub, G. C. Stocker 1758), A. sp. (Mt. Beatrice, P. I. Forster PIF14662), A. sp. (Mt. Lewis, P.I. Forster PIF15613), A. sp. (Mt. White, P.I. Forster PIF13461), A. sp. (Danbulla, L. S. Smith 10123), A. sp. (Spencer Creek, P. I. Forster PIF13701) | A. gonoclada: 2-hydroxy-4,6-dimethoxy-3,5-dimethylacetophenone. Sesquiterpenoid oils: A. floribunda, A. hillii, A. inophloia, A. minutiflora, A. pubiflora, A. shepherdii, A. sp. (Bamaga), A. sp. (Brookfield), A. sp. (McIlwraith Range), A. sp. (Pinnacle Track). Monoterpenoid oils: A. acmenoides, A. bidwillii, A. dallachiana, A. fragrantissima, A. hillii type 2, A. racemulosa, A. sp. (Byerstown Range), A. sp. (Forty Mile Scrub), A. sp. (Mt. Beatrice), A. sp. (Mt. Lewis), A. sp. (Mt. White), A. sp (Danbulla), A. sp. (Spencer Creek) | Austrobaileyaceae | 1996, [160] |
| Austromyrtus lasioclada (F. Muell.) L.S. Sm., A. sp. (Blackall Range P.R. Sharpe 5387), A sp. (Upper Mudgeeraba Creek N.B. Byrnes +4069), A. sp. (Main Range P.R. Sharpe 4877) | A. lasioclada: δ-cadinene (8–14%), germacrene-D (4–12%). A. sp. (Blackall Range P.R. Sharpe 5387): α-copaene (12–16%), β-caryophyllene (4–20%), allo-aromadendrene (3–13%). A sp. (Upper Mudgeeraba Creek N.B. Byrnes +4069): α-copaene (8–15%), β-caryophyllene (3–8%) and allo-aromadendrene (8–14%). A. sp. (Main Range P.R. Sharpe 4877): spathulenol (8–12%), allo-aromadendrene (7–9%) | Austrobaileyaceae | 1995, [161] |
| Athrotaxis cupressoides, A. selaginoides, A. laxifolia | A. cupressoides: limonene (46–56%), spathulenol (3–10%), α-acorenol (8–13%) and 8-β-hydroxyisopimarene (2–24%). A. selaginoides: limonene (40–48%), spathulenol (4–11%), α-bisabolol (6–14%), rimuene (0.8–7%), 8-β-hydroxyisopimarene (11–29%). A. laxifolia: limonene (34–58%), spathulenol (4–10%), α-acorenol (7–18%), γ-acorenol (0.1–0.5%), α-bisabolol (0.6–4%), rimuene (1–4%) and 8-β-hydroxyisopimarene (2–18%) | Cupressaceae | 2002, [162] |
| Backhousia angustifolia F. Muell, B. anisata Vickery, B. bancroftii F. M. Bailey & F. Muell., B. citriodora F. Muell, B. hughesii C. T. White, B. kingii Guymer, B. myrtifolia Hooker & Harvey, B. sciadophora F. Muell., Backhousia sp. (Didcot P.I. Forster PIF12671) | B. angustifolia: 1,8-cineole, (E)-β-ocimene, angustifolenone, angustifolionol, dehydroangustione and angustione. B. anisata: type 1, (E)-anethole, type 2, methyl chavicol. B. bancroftii: octyl acetate (0.3–61.7%), dodecyl acetate (0.2–21.0%), dodecanol (trace—22.9%), decyl acetate (0.5–39.0%), decanol (0.1–17.4%), 2,4,6-trimethoxy-3-methylacetophenone (trace—23.0%), bancroftinone (trace—90.0%). B. citriodora: type 1, citral, type 2, citronellal. B. hughesii: β-bisabolene (1.0–44.0%) unidentified (8.0–54.0%). B. kingii: α-pinene (24.0–49.0%), limonene (7.0–24.0%), 1,8-cineole (10.0–17.0%). B. myrtifolia: type 1, methyl eugenol, type 2, (E)-methyl isoeugenol, type 3, elemicin, type 4, (E)-isoelemicin. B. sciadophora: α-pinene (44.0–55.0%), β-pinene (2.4–8.0%), limonene (6.5?12.7%), linalool (2.8–6.7%). Backhousia sp. (Didcot): α-pinene (11.0%), β-pinene (5.3%), β-caryophyllene (12.0%), dodecyl acetate (8.1%), dodecanol (8.2%) | Myrtaceae | 1995, [98] |
| Backhousia anisata Vickery | Type 1: E-anethole (90+ %), methyl chavicol (<5%), Z-anethole (<0.1%). Type 2: methyl chavicol (60–75%), E-anethole (<25%), Z-anethole (<0.1%) | Myrtaceae | 1991, [163] |
| Backhousia citriodora F. Muell. | The l-citronellal type of B. citriodora described by Penfold was lost until 1996: 85–89% citronellal, 6–9% isopulegol isomers, citronellol (approx. 3%) | Myrtaceae | 2001, [100] |
| Barongia lophandra | α-Pinene (58%), β-pinene (19%) | Myrtaceae | 2003, [164] |
| Bosistoa brassii, B. floydii, B. medicinalis, B. pentacocca, B. pentacocca var. connaricarpa, B. pentacocca var. dryanderensis, B. pentacocca var. pentacocca, B. selwynii, and B. transversa | B. brassii: β–caryophyllene (2–12%), α-humulene (1–18%,), bicyclogermacrene (trace—24%). B. floydii: α-pinene (46–67%). B. medicinalis: α–pinene (13–57%), β–caryophyllene (1–9%). B. pentacocca: δ–cadinene (6–11%), α–copaene (2–7%), germacrene D (4–14%), α–cadinol (2–8%). B. transversa: α–pinene (24–83%) | Rutaceae | 2007, [165] |
| Bouchardatia neurococca (F. Muell.) Baillon | β-Caryophyllene (38.5%), α-humulene (16.1%), bicyclogermacrene (10.8%), caryophyllene oxide (13.0%) | Rutaceae | 1994, [166] |
| Brombya platynema, B. sp. (Gap Creek L.S. Smith 11116) | B. platynema: germacrene D (11–78%), β-bisabolene (0.8–22%), bicyclogermacrene (14–22%), kessane (1–17%), type 2: β-bisabolene (19–53%), curcumene (26.7%), bicyclogermacrene (7.7%), ar-curcumene (10.6%). B. sp. (Gap Creek): β-bisabolene (22.9%), ar-curcumene (14.6%), α-santalene (9.2%) | Rutaceae | 2004, [167] |
| Callistemon viminalis (Sol. ex Gaertner) G.Don ex Loudon | α-Pinene, β-pinene, myrcene, 1,8-cineole, leptospermone, flavesone. | Myrtaceae | 1997, [168] |
| Callistemon spp (high yields), low yield species, C. brachyandrus, C. montanus, C. polandii, C. teretifolius, C. sp. (Walsh’s Pyramid P.I. Forster 13767), C. sp. nov. Oakey, C. pachyphyllus, | High yielding species: 1,8-cineole (45–80%), α-pinene (2–40%), limonene (2–9%), α-terpineol (1–13%). Low yielding species: 1,8-cineole (<20%), sesquiterpenoid | Myrtaceae | 1998, [169] |
| Cananga odorata | β-Caryophyllene (34–52%), sabinene (1–20%), α-humulene (6–11%), α-pinene (1–17%) | Annonaceae | 2004, [154] |
| Choricarpia subargentea (C.T. White) L.A.S. Johnson, C. leptopetala (F. Muell.) Domin | C. subargentea: α-pinene (30–76%), limonene (2–55%), 1,8-cineole (2–20%), jensenone. C. leptopetala: α-pinene, limonene, p-cymene, 1,8-cineole | Myrtaceae | 1994, [170] |
| Cinnamomum baileyi, C. oliveri, C. baileyi, C. oliveri, C. propinquum, C. virens | C. baileyi: methyl eugenol, bicyclogermacrene. C. oliveri: camphor, safrole, methyl eugenol. C. laubatii: type 1, safrole, methyl eugenol, elemicin and bicyclogermacrene, type 2, bicyclogermacrene, β-selinene, spathulenol and γ-eudesmol. C. propinquum: p-cymene, β-eudesmol. C. virens: α-pinene, (E)-nerolidol | Lauraceae | 2001, [171] |
| Clausena brevistyla Oliv. and C. smyrelliana P.I.Forst. | C. brevistyla: type 1, myrcene (72.4%), type 2, limonene (83.3%), type 3, β-caryophyllene (19.4%), α-humulene (4.3%), bicyclogermacrene (9.5%), caryophyllene oxide (7.6%), spathulenol (10.6%). C. smyrelliana: α-pinene (73.3%), β-caryophyllene (8.5%) | Rutaceae | 2016, [172] |
| Coatesia paniculata F.Muell., syn. Geijera paniculata. | Leaf oil: α-pinene (27–57%), β-caryophyllene (4–12%) | Rutaceae | 2005, [173] |
| Coleonema pulchellum Williams | α- and β-Pinene, myrcene, β-phellandrene, linalool, terpinen-4-ol, caryophyllene, germacrene-D, bicyclogermacrene | Rutaceae | 1986, [174] |
| Corymbia dallachiana (Benth.) K.D.Hill & L.A.S.Johnson | Papuanone | Myrtaceae | 1999, [175] |
| Crowea exalata, C. saligna, C. angustifolia var. angustifolia | C. exalata: Type 1, safrole (81–88%), type 2, (E)-methyl isoeugenol (18–25%), (E)-carpacin (47–51%), type 3, safrole (27–35%), (E)-methyl isoeugenol (29–46%), α-pinene (12–25%), type 4, safrole (6–29%), asaricin (57–74%), type 5, exalatacin, croweacin (10–20%). C. saligna: croweacin (84–94%), safrole. C. angustifolia var. angustifolia: β-asarone (68%), exalatacin (13%), croweacin (7%) | Rutaceae | 1997, [176] |
| Crowea exalata F.Muell | exalaticin | Rutaceae | 2000, [11] |
| Cryptocarya bellendenkerana, C. cocosoides, C. cunninghamii, and C. lividula (Lauraceae) | C. cocosoides: bicyclogermacrene (3–26%), spathulenol (16–47%), massoia lactone (11–15%), (6-heptyl-5,6-dihydro-2H-pyran-2-one (0.3–3%) and benzyl benzoate (0.2–5%). C. cunninghamii: benzyl benzoate (80.2%). C. bellendenkerana: limonene (8.3%), β-phellandrene (11.8%), viridiflorene (9.1%). C. lividula: bicyclogermacrene (26.1%), spathulenol (21.1%), β-eudesmol (6.1%) | Lauraceae | 2016, [177] |
| Cryptocarya cunninghamii Meissner | Type 1, bicyclogermacrene (52.4%), type 2, 6-nonyl-5,6-dihydro-2H-pyran-2-one (78–88%) | Lauraceae | 1998, [178] |
| Cyathostemma micranthum | Caryophyllene oxide (26%), spathulenol (11%), benzyl benzoate (4%) | Annonaceae | 2004, [154] |
| Darwinia citriodora (Endl.) Benth | Yield: 0.5% to 1.1%: methyl myrtenate (56–76%) methyl geranate (49–75%), α-pinene (7.8%), (Z)-β-ocimene (2.0%), linalool (1.7%), bicyclogermacrene (1.3%), viridiflorol (2.3%) | Myrtaceae | 2001, [179] |
| Darwinia procera, D. fascicularis subsp. fascicularis and D. peduncularis | D. procera: myrtenyl acetate (6.1–29.6%), α-pinene (6.9–25.1%), γ-terpinene (6.2–13.6%), bicyclogermacrene (5.5–10.8%), (E)-nerolidol (3.4–9.7%). D. fascicularis ssp. fascicularis: (E)-nerolidol (33.0%), α-pinene (15.1%), γ-terpinene (10.2%). D. peduncularis: α-pinene (33.5%), γ-terpinene (23.1%), bicyclogermacrene (6.7%) | Myrtaceae | 2010, [180] |
| Decaspermum humile (Sweet ex G.Don) A.J.Scott, D. struckoilicum N.Snow & Guymer | D. struckoilicum: α-pinene (37.5%), β-caryophyllene (2.4%), α-humulene (2.2%) and α- and β-eudesmol (8.2% and 8.1%, respectively). D. humile: a-thujene (0.1–13%), α-pinene (0.2–21%), limonene (0.2–8%), myrcene (0.3–10%), β-phellandrene (0.1–5%), linalool (0.3–9%) and terpinen-4-ol (0.3–6%), β-caryophyllene (0.7–5%), aromadendrene (1–6%), viridiflorene (1–7%), δ-cadinene (0.4–14%), bicyclogermacrene (0.2–10%), globulol (1–9%) | Myrtaceae | 2005, [181] |
| Desmos goezeanus, D. spp. (Mossman River L.W. Jessup 550), D. wardianus, | D. goezeanus: benzyl benzoate, benzyl salicylate. D. spp. (Mossman River): benzyl benzoate (52%). D. wardianus: α-pinene (37%) | Annonaceae | 2002, [182] |
| Dinosperma erythrococca, D. stipitata, D. melanophloia, | D. erythrococca: type 1, geranyl acetate (80%), type 2, (E)-β-ocimene (3–28%), geranyl acetate (2–32%), linalool (2–10%), β-caryophyllene (7–11%), bicyclogermacrene (2–10%), spathulenol (1–10%), type 3, spathulenol (30%), type 4, furanoid linalool oxides (5–8%), (E)-β-ocimene (13%), γ-elemene (36.4%), type 5, myrcene (13.3%), limonene (26.3%), (E)-β-ocimene (18.5%). D. stipitata: type 1, bicyclogermacrene (22–32%), germacrene B (11–12%) evodionol methyl ether (11.5%), (E)-methyl isoeugenol (6–11%), type 2, hydrocarbon C15H,4 (9–26%), bicyclogermacrene (7–16%), γ-elemene (7–9%). D. melanophloia: methyl chavicol (59%), (E)-methyl isoeugenol (15%) | Rutaceae | 2002, [183] |
| Dinosperma longifolium T.G. Hartley (Rutaceae) | Niranin, S-methylthiocarboxylic acid-N-methyl-N-phenylethylamide, (39–62%), β-caryophyllene (5–12%), (E)-β-farnesene (5–12%), bicyclogermacrene (10–14%) | Rutaceae | 2004, [184] |
| Diselma archeri Hook.f. | Yield 0.6–0.8%: Leaves, α-pinene (45–73%), δ-3-carene (1–15%), limonene | Cupressaceae | 2003, [185] |
| Doryphora sassafras Endl., D. aromatica (F. M. Bail.) L. S. Smith | D. sassafras: methyl eugenol (27–47%), safrole (15–30%), camphor (15–19%). D. aromatica: α-, β- and γ-eudesmol, spathulenol, elemol, guaiol | Atherospermataceae | 1993, [186] |
| Drummondita calida (F.Muell.) Paul G.Wilson | α-pinene (79–86%) | Rutaceae: Boronieae | 2006, [187] |
| Dryadodaphne sp. (Mt. Lewis B.P. Hyland RFK1496) | Leaves: δ-cadinene (10.7%), globulol (4.6%), T-cadinol (3.7%). Bark and wood: guaiol, bulnesol | Atherospermataceae | 1998, [188] |
| Endressia wardellii (F.Muell.) Whiffin | E. wardellii: α-humulene (14–17%), bicyclogermacrene (17–24%) | Apiaceae | 2009, [189] |
| Eremaea pauciflora (Endl.) Druce | 1,8-Cineole (22%), eudesmols (α-, β-, γ-; 26%) | Myrtaceae | 2004, [148] |
| Eriostemon banksii A. Cunn. ex Endl, E. australasius | E. australasius and E. banksia: cis and trans-methyl-4-isoprenoxycinnamate, β-elemene (7.2–8.1%), β-caryophyllene (7.7–8.4%) sesquiterpene n.d. (12.3–15.1%), α-pinene (2.8–8.3%), β-pinene (0.2–0.8%), limonene (0.4–0.5%) | Rutaceae | 1998, [190] |
