


Buffalo on the Edge: Factors Affecting Historical Distribution and Restoration of Bison bison in the Western Cordillera, North America


Abstract
1. Introduction
2. Materials and Methods
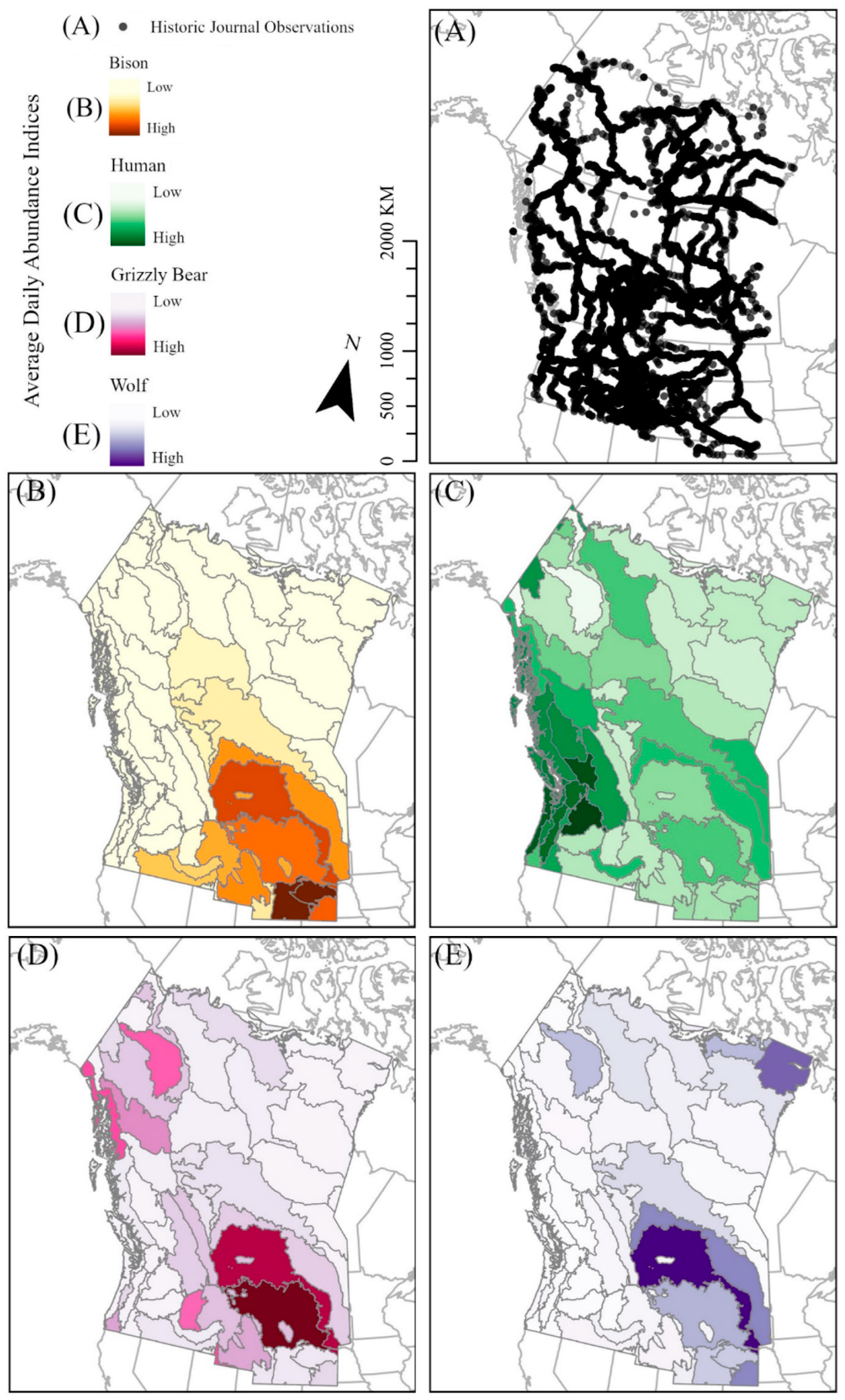
2.1. First-Person Journal Observations
2.2. Journal Data Analysis
2.2.1. Bottom-Up Effects on Bison
2.2.2. Top-Down Effects on Bison
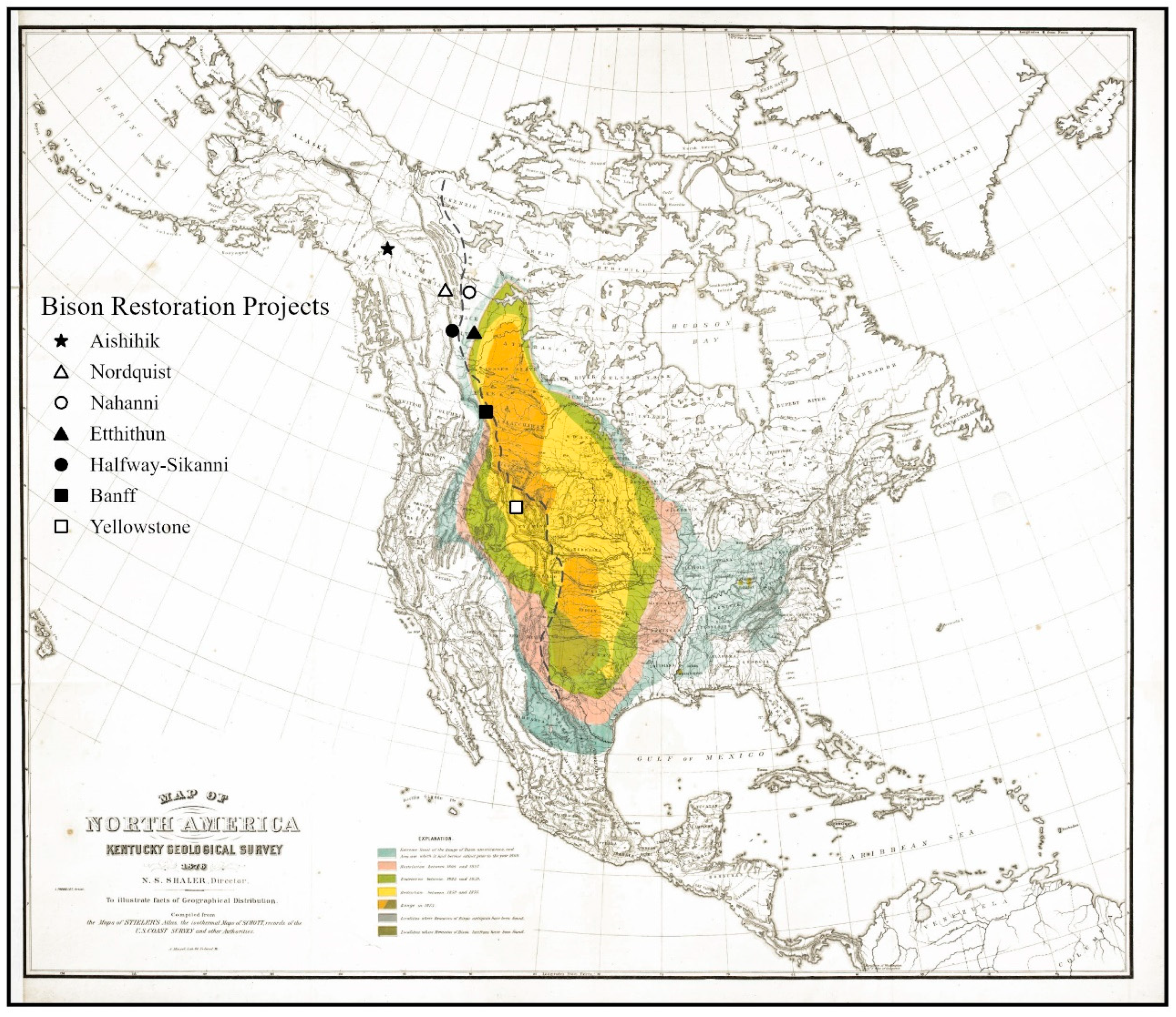
2.3. Cordilleran Bison Restoration Project Demographics and Outcomes
3. Results
3.1. Historical Range and Abundance of Bison
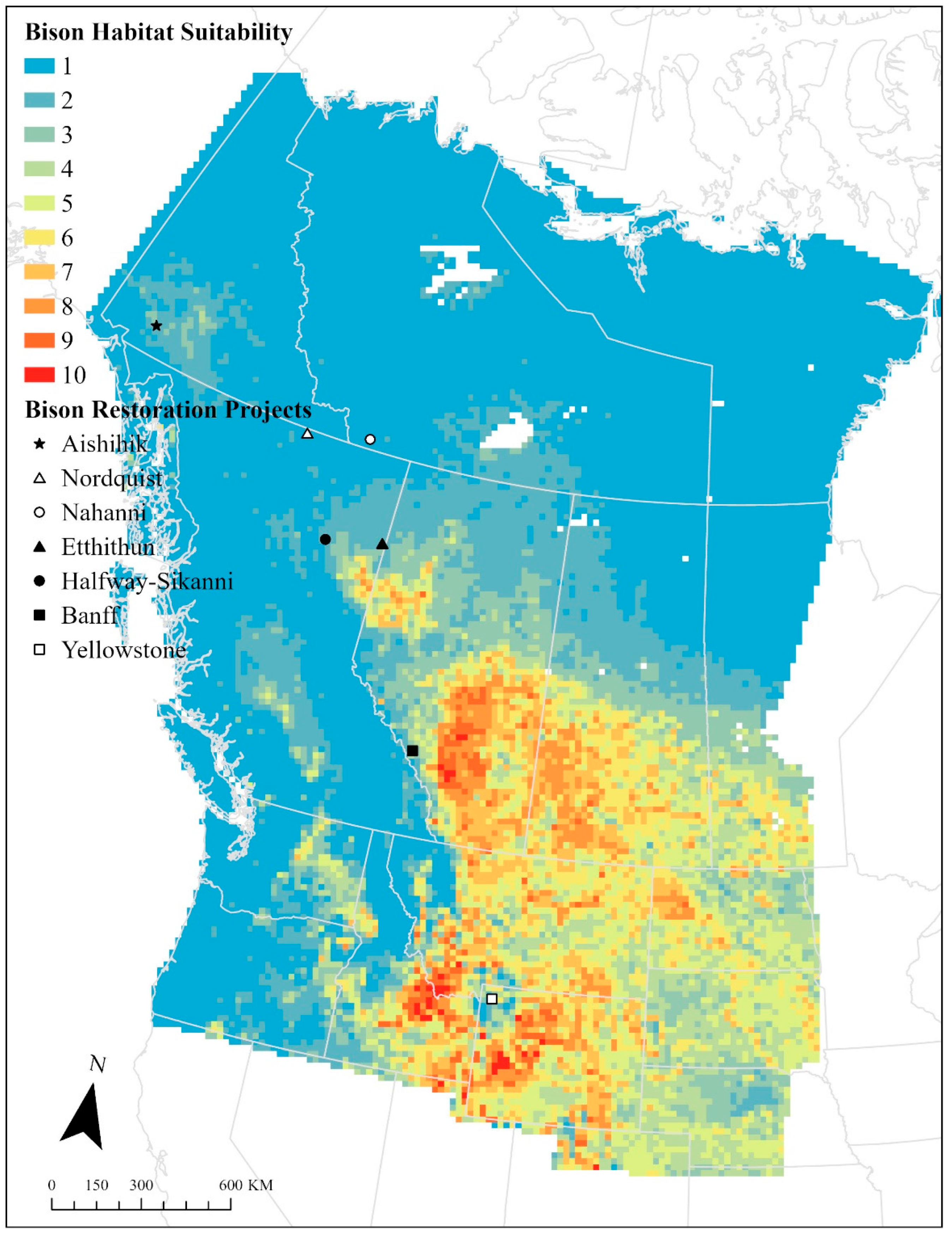
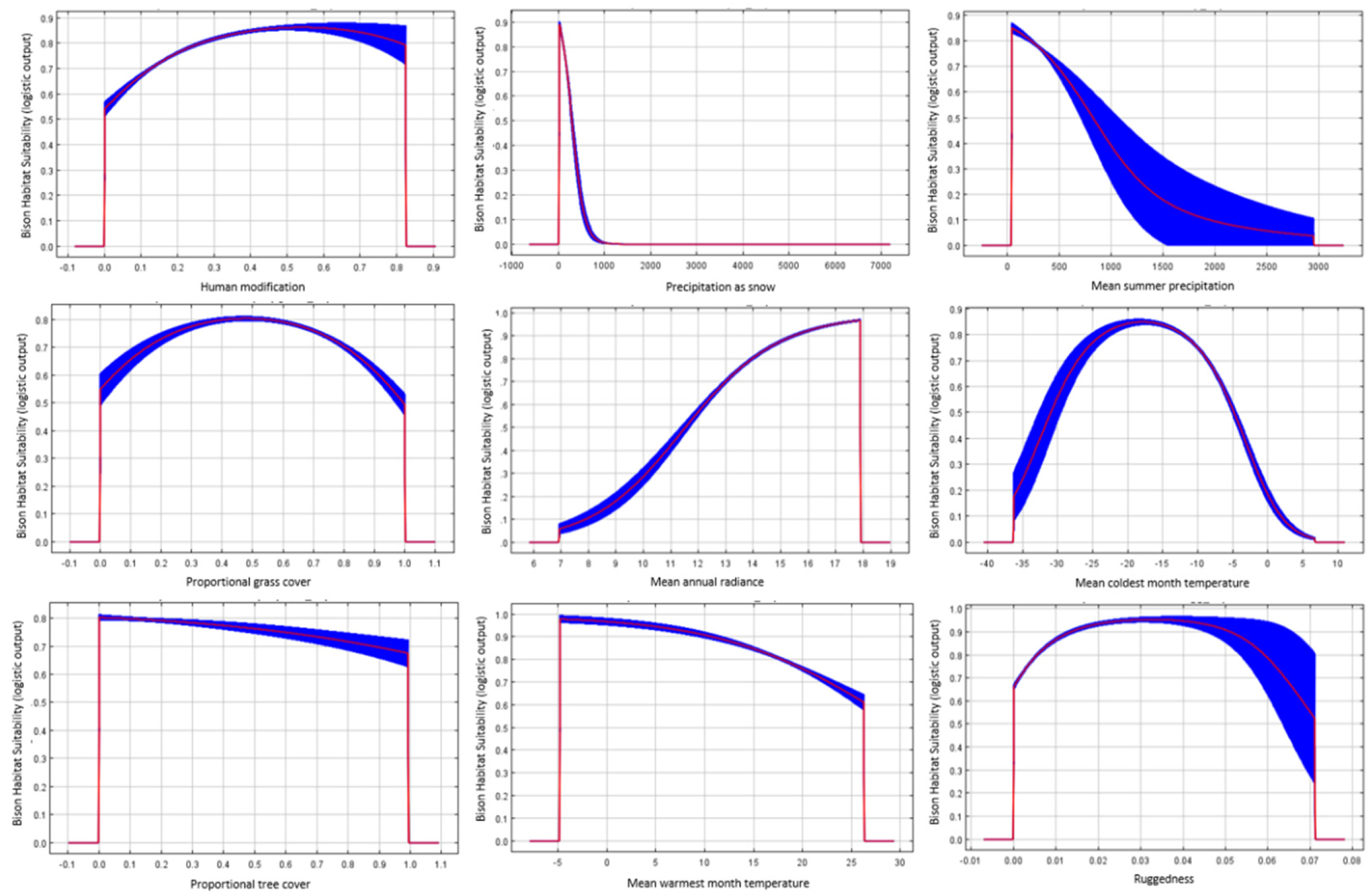
3.2. Factors Affecting Bison Distribution
