
Evaluation of Genetic Variability among Three Pistacia Species Using Internal Transcribed Spacer 1 (ITS1) Marker

 ,
,  , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. DNA Extraction
2.3. Amplification and Purification of ITS1 Region of rDNA
2.4. Sequencing and Alignment of ITS Sequences
2.5. Phylogenetic Analysis
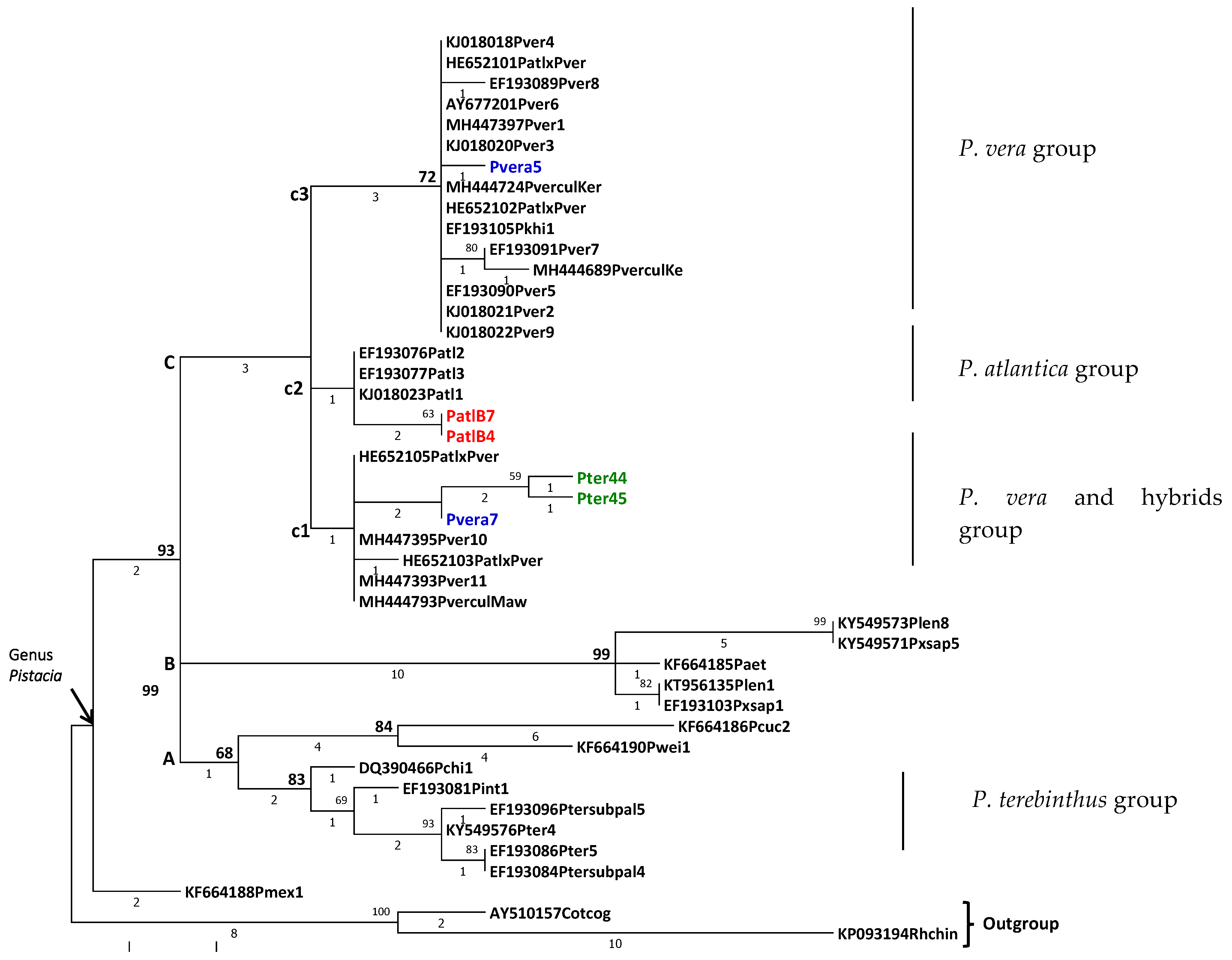
3. Results
ITS Amplification and Sequence Comparison
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ak, B.E.; Parlakçı, H. Pistacia lentiscus in the Mediterranean Region in Turkey. Acta Hortic. 2009, 818, 77–82. [Google Scholar] [CrossRef]
- Ahmed, Z.B.; Yousfi, M.; Viaene, J.; Dejaegher, B.; Demeyer, K.; Heyden, Y.V. Four Pistacia atlantica subspecies (atlantica, cabulica, kurdica and mutica): A review of their botany, ethnobotany, phytochemistry and pharmacology. J. Ethnopharmacol. 2021, 265, 113329. [Google Scholar] [CrossRef] [PubMed]
- Al-Saghir, M.G. Phylogenetic Analysis of the Genus Pistacia L. (Anacardiaceae) Based on Morphological Data. Asian J. Plant Sci. 2010, 9, 28–35. [Google Scholar] [CrossRef][Green Version]
- El Zerey-Belaskri, A. Taxonomic and botanical retrospective review of Pistacia atlantica Desf. (Anacardiaceae). Arab. J. Med. Aromat. Plants 2019, 5, 47–77. [Google Scholar] [CrossRef]
- Belhadj, S.; Derridj, A.; Auda, Y.; Gers, C.; Gauquelin, T. Analyse de la variabilité morphologique chez huit populations spontanées de Pistacia atlantica en Algérie. Can. J. Bot. 2008, 86, 520–532. [Google Scholar] [CrossRef]
- El Zerey-Belaskria, A.; Ribeirob, T.; Alcarazc, M.L.; El Zereyd, W.; Castroe, S.; Loureiroe, J.; Benhassainia, H.; Hormaza, J.I. Molecular characterization of Pistacia atlantica Desf. subsp. atlantica (Anacardiaceae) in Algeria: Genome size determination, chromosome count and genetic diversity analysis using SSR markers. Sci. Hortic. 2018, 227, 278–287. [Google Scholar] [CrossRef]
- Labdelli, A.; De La Herrán, R.; Arafeh, R.; Resentini, F.; Trainotti, L.; Halis, Y.; Adda, A.; Merah, O. Genetic Variation in Damaged Populations of Pistacia atlantica Desf. Plants 2020, 9, 1541. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. Multiple origins of the Allotetraploid Tragopogon mirus (Compositae): rDNA evidence. Syst. Bot. 1991, 16, 407–413. [Google Scholar] [CrossRef]
- Parfitt, D.E.; Badenes, M.L. Phylogeny of the genus Pistacia as determined from analysis of the chloroplast genome. Proc. Natl. Acad. Sci. USA 1997, 94, 7987–7992. [Google Scholar] [CrossRef]
- Kafkas, S.; Perl-Treves, R. Inter-specific relationships in the genus Pistacia L. (Anacardiaceae) based on RAPD fingerprints. Hortic. Sci. 2002, 37, 168–171. [Google Scholar] [CrossRef]
