

Composition and Distribution of Plankton Communities in the Atlantic Sector of the Southern Ocean

 , , ,
, , ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
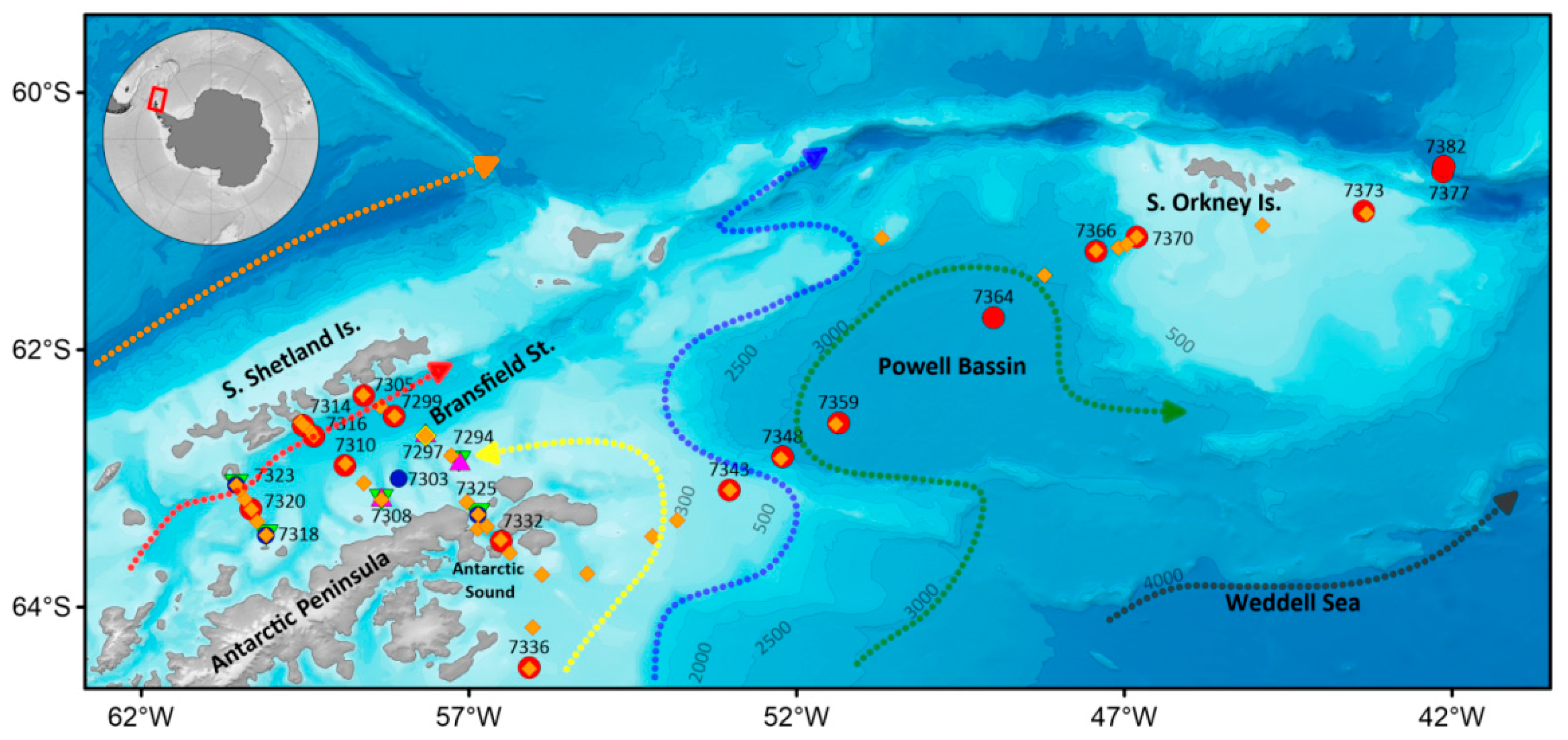
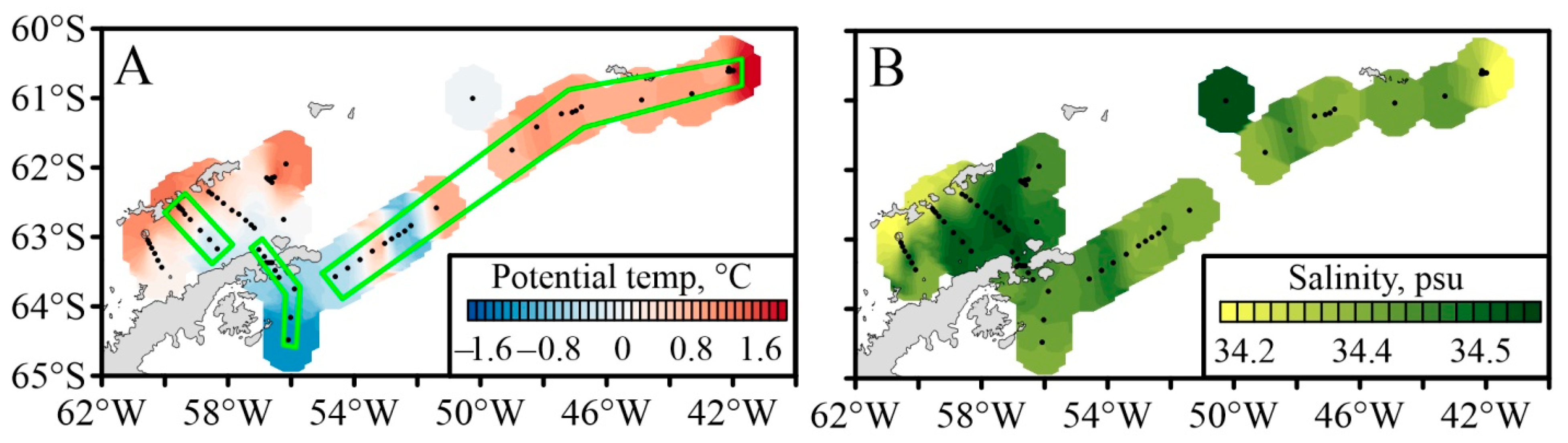
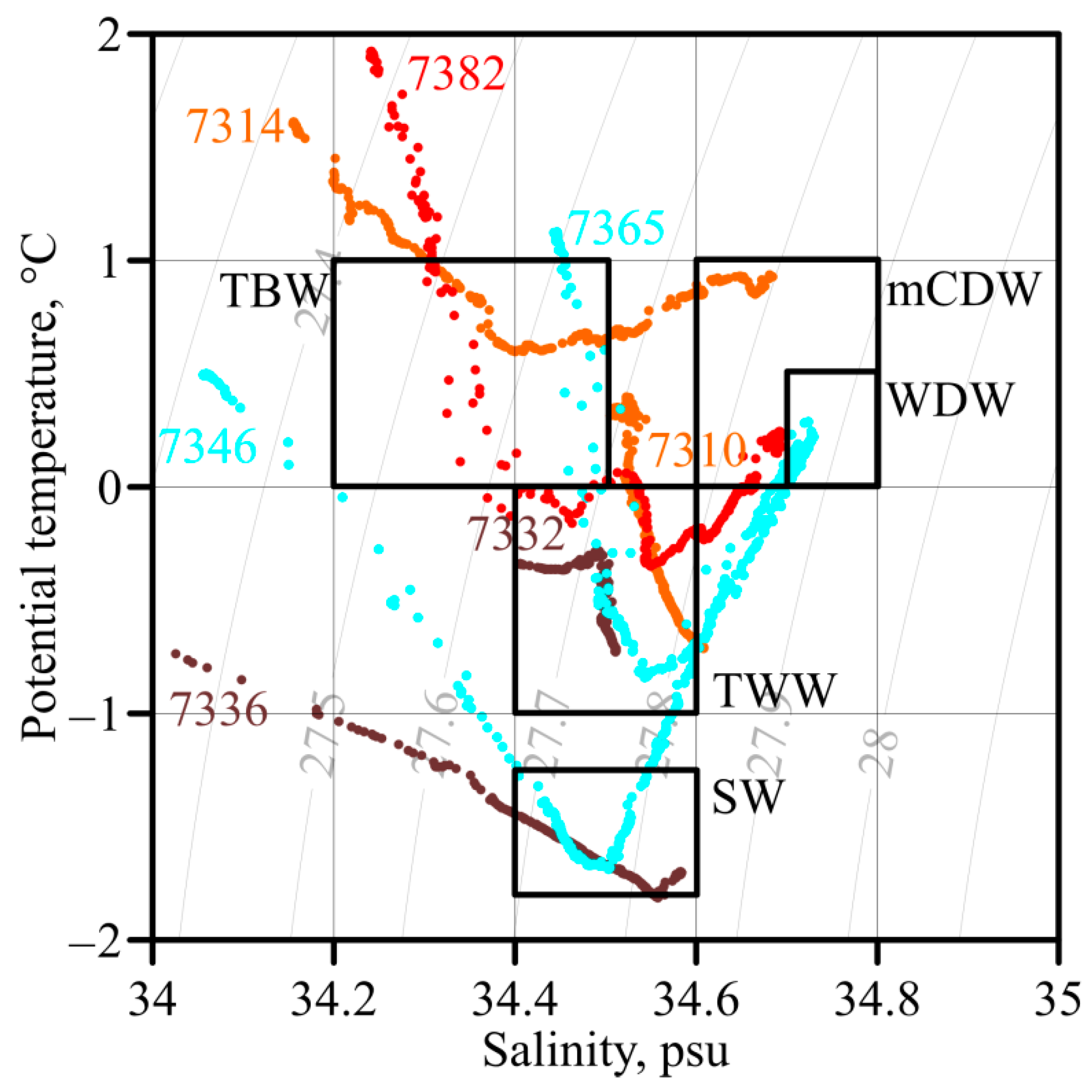
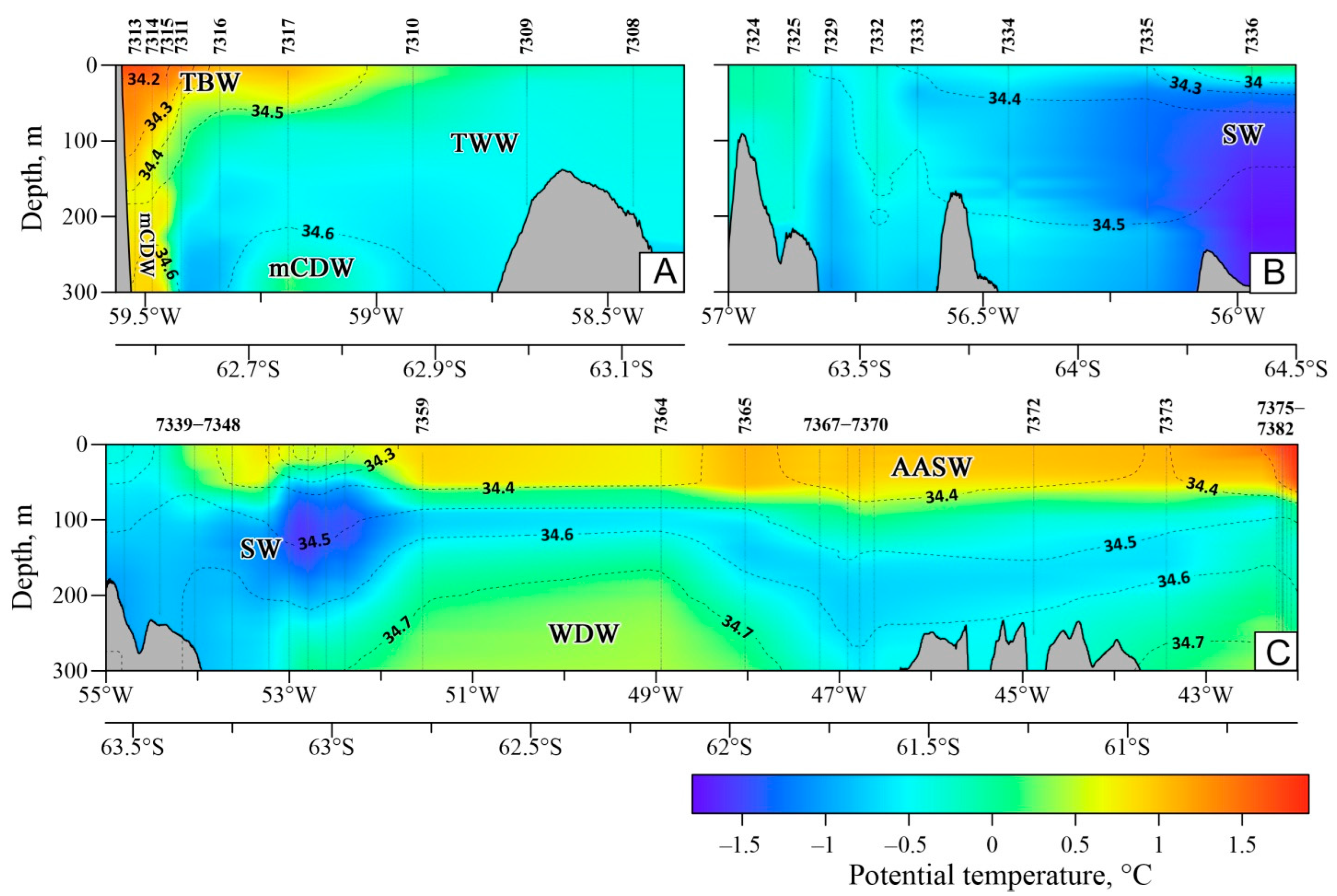
2.1. Oceanographic Measurements
2.2. Biological Sampling
2.3. Taxonomic Identification
2.4. Data Analysis
2.4.1. Phytoplankton Community Analysis