3.3. Cordilleran Bison Restoration Project Outcomes
4. Discussion
4.1. Processes Limiting Historic Bison Distribution
4.2. Implications for Bison Restoration in the Western Cordillera
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Gates, C.C.; Freese, G.H.; Gogan, P.; Kotzman, M. (Eds.) American Bison: Status Survey and Conservation Guidelines; International Union Conservation Nature: Gland, Switzerland, 2011. [Google Scholar]
- Isenberg, A.C. The Destruction of the Bison an Environmental History, 1750–1920; Cambridge University Press: Cambride, MA, USA, 2000. [Google Scholar]
- Allen, J.A. The American Bisons, Living and Extinct; Harvard College Memoirs of the Museum of Comparative Zoology; University Press: Cambridge, MA, USA, 1876; Volume 10. [Google Scholar]
- Roe, F.G. The North American Buffalo; University of Toronto Press: Toronto, ON, Canada, 1972. [Google Scholar]
- Wildlife Conservation Policy: A Reader; Geist, V., McTaggart-Cowan, I., Eds.; Detselig Enterprises: Calgary, AL, Canada, 1998. [Google Scholar]
- Hessami, M.A.; Bowles, E.; Popp, J.N.; Ford, A.T. Indigenizing the North American Model of Wildlife Conservation. FACETS 2021, 6, 1285–1306. [Google Scholar] [CrossRef]
- Gates, P.W. History of Public Land Law Development; US Government Printing Office: Washington, DC, USA, 1968; Volume 62.