| Eryngium expansum F. Muell, E. pandanifolium Cham. et Schlecht, E. rostratum Cav., E. vesiculosum Labill. | E. expansum: 7-epi-α-selinene (38.3%), cis-β-guaiene (10.8%), 2,3,6-trimethylbenzaldehyde (8.0%), (E,E)-α-farnesene (7.3%). E. pandanifolium: bornyl acetate (20.8%), β-selinene (13.8%), α-selinene (11.3%), α-muurolene (8.0%). E. rostratum: spathulenol (20.0%), β-bisabolol (8.6%), fruit oil: β-bisabolol (65.3%). E. vesiculosum: β-caryophyllene (20.3%), germacrene D (19.2%), α-humulene (8.8%) | Apiaceae | 2003, [191] |
| Eryngium paludosum (C.Moore) P.W.Michael | γ-terpinene (12.9%), β-bisabolene (12.2%), germacrene D (7.6%), myrcene (7.3%), β-caryophyllene (7.0%), limonene (6.0%), α-humulene (5.1%) | Apiaceae | 2008, [192] |
| Eryngium rosulatum P.W. Michael Ined | β-elemene (16.0%), bicyclogermacrene (12.5%), δ-elemene (7.0%), (E)-caryophyllene (5.9%) | Apiaceae | 2006, [193] |
| Eryngium vesiculosum Labill. | Caryophyllene (20.3%), germacrene D (19.2%), α-humulene (8.8%), bicyclogermacrene | Apiaceae | 2003, [194] |
| Euodia hylandii, E. pubifolia | E. hylandii: spathulenol (12–20%). E. pubifolia: spathulenol (18.3%) | Rutaceae | 2004, [195] |
| Fitzalania heteropetala F. Muell. | β-Caryophyllene (33–8%), aromadendrene (14.0%) | Annonaceae | 1997, [196] |
| Flindersia acuminata, F. australis, F. bennettiana, F. bourjotiana, F. brassii, F. brayleyana, F. collina, F. dissosperma, F. ifflaiana, F. laevicarpa, F. maculosa, F. oppositifolia, F. pimenteliana, F. schottiana, and F. xanthoxyla | F. maculosa/F. dissosperma: geijerene, pregeijerene, methyl geranate, α-pinene, β-caryophyllene and bicyclogermacrene. F. acuminata/F. australis: bicyclogermacrene, guaiol, bulnesol. F. australis type 2: β-caryophyllene, spathulenol. F. bennettiana: bicyclogermacrene. F. bourjotiana: β-caryophyllene. F. brassiii: α-cadinol, δ-cadinene. F. brayleyana: spathulenol, caryophyllene oxide. F. collina: (E,E)-farnesol. F. ifflaiana: β-caryophyllene, α-humulene, bicyclogermacrene. F. Laevicarpa: β-caryophyllene, germacrene D, bicyclogermacrene, elemol. F. oppositifolia: diverse. F. pimenteliana: β-caryophyllene, bicyclogermacrene, chemotype 2: α-pinene. F. schottiana: α-pinene, sabinene. F. xanthoxyla: bicyclogermacrene, δ-cadinene, β-caryophyllene | Rutaceae | 2005, [50] |
| Galbulimima baccata F.M.Bailey (Himantandraceae) | Elemol/hedycaryol (12–30%), α-, β- and γ-eudesmol (0.6–3%, 0.4–3% and 0.2–3%, respectively), spathulenol (1–3%) | Himantandraceae | 2005, [197] |
| Geijera linearifolia (DC.) J.M.Black | Leaf oil: spathulenol (10–17%), geranyl acetate (4–9%), bicyclogermacrene (3–6%), (E,E)-farnesol (23–30%) | Rutaceae | 2005, [173] |
| Geleznowia verrucosa Turcz | Leaves: α-pinene (80%), flower, α-pinene + eugenyl acetate | Rutaceae | 1995, [198] |
| Goniothalamus australis Jessup | Type 1, pinocarvone (10%), trans-pinocarveol (17%), type 2, α-pinene (10–11%) | Annonaceae | 2004, [154] |
| Gyrocarpus americanus Jacq., subsp. americanus | α-pinene, β-pinene, germacrene D (31%) | Hernandiaceae | 2000, [199] |
| Halfordia kendack (Montrouz.) Guillaumin S.L. | methyl eugenol and elemicin | Rutaceae | 2004, [200] |
| Haplostichanthus johnsonii F.Muell., H. sp. (Coopers Creek B.Gray 2433), H. sp. (Johnstone River L.W Jessup+ 471), H. sp. (Mt. Finnigan L.W Jessup 632), H. sp. (Rocky River Scrub P.I. Forster+ PIF10617), H. sp. (Topaz L.W. Jessup 520) | H. sp. (Rocky River Scrub): caryophyllene oxide (26.2%), humulene oxide (10.1%), spathulenol (31.6%). H. sp. (Mt. Finnigan): spathulenol (15.4%). H. sp. (Coopers Creek): β-caryophyllene (10%), γ-muurolene (12.4%), bicyclogermacrene (9.6%). H. sp. (Johnstone River): β-caryophyllene (trace—27%), α-humulene (trace—10%), caryophyllene oxide (3–19%), spathulenol (6–31%). H. sp. (Topaz): spathulenol (24–38%). H. johnsonii: spathulenol (21–36%) | Annonaceae | 2006, [201] |
| Haplostichanthus johnsonii | Yield 1.5–2%. 2,3,4,5-tetramethoxyallylbenzene (79%), elemicin (5.9%), α-copaene (5%), elemene (2%) | Annonaceae | 1992, [202] |
| Hedycarya angustifolia A.Cunn. and H. loxocarya (Benth.) W.D.Francis | Elemol and α-, β- and γ-eudesmol | Monimiaceae | 2005, [203] |
| Hernandia albiflora, H. bivalvis, H. nymphaeifolia | H. albiflora: bicydogermacrene (trace -19%), β-caryophyllene (5–9%), caryophyllene oxide (7–18%), globulol (4–6%). H. bivalvis: bicyclogermacrene (20–30%), β-caryophyllene (4–13%), α-copaene (11–13%), germacrene D (7–12%). H. nymphaeifolia: β-caryophyllene (11–44%), α-humulene (14–17%), caryophyllene oxide (5–20%) | Hernandiaceae | 2000, [199] |
| Homoranthus biflorus, H. binghiensis, H. cernuus, H. flavescens, H. montanus, H. sp. nov. Nandewar Range, H. bornhardtiensis, H. decumbens, H. homoranthoides, H. prolixus, H. decumbens, H. thomasii, H. tropicus | H. biflorus, H. binghiensis, H. cernuus type 1, H. flavescens, H. montanus and H. sp. nov. Nandewar Range type 1: β-pinene, (Z)-β-Ocimene (>40%), bicyclogermacrene, globulol. H. bornhardtiensis, H. decumbens type 1, H. homoranthoides, H. prolixus, H. sp. nov. Nandewar Range type 2: β-pinene, limonene, bicyclogermacrene, globulol. H. decumbens, H. thomasii, H. tropicus, H. sp. nov. Nandewar Range type 3: β-pinene, γ-terpinene, bicyclogermacrene, globulol. H. tropicus: methyl geranate, bicyclogermacrene, globulol | Myrtaceae | 2004, [204] |
| Homoranthus montanus Craven and S. R. Jones, H. flavescens Cunn. ex. Schauer. | H. montanus: (Z)-β-ocimene (85%). H. flavescens: (Z)-β-ocimene (69–71%) | Myrtaceae | 1998, [205] |
| Idiospermum australiense (Diels) S. T. Blake | Bicyclogermacrene (48%), caryophyllene (8%), globulol, viridiflorol, spathulenol | Calycanthaceae | 1992, [206] |
| Kibara rigidifolia A.C.Sm. | (Z)-β-Ocimene (3–12%), (E)-β-ocimene (1.5%), bicyclogermacrene, germacrene-B, guaiol, spathulenol | Monimiaceae | 1998, [188] |
| Kunzea pulchella (Lindl.) A.S.George | Globulol (83–88%) | Myrtaceae | 2004, [148] |
| Lagarostrobos franklinii (Hook.f.) Quinn | Yield 0.8–0.9%: Leaves, α-pinene (13–36%), δ-3-carene (1–17%), limonene (16–42%), 16-kaurene (3–7%), phyllocladene (4–10%), sclarene (2–23%). Wood, methyl eugenol (74%), (E)-methyl isoeugenol (2%) elemicin (24%) | Podocarpaceae | 2003, [185] |
| Leionema ambiens (F.Muell.) Paul G.Wilson | (E)-β-ocimene (>10%), (Z)-β-ocimene (0.1–4%), viridiflorene (6–7%), bicyclogermacrene (6–13%), globulol (6–7%), viridiflorol (4–6%), (E,E)-farnesol (14–23%) | Rutaceae | 2003, [117] |
| Leptospermum amboinense, L. emarginatum, L. grandiflorum, L. liversidgei, L. petersonii, L. rotundifolium, L. wooroonooran | L. amboinense: type 1, geranial (13%), sabinene (13%). L. emarginatum: α-eudesmol (7–17%), β-eudesmol (17–26%), γ-eudesmol (9–18%). L. grandiflorum: α-, β- and γ-eudesmol. L. liversidgei: citronellal (ca. 44%), neral (20%), geranial (35%). L. petersonii: type 1, citronellal, low neral/geranial, type 2, low citronellal and high neral/geranial, type 3, Penfold’s ‘variety A,’ monoterpenes, type 4, β-caryophyllene, globulol/viridiflorol/spathulenol, type 4, Penfold’s ‘variety B,’ geranyl acetate (21–38%), geraniol (21–29%). L. rotundifolium: α-pinene (16–25%), 1,8-cineole (21–28%). L. wooroonooran: α-pinene (4–11%), β-pinene (4–9%), sabinene (9–19%), β-caryophyllene (5–7%), humulene (11–20%) | Myrtaceae | 2000, [207] |
| Leptospermum arachnoides, L. crassifolium, L. deuense, L. epacridoideum, L. glabrescens, L. grandifolium, L. lanigerum, L. macrocarpum, L. nitidum, L. petraeum, L. riparium, L. spectabile, L. sphaerocarpum, L. thompsonii, L. turbinatum | Oils were dominated by α-, β- and γ-eudesmol. L. glabrescens: flavesone, leptospermone, eudesmol isomers | Myrtaceae | 1999, [208] |
| Leptospermum blakelyi, L. brevipes, L. neglectum, L. parvifolium, L. sp. (Woodgate, P. I. Forster, PIF 13959), L. multicaule, L. namadgiensis, L. microcarpum | L. brevipes, L. neglectum, L. parvifolium, L. sp. (Woodgate): α-pinene. L. blakelyi, L. multicaule, L. namadgiensis and L. sericatum: sequiterpenoid. L. divaricatum and L. microcarpum: monoterpenoid | Myrtaceae | 1998, [209] |
| Leptospermum brachyandrum (F. Muell.) Druce, L. luehmannii F. M. Bailey, L. madidum A. R. Bean subsp. madidum, L. purpurascens Joy Thomps., L. speciosum Schauer, L. whitei Cheel and L. pallidum A. R. Bean | All species produce α-pinene and lesser amounts of β-pinene, β-caryophyllene, aromadendrene, humulene, spathulenol, 1,8-cineole | Myrtaceae | 1998, [210] |
| Leptospermum coriaceum (F. Muell. Ex Miq.) Cheel, L. fastigiatum S. Moore, and L. nitens Turcz | L. coriaceum: α-pinene (25.4%), 1,8-cineole (11.5%), globulol (15.4%). L. fastigiatum and L. nitens: α-pinene (82.8% and 64.8–70.6% respectively) | Myrtaceae | 1999, [211] |
| Leptospermum spp. Eudesmol types: [L. micromyrtus Miq., L. minutifolium C.T. White, L. myrtifolium Sieber ex DC., L. rupestre Hook. f., L. sejunctum Joy Thomps.] L. novae-angliae Joy Thomps., L. rupicola Joy Thomps. Sesquiterpenoid types [L. continentale Joy Thomps., L. gregarium Joy Thomps., L. juniperinum Sm, L. obovatum Sweet., L. scoparium J. R. Forst & G. Forst. and L. squarrosum Gaertn.] | Eudesmol types dominated by α-, β- and γ-eudesmol. L. myrtifolium type 2: (E,Z)-farnesal (5.8%), (E,E)-farnesal (12.9%), 2,3-dihydro-(E)-farnesol (10.3%), (E,E)-farnesol (26.5%). L. novae-angliae: (E)-nerolidol (50%). L. rupicola: α- and β-pinene | Myrtaceae | 1999, [101] |
| Leptospermum morrisonlii, L. oreophilum, L. variabile, Leptospermum sp. (Mt Maroon, A.R. Bean 6665), L. polygalifolium, ssp. polygalifolium, montanum, howense, cismontanum, transmontanum, tropicum and ‘wallum,’ L. madidum spp. sativum | L. morrisonlii: grandiflorone. L. oreophilum: (E,E)-farnesol. L. variabile: type 1, geranyl acetate, β-caryophyllene, humulene, type 2, 1,8-cineole, type 3, α-pinene, β-caryophyllene, α-, β- and γ-eudesmol. L. sp. (Mt Maroon): type 1, β-caryophyllene, humulene, type 2, β-caryophyllene, δ-cadinene, calamenene, sesquiterpene n.d. L. polygalifolium, ssp. polygalifolium, montanum and howense: α-, β-, and γ-eudesmol. L. ssp. cismontanum, transmontanum, tropicum and ‘wallum’: 1,8-cineole. L. madidum spp. sativum: α-pinene, β-pinene, γ-terpinene α-, β-, and γ-eudesmol | Myrtaceae | 2000, [212] |
| Leptospermum scoparium | Isoleptospermone | Myrtaceae | 1999, [175] |
| Levieria acuminata (F.Muell.) Perkins | n-Dodecanal (28%), δ-cadinene (5.8%), calamenene (5.7%) | Monimiaceae | 1998, [188] |
| Lindera queenslandica B. Hyland | γ-Elemene (21.4%), α-copaene (17.9%), β-caryophyllene (7.4%), α-humulene (9.0%) | Lauraceae | 1999, [213] |
| Lindsayomyrtus racemoides (Greves) Craven | β-Caryophyllene (7.4–13%), humulene (5.2–11.6%), β-trans-ocimene (5.0–7.3%) | Myrtaceae | 1996, [214] |
| Lophostemon Schott spp. | α-Pinene, aromadendrene, allo-aromadendrene, globulol, spathulenol. | Myrtaceae | 2000, [215] |
| Lunasia amara Blanco var. amara | γ-Elemene (0.7–19%), germacrene-D (18–51%), bicyclogermacrene (7–26%), bicycloelemene (1–2%), β-bourbonene (0.7–3%), γ-elemene (4–9%), α-farnesene (1–3%), δ-cadinene (3–5%) | Rutaceae | 1997, [216] |
| Lycopus australis R.Br. | β-phellandrene (26–40%), β-caryophyllene (7–16%), α-humulene (18–30%) | Lamiaceae | 2005, [217] |
| Lysicarpus angustifolius (Hook.) Druce | α-Pinene, β-pinene, limonene (8%), α-terpineol (2–5%), viridiflorene (5–8%), aromadendrene (5–8%), globulol (4–5%), spathulenol (2–5%) | Myrtaceae | 1994, [218] |
| Malleostemon tuberculatus (E.Pritz.) J.W.Green | α-Pinene (33%), 1,8-cineole (21%), E,E-farnesol (6%) | Myrtaceae | 2004, [148] |