- Basha, A.I.; Padulosi, S.; Chabane, K.; Hadj-Hassan, A.; Dulloo, E.; Pagnotta, M.A.; Porceddu, E. Genetic diversity of Syrian pistachio varieties evaluated by AFLP markers. Genet. Resour. Crop Evol. 2007, 54, 1807–1816. [Google Scholar] [CrossRef]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F.; Campbell, C.S.; Donoghue, M.J. The ITS region of nuclear ribosomal DNA: A valuable source of evidence on angiosperm phylogeny. Ann. Mo. Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods Applications; Innis, M.A., Gelfand, D.H., Shinsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Baldwin, B.G. Phylogenetic utility of the internal transcribed spacer of nuclear ribosomal DNA in plants: An example from the Compositae. Mol. Phylogenet. Evol. 1992, 1, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Suh, Y.; Thien, L.B.; Reeve, H.E.; Zimmer, E.A. Molecular evolution and phylogenetic implications of ribosomal DNA in Winteraceae. Am. J. Bot. 1993, 80, 1042–1055. [Google Scholar] [CrossRef]
- Yi, T.; Wen, J.; Golan-Glodhirsh, A.; Parfitt, D.E. Phylogenetics and reticulate evolution in Pistacia (Anacardiaceae). Am. J. Bot. 2008, 95, 241–251. [Google Scholar] [CrossRef]
- Aznarte-Mellado, C.; Sola-Campoy, P.J.; Robles, F.; Rejón, C.R.; Herrán, R.; Navajas-Pérez, R. Molecular characterization of the interspecific hybrid Pistacia vigros (P. vera L. × P. atlantica Desf.). Sci. Hort. 2014, 179, 180–183. [Google Scholar] [CrossRef]
- Alaei, H.; Mohammadi, A.H.; Dehghani, A. Molecular characterization of the rDNA-ITS sequence and a PCR diagnostic technique for Pileolaria terebinthi, the cause of pistachio rust. Phytopathol. Mediterr. 2012, 51, 488–495. [Google Scholar] [CrossRef]
- Abuduli, A.; Aydin, Y.; Sakiroglu, M.; Onay, A.; Ercisli, S.; Uncuoglu, A.A. Molecular Evaluation of Genetic Diversity in Wild-Type Mastic Tree (Pistacia lentiscus L.). Biochem. Genet. 2016, 54, 619–635. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bul. 1987, 19, 11–15. [Google Scholar]
- Kafkas, S. Phylogenetic analysis of the genus Pistacia by AFLP markers. Plant Syst. Evol. 2006, 262, 113–124. [Google Scholar] [CrossRef]
- Al-Sousli, M.; Faory, H.; Nakar, M.; Zaid, S.; Al-Safadi, B.; Al-Saghir, M. Genetic Relationships among Some Pistacia Species (Anacardiaceae) in Syria. Middle East J. Sci. Res. 2014, 21, 1487–1496. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software versions 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Pearson, W.R.; Lipman, D.J. Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA 1988, 85, 2444–2448. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Esmailpour, A. Distribution, use and conservation of pistachio in Iran. In Towards a Comprehensive Documentation and Use of Pistacia Genetic Diversity in Central and West Asia, North Africa and Europe. Report of the IPGRI Workshop; 14–17 December 1998, Ibrid, Jordan; Padulosi, S., Hadj-Hassan, A., Eds.; IPGRI: Rome, Italy, 2001; p. 16. [Google Scholar]
- Pazouki, L.; Mardi, M.; Shanjani, P.S.; Hagidimitriou, M.; Pirseyedi, S.M.; Naghavi, M.R.; Avanzato, D.; Vendramin, E.; Kafkas, S.; Ghareyazi, B.; et al. Genetic diversity and relationships among Pistacia species and cultivars. Conserv. Genet. 2010, 11, 311–318. [Google Scholar] [CrossRef]