- Donahue, D.L. The Western Range Revisited: Removing Livestock from Public Lands to Conserve Native Biodiversity; University of Oklahoma Press: Norman, OK, USA, 1999; Volume 5. [Google Scholar]
- Bryner, G. Balancing preservation and logging: Public lands policy in British Columbia and the Western United States. Policy Stud. J. 1999, 27, 307–327. [Google Scholar] [CrossRef]
- Hebblewhite, M.; Hilty, J.A.; Williams, S.; Locke, H.; Chester, C.; Johns, D.; Kehm, G.; Francis, W.L. Can a large-landscape conservation vision contribute to achieving biodiversity targets? Conserv. Sci Pract. 2022, 4, e588. [Google Scholar] [CrossRef]
- Daubenmire, R. The western limits of the range of the American Bison. Ecology 1985, 66, 622–624. [Google Scholar] [CrossRef]
- Van Vuren, D. Bison west of the Rocky Mountains: An alternative explanation. Northwest Sci. 1987, 61, 65–69. [Google Scholar]
- Wendt, J.A.F.; McWethy, D.B.; Widga, C.; Shuman, B.N. Large-scale climatic drivers of Bison distribution and abundance in North America since the Last Glacial Maximum. Quat. Sci. Rev. 2022, 284, 107472. [Google Scholar] [CrossRef]
- Bailey, J.A. Historic distribution and abundance of Bison in the Rocky Mountains of the United States. Intermt. J. Sci. 2016, 22, 36–53. [Google Scholar]
- Reeves, B.O.K. Bison killing in the Southwestern Alberta Rockies. Plains Anthropol. 1978, 23, 63–78. [Google Scholar] [CrossRef]
- Zedeño, M.N. Bison hunters and the Rocky Mountains: An evolving partnership. Quat. Int. 2017, 461, 80–101. [Google Scholar] [CrossRef]
- Zedeño, M.N.; Ballenger, J.A.M.; Murray, J.R. Landscape engineering and organizational complexity among late prehistoric Bison hunters of the Northwestern Plains. Curr. Anthropol. 2014, 55, 23–58. [Google Scholar] [CrossRef]
- Mack, R.N.; Thompson, J.N. Evolution in Steppe with few large, hooved mammals. Am. Nat. 1982, 119, 757–773. [Google Scholar] [CrossRef]
- Nabokov, P.; Loendorf, L. Restoring a Presence: American Indians and Yellowstone National Park; University of Oklahoma Press: Noman, OK, USA, 2004. [Google Scholar]
- Binnema, T.; Niemi, M. ‘Let the line be drawn now’: Wilderness, conservation, and the exclusion of Aboriginal people from Banff National Park in Canada. Environ. Hist. 2006, 11, 724–750. [Google Scholar] [CrossRef]
- Boyce, M.S. Natural regulation or control of nature? In The Greater Yellowstone Ecosystem: Redefining America’s Wilderness Heritage; Keiter, B., Boyce, M.S., Eds.; Yale University Press: New Haven, CN, USA, 1991; pp. 183–207. [Google Scholar]
- Wagner, F.H. Yellowstone’s Destabilized Ecosystem: Elk Effects, Science, and Policy Conflict; Oxford University Press: Oxford, MI, USA, 2006. [Google Scholar]
- Beschta, R.L.; Ripple, W.J.; Kauffman, J.B.; Painter, L.E. Bison limit ecosystem recovery in Northern Yellowstone. Food Webs 2020, 23, e00142. [Google Scholar] [CrossRef]
- Kay, C.E. The condition and trend of Aspen, Willows, and associated species on the Northern Yellowstone Range. Rangelands 2018, 40, 202–211. [Google Scholar] [CrossRef]
- Yonk, R.M.; Mosley, J.C.; Husby, P.O. Human influences on the Northern Yellowstone Range. Rangelands 2018, 40, 177–188. [Google Scholar] [CrossRef]
- White, C.A.; Olmsted, C.E.; Kay, C.E. Aspen, Elk, and Fire in the Rocky Mountain National Parks of North America. Wildl. Soc. Bull. 1998, 26, 449–462. [Google Scholar]
- Wyman, T. Grizzly Bear Predation on a Bull Bison in Yellowstone National Park. Ursus 2002, 13, 375–377. [Google Scholar]
- Carbyn, L.N.; Trottier, T. Descriptions of Wolf Attacks on Bison Calves in Wood Buffalo National Park. ARCTIC 1988, 41, 297–302. [Google Scholar] [CrossRef]
- Smith, D.W.; Mech, L.D.; Meagher, M.; Clark, W.E.; Jaffe, R.; Phillips, M.K.; Mack, J.A. Wolf-Bison Interactions in Yellowstone National Park. J. Mammal. 2000, 81, 1128–1135. [Google Scholar] [CrossRef]
- Kay, C.E. Were Native people keystone predators? A continuous-time analysis of wildlife observations made by Lewis and Clark. Can. Field Nat. 2007, 121, 1–16. [Google Scholar] [CrossRef]
- Wiken, E.; Nava, F.J.; Griffith, G. North American Terrestrial Ecoregions—Level III; Commission for Environmental Cooperation: Montreal, QC, Canada, 2011. [Google Scholar]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An ecoregion-based approach to protecting half the terrestrial realm. BioScience 2017, 67, 534–545. [Google Scholar] [CrossRef]
- Smith, J.R.; Hendershot, J.N.; Nova, N.; Daily, G.C. The biogeography of ecoregions: Descriptive power across regions and taxa. J. Biogeogr. 2020, 47, 1413–1426. [Google Scholar] [CrossRef]
- Vynne, C.; Gosling, J.; Maney, C.; Dinerstein, E.; Lee, A.T.L.; Burgess, N.D.; Fernández, N.; Fernando, S.; Jhala, H.; Jhala, Y.; et al. An ecoregion-based approach to restoring the world’s intact large mammal assemblages. Ecography 2022, 4, 1–12. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudık, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AL, Canada, 4–8 July 2004; pp. 662–665. [Google Scholar]
- Venne, S.; Currie, D.J. Can habitat suitability estimated from MaxEnt predict colonizations and extinctions? Divers. Distrib. 2021, 27, 873–886. [Google Scholar] [CrossRef]
- Bleyhl, B.; Sipko, T.; Trepet, S.; Bragina, E.; Leitão, P.J.; Radeloff, V.C.; Kuemmerle, T. Mapping seasonal european Bison habitat in the Caucasus Mountains to identify potential reintroduction sites. Biol. Conserv. 2015, 191, 83–92. [Google Scholar] [CrossRef]
- Kuemmerle, T.; Perzanowski, K.; Chaskovskyy, O.; Ostapowicz, K.; Halada, L.; Bashta, A.-T.; Kruhlov, I.; Hostert, P.; Waller, D.M.; Radeloff, V.C. European Bison habitat in the Carpathian Mountains. Biol. Conserv. 2010, 143, 908–916. [Google Scholar] [CrossRef]
- Su, H.; Bista, M.; Li, M. Mapping habitat suitability for Asiatic Black Bear and Red Panda in Makalu Barun National Park of Nepal from Maxent and GARP models. Sci. Rep. 2021, 11, 14135. [Google Scholar] [CrossRef]
- Esri Inc. ArcGIS Pro (Version 2.4.0); Esri Inc.: Redlands, CA, USA, 2019. [Google Scholar]
- Rockström, J.; Steffen, W.; Noone, K.; Persson, Å.; Chapin, F.S.; Lambin, E.F.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. A safe operating space for humanity. Nature 2009, 461, 472–475. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Oakleaf, J.R.; Theobald, D.M.; Baruch-Mordo, S.; Kiesecker, J. Global Human Modification of Terrestrial Systems; NASA Socioeconomic Data Applications Center SEDAC: New York, NY, USA, 2020. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Oakleaf, J.R.; Theobald, D.M.; Baruch-Mordo, S.; Kiesecker, J. Managing the Middle: A Shift in Conservation Priorities Based on the Global Human Modification Gradient. Glob. Chang. Biol. 2019, 25, 811–826. [Google Scholar] [CrossRef]
- Canada Centre for Remote Sensing (CCRS). Earth Sciences Sector, Natural Resources Canada. 2005 Land Cover of North America at 250 Meters; Canada Centre for Remote Sensing (CCRS): Ottawa, ON, Canada, 2013; p. 3.