| Medicosma cunninghamii, M. elliptica, M. fareana, M. glandulosa, M. obovata, M. riparia, M. sessiliflora, M. sp. (East Mulgrave River R.L. Jago + 3696), M. sp. (Karnak P.I. Forster+ PIF15541), M. sp. (Mt Mellum P.I. Forster + PIF25572) | M. cunninghamii: evodionol, evodionol methyl ether, α-pinene, myrcene, ocimene. M. elliptica: α-pinene (0.6–29%), sabinene (12–55%), myrcene (8–16%), limonene (0.4–13%). M. fareana: bicyclogermacrene (15–44%). M. glandulosa: α-pinene (t-35%), β-caryophyllene (t-15%), aromadendrene (t-10%), (E,E)-α-farnesene (1–13%), bicyclogermacrene (1–13%), globulol (1–8%), spathulenol (3–10%). M. obovate: α-pinene (1–12%), limonene (10–13%), (E)-β-ocimene (14–23%), β-caryophyllene (17–19%), α-humulene (6–14%). M. riparia: α-selinene (7–15%), evodionol (1–3%), evodionol methyl ether (30–56%). M. sessiliflora: β-caryophyllene (3–11%), aromadendrene (5–14%), humulene (8–19%), spathulenol (3–12%). M. sp. (East Mulgrave) bicyclogermacrene (17–21%). M. sp. (Karnak): α-pinene (1–40%), bicyclogermacrene (3–8%), spathulenol (10–22%). M. sp. (Mt Mellum): α-pinene (38–54%) (Z)-β-ocimene (10–13%) | Rutaceae | 2004, [219] |
| Melicope affinis, M. bonwickii, M. broadbentiana, M. elleryana, M. fellii, M. hayesii, M. jonesii, M. micrococca, M. peninsularis, M. rubra, M. vitiflora, M. xanthoxyloides | M. affinis: bicyclogermacrene (7–18%), bisabolene (t-9%). M. bonwickii: zierone (0.3–3%). M. broadbentiana: α-pinene (21–76%), limonene (0.6–28%). M. elleryana: zierone (26–42%), allo-evodione (4–10%), evodione (10–22%). M. fellii: β-caryophyllene (9.9%), α-humulene (8.4%), caryophyllene oxide (7.4%). M. hayesii: bicyclogermacrene (22.8%), germacrene D (13.9%), (E,E)-α-farnesene (9.2%), globulol (10.6%). M. jonesii: sesquiterpenic. M. micrococca: α-pinene (1–46%), (E)-(β-ocimene (t-10%), (β-caryophyllene (0.4–15%), bicyclogermacrene (t-11%), caryophyllene oxide (0.3–23%), spathulenol (1–12%). M. peninsularis: β-caryophyllene (30–49%), α-humulene (26?35%). M. rubra and M. vitiflora: sabinene (31.1%), γ-terpinene, germacrene D (22.6%), sabinene (0.1–54%), limonene (1–47%). M. xanthoxyloides: β-caryophyllene (13–47%), spathulenol (1–18%), α-pinene (t-15%) | Rutaceae | 2004, [195] |
| Melicope contermina, M. polybotrya | M. conterminal: limonene (33%), elemol (23%). M. polybotrya: geijerene (41%), pregeijerene (38%), limonene (9%) | Rutaceae | 2004, [220] |
| Melicope melanophloia C.T. White | Type 1, methyl chavicol (5–13%), methyl eugenol (51–67%), type 2, α-pinene (34–37%), myrcene, α-phellandrene, limonene, 1,8-cineole (4–12%), type 3, limonene (1–8%), (Z)-β-ocimene (12–18%), (E)-β-ocimene (23–56%), 2-hydroxy-4,6-dimethoxyacetophenone (4–6%) | Rutaceae | 1997, [221] |
| Melodorum sp. (Font Hills G. Sankowsky 380), M. sp. (Stone Crossing L.W. Jessup 814), M. sp. (Topaz G. Sankowsky + 244), M. sp. (Claudie River B.P.Hyland 21171V), M. uhrii, M. leichhardtii, | M. sp. (Font Hills): α-eudesmol (9–5%), β-eudesmol (7–11%), β-caryophyllene (10–16%), bicyclogermacrene (1–9%) and α-pinene (14–15%). M. sp. (Stone Crossing): benzyl benzoate (20%), benzyl salicylate (2.7%). M. sp. (Topaz): bicyclogermacrene (34–50%), β-caryophyllene (11–16%), spathulenol (2–10%). M. sp. (Claudie River): bicyclogermacrene (29.3%), β-caryophyllene (26.7%). M. uhrii: bicyclogermacrene (45%), benzyl benzoate (5%). M. leichhardtii: germacrene D (6–10%), bicyclogermacrene (15–19%), (Z)-β-ocimene (6?8%), (E)-β-ocimene (2–5%) | Annonaceae | 2004, [222] |
| Mentha diemenica Sprengel | 1% Yield, menthone (32%), neomenthyl acetate (0.0–18.3%), pulegone (25–44%), neomenthol (2.5–9.0%), menthyl acetate (2.7–5.7%), menthol (1.8–2.7%), isomenthone (1–3%) | Lamiaceae | 1996, [223] |
| Mentha grandiflora Benth | trans-piperitone oxide (21%), piperitenone oxide (36%), pulegone (19%), menthone (10%) | Lamiaceae | 1997, [224] |
| Microcachrys tetragona (Hook.) Hook.f. | Yield 0.1–0.5%: α-pinene (44–50%), limonene (10–13%) | Podocarpaceae | 2003, [185] |
| Micromelum minutum (G.Forst.) Wight & Arn. | M. minutum: δ-elemene (3.3–18.6%), β-caryophyllene (4.8–30.3%), germacrene-D (2–15.8%), germacrene-B (3.9–34.3%) | Rutaceae | 2016, [172] |
| Micromyrtus striata J. W. Green | Isoamylisovalerate (23.0–48.7%), α-pinene (5.9–27.7%) | Myrtaceae | 1991, [225] |
| Microstrobos fitzgeraldii, M. niphophilus | M. fitzgeraldii: α-pinene (22.7%), myrcene (24.1%), hibaene (27.0%). M. niphophilus: α-pinene (26.9%), limonene (30.5%) myrcene (20.8%) | Podocarpaceae | 2001, [226] |
| Miliusa traceyi, M. horsfieldii, M. brahei | Miliusa traceyi: α-pinene (18.7%), β-pinene (18.6%), β-caryophyllene (13.5%). M. horsfieldii: β-caryophyllene (20.2%), caryophyllene oxide (12.5%). M brahei: β-caryophyllene (12.8%), α-humulene (11.3%), bicyclogermacrene (12.9%) | Annonaceae | 2004, [227] |
| Mitrantia bilocularis Peter G.Wilson & B.Hyland | β-Caryophyllene (29%) and globulol (13%). | Myrtaceae | 2003, [164] |
| Mitrephora zippeliana Mig. | Type 1, α-pinene (13%), β-pinene (15%), caryophyllene oxide (10%), spathulenol (10%), β-caryophyllene (8%), type 2, β-caryophyllene (18%), α-humulene (7%), γ-curcumene (4%), bicyclogermacrene (4%), ar-curcumene (5%), caryophyllene oxide (3%), spathulenol (5%) | Annonaceae | 2004, [154] |
| Murraya paniculata (L.) Jack. | Small leaves variant: High yield. Big leaves variant: small yield. Both germacrene-D, E-nerolidol | Rutaceae | 1994, [228] |
| Neolitsea australiensis, N. brassii | N. australiensis: bicyclogermacrene (12–16%), guaiol (13–17%). N. brassii: bicyclogermacrene (11–15%), cubenol (6–10%), guaiol (7–10%). Northern chemotypes: germacrone (50%), bicyclogermacrene (12–35%), spathulenol (4–38%), type 2, furanogermenone (43%). Southern chemotypes: γ-eudesmol (3–30%), spathulenol (5–30%) | Lauraceae | 2002, [229] |
| Neofabricia myrtifolia, N. mjoebergii, N. sericisepala | N. mjoebergii: caryophyllene, humulene, β-pinene. N. sericisepala: over 50% α-pinene, caryophyllene. N. myrtifolia: type 1 α-pinene (>60%) caryophyllene, type 2, α-pinene, caryophyllene (up to 60%) | Myrtaceae | 1992, [230] |
| Osbornia octodonta F. Muell. | α-Pinene (35%), 1, 8-cineole (24%), α-terpineol (11%) | Myrtaceae | 1993, [231] |
| Palmeria F.Muell., spp. | Elemol, spathulenol, bicyclogermacrene, ishwarane | Monimiaceae | 2004, [232] |
| Pentaceras australe (F. Muell.) Benth. | α-Bisabolol (0.3–18%), γ-elemene (11–25%), germacrene D (13–25%), sesquicineole (4–7%) | Rutaceae | 2002, [233] |
| Phebalium distans, P. glandulosum subsp. glandulosum, P. longifolium, P. nottii, P. squamulosum subsp. squamulosum, P. squamulosum subsp. gracile, P. whitei, and P. woombye | P. distans: α-pinene (0.8–42%), bicyclogermacrene (12–22%). P. glandulosum subsp. glandulosum: dihydrotagetone, 75–95%. P. longifolium: β-caryophyllene (9–20%), bicyclogermacrene (7–23%). P. nottii: α-pinene (25–42%), guaiol (t-28%), bulnesol (nil to 34%). P. squamulosum subsp. squamulosum: α-phellandrene (12–25%), β-phellandrene (14–15%), bicyclogermacrene (2–12%), elemol/hedycaryol (12–36%). P. squamulosum subsp. gracile: geijerene (4–8%), α-pinene (44–50%), guaiol (9–11%). P. whitei: α-pinene (22–42%), limonene (1–12%), bicyclogermacrene (11–20%). P. woombye: α-pinene (10–21%), β-phellandrene (12–20%), bicyclogermacrene (11–20%), germacrene D (5–10%) | Rutaceae | 2006, [116] |
| Phyllocladus aspleniifolius (Labill.) Hook.f. | Yield 0.5–1%: α-pinene (44–55%), phyllocladene (15–28%), 8-β-hydroxyisopimarene (5–7%) | Podocarpaceae | 2003, [185] |
| Pilidiostigma glabrum Burret, P. recurvum (C.T. White) A.J. Scott, P. rhytisperma (F. Muell.) Burret, P. tetramerum L.S. Sm., P. tropicum L.S. Sm. and Pilidiostigma sp. (Mt Lewis G.P. Guymer 2024) | All species: aromadendrene, allo-aromadendrene, β-caryophyllene, α-copaene, viridiflorene, bicyclogermacrene, δ- and γ-cadinene and globulol | Myrtaceae | 1999, [234] |
| Pistacia lentiscus L. | α-Pinene, β-pinene, myrcene, limonene, thymol, carvacrol | Anacardiaceae | 1990, [235] |
| Pitaviaster haplophyllus (F. Muell.) T. G. Hartley | Germacrene D (10–28%), germacrene B (1–30%), bicyclogermacrene (2–10%), elemol (5–21%) | Rutaceae | 2002, [236] |
| Podocarpus dispermus, P. drouynianus, P. elatus, P. grayae, P. smithii, P. spinulosus | P. dispermus: isopimara-9(11),15-diene (10.3%). P. drouynianus: bicyclogermacrene (57%). P. elatus: α-pinene, β-caryophyllene, bicyclogermacrene. P. grayae: Bicyclogermacrene (27–44%), germacrene D (4–11%), spathulenol (3–11%). P. lawrencei: α-pinene (14–18%), β-caryophyllene (3–15%), spathulenol (0.8–19%), rimuene (0.2–30%), sandarocopimara-8(14), 15-diene (0.1–20%), beyerene (7–9%). P. smithii: α-copaene (4–8%), β-caryophyllene (5%), cadinene (9–11%). P. spinulosus: limonene (13–16%), bicyclogermacrene (15–18%) and viridiflorol (16–18%) | Podocarpaceae | 2004, [237] |
| Polyalthia australis, P. michaelii, P. nitidissima, P. sp. (Wyvuri B.P.Hyland RFK2632) | P. australis: β-caryophyllene (4–15%), germacrene D (1–13%), bicyclogermacrene (1–10%), caryophyllene oxide (6–10%), spathulenol (7–24%). P. michaelii: spathulenol (42.2%). P. nitidissima: type 1, α-pinene (10–12%), limonene (4–6%), (E)-β-ocimene (0.7–2%), δ-cadinene (4–8%), spathulenol (15–17%), type 2, spathulenol (22–28%), bicyclogermacrene (4–24%), β-caryophyllene (2–10%). P. sp. (Wyvuri): globulol (1–34%), spathulenol (4–6%), ledol (1–10%), germacrene B (22–39%) | Annonaceae | 2001, [238] |
| Polygonum odoratum Lour | Decanal (27.73%), dodecanal (44.05%), decanol (10.88%) | Polygonaceae | 1997, [239] |
| Pseuduvaria mulgraveana var. mulgraveana, P. mulgraveana var. glabrescens, P. hylandii, P. villosa, and P. froggattii | P. mulgraveana var. mulgraveana and P. mulgraveana var. glabrescens: elemicin (87%) and methyl eugenol (61%). P. villosa: β-caryophyllene (3–13%) and α-copaene (4–11%). P. froggattii: caryophyllene oxide (2–22%), froggatt ether (0.1–18%), spathulenol (9–18%). P. hylandii: β-caryophyllene (22–28%), α-himachalene (2–10%), α-humulene (8–9%) | Annonaceae | 2004, [240] |
| Ristantia gouldii, R. pachysperma, R. waterhousei | R. gouldii: α-pinene (39–48%), β-pinene (19–34%). R. pachysperma: (E)-nerolidol (42%). R. waterhousei: α-Pinene (53–66%) | Myrtaceae | 2003, [164] |
| Rhodamnia dumicola, R. pauciovulata, R. rubescens and R. sp. (McIlwraith Range, L.J. Webb + 9527), R. australis, R. blairiana, R. costata, R. dumicola, R. argentea, R. whiteana, R. sp. (Cape York, L. S. Smith 12538), R. glabrescens, R. sessiliflora, and R. spongiosa, R. maideniana, R. sp. (Calliope, N. Gibson 1335) | R. dumicola, R. pauciovulata, R. rubescens and R. sp. (McIlwraith Range): α-pinene. R. australis, R. blairiana, R. costata, R. dumicola, R. argentea, R. whiteana and R. sp. (Cape York): α-, β-, and γ-eudesmol, α- and β-pinene. R. glabrescens, R. argentea type 2, R. sessiliflora, R. spongiosa: β-caryophyllene, globulol, viridiflorol, spathulenol. R. maideniana, R. sp. (Calliope): unidentified oxygenated sesquiterpenes | Myrtaceae | 1997, [241] |
| Rhodomyrtus canescens C. T. White & W. D. Francis, R. effussa Guymer., R. macrocarpa Benth., R. pervagata Guymer., R. psidioides (G. Don.) Benth., R. sericea Burret., R. trineura (F. Muell.) F. Muell. ex Benth. subsp. trineura, R. trineura subsp. capensis Guymer | R. canescens: α-pinene (20–23%), β-pinene (6–10%), aromadendrene (12?17%). R. effussa: globulol (11–22%), viridiflorol (8–10%), spathulenol (5–18%). R. macrocarpa: β-caryophyllene (9–44%), aromadendrene (6–11%), globulol (8–10%). R. pervagata: α-pinene (27–35%), β-pinene (18–24%). R. psidioides: α-pinene (28–66%), limonene (1–24%). R. sericea: α-pinene (28%), β-pinene (21%), β-caryophyllene (13%). R. trineura subsp. trineura: β-caryophyllene (16–29%), caryophyllene oxide (2–12%), globulol (7–10%). R. trineura subsp. capensis: α-pinene (tr-26%), globulol (9–19%), viridiflorol (5–12%), spathulenol (4–7%) | Myrtaceae | 1997, [242] |