- Kafkas, S.; Perl-Treves, R. Morphological and molecular phylogeny of Pistacia species in Turkey. Theor. Appl. Genet. 2001, 102, 908–915. [Google Scholar] [CrossRef]
- Álvarez, I.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phylogenet. Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef]
- Wissemann, V. Hybridization and the evolution of the nrITS spacer region. In Plant Genome: Biodiversity and Evolution; Sharma, A.K., Sharma, A., Eds.; Science Publishers, Inc.: Enfield, NH, USA, 2003; pp. 57–66. [Google Scholar]
- Sharma, S.; Rustgi, S.; Balyan, H.S.; Gupta, P.K. Internal transcribed spacer (ITS) sequences of ribosomal DNA of wild barley and their comparison with ITS sequences in common wheat. Barley Genet. Newlet. 2002, 32, 38–45. [Google Scholar]
- Talebi, M.; Akbari, M.; Zamani, M.; Sayed-Tabatabaei, B.E. Molecular polymorphism in Pistacia vera L. using non-coding regions of chloroplast DNA. J. Genet. Engin. Biotechnol. 2016, 14, 31–37. [Google Scholar] [CrossRef]
- Muir, P.; Li, S.; Lou, S.; Wang, D.; Spakowicz, D.J.; Salichos, L.; Zhang, J.; Weinstock, G.M.; Isaacs, F.; Rozowsky, J.; et al. The real cost of sequencing: Scaling computation to keep pace with data generation. Genome Biol. 2016, 17, 53. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| N° | Abbreviation | BLAST GenBank Access Number | Species Name |
|---|---|---|---|
| 1 | KJ018023Patl1 | KJ018023 | Pistacia atlantica Desf. |
| 2 | EF193076Patl2 | EF193076 | Pistacia atlantica Desf. |
| 3 | EF193077Patl3 | EF193077 | Pistacia atlantica Desf. |
| 4 | HE652105PatlxPver | HE652105 | P. atlantica × P. vera cultivar BIO200, clone BIO200_5 |
| 5 | HE652103PatlxPver | HE652103 | P. atlantica × P. vera cultivar BIO200, clone BIO200_3 |
| 6 | HE652102PatlxPver | HE652102 | P. atlantica × P. vera cultivar BIO200, clone BIO200_2 |
| 7 | HE652101PatlxPver | HE652101 | P. atlantica × P. vera cultivar BIO200, clone BIO200_1 |
| 8 | KF664185Paet | KF664185 | Pistacia aethiopica voucher Yi12041(KUN) |
| 9 | DQ390466Pchi1 | DQ390466 | Pistacia chinensis |
| 10 | KF664186Pcuc2 | KF664186 | Pistacia cucphuongensis voucher SK Dell 1100(US) |
| 11 | EF193081Pint1 | EF193081 | Pistacia integerrima voucher Parfitt 54 |
| 12 | EF193105Pkhi1 | EF193105 | Pistacia khinjuk |
| 13 | KT956135Plen1 | KT956135 | Pistacia lentiscus L. |
| 14 | KY549573Plen8 | KY549573 | Pistacia lentiscus L. |
| 15 | KF664188Pmex1 | KF664188 | Pistacia mexicana voucher Wen8675(US) |
| 16 | KY549576Pter4 | KY549576 | Pistacia terebinthus L. |
| 17 | EF193084Ptersubpal4 | EF193084 | Pistacia terebinthus L. |
| 18 | EF193096Ptersubpal5 | EF193096 | P. terebinthus subsp. palaestina voucher Golan 1.222 clone 1 |
| 19 | EF193086Pter5 | EF193086 | Pistacia terebinthus voucher Golan 1.350 |
| 20 | KJ018021Pver2 | KJ018021 | Pistacia vera L. |