- U.S. Geological Survey. North America Elevation 1-Kilometer Resolution GRID; U.S. Geological Survey: Reston, VA, USA, 2007.
- Poole, K.G.; Stuart-Smith, K.; Teske, I.E. Wintering strategies by Mountain Goats in interior mountains. Can. J. Zool. 2009, 87, 273–283. [Google Scholar] [CrossRef]
- Mahony, C.R.; Wang, T.; Hamann, A.; Cannon, A.J. A global climate model ensemble for downscaled monthly climate normals over North America. Int. J. Climatol. 2022, 42, 5871–5891. joc.7566. [Google Scholar] [CrossRef]
- Sheppard, A.H.C.; Hecker, L.J.; Edwards, M.A.; Nielsen, S.E. Determining the influence of snow and temperature on the movement rates of Wood Bison (Bison Bison Athabascae). Can. J. Zool. 2021, 99, 489–496. [Google Scholar] [CrossRef]
- Meagher, M. Range expansion by Bison of Yellowstone National Park. J. Mammal. 1989, 70, 670–675. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. Kuenm: An R package for detailed development of Ecological Niche Models using Maxent. PeerJ 2019, 7, 2–15. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Edwards, T.C.; Guisan, A.; McMahon, S.M.; Normand, S.; Thuiller, W.; Wüest, R.O.; Zimmermann, N.E.; Elith, J. What do we gain from simplicity versus complexity in Species Distribution Models? Ecography 2014, 37, 1267–1281. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking Receiver Operating Characteristic Analysis Applications in Ecological Niche Modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Anderson, R.P.; Lew, D.; Peterson, A.T. Evaluating Predictive Models of Species’ Distributions: Criteria for Selecting Optimal Models. Ecol. Model. 2003, 162, 211–232. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological Niche Modeling in Maxent: The Importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression: Hosmer/Applied Logistic Regression; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000; ISBN 9780471722144. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference; R Core Team R: Vienna, Austria, 2022. [Google Scholar]
- Lüdecke, D.; Ben-Shachar, M.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R package for assessment, comparison and testing of statistical Mmodels. JOSS 2021, 6, 3139. [Google Scholar] [CrossRef]
- RStudio Team RStudio. Integrated Development Environment for R 2022; RStudio Team RStudio: Boston, MA, USA, 2022. [Google Scholar]
- R Core Team, R. A Language and Environment for Statistical Computing 2022; R Core Team R: Vienna, Austria, 2022. [Google Scholar]
- Harper, W.L.; Elliot, J.P.; Hatter, I.; Schwantje, H. Management Plan for Wood Bison in British Columbia; Wildlife Bulletin; British Columbia, Ministry of Environment, Lands, and Parks, Wildlife Branch: Victoria, BC, Canada, 2000; p. 43. ISBN 0772642001. [Google Scholar]
- Plumb, G.E.; White, P.J.; Coughenour, M.B.; Wallen, R.L. Carrying capacity, migration, and dispersal in Yellowstone Bison. Biol. Conserv. 2009, 142, 2377–2387. [Google Scholar] [CrossRef]
- Mosley, J.C.; Mundinger, J.G. History and Status of Wild Ungulate Populations on the Northern Yellowstone Range. Rangelands 2018, 40, 189–201. [Google Scholar] [CrossRef]
- Laskin, D.N.; Watt, D.; Whittington, J.; Heuer, K. Designing a fence that enables free passage of wildlife while containing reintroduced Bison: A multispecies evaluation. Wildl. Biol. 2020, 2020, 1–14. [Google Scholar] [CrossRef]
- Heuer, K. Update to Stakeholders on Banff Bison Restoration Project 2021; Government of Canada: Banff, AB, Canada, 2021. [Google Scholar]
- Steenweg, R.; Hebblewhite, M.; Gummer, D.; Low, B.; Hunt, B. Assessing potential habitat and carrying capacity for reintroduction of Plains Bison (Bison Bison Bison) in Banff National Park. PLoS ONE 2016, 11, e0150065. [Google Scholar] [CrossRef]
- Rowe, M. Halfway-Sikanni Plains Bison Inventory; BC Ministry of Environment: Fort St. John, BC, Canada, 2006.
- COSEWIC. COSEWIC Assessment and Status Report on the Plains Bison (Bison Bison Bison) and the Wood Bison (Bison Bison Athabascae) in Canada; Committee on the Status of Endangered Wildlife in Canada: Ottawa, ON, Canada, 2013. [Google Scholar]
- Hoar, B.R. Does the Pink Mountain Region Contain Sufficient Habitat Suitable for Plains Bison in Summer and Winter? BC Ministry of Forests, Lands, Natural Resource Operations and Rural Development: Fort St. John, BC, Canada, 2021.