| Sarcomelicope simplicifolia subsp. simplicifolia (Endl.) T.G. Hartley | β-Caryophyllene (14–22%), bicyclogermacrene (10–42%), α-copaene (1–8%), γ-elemene (1–3%), α-humulene (1–4%), δ-cadinene (1–7%), δ-elemene (0.2–2%), spathulenol (1–5%) | Rutaceae | 1997, [216] |
| Sphaerantia chartacea Peter G.Wilson & B.Hyland | β-Bisabolene (t-25%), bicyclogermacrene (15–40%), globulol (6–10%) | Myrtaceae | 2003, [164] |
| Sphaerantia discolor Peter G.Wilson & B.Hyland | Bicyclogermacrene (19%), globulol (15%), viridiflorene (14%), viridiflorol (8%), ar-curcumene (9%) | Myrtaceae | 2003, [164] |
| Steganthera australiana, S. cooperorum, S. hirsute, S. laxiflora subsp. laxiflora, S. laxiflora subsp. lewisensis, and S. macooraia | S. australiana: sesquiterpenoid. S. cooperorum: aromadendrene (6–10%), viridiflorene (7–8%), globulol (7–9%), b-eudesmol (8–11%); dodecanal (2–4%), type 2, elemol (30–42%), hedycaryol (approx. 8%), α-, β- and γ-eudesmol (10–14%, 10–14%, 5–9%, respectively). S. hirsute: bicyclogermacrene (37–56%), β-caryophyllene (2–17%), germacrene D (2–22%). S. laxiflora subsp. laxiflora: viridiflorene (13–20%). S. macooraia: guaiol (2–23%), bulnesol (0.7–10%), elemol (10–22%). S. laxiflora subsp. lewisensis: viridiflorene (11.8%), 2-dodecenal (6.3%) | Monimiaceae | 2009, [189] |
| Sundacarpus amarus (Blume) C.N.Page | β-Selinene (67–77%), bicyclogermacrene (9–11%) | Podocarpaceae | 2000, [243] |
| Syncarpia glomulifera (Sm.) Nied. subsp. glomulifera, S. glomulifera subsp. glabra (Benth.) A. R. Bean, S. verecunda A. R. Bean, S. hillii F. M. Bailey | S. glomulifera (Sm.) Nied. subsp. glomulifera, S. glomulifera subsp. glabra (Benth.) A. R. Bean and S. verecunda A. R. Bean: α-pinene (30–50%), α-thujene (11–27%), aromadendrene (1–13%), globulol (3–8%). S. hillii: hillyl acetate (53–80%), hillone (6–12%), α-pinene (2–22%) | Myrtaceae | 1996, [244] |
| Syncarpia hillii F.M.Bailey | Hillyl acetate, 7-acetoxy-2,2,6,6,8,8-hexamethyl-5-oxo-5,6,7,8-tetrahydro-2H-chromen, | Myrtaceae | 1994, [245] |
| Thaleropia queenslandica (L.S.Sm.) Peter G.Wilson | α-Pinene (61–83%), spathulenol (1–4%), bicyclogermacrene (1–3%) | Myrtaceae | 1997, [246] |
| Thryptomene australis, T. kochii | T. australis: geranic acid (52%), α-pinene (22%). T. kochii: α-pinene (58–60%) | Myrtaceae | 2004, [148] |
| Thryptomene hexandra, T. parviflora, T. oligandra | T. hexandra and T. parviflora: 1,8-cineole (up to 70%), α-pinene (up to 16%). T. oligandra: γ-terpinene (32–62%), p-cymene (25–50%), α-pinene (1–20%) | Myrtaceae | 2000, [247] |
| Trachymene incisa Rudge | β-selinene, bicyclogermacrene, γ-bisabolene, α-pinene, β-caryophyllene | Araliaceae | 2021, [248] |
| Tristaniopsis collina, T. exiliflora, T. laurina, T. neriifolia | T. collina: α-pinene (trace—35–9%), myrcene (5.6–29.3%), cubenol (0–16.6%). T. exiliflora: β-caryophyllene (13.4%), δ-cadinene (16.4%). T. laurina: type 1, α-pinene (79.4%), type 2, limonene (13.0%), globulol (9.7%). T. neriifolia: α-pinene (24.4%), α-eudesmol (17.8%), β-eudesmol (17.2%) and γ-eudesmol (28.0%) | Myrtaceae | 1999, [249] |
| Uromyrtus australis A. J. Scott, U. metrosideros (F. M. Bailey), A. J. Scott, U. sp. (Tinaroo Range G. P. Guymer 2034), U. sp. (McPherson Range G. P. Guymer 2000) | U. australis: β-caryophyllene (21%), β-, α- and γ-eudesmol (13, 9 and 11% respectively). U. metrosideros: α-pinene (13–20%), β-pinene (33–42%), α-terpineol (3–7%), spathulenol (9–15%). U. sp. (Tinaroo Range): bicyclogermacrene (14–23%), globulol (9–12%), viridiflorol (5–7%) and spathulenol (3–4%). U. sp. (McPherson Range): terpinen-4-ol (8–13%), α-pinene (3–10%) | Myrtaceae | 1996, [250] |
| Uvaria rufa, U. concave | U. rufa: α-humulene (50%), benzyl benzoate U. concave: spathulenol (32%) | Annonaceae | 2004, [154] |
| Vitex limonifolia Wall | Caryophyllene (43%), caryophyllene oxide (13%), α-pinene (6%) | Lamiaceae | 1990, [251] |
| Vitex trifolia L., V. trifolia L. var. simplicifolia Cham | V. trifolia: 1,8-cineole, terpinyl acetate, sabinene, α-pinene, caryophyllene. V. trifolia var. simplicifolia: 1,8-cineole, α-pinene, terpinyl acetate, sabinene | Lamiaceae | 1991, [252] |
| Viticipremna queenslandica Munir (syn. Vitex queenslandica) | β-caryophyllene (27–34%), germacrene D (1–16%), bicyclogermacrene (9–15%), spathulenol (2–6%) | Lamiaceae | 2008, [253] |
| Waterhousea floribunda, W. hedraiophylla, W. unipunctata, W. mulgraveana | W. floribunda: α- and β-pinene (17–21%). W. hedraiophylla: spathulenol, calamenene, δ-cadinene, bicyclogermacrene. W. unipunctata: germacrene D (8.2–25.6%), bicyclogermacrene (3.2–12.3%), globulol (0.4–15.8%), viridiflorol (0.2–9.6%). W. mulgraveana: 1,3,5-trimethoxybenzene, 2,4,6-trimethoxyethylbenzene, 2,4,6-trimethoxystyrene, 2,4,6-trimethoxy-3,5-dirnethylstyrene | Myrtaceae | 2002, [254] |
| Wollemia nobilis W.G.Jones, K.D.Hill & J.M.Allen | (+)−16-Kaurene (60%), α-pinene (9%), germacrene-D (8%) | Araucariaceae | 2000, [147] |
| Welchiodendron longivalve (F. Muell.) Peter G. Wilson & J. T. Waterh | β-Trans-ocimene (17–28%), caryophyllene (39–57%), humulene (7–22%) | Myrtaceae | 1996, [214] |
| Wilkiea angustifolia, W. austroqueenslandica, W. cordata, W. huegeliana, W. longipes, W. macrophylla, W. pubescens, W. rigidifolia, W. smithii, W. sp. (McDowall Range J.G.Tracey 14552), W. sp. (Palmerston B.P.Hyland 80) and W. sp. (Russell Gorge S.J.Dansie 1909) | W. angustifolia: β-eudesmol (35%). W. austroqueenslandica: (E)- β-ocimene (5–9%), germacrene D (7–18%), bicyclogermacrene (23–26%). W. cordata: β-eudesmol (22–25%), spathulenol (11.3%). W. huegeliana: α-pinene (10–12%), viridiflorene (4–9%). W. longipes: (Z)-β-ocimene (11–15%). W. macrophylla: dodecanal (2–35%), (2)-dodecenal (0.1–2%), β-caryophyllene (14–24%). W. pubescens: sesquiterpenes. W. rigidifolia: (Z)-β-ocimene (3.4–11.5%), bicyclogermacrene (3.2–17.4%), germacrene B (0.3–14.5%). W. smithii: β-caryophyllene (10.4%), bicyclogermacrene (9.6%), caryophyllene oxide (8.9%), spathulenol (16.7%). W. sp. (McDowall Range): selina-6-en-4-ol (30.7%), germacrene D-4-ol (18.6%). W. sp. (Palmerston): spathulenol (22%). W. sp. (Russell) elemol (19.2%) | Monimiaceae | 2009, [189] |
| Xylopia maccreae (F.Muell.) L.S. Sm., Xylopia sp. (Bertiehaugh Homestead C. Dalliston CC173) | X. maccreae: bicyclogermacrene (34%), β-cubebene, β-caryophyllene, germacrene D. Xylopia sp. (Bertiehaugh Homestead): spathulenol (31%), globulol, viridiflorol, caryophyllene oxide | Annonaceae | 1998, [255] |
| Zygogynum howeanum, Z. semecarpoides var. semecarpoides, Z. queenslandianum subsp. queenslandianum, Z. queenslandianum subsp. australe | Z. howeanum: β-caryophyllene (48.0%), α-humulene (16.5%) and (Z)-β-farnesene (3.2%). Z. semecarpoides var. semecarpoides spathulenol (24.4%). Z. queenslandianum subsp. queenslandianum: bicyclogermacrene (10.4%), globulol (7.7%), viridiflorol (4.5%), spathulenol (11.5%). Z. queenslandianum subsp. austral: elemol (5.1%), bicyclogermacrene, spathulenol | Winteraceae | 1994, [256] |
6. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, N.D.; Watt, R.A.; Moffat, A.C. A near-infrared method for the assay of cineole in eucalyptus oil as an alternative to the official BP method. J. Pharm. Pharm. 2001, 53, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Sadgrove, N.J.; Jones, G.L. A contemporary introduction to essential oils: Chemistry, bioactivity and prospects for Australian agriculture. Agriculture 2015, 5, 48–102. [Google Scholar] [CrossRef] [Green Version]
- Sadgrove, N.J. Honest nutraceuticals, cosmetics, therapies, and foods (NCTFs): Standardization and safety of natural products. Crit. Rev. Food. Sci. Nutr. 2021, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Zhou, L.; Huang, Y.; Wang, Y.; Hao, X.; Wang, J. Antimicrobial activity of globulol isolated from the fruits of Eucalyptus globulus Labill. Nat. Prod. Res. 2008, 22, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Mulyaningsih, S.; Sporer, F.; Reichling, J.; Wink, M. Antibacterial activity of essential oils from Eucalyptus and of selected components against multidrug-resistant bacterial pathogens. Pharm. Biol. 2011, 49, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Mulyaningsih, S.; Sporer, F.; Zimmermann, S.; Reichling, J.; Wink, M. Synergistic properties of the terpenoids aromadendrene and 1,8-cineole from the essential oil of Eucalyptus globulus against antibiotic-susceptible and antibiotic-resistant pathogens. Phytomedicine 2010, 17, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- McLellan, R.C.; Dixon, K.; Watson, D.M. Prolific or precarious: A review of the status of Australian sandalwood (Santalum spicatum [R.Br.] A.DC., Santalaceae). Rangel. J. 2021, 43, 211–222. [Google Scholar] [CrossRef]
- Howes, M.J.; Simmonds, M.S.; Kite, G.C. Evaluation of the quality of sandalwood essential oils by gas chromatography-mass spectrometry. J. Chromatogr. A 2004, 1028, 307–312. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Klepp, J.; Legendre, S.V.A.-M.; Lyddiard, D.; Sumby, C.J.; Greatrex, B.W. Revision of the phytochemistry of Eremophila sturtii and E. mitchellii. J. Nat. Prod. 2018, 81, 405–409. [Google Scholar] [CrossRef]
- Beattie, K.; Waterman, P.G.; Forster, P.I.; Thompson, D.R.; Leach, D.N. Chemical composition and cytotoxicity of oils and eremophilanes derived from various parts of Eremophila mitchellii Benth. (Myoporaceae). Phytochemistry 2011, 71, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Southwell, I.A.; Brophy, J.J. Essential oil isolates from the Australian flora. Part 3. J. Essent. Oil Res. 2000, 12, 267–278. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Jones, G.L.; Greatrex, B.W.; Watson, K. Ethnopharmacology and Chemotaxonomy of Essential Oil Yielding Australian Plants. Ph.D. Thesis, University of New England, Armidale, Australia, 2014. [Google Scholar]
- Jones, G.L.; Sadgrove, N.J. Ethnopharmacology in Australia and Oceania. In Ethnopharmacology—A Reader; Heinrich, M., Jäger, A.K., Eds.; John Wiley & Sons: Chichester, UK, 2015. [Google Scholar]
- Sadgrove, N.J.; Jones, G.L. From Petri Dish to Patient: Bioavailability Estimation and Mechanism of Action for Antimicrobial and Immunomodulatory Natural Products. Front. Microbiol. 2019, 10, 2470. [Google Scholar] [CrossRef] [PubMed]
- Langat, M.K.; Mayowa, Y.; Sadgrove, N.J.; Danyaal, M.; Prescott, T.A.K.; Kami, T.; Schwikkard, S.; Barker, J.; Cheek, M. Multi-layered antimicrobial synergism of (E)-caryophyllene with alkaloids and 13S-hydroxy-9Z,11E,15E-octadecatrienoic acid from the leaves of Vepris gossweileri (I. Verd.) Mziray. Nat. Prod. Res. 2021; (in press). [Google Scholar] [CrossRef]
- Sadgrove, N.J. Southern Africa as a ‘cradle of incense’ in wider African aromatherapy. Sci. Afr. 2020, 9, e00502. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Padilla-Gonzalez, G.F.; Leuner, O.; Melnikovova, I.; Fernandez-Cusimamani, E. Pharmacology of Natural Volatiles and Essential Oils in Food, Therapy, and Disease Prophylaxis. Front Pharm. 2021, 12, 740302. [Google Scholar] [CrossRef]
- Sadgrove, N.; Jones, G.L.; Greatrex, B.W. Isolation and characterisation of (−)-genifuranal: The principal antimicrobial component in traditional smoking applications of Eremophila longifolia (Scrophulariaceae) by Australian Aboriginal peoples. J. Ethnopharmacol. 2014, 154, 758–766. [Google Scholar] [CrossRef]