| 21 | KJ018020Pver3 | KJ018020 | Pistacia vera L. |
| 22 | KJ018022Pver9 | KJ018022 | Pistacia vera L. |
| 23 | KJ018018Pver4 | KJ018018 | Pistacia vera L. |
| 24 | AY677201Pver6 | KJ018022 | Pistacia vera L. |
| 25 | MH447397Pver1 | MH447397 | Pistacia vera L. |
| 26 | MH447395Pver10 | MH447395 | Pistacia vera L. |
| 27 | MH447393Pver11 | MH447393 | Pistacia vera L. |
| 28 | MH444793PverculMaw | MH444793 | Pistacia vera L. |
| 29 | MH444724PverculMaw | MH444724 | Pistacia vera L. |
| 30 | MH444689PverculKe | MH444689 | Pistacia vera L. |
| 31 | EF193089Pver8 | EF193089 | P. vera voucher Wen 7099 clone 1 |
| 32 | EF193090Pver5 | EF193090 | P. vera voucher Wen 7099 clone 2 |
| 33 | EF193091Pver7 | EF193091 | P. vera voucher Wen 7099 clone 3 |
| 34 | KF664190Pwei1 | KF664190 | Pistacia weinmaniifolia |
| 35 | EF193103Pxsap1 | EF193103 | Pistacia × saportae voucher T10 clone 4 |
| 36 | KY549571Pxsap5 | KY549571 | Pistacia × saportae |
| 37 | AY510157Cotcog | AY510157 | Cotinus coggygria |
| 38 | KP093194Rhchin | KP093194 | Rhus chinensis |
| 39 | PatlB7 (in this study) | ON026857 | Pistacia atlantica Desf.B7 |
| 40 | PatlB4 (in this study) | ON026858 | Pistacia atlantica Desf.B4 |
| 41 | Pter44 (in this study) | ON026856 | Pistacia terebinthus L.44 |
| 42 | Pter45 (in this study) | ON026855 | Pistacia terebinthus L.45 |
| 43 | Pvera5 (in this study) | ON026854 | Pistacia vera L.5 |
| 44 | PVera7 (in this study) | ON026853 | Pistacia vera L.7 |
| Type of Site | Number of Sequences |
|---|---|
| Total number of sites | 248 |
| Number of sites retained (C) | 185 (74.60%) |
| Number of variable sites (V) | 61 (24.60%) |
| Number of informative sites (Pi) (Parsim-info) | 40 (16.13%) |
| Number of Singleton sites (S) | 21 (8.47%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labdelli, A.; De La Herrán, R.; Resentini, F.; Trainotti, L.; Tahirine, M.; Merah, O. Evaluation of Genetic Variability among Three Pistacia Species Using Internal Transcribed Spacer 1 (ITS1) Marker. Diversity 2022, 14, 1051. https://doi.org/10.3390/d14121051
Labdelli A, De La Herrán R, Resentini F, Trainotti L, Tahirine M, Merah O. Evaluation of Genetic Variability among Three Pistacia Species Using Internal Transcribed Spacer 1 (ITS1) Marker. Diversity. 2022; 14(12):1051. https://doi.org/10.3390/d14121051
Chicago/Turabian StyleLabdelli, Amina, Roberto De La Herrán, Francesca Resentini, Livio Trainotti, Mohammed Tahirine, and Othmane Merah. 2022. "Evaluation of Genetic Variability among Three Pistacia Species Using Internal Transcribed Spacer 1 (ITS1) Marker" Diversity 14, no. 12: 1051. https://doi.org/10.3390/d14121051
APA StyleLabdelli, A., De La Herrán, R., Resentini, F., Trainotti, L., Tahirine, M., & Merah, O. (2022). Evaluation of Genetic Variability among Three Pistacia Species Using Internal Transcribed Spacer 1 (ITS1) Marker. Diversity, 14(12), 1051. https://doi.org/10.3390/d14121051