- Rowe, M.; Backmeyer, R. Etthithun Wood Bison Inventory; BC Ministry of Environment: Fort St. John, BC, Canada, 2006.
- Leverkus, S.E.R. Conservation of Biodiversity in Northern Canada through Ecological Processes and Cultural Landscapes. Ph.D. Dissertation, Oklahoma State University, Stillwater, OK, United States, 2015. [Google Scholar]
- Lewis, C.J.; Das Gupta, S. Wood Bison—Etthithun Herd Minimum Population Survey Results: 22–25 February 2021; British Columbia Ministry of Forests, Lands, Natural Resource Operations and Rural Development: Peace Region, BC, Canada, 2021; pp. 1–30. [Google Scholar]
- Rowe, M. Nordquist Wood Bison Inventory; Peace Region Technical Report; BC Ministry of Environment: Fort St. John, BC, Canada, 2007.
- Jung, T.S. Extralimital movements of reintroduced Bison (Bison Bison): Implications for potential range expansion and human-wildlife conflict. Eur. J. Wildl. Res. 2017, 63, 35. [Google Scholar] [CrossRef]
- Theissen, C. Peace Wood Bison Project: Annual Report 2009/2010; BC Ministry of Environment: Fort St. John, BC, Canada, 2010.
- Larter, N.C.; Allaire, D.G. History and Current Status of the Nahanni Wood Bison Population; File Report; Government of the Northwest Territories: Yellowknife, NT, Canada, 2007.
- Larter, N.C.; Allaire, D.G. Population Survey of the Nahanni Wood Bison Population; Manuscript Report; Environment and Natural Resources, Government of the Northwest Territories: Yellowknife, NT, Canada, 2013. [Google Scholar]
- Markel, C.; Clark, D. Developing policy alternatives for the management of Wood Bison (Bison Bison Athabascae) in Kluane National Park. North. Rev. 2012, 36, 53–75. [Google Scholar]
- Clark, D.A.; Workman, L.; Jung, T.S. Impacts of reintroduced bison on First Nations People in Yukon, Canada: Finding common ground through participatory research and social learning. Conserv. Soc. 2016, 14, 1–12. [Google Scholar] [CrossRef]
- Jung, T.S.; Czetwertynski, S.M.; Schmiegelow, F.K.A. Boreal Forest titans do not clash: Low overlap in winter habitat selection by Moose (Alces Americanus) and reintroduced Bison (Bison Bison). Eur. J. Wildl. Res. 2018, 64, 25. [Google Scholar] [CrossRef]
- Jung, T.S.; Hegel, T.M.; Stotyn, S.A.; Czetwertynski, S.M. Co-occurrence of reintroduced and resident ungulates on a shared winter range in Northwestern Canada. Écoscience 2015, 22, 7–16. [Google Scholar] [CrossRef]
- Fischer, L.A.; Gates, C.C. Competition potential between sympatric Woodland Caribou and Wood Bison in Southwestern Yukon, Canada. Can. J. Zool. 2005, 83, 1162–1173. [Google Scholar] [CrossRef]
- White, C.A.; Langemann, E.G.; Gates, C.C.; Kay, C.E.; Shury, T.; Hurd, T.E. Plains Bison restoration in the Canadian Rocky Mountains: Ecological and management considerations. In Proceedings of the George Wright Biannual Conference on Research and Resource Management in National Parks and on Public Lands 11, Hancock, MI, USA, 16–20 April 2001; George Wright Society: Washington, DC, USA, 2001; Volume 11, pp. 152–160. [Google Scholar]
- Mosley, J.C.; Fidel, J.; Hunter, H.E.; Husby, P.O.; Kay, C.E.; Mundinger, J.G.; Yonk, R.M. An ecological assessment of the Northern Yellowstone Range: Synthesis and call to action. Rangelands 2018, 40, 224–227. [Google Scholar] [CrossRef]
- Cronin, M.A.; MacNeil, M.D.; Vu, N.; Leesburg, V.; Blackburn, H.D.; Derr, J.N. Genetic Variation and Differentiation of Bison (Bison Bison) Subspecies and Cattle (Bos Taurus) Breeds and Subspecies. J. Hered. 2013, 104, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.A.; Strobeck, C. Genetic Variation within and Relatedness among Wood and Plains Bison Populations. Genome 1999, 42, 14. [Google Scholar] [CrossRef]
- Geist, V. Buffalo Nation: History and Legend of the North American Bison; Fifth House: Saskatoon, SK, Canada, 1996. [Google Scholar]
- Laliberte, A.S.; Ripple, W.J. Wildlife encounters by Lewis and Clark: A spatial analysis of interactions between Native Americans and wildlife. BioScience 2003, 53, 994–1003. [Google Scholar] [CrossRef]
- Binnema, T. Common and Contested Ground: A Human and Environmental History of the Northwestern Plains; University of Oklahoma Press: Norman, OK, USA, 2001. [Google Scholar]
- Beaudoin, A.B. A.B. A Bison’s view of landscape and the paleoenvironment. In Bison and People on the North American Great Plains; Cunfer, G., Waiser, W.A., Eds.; Texas A&M University Press: College Station, TX, USA, 2016; pp. 51–89. [Google Scholar]
- Binnema, T. A Fur trade historian’s view of seasonal Bison movements on the Northern Great Plains. In Bison and People on the American Great Plains: A Deep Environmental History; Cunfer, G., Waiser, W.A., Eds.; Texas A&M University Press: College Station, TX, USA, 2016; pp. 159–177. [Google Scholar]
- Cunfer, G.; Waiser, W.A. (Eds.) Bison and People on the North American Great Plains: A Deep Environmental History, 1st ed.; Connecting the Greater West; Texas A&M University Press: College Station, TX, USA, 2016. [Google Scholar]
- Carlson, K.; Bement, L. Organization of bison hunting at the Pleistocene/Holocene transition on the Plains of North America. Quat. Int. 2013, 297, 93–99. [Google Scholar] [CrossRef]
- Bayham, J.; Cole, K.E.; Bayham, F.E. Social boundaries, resource depression, and conflict: A bioeconomic model of the Intertribal Buffer Zone. Quat. Int. 2017, 518, 69–82. [Google Scholar] [CrossRef]