- Sadgrove, N.; Jones, G.L. A possible role of partially pyrolysed essential oils in Australian Aboriginal traditional ceremonial and medicinal smoking applications of Eremophila longifolia (R. Br.) F. Muell (Scrophulariaceae). J. Ethnopharmacol. 2013, 147, 638–644. [Google Scholar] [CrossRef]
- Fang, J.-M.; Chen, Y.-C.; Wang, B.-W.; Cheng, Y.-S. Terpenes from the heartwood of Juniperus chinensis. Phytochemistry 1996, 41, 1361–1365. [Google Scholar] [CrossRef]
- Lee, C.-H.; Park, J.-M.; Song, H.-Y.; Jeong, E.-Y.; Lee, H.-S. Acaricidal activities of major constituents of essential oil of Juniperus chinensis leaves against house dust and stored food mites. J. Food Prot. 2009, 72, 1686–1691. [Google Scholar] [CrossRef]
- Tunalier, Z.; Kirimer, N.; Baser, K.H.C. A potential new source of cedarwood oil: Juniperus foetidissima Willd. J. Essent. Oil Res. 2004, 16, 233–235. [Google Scholar] [CrossRef]
- Soković, M.; Ristic, M.; Grubisic, D. Chemical Composition and Antifungal Activity of the Essential Oil from Juniperus excelsa Berries. Pharm. Biol. 2004, 42, 328–331. [Google Scholar] [CrossRef]
- Fang, J.-M.; Sou, Y.-C.; Chiu, Y.-H.; Cheng, Y.-S. Diterpenes from the bark of Juniperus chinensis. Phytochemistry 1993, 34, 1581–1584. [Google Scholar]
- Oda, J.; Ando, N.; Nakajima, Y.; Inouye, Y. Studies on Insecticidal Constituents of Juniperus recurva Buch. Agric. Biol. Chem. 1977, 41, 201–204. [Google Scholar] [CrossRef]
- Richmond, G.S. A review of the use of Eremophila (Myoporaceae) by Australian Aborigines. J. Adel. Bot. Gard. 1993, 15, 101–107. [Google Scholar]
- Maiden, J.H. Forestry Handbook. Part 2. Some of the Principal Commercial Trees of New South Wales; William Applegate Gullick; Government Printer: Sydney, Australia, 1917. [Google Scholar]
- Maiden, J.H. The Useful Native Plants of Australia; Published by Compendium in 1975; Alexander Bros Vic: Melbourne, Australia, 1889. [Google Scholar]
- Cribb, A.B.; Cribb, J.W. Useful Wild Plants in Australia; William Collins Pty. Ltd.: Sydney, Australia, 1981. [Google Scholar]
- Beattie, K.D. Phytochemical Studies and Bioactivity of Centipeda and Eremophila Species. Ph.D. Thesis, Southern Cross University, Lismore, Australia, 2009. [Google Scholar]
- Govaerts, R.; Nic Lughadha, E.; Black, N.; Turner, R.; Paton, A. The World Checklist of Vascular Plants, a continuously updated resource for exploring global plant diversity. Sci. Data 2021, 8, 215. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Senbill, H.; Van Wyk, B.-E.; Greatrex, B.W. New labdanes with antimicrobial and acaricidal activity: Terpenes of Callitris and Widdringtonia (Cupressaceae). Antibiotics 2020, 9, 173. [Google Scholar] [CrossRef] [Green Version]
- Hay, R. Demodex and skin infection: Fact or fiction. Curr. Opin. Infect. Dis. 2010, 23, 103–105. [Google Scholar] [CrossRef]
- Kiran, S.R.; Reddy, A.S.; Devi, P.S.; Reddy, K.J. Insecticidal, antifeedant and oviposition deterrent effects of the essential oil and individual compounds from leaves of Chloroxylon swietenia DC. Pest. Manag. Sci. 2006, 62, 1116–1121. [Google Scholar] [CrossRef]
- Ali, J.G.; Alborn, H.T.; Campos-Herrera, R.; Kaplan, F.; Duncan, L.W.; Rodriguez-Saona, C.; Koppenhofer, A.M.; Stelinski, L.L. Subterranean, herbivore-induced plant volatile increases biological control activity of multiple beneficial nematode species in distinct habitats. PLoS ONE 2012, 7, e38146. [Google Scholar] [CrossRef] [Green Version]
- Winkelman, W.J. Aromatherapy, botanicals, and essential oils in acne. Clin. Derm. 2018, 36, 299–305. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Simmonds, M.S.J. Topical and nutricosmetic products for healthy hair and dermal antiaging using “dual-acting” (2 for 1) plant-based peptides, hormones, and cannabinoids. FASEB Bioadv. 2021, 3, 601–610. [Google Scholar] [CrossRef]
- Sadgrove, N.J. The new paradigm for androgenetic alopecia and plant-based folk remedies: 5α-reductase inhibition, reversal of secondary microinflammation and improving insulin resistance. J. Ethnopharmacol. 2018, 227, 206–236. [Google Scholar] [CrossRef] [PubMed]
- Hulley, I.M.; Van Vuuren, S.F.; Sadgrove, N.J.; Van Wyk, B.-E. Antimicrobial activity of Elytropappus rhinocerotis (Asteraceae) against micro-organisms associated with foot odour and skin ailments. J. Ethnopharmacol. 2019, 228, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Sadgrove, N.; Mijajlovic, S.; Tucker, D.J.; Watson, K.; Jones, G.L. Characterization and bioactivity of essential oils from novel chemotypes of Eremophila longifolia (F. Muell) (Myoporaceae): A highly valued traditional Australian medicine. Flavour Fragr. J. 2011, 26, 341–350. [Google Scholar] [CrossRef]
- Carson, C.F.; Hammer, K.A.; Riley, T.V. Melaleuca alternifolia (Tea Tree) oil: A review of antimicrobial and other medicinal properties. Clin. Microbiol. Rev. 2006, 19, 50–62. [Google Scholar] [CrossRef] [Green Version]
- Bejar, E. Tea Tree Oil (Melaleuca alternifolia and M. linariifolia). Available online: www.botanicaladulterants.org (accessed on 10 May 2021).
- Wilkinson, J.M.; Cavanagh, H.M.A. Antibacterial activity of essential oils from Australian native plants. Phytother. Res. 2005, 19, 643–646. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Padilla-González, G.F.; Telford, I.R.H.; Greatrex, B.W.; Jones, G.L.; Andrew, R.; Bruhl, J.J.; Langat, M.K.; Melnikovova, I.; Fernandez-Cusimamani, E. Prostanthera (Lamiaceae) as a ‘Cradle of Incense’: Chemophenetics of Rare Essential Oils from Both New and Forgotten Australian ‘Mint Bush’ Species. Plants 2020, 9, 1570. [Google Scholar] [CrossRef]
- Collins, T.L.; Jones, G.L.; Sadgrove, N. Volatiles from the rare Australian desert plant Prostanthera centralis B.J.Conn (Lamiaceae): Chemical composition and antimicrobial activity. Agriculture 2014, 4, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Sadgrove, N.J.; Greatrex, B.W.; Jones, G.L. α-Cyclodextrin encapsulation enhances antimicrobial activity of cineole-rich essential oils from Australian species of Prostanthera (Lamiaceae). Nat. Volatiles Essent. Oils 2015, 2, 30–38. [Google Scholar]
- Sadgrove, N.; Jones, G.L. Antimicrobial activity of essential oils and solvent extracts from Zieria species (Rutaceae). Nat. Prod. Commun. 2013, 8, 741–745. [Google Scholar] [CrossRef] [Green Version]
- Tavares, A.C.; Goncalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Lopes, M.C.; Canhoto, J.; Salgueiro, L.R. Essential oil of Daucus carota subsp. halophilus: Composition, antifungal activity and cytotoxicity. J. Ethnopharmacol. 2008, 119, 129–134. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Gonçalves-Martins, M.; Jones, G.L. Chemogeography and antimicrobial activity of essential oils from Geijera parviflora and Geijera salicifolia (Rutaceae): Two traditional Australian medicinal plants. Phytochemistry 2014, 104, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of the Australian Species of Flindersia (Rutaceae). J. Essent. Oil Res. 2005, 17, 388–395. [Google Scholar] [CrossRef]
- Tonzibo, Z.F.; Wognin, E.; Chalchat, J.C.; N’Guessan, Y.T. Chemical Investigation of Chromolaena odorata L. King Robinson from Ivory Coast. J. Essent. Oil Bear. Plants 2007, 10, 94–100. [Google Scholar] [CrossRef]
- Dellar, J.E.; Cole, M.D.; Gray, A.I.; Gibbons, S.; Waterman, P.G. Antimicrobial sesquiterpenes from Prostanthera aff. melissifolia and P. rotundifolia. Phytochemistry 1994, 36, 957–960. [Google Scholar] [CrossRef]
- Sakkas, H.; Economou, V.; Gousia, P.; Bozidis, P.; Sakkas, V.A.; Petsios, S.; Mpekoulis, G.; Ilia, A.; Papadopoulou, C. Antibacterial Efficacy of Commercially Available Essential Oils Tested Against Drug-Resistant Gram-Positive Pathogens. Appl. Sci. 2018, 8, 2201. [Google Scholar] [CrossRef] [Green Version]
- Tighe, S.; Gao, Y.-Y.; Tseng, S.C.G. Terpinen-4-ol is the Most Active Ingredient of Tea Tree Oil to Kill Demodex Mites. Transl. Vis. Sci. Technol. 2013, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.-J.; Jin, L.-Y.; Yu, Q.; Liu, H.; Khan, A.; Yan, H.; Hao, X.-J.; An, L.-K.; Liu, H.-Y. Eucalypglobulusals A–J, Formyl-Phloroglucinol–Terpene Meroterpenoids from Eucalyptus globulus Fruits. J. Nat. Prod. 2018, 81, 2638–2646. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Oliveira, T.B.; Khumalo, G.P.; Vuuren, S.F.v.; van Wyk, B.-E. Antimicrobial Isoflavones and Derivatives from Erythrina (Fabaceae): Structure Activity Perspective (Sar & Qsar) on Experimental and Mined Values Against Staphylococcus Aureus. Antibiotics 2020, 9, 223. [Google Scholar] [CrossRef]
- Bharate, S.B.; Bhutani, K.K.; Khan, S.I.; Tekwani, B.L.; Jacob, M.R.; Khan, I.A.; Singh, I.P. Biomimetic synthesis, antimicrobial, antileishmanial and antimalarial activities of euglobals and their analogues. Bioorg. Med. Chem. 2006, 14, 1750–1760. [Google Scholar] [CrossRef]
- Usuki, Y.; Deguchi, T.; Iio, H. A New Concise Synthesis of (+)-Ipomeamarone, (−)-Ngaione, and Their Stereoisomers. Chem. Lett. 2014, 43, 1882–1884. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Padilla-Gonzalez, G.F.; Green, A.; Langat, M.K.; Mas-Claret, E.; Lyddiard, D.; Klepp, J.; Legendre, S.V.A.; Greatrex, B.W.; Jones, G.L.; et al. The Diversity of Volatile Compounds in Australia’s Semi-Desert Genus Eremophila (Scrophulariaceae). Plants 2021, 10, 785. [Google Scholar] [CrossRef] [PubMed]
- Sadgrove, N.J.; Jones, G.L. Cytogeography of essential oil chemotypes of Eremophila longifolia F. Muell (Schrophulariaceae). Phytochemistry 2014, 105, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Homburger, F.; Kelley, T.; Friedler, G.; Rusfield, A.B. Toxic and Possible Carcinogenic Effects of 4-Allyl-1,2-methylene-dioxybenzene (Safrole) in Rats on Deficient Diets. Med. Experimentalis. Int. J. Exp. Med. 1961, 4, 1–11. [Google Scholar]
- Miller, E.C.; Swanson, A.B.; Phillips, D.H.; Fletcher, T.L.; Liem, A.; Miller, J.A. Structure-Activity Studies of the Carcinogenicities in the Mouse and Rat of Some Naturally Occuring Synthetic Alkenylbenzene Derivatives Related to Safrole and Estragole. Cancer Res. 1983, 43, 1124–1134. [Google Scholar]
- Kalbhen, D.; Abbo, A. Nutmeg as a narcotic. A contribution to the chemistry of pharmacology of nutmeg (Myristica fragrans). Angew. Chem. Int. 1971, 10, 370–374. [Google Scholar] [CrossRef]
- Beyer, J.; Ehlers, D.; Maurer, H.H. Abuse of Nutmeg (Myristica fragrans Houtt.): Studies on the metabolism and the toxicologic detection of its ingredients elemicin, myristicin, and safrole in rat and human urine using gas chromagrography/mass spectrometery. Ther. Drug Monit. 2006, 28, 568–575. [Google Scholar] [CrossRef]
- Yonghua, D.; Chuan, W.; Lijun, Z.; Ping, Z.; Qin, W.; Zhongqiong, Y.; Qinjiu, J.; Jihong, J. Effects of sub-chronic intoxication of 1,8-cineole on blood biochemical indexes in mice. Anim. Husb. Feed Sci. 2015, 7, 167–170. [Google Scholar]
- Frosch, P.J.; Johansen, J.D.; Menne, T.; Pirker, C.; Rastogi, S.C.; Andersen, K.E.; Bruze, M.; Goossens, A.; Lepoittevin, J.P.; White, I.R. Further important sensitizers in patients sensitive to fragrances. Contact Dermat. 2002, 47, 279–287. [Google Scholar] [CrossRef]
- Jacobs, M.R.; Hornfeldt, C.S. Melaleuca oil poisoning. J. Toxicol. Clin. Toxicol. 1994, 32, 461–464. [Google Scholar] [CrossRef]
- Zidorn, C. Plant chemophenetics—A new term for plant chemosystematics/plant chemotaxonomy in the macro-molecular era. Phytochemistry 2019, 163, 147–148. [Google Scholar] [CrossRef]