2.4.2. Mesozooplankton Community Analysis
2.4.3. Macrozooplankton Community Analysis
2.4.4. Ichthyoplankton Community Analysis
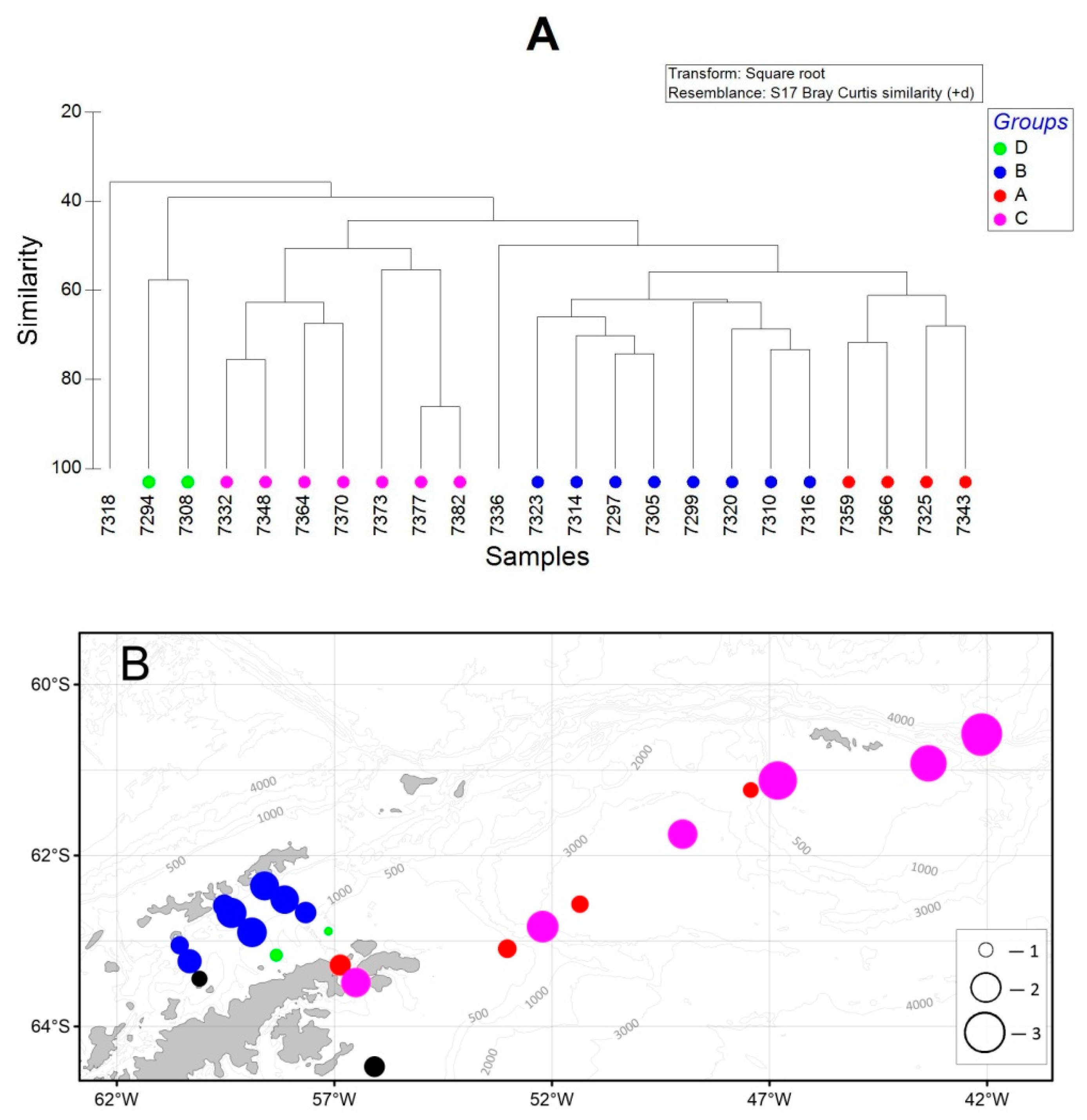
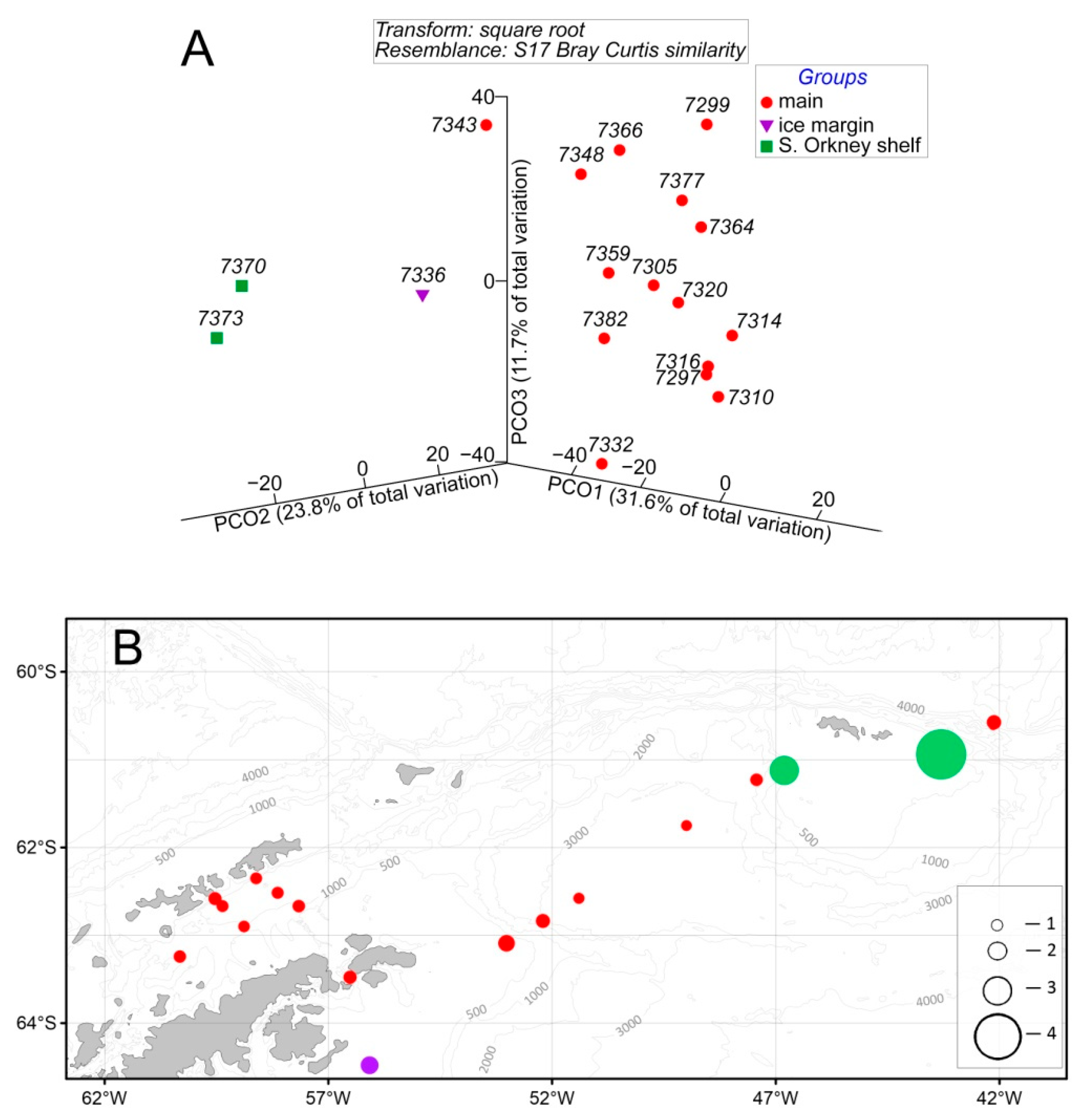
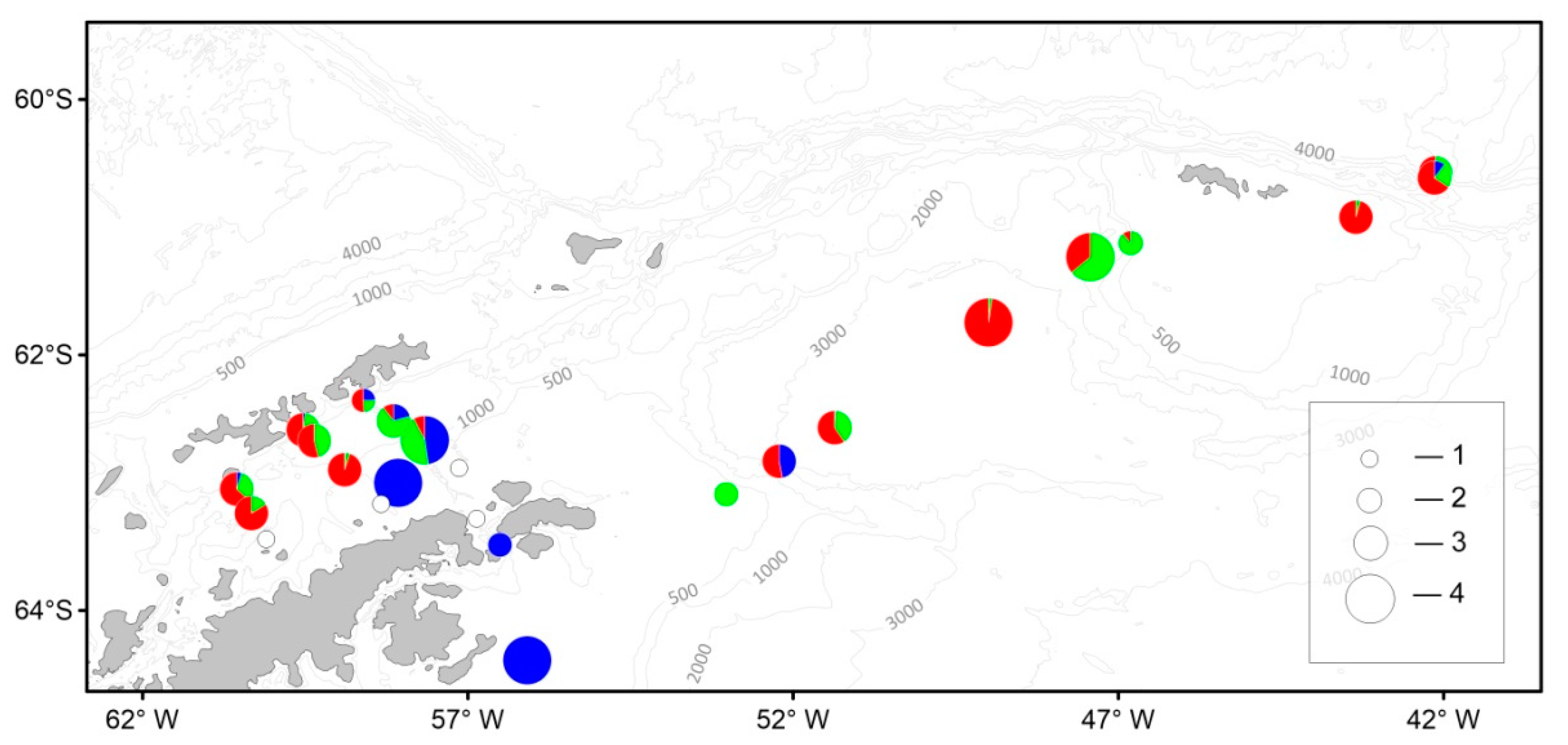
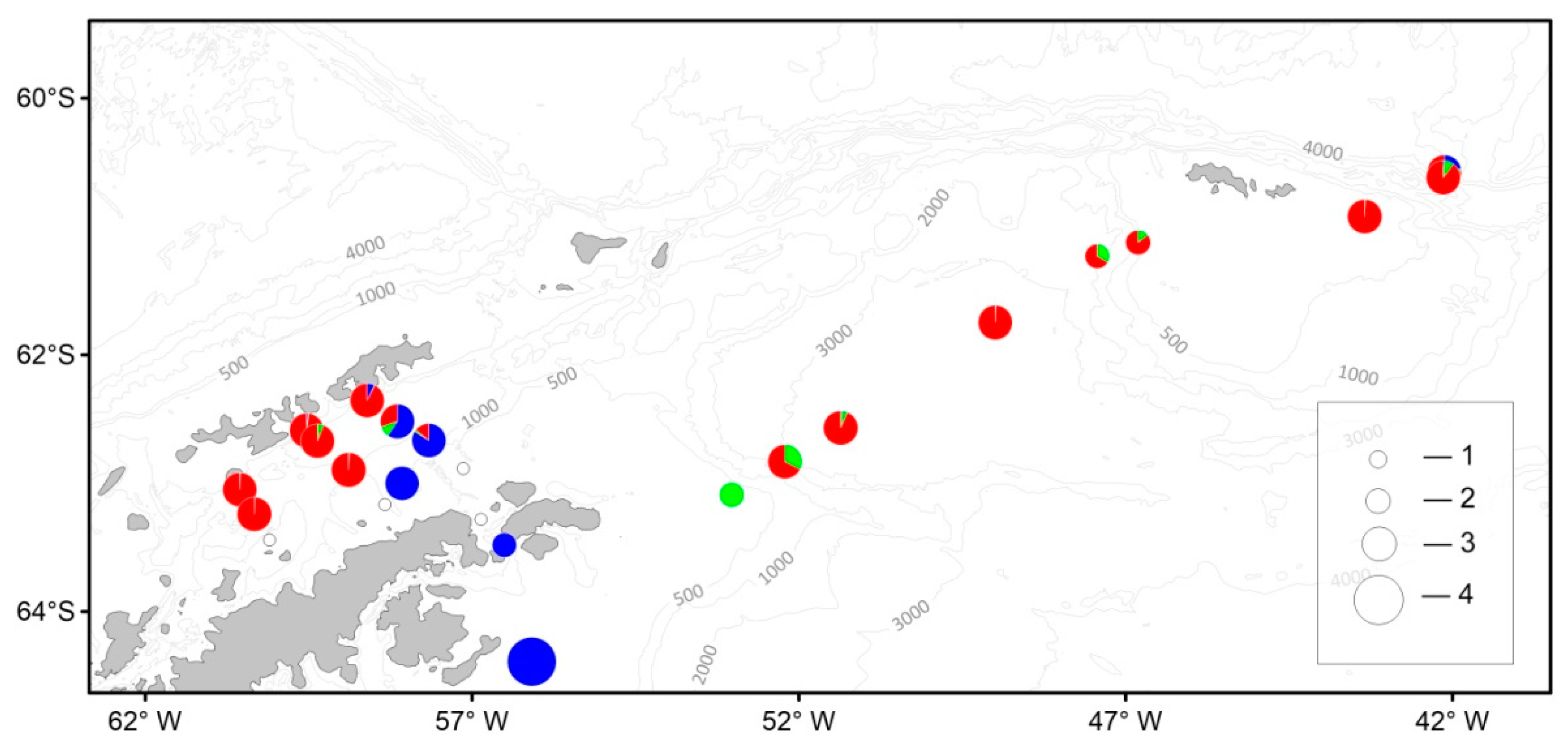
3. Results
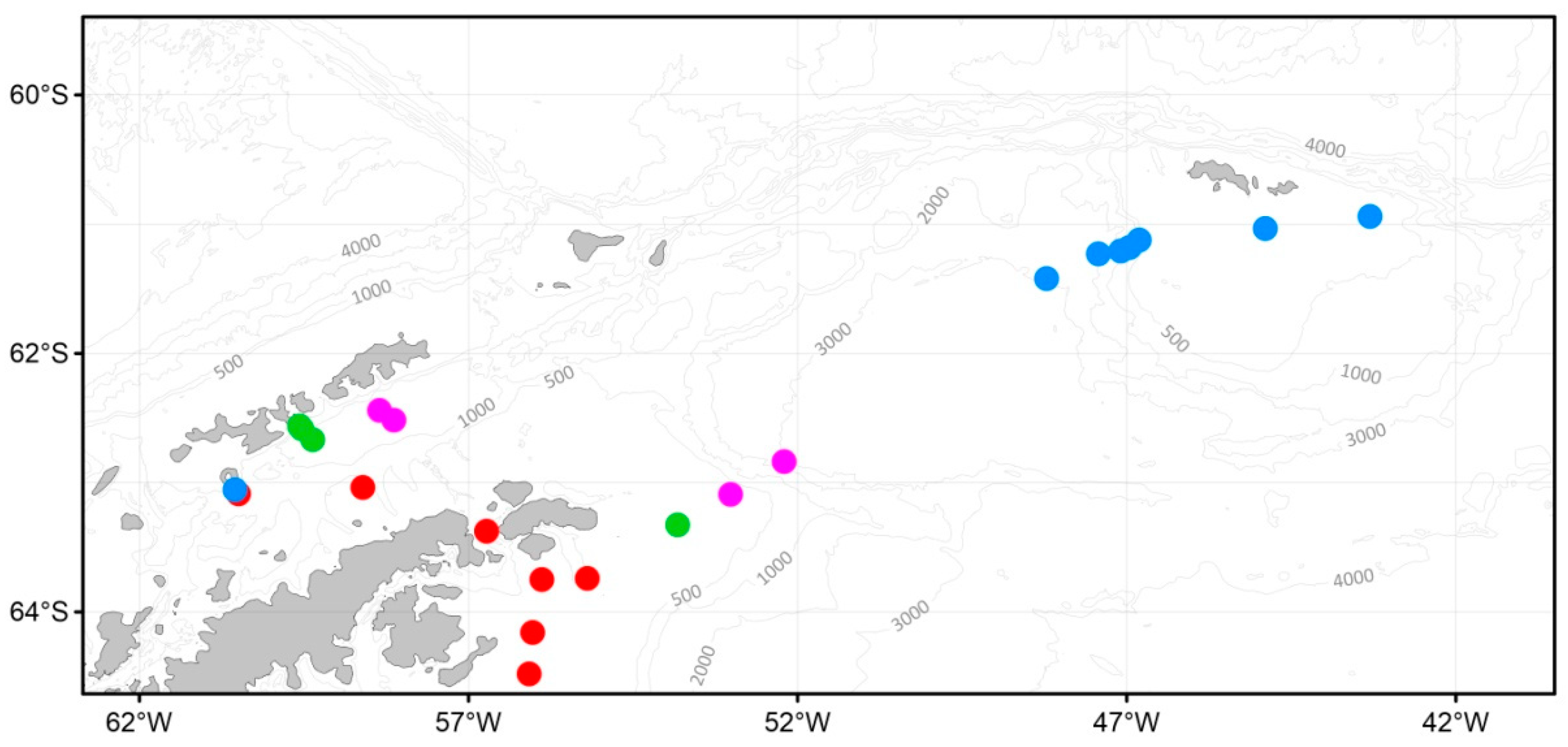
3.1. Oceanographic Characteristics
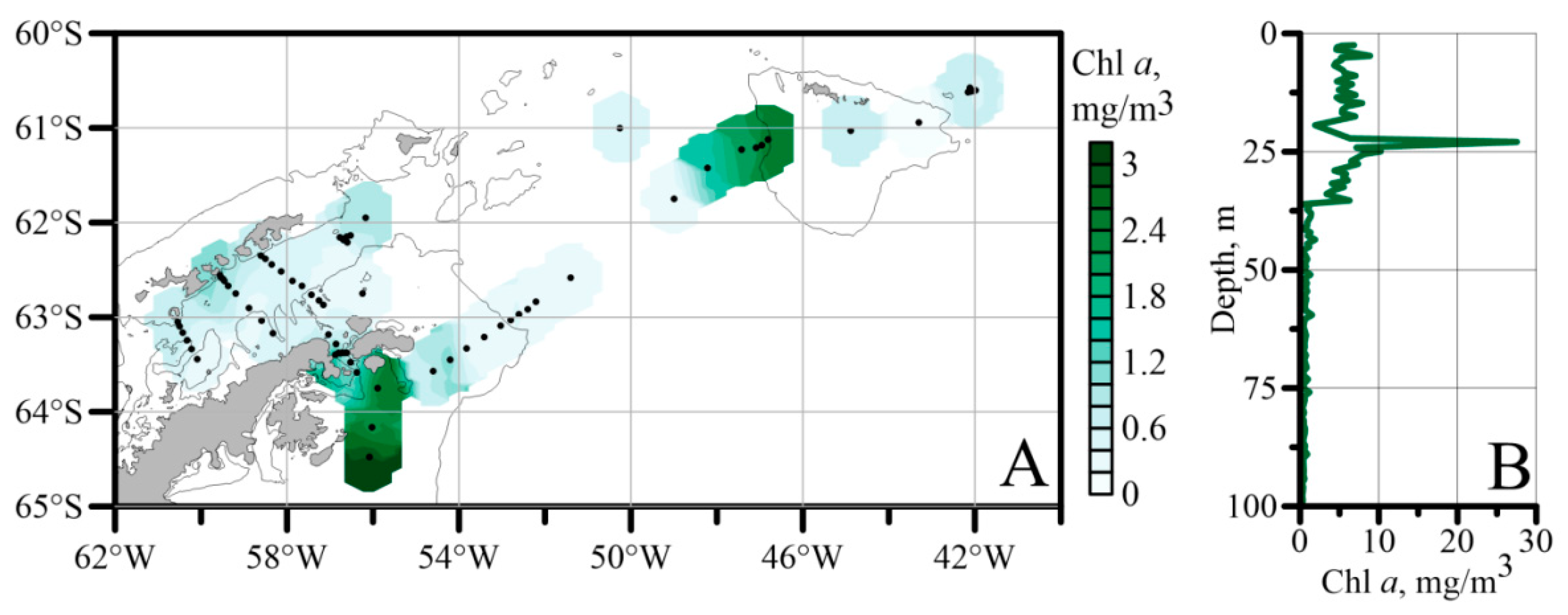
3.2. Phytoplankton
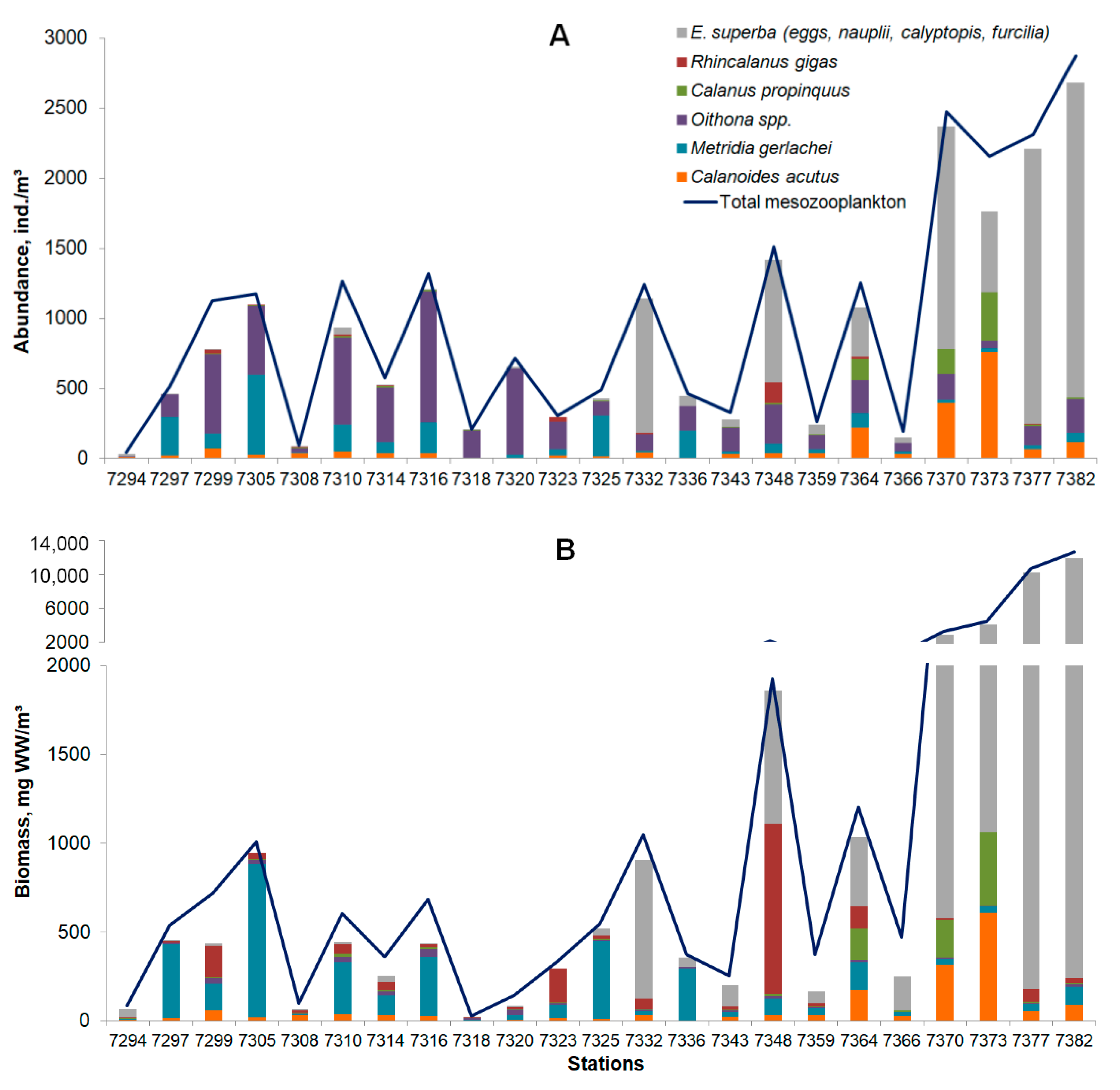