- Moodie, D.W.; Ray, A.J. Buffalo migrations on the Canadian Plains. Plains Anthropol. 1976, 21, 45–46. [Google Scholar] [CrossRef]
- Morgan, R.G. Bison movement patterns on the Canadian Plains: An ecological analysis. Plains Anthropol. 1980, 25, 143–160. [Google Scholar] [CrossRef]
- Geist, V. Phantom Subspecies: The Wood Bison (Bison Bison “Athabascae” Rhoads 1897) Is Not a Valid Taxon, but an Ecotype. Arctic 1991, 44, 283–300. [Google Scholar] [CrossRef]
- Kay, C.E. Aboriginal overkill: The role of Native Americans in structuring Western Ecosystems. Hum. Nat. 1994, 5, 359–398. [Google Scholar] [CrossRef]
- Martin, P.S.; Szuter, C.R. Megafauna of the Columbia Basin, 1800–1840, Lewis and Clark in a game Sink. In Northwest Lands, Northwest Peoples: Readings in Environmental History; Goble, D.D., Hirt, P.W., Eds.; University of Washington Press: Seattle, WA, USA, 1999. [Google Scholar]
- Arthur, G. An introduction to the ecology of early historic communal Bison hunting among the Northern Plains Indians. Can. Natl. Mus. Can. Natl. Mus. Man Mercury Ser. Archaeol. Surv. Can. Pap. 1975, 37, 144. [Google Scholar]
- Brink, J. Imagining Head-Smashed-In: Aboriginal Buffalo Hunting on the Northern Plains; Athabasca University Press: Edmonton, AL, Canada, 2008. [Google Scholar]
- White, C.A. Historical Ecology of Bison Movement Corridors, Western Cordillera, North America: A Cross Watershed Comparison; CW and Associates: Canmore, AB, Canada, 2018. [Google Scholar]
- Grayson, D.K. Holocene Bison in the Great Basin, Western USA. Holocene 2006, 16, 913–925. [Google Scholar] [CrossRef]
- Lupo, K.; Schmitt, D.N. On Late Holocene Variability in Bison Populations in the Northeastern Great Basin. J. Calif. Great Basin Anthropol. 1997, 19, 55–69. [Google Scholar]
- Lyman, R.L. Late-Quaternary Diminution and Abundance of Prehistoric Bison (Bison Sp.) in Eastern Washington State, USA. Quat. Res. 2004, 62, 76–85. [Google Scholar] [CrossRef]
- Hunn, E.S.; Selam, J. Nch’i-Wana, “The Big River”: Mid-Columbia Indians and Their Land; University of Washington Press: Seattle, WA, USA, 1990. [Google Scholar]
- Prentiss, W.C.; Kuijt, I. Complex Hunter Gatherers Evolution Organization of Prehistoric Communities Plateau of Northwestern North America; Utah State University Press: Salt Lake City, UT, USA, 2004. [Google Scholar]
- Prentiss, A.M.; Kuijt, I. People of the Middle Fraser Canyon an Archaeological History; University of British Columbia Press: Vancouver, BC, Canada, 2014. [Google Scholar]
- Tremayne, A.H.; Winterhalder, B. Large mammal biomass predicts the changing distribution of hunter-gatherer settlements in Mid-Late Holocene Alaska. J. Anthropol. Archaeol. 2017, 45, 81–97. [Google Scholar] [CrossRef]
- Doering, B.N. Subarctic landscape adaptations and paleodemography: A 14,000-year history of climate change and human settlement in Central Alaska and Yukon. Quat. Sci. Rev. 2021, 268, 107139. [Google Scholar] [CrossRef]
- Klein, L. Demystifying the opposition: The Hudson’s Bay Company and the Tlingit. Arct. Anthropol. 1987, 24, 101–114. [Google Scholar]
- Cloud, J.; Williams, L. The Tlingit Map of 1869. Exped. Mag. 2012, 54, 10–18. [Google Scholar]
- Christianson, A.C.; Sutherland, C.R.; Moola, F.; Gonzalez Bautista, N.; Young, D.; MacDonald, H. Centering Indigenous voices: The role of fire in the Boreal Forest of North America. Curr. For. Rep. 2022, 8, 257–276. [Google Scholar] [CrossRef]
- Joseph, L.; Turner, N.J. “The old foods are the new foods!”: Erosion and revitalization of Indigenous Food Systems in Northwestern North America. Front. Sustain. Food Syst. 2020, 4, 596237. [Google Scholar] [CrossRef]
- Peacock, S.M.; Turner, N.J. ‘Just like a garden,’ traditional resource management and biodiversity conservation on the Interior Plateau of British Columbia. In Biodiversity and Native America; Minnis, P.E., Elisens, W.J., Eds.; University of Oklahoma Press: Norman, OK, USA, 2000; pp. 133–179. [Google Scholar]
- Fortin, D.; Fryxell, J.M.; O’Brodovich, L.; Frandsen, D. Foraging ecology of Bison at the landscape and plant community levels: The applicability of energy maximization principles. Oecologia 2003, 134, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Harvey, L.; Fortin, D. Spatial heterogeneity in the strength of plant-herbivore interactions under predation risk: The tale of Bison foraging in Wolf country. PLoS ONE 2013, 8, 73324. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M. Seasonal patterns of Bison diet across climate gradients in North America. Sci. Rep. 2021, 11, 6829. [Google Scholar] [CrossRef] [PubMed]
- Hecker, L.J.; Coogan, S.C.P.; Nielsen, S.E.; Edwards, M.A. Latitudinal and seasonal plasticity in American Bison Bison Bison diets. Mam. Rev. 2021, 51, 193–206. [Google Scholar] [CrossRef]
- Holt, R.D.; Bonsall, M.B. Apparent competition. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 447–471. [Google Scholar] [CrossRef]
- Jung, T.S. Investigating local concerns regarding large mammal restoration: Group size in a growing population of reintroduced Bison (Bison Bison). Glob. Ecol. Conserv. 2020, 24, 01303. [Google Scholar] [CrossRef]
- Packer, C.; Holt, R.D.; Hudson, P.J.; Lafferty, K.D.; Dobson, A.P. Keeping the herds healthy and alert: Implications of predator control for infectious disease. Ecol. Lett. 2003, 6, 797–802. [Google Scholar] [CrossRef]