- Ramos, Y.J.; da Costa-Oliveira, C.; Candido-Fonseca, I.; de Queiroz, G.A.; Guimarães, E.F.; Defaveri, A.C.; Sadgrove, N.J.; Moreira, D.d.L. Advanced Chemophenetic Analysis of Essential Oil from Leaves of Piper gaudichaudianum Kunth (Piperaceae) Using a New Reduction-Oxidation Index to Explore Seasonal and Circadian Rhythms. Plants 2021, 10, 2116. [Google Scholar] [CrossRef] [PubMed]
- Della, E.W.; Jefferies, P.R. The Chemistry of Eremophila Species. 111. The Essential oil of Eremophila longifolia F. Muell. Aust. J. Chem. 1961, 14, 663–664. [Google Scholar] [CrossRef] [Green Version]
- Chinnock, R. Eremophila and Allied Genera. A Monograph of the Myoporaceae; Rosenberg Publishing: Kenthurst, Australia, 2007. [Google Scholar]
- Sadgrove, N.J.; Telford, I.R.H.; Greatrex, B.W.; Jones, G.L. Composition and antimicrobial activity of essential oils from the Phebalium squamulosum species complex (Rutaceae) in New South Wales, Australia. Phytochemistry 2014, 97, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Sadgrove, N.; Telford, I.R.H.; Greatrex, B.W.; Dowell, A.; Jones, G.L. Dihydrotagetone, an unusual fruity ketone, is found in enantiopure and enantioenriched forms in additional Australian native taxa of Phebalium (Rutaceae: Boronieae). Nat. Prod. Commun. 2013, 8, 737–740. [Google Scholar] [CrossRef] [Green Version]
- Lassak, E.V.; Southwell, I.A. Occurrence of some unusual compounds in the leaf oils of Eriostemon obovalis and Phebalium glandulosum subsp. glandulosum. Aust. J. Chem. 1974, 27, 2703–2705. [Google Scholar] [CrossRef]
- Telford, I.R.H.; Bruhl, J.J. Phebalium verrucosum (Rutaceae: Boronieae), new status for a taxon excluded from P. squamulosum on morphological and phytochemical evidence. Telopea 2014, 16, 127–132. [Google Scholar] [CrossRef]
- Sadgrove, N.J. Comparing essential oils from Australia’s ‘Victorian Christmas Bush’ (Prostanthera lasianthos Labill., Lamiaceae) to closely allied new species: Phenotypic plasticity and taxonomic variability. Phytochemistry 2020, 176, 112403. [Google Scholar] [CrossRef]
- Conn, B.J.; Henwood, M.J.; Proft, K.M.; Scott, J.A.; Wilson, T.C.; Howes, R.S. An integrative taxonomic approach resolves the Prostanthera lasianthos (Lamiaceae) species complex. Aust. Syst. Bot. 2021, 34, 438–476. [Google Scholar] [CrossRef]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Collins, T.L.; Andrew, R.L.; Bruhl, J.J. Morphological, phytochemical and molecular analyses define species limits in Eucalyptus magnificata (Myrtaceae) and lead to the discovery of a new rare species. Aust. Syst. Bot. 2019, 32, 12–28. [Google Scholar] [CrossRef]
- Collins, T.L.; Andrew, R.L.; Greatrex, B.W.; Bruhl, J.J. Reliable analysis of volatile compounds from small samples of Eucalyptus magnificata (Myrtaceae). Aust. Syst. Bot. 2018, 31, 232–240. [Google Scholar] [CrossRef]
- Toyota, M.; Koyama, H.; Mizutani, M.; Asakawa, Y. (−)-ent-spathulenol isolated from liverworts is an artefact. Phytochemistry 1996, 41, 1347–1350. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Telford, I.R.H.; Padilla-González, G.F.; Greatrex, B.W.; Bruhl, J.J. GC–MS ‘chemophenetics’ on Australian pink-flowered Phebalium (Rutaceae) using herbarium leaf material demonstrates phenetic agreement with putative new species. Phytochem. Lett. 2020, 38, 112–120. [Google Scholar] [CrossRef]
- Akhtar, M.A.; Raju, R.; Beattie, K.D.; Bodkin, F.; Munch, G. Medicinal Plants of the Australian Aboriginal Dharawal People Exhibiting Anti-Inflammatory Activity. Evid.-Based Complement. Altern. Med. 2016, 2016, 2935403. [Google Scholar] [CrossRef] [PubMed]
- Lassak, E.V.; McCarthy, T. Australian Medicinal Plants; Methuen Australia Pty. Ltd.: North Rhyde, Australia, 2011. [Google Scholar]
- Sadgrove, N.J.; Lyddiard, D.; Collins, T.L.; Greatrex, B.W.; Jones, G.L. Genifuranal and other derivatives: Smoking desert plants. Acta Hortic. 2016, 1125, 181–188. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Jones, G.L. Reviewing the importance of aromatic medicinal plants in the traditional pharmacopoeia of Australian Aboriginal people. Acta Hortic. 2014, 1125, 297–302. [Google Scholar] [CrossRef]
- Nsangou, M.F.; Happi, E.N.; Fannang, S.V.; Atangana, A.F.; Waffo, A.F.K.; Wansi, J.D.; Isyaka, S.M.; Sadgrove, N.J.; Sewald, N.; Langat, M.K. Chemical Composition and Synergistic Antimicrobial Effects of a Vegetatively Propagated Cameroonian Lemon, Citrus x limon (L.) Osbeck. ACS Food Sci. Technol. 2021, 1, 354–361. [Google Scholar] [CrossRef]
- Sadgrove, N.; Hitchock, M.; Watson, K.; Jones, G.L. Chemical and biological characterization of novel essential oils from Eremophila bignoniiflora (F. Muell) (Myoporaceae): A traditional Aboriginal Australian bush medicine. Phytother. Res. 2013, 27, 1508–1516. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Jones, G.L. Phytochemical variability of Pittosporum angustifolium Lodd. (Pittosporaceae): A traditional and contemporary Aboriginal Australian medicine. Acta Hortic. 2014, 1125, 303–308. [Google Scholar] [CrossRef]
- Cribb, A.B.; Cribb, J.W. Wild Medicine in Australia; William Collins, Pty. Ltd.: Sydney, Australia, 1981. [Google Scholar]
- Baker, R.T.; Smith, H.G. On a new species of Prostanthera and its essential oil. J. Proc. R. Soc. NSW 1912, 46, 103–110. [Google Scholar]
- Conn, B.J. A taxonomic revision of Prostanthera Labill. section Klanderia (F.v.Muell.) Benth. (Labiatae). J. Adel. Bot. Gard. 1984, 6, 207–348. [Google Scholar]
- Southwell, I.A.; Tucker, D.J. cis-Dihydroagarofuran from Prostanthera sp. aff. ovalifolia. Phytochemistry 1993, 22, 857–862. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Collins, T.L.; Legendre, S.V.A.-M.; Klepp, J.; Jones, G.L.; Greatrex, B.W. The Iridoid Myodesert-1-ene and Elemol/Eudesmol are found in Distinct Chemotypes of the Australian Aboriginal Medicinal Plant Eremophila dalyana(Scrophulariaceae). Nat. Prod. Commun. 2016, 11, 1211–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penfold, A.R. Natural chemical resources of Australian plant products. Part II. J. Chem. Educ. 1932, 9, 429. [Google Scholar] [CrossRef]
- Penfold, A.R. The natural chemical resources of Australia Plant Products. Part 1. J. Chem. Educ. 1929, 6, 1195–1205. [Google Scholar] [CrossRef]
- Penfold, A.R.; Willis, J.L. The essential oil industry of Australia. Econ. Bot. 1954, 8, 316–336. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Forster, P.I. Leaf Oils of the Genus Backhousia (Myrtaceae). J. Essent. Oil Res. 1995, 7, 237–254. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Van Wyk, B.-E. Major volatile compounds in the essential oil of the aromatic culinary herb Pelargonium crispum (Geraniaceae). Nat. Volatiles Essent. OIls 2018, 5, 23–28. [Google Scholar]
- Doran, J.C.; Brophy, J.J.; Lassak, E.V. Backhousia citriodora F. Muell.—Rediscovery and chemical characterization of the L-citronellal form and aspects of its breeding system. Flavour Fragr. J. 2001, 16, 325–328. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Bean, A.R.; Forster, P.I.; Lepschi, B.J. Leaf essential oils of the genus Leptospermum (Myrtaceae) in eastern Australia. Part 5. Leptospermum continentale and allies. Flavour Fragr. J. 1999, 14, 98–104. [Google Scholar] [CrossRef]
- Diatloff, E. Effects of applied nitrogen fertiliser on the chemical composition of the essential oil of three Leptospermum spp. Aust. J. Exp. Agric. 1990, 30, 681–685. [Google Scholar] [CrossRef]
- Caputo, L.; Smeriglio, A.; Trombetta, D.; Cornara, L.; Trevena, G.; Valussi, M.; Fratianni, F.; De Feo, V.; Nazzaro, F. Chemical Composition and Biological Activities of the Essential Oils of Leptospermum petersonii and Eucalyptus gunnii. Front. Microbiol. 2020, 11, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, R.S.; Padalia, R.C.; Chauhan, A.; Singh, A.; Yadav, A.K. Volatile constituents of essential oil and rose water of damask rose (Rosa damascena Mill.) cultivars from North Indian hills. Nat. Prod. Res. 2011, 25, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Reichling, J.; Koch, C.; Stahl-Biskup, E.; Sojka, C.; Schnitzler, P. Virucidal activity of a beta-triketone-rich essential oil of Leptospermum scoparium (manuka oil) against HSV-1 and HSV-2 in cell culture. Planta Med. 2005, 71, 1123–1127. [Google Scholar] [CrossRef]
- Killeen, D.P.; van Klink, J.W.; Smallfield, B.M.; Gordon, K.C.; Perry, N.B. Herbicidal beta-triketones are compartmentalized in leaves of Leptospermum species: Localization by Raman microscopy and rapid screening. New Phytol. 2015, 205, 339–349. [Google Scholar] [CrossRef]
- Hellyer, R.O.; Pinhey, J.T. The structure of grandiflorone, a new β-triketone. J. Chem. Soc. C Org. 1966, 1496–1498. [Google Scholar] [CrossRef]
- Thomas, J.; Narkowicz, C.K.; Jacobson, G.A.; Davies, N.W. An examination of the essential oils of Tasmanian Kunzea ambigua, other Kunzea spp. and commercial Kunzea oil. J. Essent. Oil Res. 2010, 22, 381–385. [Google Scholar] [CrossRef]
- Penfold, A.R. The essential oil of Eriostemon myoporoides (De Candolle). J. Proc. R. Soc. NSW 1925, 59, 206–211. [Google Scholar]
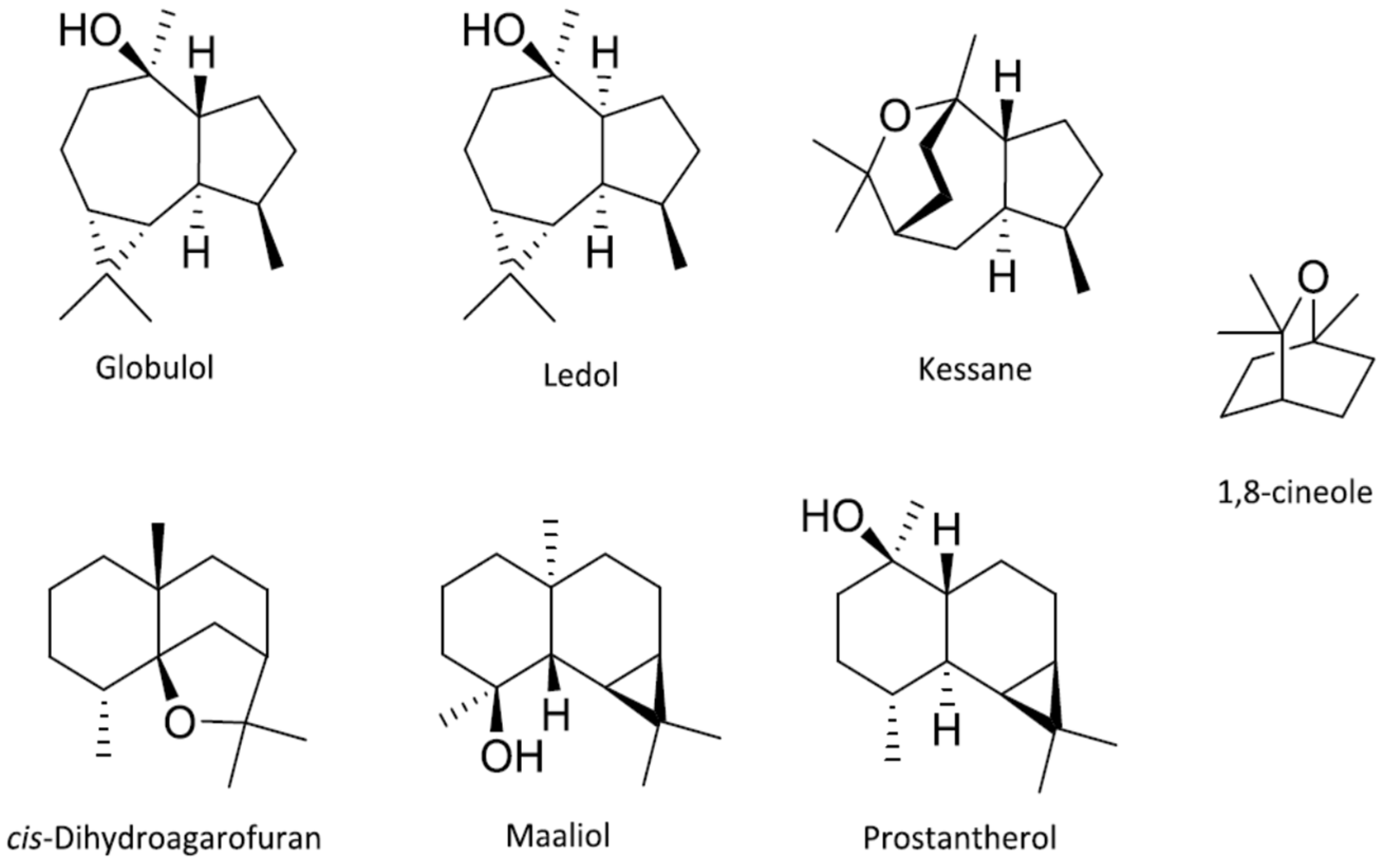
- Hellyer, R.O. Occurence of maaliol, elemol, and globulol in some Australian essential oils. Aust. J. Chem. 1962, 15, 157. [Google Scholar] [CrossRef] [Green Version]
- Curkic, A. Phytochemistry and Pharmacology of Volatile Components of Calytrix Exstipulata & Cymbopogon Bombycinus; A 449, Matrikelnummer: 0505534; Universitat Wien: Vienna, Austria, 2012. [Google Scholar]
- Doimo, L.; Bartley, J.P.; Michael, G.D. Cassinia quinquefaria R. Br. Flower and Leaf Essential Oils. J. Essent. Oil Res. 2000, 12, 702–704. [Google Scholar] [CrossRef]
- Doimo, L.; Bartley, J.P.; Michael, G.D. Cassinia laevis R. Br. Flower and Leaf Essential Oils. J. Essent. Oil Res. 2001, 13, 78–79. [Google Scholar] [CrossRef]
- Boland, D.J.; Brophy, J.J.; House, A.P.N. Eucalyptus leaf Oils: Use, Chemistry, Distillation and Marketing; Inkata Press—Commonwealth Scientific and Industrial Research Organization: Sydney, Australia, 1991. [Google Scholar]
- Kim, J.-K.; Kang, C.-S.; Lee, J.-K.; Kim, Y.-R.; Han, H.-Y.; Yun, H.K. Evaluation of Repellency Effect of Two Natural Aroma Mosquito Repellent Compounds, Citronella and Citronellal. Entomol. Res. 2005, 35, 117–120. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Leaf Essential Oils of the Queensland Species of Phebalium (Rutaceae: Boronieae). J. Essent. Oil Res. 2006, 18, 386–391. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. What is the Smell of the “Fruit Salad Plant”?: The Leaf Oil of Leionema ambiens (Rutaceae). J. Essent. Oil Res. 2006, 18, 131–133. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Jones, G.L. Medicinal compounds, chemically and biologically characterised from extracts of Australian Callitris endlicheri and C. glaucophylla (Cupressaceae): Used traditionally in Aboriginal and colonial pharmacopoeia. J. Ethnopharmacol. 2014, 153, 872–883. [Google Scholar]
- Low, T. Bush Medicine: A pharmacopoeia of Natural Remedies; Greenhouse Publications Pty Ltd.: Richmond, Australia, 1990. [Google Scholar]
- Latz, P. Bushfires and Bushtucker: Aboriginal plant Use in Central Australia; IAD Press: Alice Springs, Australia, 2004. [Google Scholar]
- Low, T. Bush Tucker: Australia’s Wild Food Harvest; Angus and Robertson Publishers: Sydney, Australia, 1989. [Google Scholar]
- Metcalfe, C.R. The Structure of Some Sandalwoods and Their Substitutes and of Some Other Little Known Scented Woods. Bull. Misc. Inf. R. Bot. Gard. Kew 1935, 1935, 165–195. Available online: https://www.jstor.org/stable/pdf/4107533.pdf (accessed on 7 December 2021). [CrossRef]
- RBGK. World Checklist of Vascular Plants. Available online: https://wcvp.science.kew.org/ (accessed on 13 January 2022).