3.3. Mesozooplankton
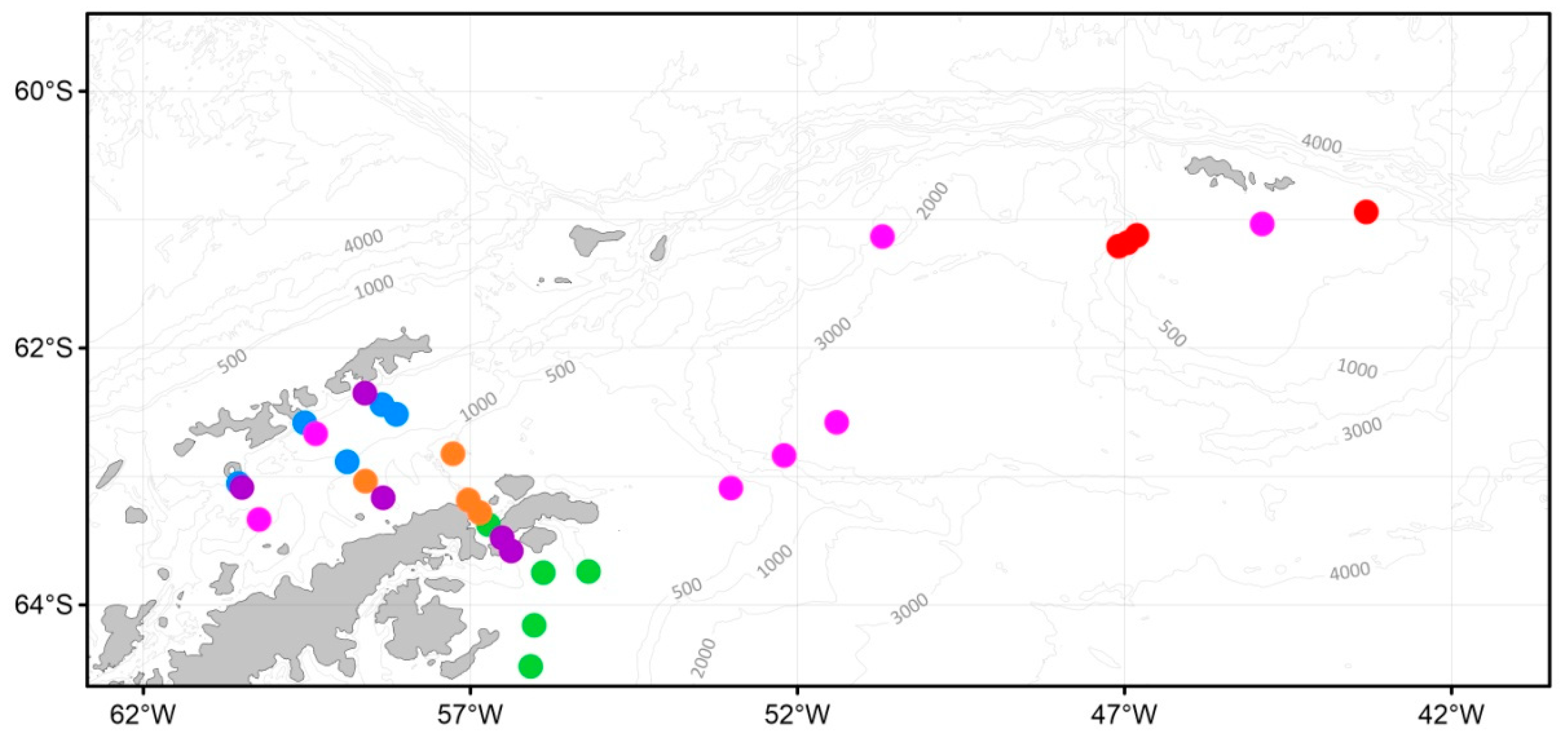
3.4. Macrozooplankton
3.5. Ichthyoplankton
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anisimov, O.A.; Vaughan, D.G.; Callaghan, T.V.; Furgal, C.; Marchant, H.; Prowse, T.D.; Vilhjalmsson, H.; Walsh, J.E. 2007: Polar regions (Arctic and Antarctic). In Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 653–685. [Google Scholar]
- Stammerjohn, S.E.; Martinson, D.G.; Smith, R.C.; Yuan, X.; Rind, D. Trends in