- Schusler, T.M.; Chase, L.C.; Decker, D.J. Community-based comanagement: Sharing responsibility when tolerance for wildlife is exceeded. Hum. Dimens. Wildl. 2000, 5, 34–49. [Google Scholar] [CrossRef]
- Redford, K.H.; Aune, K.; Plumb, G. Hope is a Bison: Editorial. Conserv. Biol. 2016, 30, 689–691. [Google Scholar] [CrossRef]
- Thornton, T.; Deur, D.; Kitka, H. Cultivation of Salmon and other marine resources on the Northwest Coast of North America. Hum. Ecol. 2015, 43, 189–199. [Google Scholar] [CrossRef]
- Kimmerer, R.W.; Lake, F.K. Maintaining the mosaic—The role of Indigenous burning in land management. J. For. 2001, 99, 36–41. [Google Scholar] [CrossRef]
- Hoffman, K.M.; Davis, E.L.; Wickham, S.B.; Schang, K.; Johnson, A.; Larking, T.; Lauriault, P.N.; Quynh Le, N.; Swerdfager, E.; Trant, A.J. Conservation of Earth’s biodiversity is embedded in Indigenous fire stewardship. Proc. Natl. Acad. Sci. USA 2021, 118, e2105073118. [Google Scholar] [CrossRef] [PubMed]
- White, C.A.; Perrakis, D.D.B.; Kafka, V.G.; Ennis, T. Burning at the edge: Integrating biophysical and eco-cultural fire processes in Canada’s Parks and Protected Areas. Fire Ecol. 2011, 7, 74–106. [Google Scholar] [CrossRef]
- Reid, A.J.; Eckert, L.E.; Lane, J.; Young, N.; Hinch, S.G.; Darimont, C.T.; Cooke, S.J.; Ban, N.C.; Marshall, A. “Two-Eyed Seeing”: An Indigenous framework to transform fisheries research and management. Fish Fish 2021, 22, 243–261. [Google Scholar] [CrossRef]
- Artelle, K.A.; Zurba, M.; Bhattacharyya, J.; Chan, D.E.; Brown, K.; Housty, J.; Moola, F. Supporting resurgent Indigenous-led governance: A nascent mechanism for just and effective conservation. Biol. Conserv. 2019, 240, 108284. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictors | Estimate | Std. Error | p |
|---|---|---|---|
| (Intercept) | −2.76 | 0.192 | <0.001 |
| MaxEnt habitat suitability | 1.33 | 0.0575 | <0.001 |
| Human abundance index | −0.224 | 0.0404 | <0.001 |
| Grizzly bear abundance index | 0.0827 | 0.0291 | 0.0045 |
| Wolf abundance index | 0.201 | 0.0361 | <0.001 |
| Location and Ecotype [References] | Estimated Range (km2) | Population Estimate (Year) | Estimated Carrying Capacity (K, Bison/km2) | Observed Annual Rate of Increase (%) at ≤50% K | Remarks |
|---|---|---|---|---|---|
| Yellowstone, WY, MT Plains Bison [22,62,63] | 9400 | 4500 (2018) | 0.405 | 12 | All native predators except humans. K determined by habitat quality and demographics, population c. 2010 > 75% of K (counts of ~4000), large impacts on grasslands, woody plants, and hydrologic indicators, emigration rate out of park is increasing. |
| Banff, AB Plains Bison [64,65,66] | 1200 | 80 (2020) | 0.480 | 30 | All native predators except humans. K determined by habitat quality, current increase rate very high due to high female-to-male ratio of the founder herd, current population < 100 but increasing rapidly. |
| Halfway-Sikanni, BC Plains Bison [67,68,69] | 3200 | 1013 (2014) | 0.500 | 10–38 | All native predators present, including humans. A total of 48 plains bison escaped in 1971, population ~1300 in 2006, K socially determined to minimize range expansion. |
| Etthithun, BC & AB Wood Bison [61,68,70,71,72] | 5000 | 193 (2010) | No information | 15–20 | All native predators present. First Nations allocated permits to take bison to discourage range expansion due to conflicts with industry and agriculture. |
| Nordquist, BC Wood Bison [61,68,71,73,74,75] | 1400 | 124–142 (2010) | No information | <5% | All native predators present, including humans. A total of 49 bison released in 1995 at Aline Lake, by 2010, range mainly along 170 km Alaska Highway, population limited by traffic accidents. |
| Nahanni, NT & BC Wood Bison [76,77] | 11,700 | 431 (2011) | No information | ~8% | All native predators present, including humans. Slow growth since establishment, limiting factors appear to be drowning, traffic accidents, hunting, and perhaps tooth wear due to diet high in silica. |
| Aishihik, YT Wood Bison [78,79,80,81,82] | 11,000 | 1106–1385 (2011) | 0.125 | 10–20 | All native predators present. Socially determined carrying capacity to minimize conflicts with Indigenous land uses and competition with other species. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farr, J.J.; White, C.A. Buffalo on the Edge: Factors Affecting Historical Distribution and Restoration of Bison bison in the Western Cordillera, North America. Diversity 2022, 14, 937. https://doi.org/10.3390/d14110937
Farr JJ, White CA. Buffalo on the Edge: Factors Affecting Historical Distribution and Restoration of Bison bison in the Western Cordillera, North America. Diversity. 2022; 14(11):937. https://doi.org/10.3390/d14110937
Chicago/Turabian StyleFarr, Jonathan James, and Clifford A. White. 2022. "Buffalo on the Edge: Factors Affecting Historical Distribution and Restoration of Bison bison in the Western Cordillera, North America" Diversity 14, no. 11: 937. https://doi.org/10.3390/d14110937
APA StyleFarr, J. J., & White, C. A. (2022). Buffalo on the Edge: Factors Affecting Historical Distribution and Restoration of Bison bison in the Western Cordillera, North America. Diversity, 14(11), 937. https://doi.org/10.3390/d14110937