- Braun, N.A.; Sim, S.; Kohlenberg, B.; Lawrence, B.M. Hawaiian Sandalwood: Oil Composition of Santalum paniculatum and Comparison with Other Sandal Species. Nat. Prod. Commun. 2014, 9, 1365–1368. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Chen, Y.; Shang, L.; Li, G. Assessment of essential oils from five Santalum species using ATR-fourier transform mid-infrared spectroscopy and GC-MS combined with chemometric analysis. J. Essent. Oil Res. 2020, 32, 150–157. [Google Scholar] [CrossRef]
- Penfold, A.R.; Morrison, F.R. Some Australian Essential Oils in Insecticides and Repellents; Fisher, Knight and Company: St Albans, UK, 1952. [Google Scholar]
- Cook, E.; Bird, T.; Peterson, M.; Barbetti, M.; Buckley, B.; D’Arrigo, R.; Francey, R.; Tans, P. Climatic change in tasmania inferred from a 1089-year tree-ring chronology of huon pine. Science 1991, 253, 1266–1268. [Google Scholar] [CrossRef]
- Drew, D.M.; Allen, K.; Downes, G.M.; Evans, R.; Battaglia, M.; Baker, P. Wood properties in a long-lived conifer reveal strong climate signals where ring-width series do not. Tree Physiol. 2013, 33, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Sadgrove, N.; Jones, G.L. Chemical and biological characterisation of solvent extracts and essential oils from leaves and fruit of two Australian species of Pittosporum (Pittosporaceae) used in aboriginal medicinal practice. J. Ethnopharmacol. 2013, 145, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Southwell, I.A. Methoxystyrenes from the genus Zieria. Phytochemistry 1981, 20, 1448–1450. [Google Scholar] [CrossRef]
- Flynn, T.M.; Southwell, I.A. Essential oil constituents of the genus Zieria. Phytochemistry 1987, 26, 1673–1686. [Google Scholar] [CrossRef]
- Southwell, I.A.; Armstrong, J.A. Chemical variation within the genus Zieria. Phytochemistry 1987, 26, 1687–1692. [Google Scholar] [CrossRef]
- Johnson, J.B.; Batley, R.; Manson, D.; White, S.; Naiker, M. Volatile compounds, phenolic acid profiles and phytochemical content of five Australian finger lime (Citrus australasica) cultivars. LWT 2022, 154, 112640. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of the Australian Species of Citrus (Rutaceae). J. Essent. Oil Res. 2001, 13, 264–268. [Google Scholar] [CrossRef]
- Southwell, I.A.; Brophy, J.J. Differentiation within the Australian Tasmannia by essential oil comparison. Phytochemistry 1992, 31, 3073–3081. [Google Scholar] [CrossRef]
- Cock, I.E. The phytochemistry and chemotherapeutic potential of Tasmannia lanceolata (Tasmanian pepper): A review. Pharmacogn. Commun. 2013, 3, 13–25. [Google Scholar]
- Khumalo, G.P.; Sadgrove, N.J.; Van Vuuren, S.F.; Van Wyk, B.-E. Antimicrobial activity of volatile and non-volatile isolated compounds and extracts from the bark and leaves of Warburgia salutaris (Canellaceae) against skin and respiratory pathogens. S. Afr. J. Bot. 2019, 122, 547–550. [Google Scholar] [CrossRef]
- Sultana, R.; Hossain, R.; Adhikari, A.; Ali, Z.; Yousuf, S.; Choudhary, M.I.; Ali, M.Y.; Zaman, M.S. Drimane-type sesquiterpenes from Polygonum hydropiper. Planta Med. 2011, 77, 1848–1851. [Google Scholar] [CrossRef]
- Lowe, R.F.; Russell, M.F.; Southwell, I.A.; Robinson, C.J.; Day, J. Composition of an Essential Oil from Agonis fragrans J.R. Wheeler et NGMarchant. J. Essent. Oil Res. 2007, 19, 342–344. [Google Scholar] [CrossRef]
- Shellie, R.; Mondello, L.; Dugo, G.; Marriott, P. Enantioselective gas chromatographic analysis of monoterpenes in essential oils of the family Myrtaceae. Flavour Fragr. J. 2004, 19, 582–585. [Google Scholar] [CrossRef]
- Brophy, J.J.; Craven, L.A.; Doran, J.C. Melaleucas: Their Botany, Essential Oils and Uses; ACIAR Monograph No. 156; Australian Centre for International Agricultural Research: Canberra, Australia, 2013. [Google Scholar]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Essential Oil of Acmenosperma claviflorum (Myrtaceae). J. Essent. Oil Res. 1999, 11, 162–164. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I.; Southwell, I.A. Leaf Oils of the Genus Acradenia (Rutaceae). J. Essent. Oil Res. 2001, 13, 136–139. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Leaf Essential Oils of the Australian Species of Acronychia (Rutaceae). J. Essent. Oil Res. 2004, 16, 597–607. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J. Essential Oil of the Leaves and Flowers of Actinodium cunninghamii Schauer (Myrtaceae). J. Essent. Oil Res. 1994, 6, 639–640. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; O’Sullivan, W. Chemistry of the Australian gymnosperms: Part VII. The leaf oils of the genus Actinostrobus. Biochem. Syst. Ecol. 2004, 32, 867–873. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Wu, M.Z.; Fookes, C.J.R.; Forster, P.I. The steam volatile oil of Wollemia nobilis and its comparison with other members of the Araucariaceae (Agathis and Araucaria). Biochem. Syst. Ecol. 2000, 28, 563–578. [Google Scholar] [CrossRef]
- Lassak, E.V.; Brophy, J.J. Steam volatile leaf oils of some Western Australian species of the family Myrtaceae. Flavour Fragr. J. 2004, 19, 12–16. [Google Scholar] [CrossRef]
- Brophy, J.J.; Boland, D.J. The leaf essential oil of Allosyncarpia ternata S. T. Blake. Flavour Fragr. J. 1992, 7, 117–119. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Craven, L.A. The Leaf Essential Oil of Angasomyrtus salina Trudgen & Keighery. J. Essent. Oil Res. 1994, 6, 69–71. [Google Scholar] [CrossRef]
- Dunlop, P.J.; Bignell, C.M.; Brooker, M.I.H.; Brophy, J.J.; Brynn Hibbert, D. Use of gas chromatograms of essential leaf oils to compare eight taxa of genus Angophora (Myrtaceae): Possible relationships to the genus Eucalyptus. Biochem. Syst. Ecol. 1999, 27, 815–830. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Variation in Archirhodomyrtus beckleri (F. Muell.) A.J. Scott (Myrtaceae): Evidence from Volatile Oils. Flavour Fragr. J. 1996, 11, 11–14. [Google Scholar] [CrossRef]
- Boland, D.J.; Brophy, J.J.; Goldsack, R.J. The leaf essential oil of Arillastrum gummiferum (Brongriart & Gris) Pancher ex Baillon. Flavour Fragr. J. 1994, 9, 47–49. [Google Scholar] [CrossRef]
- Brophy, J.; Goldsack, R.; Forster, P. Essential Oils from the Leaves of Some Queensland Annonaceae. J. Essent. Oil Res. 2004, 16, 95–100. [Google Scholar] [CrossRef]
- Brophy, J.J.; Clarkson, J.R.; Craven, L.A.; Forresters, R.I. The essential oils of Australian Members of the Genus Asteromyrtus (Myrtaceae). Biochem. Syst. Ecol. 1994, 22, 409–417. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Leaf Essential Oil of Austrobaileya scandens C. White. J. Essent. Oil Res. 1994, 6, 301–303. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Essential Oil of Austromatthaea elegans L.S. Smith (Monimiaceae) Leaves. J. Essent. Oil Res. 1995, 7, 585–588. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. (E)-β-Ocimene from Two Species of Austromyrtus (Myrtaceae). J. Essent. Oil Res. 1995, 7, 1–4. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Forster, P.I. The essential oils of Australian Austromyrtus sens. lat. Part 1. The A. dulcis group. Flavour Fragr. J. 1995, 10, 69–73. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Forster, P.I. The Essential Oils of Australian Austromyrtus sens. lat. Part 3. The Austromyrtus bidwillii Group. Flavour Fragr. J. 1996, 11, 275–287. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The essential oils of Australian Austromyrtus sens. lat. Part 2. The Austromyrtus lasioclada group. Flavour Fragr. J. 1995, 10, 293–296. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Rozefelds, A.C. The Chemistry of the Australian Gymnosperms—Part 4: The Leaf Oils of the Genus Athrotaxis D. Don (Cupressaceae). J. Essent. Oil Res. 2002, 14, 109–113. [Google Scholar] [CrossRef]
- Brophy, J.J.; Boland, D.J. The leaf essential oil of two chemotypes of Backhousia anisata vickery. Flavour Fragr. J. 1991, 6, 187–188. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Leaf Oils of the Genera Barongia, Mitrantia, Sphaerantia and Ristantia (Myrtaceae). J. Essent. Oil Res. 2003, 15, 226–230. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Essential Oils from the Leaves of Bosistoa F. Muell. ex Benth. (Rutaceae). J. Essent. Oil Res. 2007, 19, 249–254. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Essential Oil of Bouchardatia neurococca (Rutaceae) Leaves. J. Essent. Oil Res. 1994, 6, 505–506. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.i. Leaf Oils of the Genus Brombya (Rutaceae). J. Essent. Oil Res. 2004, 16, 402–404. [Google Scholar] [CrossRef]
- Brophy, J.J.; Forster, P.I.; Goldsack, R.J.; Hibbert, D.B.; Punruckvong, A. Variation in Callistemon viminalis (Myrtaceae): New evidence from leaf essential oils. Aust. Syst. Bot. 1997, 10, 1–13. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I.; Craven, L.A.; Lepschi, B.J. The Leaf Essential Oils of the Australian Members of the Genus Callistemon (Myrtaceae). J. Essent. Oil Res. 1998, 10, 595–606. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The essential oils of Choricarpia leptopetala (F. Muell.) Domin and C. subargentea (C.T. White) L.A.S. Johnson (Myrtaceae). Flavour Fragr. J. 1994, 9, 7–10. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of the Australian Species of Cinnamomum (Lauraceae). J. Essent. Oil Res. 2001, 13, 332–335. [Google Scholar] [CrossRef]
- Brophy, J.J.; Forster, P.I.; Goldsack, R.J. Leaf oils of the Australian species of Clausena and Micromelum (Rutaceae). J. Essent. Oil Res. 2016, 28, 406–412. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The leaf oils of Coatesia and Geijera (Rutaceae) from Australia. J. Essent. Oil Res. 2005, 17, 169–174. [Google Scholar] [CrossRef]
- Brophy, J.J.; Lassak, E.V. Volatile leaf oil of Coleonema pulchellum Williams (Rutaceae). Flavour Fragr. J. 1986, 1, 155–157. [Google Scholar] [CrossRef]
- van Klink, J.W.; Brophy, J.J.; Perry, N.B.; Weavers, R.T. β-Triketones from Myrtaceae: Isoleptospermone from Leptospermum scoparium and Papuanone from Corymbia dallachiana. J. Nat. Prod. 1999, 62, 487–489. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Punruckvong, A.; Forster, P.I.; Fookes, C.J.R. Essential Oils of the Genus Crowea (Rutaceae). J. Essent. Oil Res. 1997, 9, 401–409. [Google Scholar] [CrossRef]
- Brophy, J.J.; Forster, P.I.; Goldsack, R.J. Coconut Laurels: The Leaf Essential Oils from Four Endemic Australian Cryptocarya Species: C. bellendenkerana, C. cocosoides, C. cunninghamii and C. lividula (Lauraceae). Nat. Prod. Commun. 2016, 11, 255–258. [Google Scholar] [CrossRef] [Green Version]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Essential Oil of Cryptocarya cunninghamii Meissner (Lauraceae). J. Essent. Oil Res. 1998, 10, 73–75. [Google Scholar] [CrossRef]
- Southwell, I.A.; Brophy, J.J.; Tucker, D.J. Darwinia citriodora (Myrtaceae), a New Source of Methyl Myrtenate and Methyl Geranate. J. Essent. Oil Res. 2001, 13, 58–60. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Pala-Paul, J.; Copeland, L.M.; Lassak, E.V. Essential oil composition of three Australian endemic species of Darwinia (Myrtaceae). Nat. Prod. Commun. 2010, 5, 1833–1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of the Australian Species of Decaspermum (Myrtaceae). J. Essent. Oil Res. 2005, 17, 611–613. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Essential Oils of the Australian Species of Desmos (Annonaceae). J. Essent. Oil Res. 2002, 14, 298–301. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of Dinosperma erythrococca and D. stipitata (Rutaceae). J. Essent. Oil Res. 2002, 14, 443–446. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Rediscovery and Leaf Oil Chemistry of Dinosperma longifolium T.G. Hartley (Rutaceae). J. Essent. Oil Res. 2004, 16, 350–352. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Rozefelds, A.C. Chemistry of the Australian Gymnosperms—Part 5: Leaf Essential Oils of Some Endemic Tasmanian Gymnosperms: Diselma archeri, Lagarostrobos franklinii, Microcachrys tetragona and Phyllocladus aspleniifolius. J. Essent. Oil Res. 2003, 15, 217–220. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; House, A.P.N.; Lassak, E.V. The Essential Oils of the Genus Doryphora. J. Essent. Oil Res. 1993, 5, 581–586. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Essential Oils of Drummondita calida (Rutaceae: Boronieae). J. Essent. Oil Res. 2006, 18, 652–653. [Google Scholar] [CrossRef]
- Brophy, J.J.; Forster, P.I.; Goldsack, R.J. Essential oils of some Australian monimiaceae. Flavour Fragr. J. 1998, 13, 273–276. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. A Preliminary Investigation of the Leaf Essential Oils of the Australian Species of Endressia, Steganthera and Wilkiea (Monimiaceae). J. Essent. Oil Res. 2009, 21, 115–122. [Google Scholar] [CrossRef]
- Bayly, M.J.; Brophy, J.J.; Forster, P.I.; Goldsack, R.J.; Wilson, P.G. Reinstatement of Eriostemon banksii (Rutaceae), with a Report on the Composition of Leaf Essential Oils in E. banksii and E. australasius s. str. Aust. Syst. Bot. 1998, 11, 13–22. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Copeland, L.M.; Palá-Paúl, J. Essential Oil of Eryngium L. Species from New South Wales (Australia). J. Essent. Oil Res. 2003, 15, 392–397. [Google Scholar] [CrossRef]
- Palá-Paúl, J.; Copeland, L.M.; Brophy, J.J.; Goldsack, R.J. Essential Oil Composition of Eryngium paludosum (Moore et Betche) P.W.Michael: An Endemic Species from Eastern Australia. J. Essent. Oil Res. 2008, 20, 416–419. [Google Scholar] [CrossRef]
- Palá-Paúl, J.; Copeland, L.M.; Brophy, J.J.; Goldsack, R.J. Essential oil composition of Eryngium rosulatum P.W. Michael ined.: A new undescribed species from eastern Australia. Biochem. Syst. Ecol. 2006, 34, 796–801. [Google Scholar] [CrossRef]
- Palá-Paúl, J.; Brophy, J.J.; Goldsack, R.J.; Copeland, L.M.; Pérez-Alonso, M.J.; Velasco-Negueruela, A. Essential oil composition of the seasonal heterophyllous leaves of Eryngium vesiculosum from Australia. Aust. J. Bot. 2003, 51, 497–501. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Composition of the Leaf Oils of the Australian Species of Euodia and Melicope (Rutaceae). J. Essent. Oil Res. 2004, 16, 286–293. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Leaf Essential Oil of Fitzalania heteropetala F. Muell. (Annonaceae). J. Essent. Oil Res. 1997, 9, 93–94. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Essential Oil of Galbulimima baccata (Himantandraceae) from Queensland, Australia. J. Essent. Oil Res. 2005, 17, 536–538. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J. Essential Oil of the Leaves and Flowers of Geleznowia verrucosa Turcz. (Rutaceae). J. Essent. Oil Res. 1995, 7, 663–665. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Leaf Essential Oils of the Australian Species of Gyrocarpus and Hernandia (Hernandiaceae). J. Essent. Oil Res. 2000, 12, 717–722. [Google Scholar] [CrossRef]
- Forster, P.I.; Brophy, J.J.; Goldsack, R.J. Variation in Australian populations of Halfordia kendack S.L. (Rutaceae): Evidence from leaf essential oils. Aust. Syst. Bot. 2004, 17, 571–580. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Leaf Oils of the Australian Species of the Genus Haplostichanthus (Annonaceae). J. Essent. Oil Res. 2006, 18, 64–67. [Google Scholar] [CrossRef]
- Brophy, J.J.; Fookes, C.J.R.; House, A.P.N. The Leaf Essential Oil of Haplostichanthus johnsonii F. Muell. J. Essent. Oil Res. 1992, 4, 315–316. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of the Australian Species of Hedycarya (Monimiaceae). J. Essent. Oil Res. 2005, 17, 432–436. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Copeland, L.M. Leaf Oils of the Genus Homoranthus (Myrtaceae). J. Essent. Oil Res. 2004, 16, 46–60. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Cornwell, C.P.; Leach, D.N.; Wyllie, S.G.; Forster, P.I.; Fookes, C.J.R. (Z)-β-Ocimene from Two Species of Homoranthus (Myrtaceae). J. Essent. Oil Res. 1998, 10, 229–233. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J. The leaf essential oil of Idiospermum australiense (Diels) S. T. Blake (idiospermaceae). Flavour Fragr. J. 1992, 7, 79–80. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Punruckvong, A.; Bean, A.R.; Forster, P.I.; Lepschi, B.J.; Doran, J.C.; Rozefelds, A.C. Leaf essential oils of the genus Leptospermum (Myrtaceae) in eastern Australia. Part 7. Leptospermum petersonii, L. liversidgei and allies. Flavour Fragr. J. 2000, 15, 342–351. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Bean, A.R.; Forster, P.I.; Lepschi, B.J. Leaf essential oils of the genus Leptospermum (Myrtaceae) in eastern Australia. Part 3. Leptospermum arachnoides and allies. Flavour Fragr. J. 1999, 14, 85–91. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Bean, A.R.; Forster, P.I.; Lepschi, B.J. Leaf essential oils of the genus Leptospermum (Myrtaceae) in eastern Australia. Part 2. Leptospermum blakelyi and allies. Flavour Fragr. J. 1998, 13, 353–358. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I.; Bean, A.R.; Clarkson, J.R.; Lepschi, B.J. Leaf essential oils of the genus Leptospermum (Myrtaceae) in Eastern Australia. Part 1. Leptospermum brachyandrum and Leptospermum pallidum groups. Flavour Fragr. J. 1998, 13, 19–25. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Lassak, E.V. Leaf Essential Oils of Some Leptospermum (Myrtaceae) Species from Southern and Western Australia. J. Essent. Oil Res. 1999, 11, 1–5. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Bean, A.R.; Forster, P.I.; Lepschi, B.J. Leaf essential oils of the genus Leptospermum (Myrtaceae) in eastern Australia, Part 6. Leptospermum polygalifolium and allies. Flavour Fragr. J. 2000, 15, 271–277. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Essential Oil of Lindera queenslandica (Lauraceae). J. Essent. Oil Res. 1999, 11, 453–455. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I.; Clarkson, J.R. The Essential Oil of Welchiodendron longivalve (F. Muell.) Peter G. Wilson & J. T. Waterh. and Lindsayomyrtus racemoides (Greves) Craven (Myrtaceae) Leaves. Flavour Fragr. J. 1996, 11, 67–70. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Essential oils of the genus Lophostemon (Myrtaceae). Flavour Fragr. J. 2000, 15, 17–20. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I.; Hutton, I. Leaf Essential Oils of Lunasia amara var. amara and Sarcomelicope simplicifolia subsp. simplicifolia (Rutaceae) from Australia. J. Essent. Oil Res. 1997, 9, 141–144. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Leaf Essential Oils of Lycopus australis (Lamiaceae), the Australian Gipsywort. J. Essent. Oil Res. 2005, 17, 133–134. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Essential Oil of Lysicarpus angustifolius (Hook.) Druce (Myrtaceae). J. Essent. Oil Res. 1994, 6, 139–143. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.l. The Leaf Oils of the Australian Species of Medicosma (Rutaceae). J. Essent. Oil Res. 2004, 16, 161–166. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Hutton, I. Leaf Oils of the Endemic Melicope (Rutaceae) of Lord Howe Island. J. Essent. Oil Res. 2004, 16, 449–452. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Chemotype Variation in the Leaf Essential Oils of Melicope melanophloia C.T. White (Rutaceae). J. Essent. Oil Res. 1997, 9, 279–282. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of the Queensland Species of Melodorum (Annonaceae). J. Essent. Oil Res. 2004, 16, 483–486. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Lawrence, B.M.; Forster, P.I. Essential Oil of Mentha diemenica (Lamiaceae). J. Essent. Oil Res. 1996, 8, 179–181. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I.; Fookes, C.J.R. Leaf Essential Oil of Mentha grandiflora Benth. (Lamiaceae). J. Essent. Oil Res. 1997, 9, 459–461. [Google Scholar] [CrossRef]
- Southwell, I.A.; Brophy, J.J. Isoamyl Isovalerate from Essential Oil of Micromyrtus striata J. W. Green. J. Essent. Oil Res. 1991, 3, 281–283. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Rozefelds, A.C. Chemistry of Australian Gymnosperms. Part III. Leaf Oils of the Genus Microstrobos (Podocarpaceae). J. Essent. Oil Res. 2001, 13, 108–109. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of the Australian Species of Miliusa (Annonaceae). J. Essent. Oil Res. 2004, 16, 253–255. [Google Scholar] [CrossRef]
- Brophy, J.; Forster, P.; Goldsack, R. Diversity in Australian populations of Murraya paniculate (Rutaceae): New evidence from volatile leaf oils. Aust. Syst. Bot. 1994, 7, 409–418. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Forster, P.I. The Leaf Oils of the Australian Species of Neolitsea (Lauraceae). J. Essent. Oil Res. 2002, 14, 191–195. [Google Scholar] [CrossRef]
- Brophy, J.J.; Clarkson, J.R. The essential oils of the genus Neofabricia (Myrtaceae). Biochem. Syst. Ecol. 1992, 20, 689–696. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Clarkson, J.R. The Essential Oil of Osbornia octodonta F. Muell. J. Essent. Oil Res. 1993, 5, 1–5. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Essential Oils from the Leaves of the Australian Species of Palmeria (Monimiaceae). J. Essent. Oil Res. 2004, 16, 312–317. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Forster, P.I. The Essential Oils of Pentaceras australe (Rutaceae). J. Essent. Oil Res. 2002, 14, 348–350. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Punruckvong, A.; Forster, P.I. The leaf essential oils of Pilidiostigma (Myrtaceae). Flavour Fragr. J. 1999, 14, 143–146. [Google Scholar] [CrossRef]
- Wyllie, S.G.; Brophy, J.J.; Sarafis, V.; Hobbs, M. Volatile Components of the Fruit of Pistacia Lentiscus. J. Food Sci. 1990, 55, 1325–1326. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Essential Oil of Pitaviaster haplophyllus (F. Muell.) T. G. Hartley. J. Essent. Oil Res. 2002, 14, 130–131. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I.; Rozefelds, A.C. Chemistry of the Australian Gymnosperms. Part 6. Leaf Oils of the Australian Species of Genus Podocarpus. J. Essent. Oil Res. 2004, 16, 342–346. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Leaf Oils of the Australian Species of Polyalthia (Annonaceae). J. Essent. Oil Res. 2001, 13, 5–7. [Google Scholar] [CrossRef]
- Hunter, M.V.; Brophy, J.J.; Ralph, B.J.; Bienvenu, F.E. Composition of Polygonum odoratum Lour. from Southern Australia. J. Essent. Oil Res. 1997, 9, 603–604. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Hook, J.M.; Fookes, C.J.R.; Forster, P.I. The Leaf Essential Oils of the Australian Species of Pseuduvaria (Annonaceae). J. Essent. Oil Res. 2004, 14, 362–366. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The leaf essential oils of the Australian species of Rhodamnia (Myrtaceae). Flavour Fragr. J. 1997, 12, 345–354. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Essential Oils of the Australian Species of Rhodomyrtus (Myrtaceae). Flavour Fragr. J. 1997, 12, 103–108. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Forster, P.I. Essential Oils of Australian Gymnosperms. Part 1. The Leaf Oil of Sundacarpus amarus (Blume) C.N. Page (Podocarpaceae). J. Essent. Oil Res. 2000, 12, 421–423. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Bean, A.R.; Forster, P.I.; Fookes, C.J.R. The Leaf Essential Oils of the Genus Syncarpia Ten. (Myrtaceae). Flavour Fragr. J. 1996, 11, 361–366. [Google Scholar] [CrossRef]
- Brophy, J.J.; Craig, D.C.; Goldsack, R.J.; Fookes, C.J.R. Hillyl acetate, a keto-acetate from the leaf steam volatiles ofSyncarpia hillii. Phytochemistry 1994, 37, 1645–1647. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Essential Oil of Thaleropia queenslandica (Myrtaceae). J. Essent. Oil Res. 1997, 9, 587–588. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I.; Clarkson, J.R. The Essential Oils of the Queensland Species of Thryptomene (Myrtaceae). J. Essent. Oil Res. 2000, 12, 11–13. [Google Scholar] [CrossRef]
- Pala-Paul, J.; Copeland, L.M.; Brophy, J.J. The Essential Oil Composition of Trachymene incisa Rudge subsp. incisa Rudge from Australia. Plants 2021, 10, 601. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Essential Oils of Australian Species of the Genera Tristaniopsis and Tristania (Myrtaceae). J. Essent. Oil Res. 1999, 11, 661–665. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Essential Oils of the Australian Species of Uromyrtus (Myrtaceae). Flavour Fragr. J. 1996, 11, 133–138. [Google Scholar] [CrossRef]
- Suksamrarn, A.; Aphaijitt, S.; Brophy, J.J. The volatile leaf oil of Vitex limonifolia Wall. Flavour Fragr. J. 1990, 5, 53–55. [Google Scholar] [CrossRef]
- Suksamrarn, A.; Werawattanametin, K.; Brophy, J.J. Variation of essential oil constituents in Vitex trifolia species. Flavour Fragr. J. 1991, 6, 97–99. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Essential Oils of Viticipremna queenslandica (Lamiaceae). J. Essent. Oil Res. 2008, 20, 403–404. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Fookes, C.J.R.; Forster, P.I. Leaf Essential Oils of the Genus Waterhousea (Myrtaceae). J. Essent. Oil Res. 2002, 14, 31–34. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Essential Oils of the Australian Species of Xylopia (Annonaceae). J. Essent. Oil Res. 1998, 10, 469–472. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Goldsack, G.; House, A.P.N. Leaf Essential Oils of the Australian Members of the Genus Zygogynum. J. Essent. Oil Res. 1994, 6, 353–361. [Google Scholar] [CrossRef]
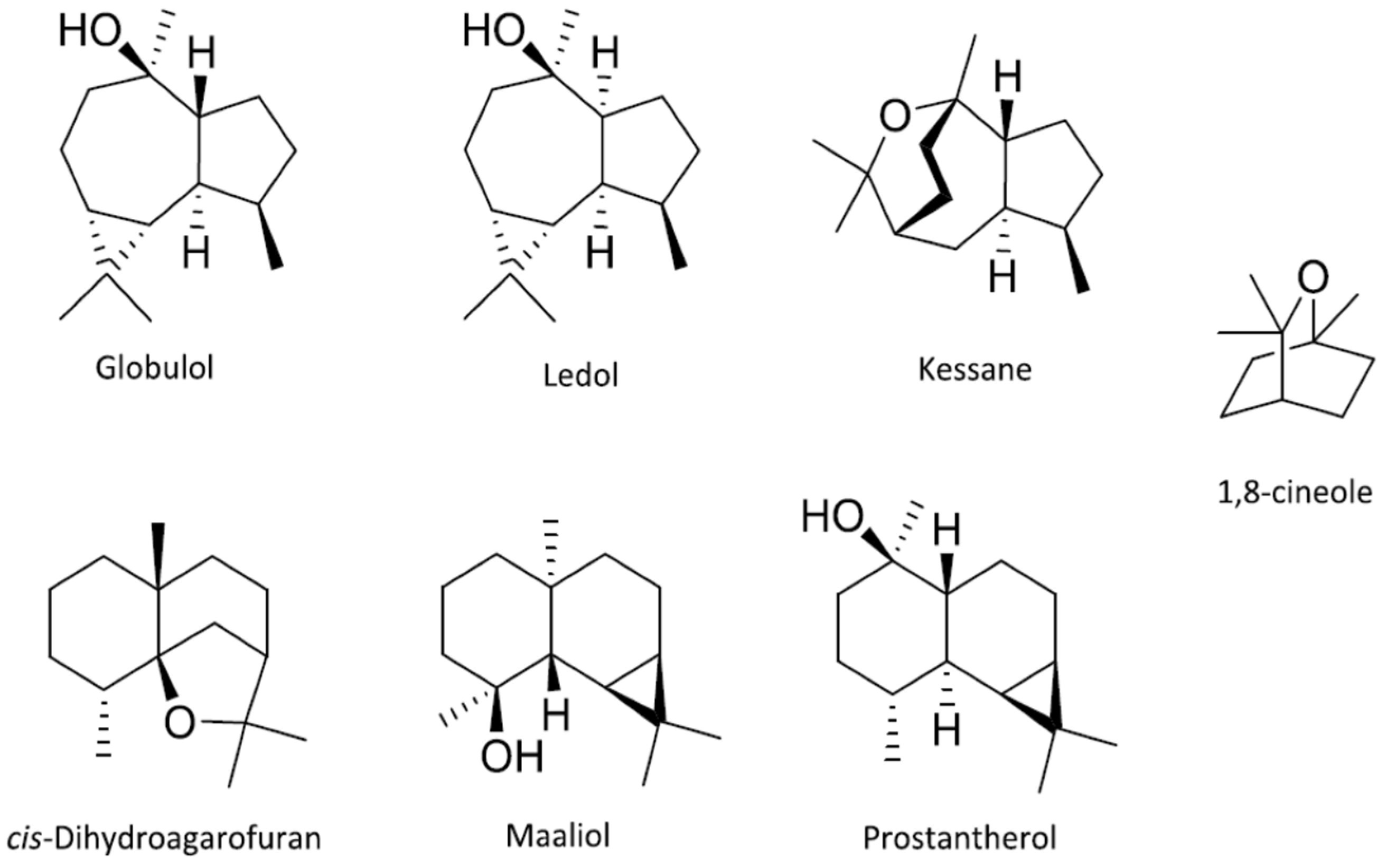
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadgrove, N.J. Purely Australian Essential Oils Past and Present: Chemical Diversity, Authenticity, Bioactivity, and Commercial Value. Diversity 2022, 14, 124. https://doi.org/10.3390/d14020124
Sadgrove NJ. Purely Australian Essential Oils Past and Present: Chemical Diversity, Authenticity, Bioactivity, and Commercial Value. Diversity. 2022; 14(2):124. https://doi.org/10.3390/d14020124
Chicago/Turabian StyleSadgrove, Nicholas John. 2022. "Purely Australian Essential Oils Past and Present: Chemical Diversity, Authenticity, Bioactivity, and Commercial Value" Diversity 14, no. 2: 124. https://doi.org/10.3390/d14020124
APA StyleSadgrove, N. J. (2022). Purely Australian Essential Oils Past and Present: Chemical Diversity, Authenticity, Bioactivity, and Commercial Value. Diversity, 14(2), 124. https://doi.org/10.3390/d14020124