Antarctic Annual Sea Ice Retreat and Advance and Their Relation to El Nino–Southern Oscillation and Southern Annular Mode Variability. J. Geophys. Res. 2008, 113, C03S90. [Google Scholar] [CrossRef]
- Meredith, M.P.; King, J.C. Rapid climate change in the ocean west of the Antarctic Peninsula during the second half of the 20th century. Geophys. Res. Lett. 2005, 32, L19604. [Google Scholar]
- Voronina, N.M. Comparative abundance and distribution of major filter–feeders in the Antarctic pelagic zone. J. Mar. Syst. 1998, 17, 375–390. [Google Scholar] [CrossRef]
- Hewes, C.D.; Reiss, C.S.; Holm-Hansen, O.A. Quantitative analysis of sources for summertime phytoplankton variability over 18 years in the South Shetland Islands (Antarctica) region. Deep-Sea Res. II 2009, 56, 1230–1241. [Google Scholar] [CrossRef]
- Loeb, V.J.; Santora, J.A. Population dynamics of Salpa thompsoni near the Antarctic Peninsula: Growth rates and interannual variations in reproductive activity (1993–2009). Prog. Oceanogr. 2012, 96, 93–107. [Google Scholar] [CrossRef]
- Garcia, M.D.; Dutto, M.S.; Chazarreta, C.J.; Berasategui, A.A.; Schoss, I.R.; Hoffmeyer, M.S. Micro- and mesozooplankton successions in the Antarctic coastal environment during a warm year. PLoS ONE 2020, 15, 5. [Google Scholar]
- Pakhomov, E.A.; Froneman, P.W.; Perissinotto, R. Salp/krill interactions in the Southern Ocean: Spatial segregation and implications for the carbon flux. Deep-Sea Res. II 2002, 49, 1881–1907. [Google Scholar] [CrossRef]
- Rintoul, S.R.; Sparrow, M.; Meredith, M.P.; Wadley, V.; Speer, K.; Hofmann, E.; Summerhayes, C.; Urban, E.; Bellerby, R.; Ackley, S. The Southern Ocean Observing System: Initial Science and Implementation Strategy; SCAR and SCOR: Cambridge, UK, 2012; 74p, ISBN 978-0-948277-27-6. [Google Scholar]
- Trathan, P.N.; Hill, S.L. The importance of krill predation in the Southern Ocean. In The Biology of Antarctic Krill: Euphausia Superba Dana 1850, Advances in Polar Ecology; Siegel, V., Ed.; Springer: Cham, Switzerland, 2016; pp. 321–350. [Google Scholar] [CrossRef]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate change and marine plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]
- Edwards, M.; Bresnan, E.; Cook, K.; Heath, M.; Helaouet, P.; Lynam, C.; Raine, R.; Widdicombe, C. Impacts of climate change on plankton. MCCIP Sci. Rev. 2013, 2013, 98–112. [Google Scholar]
- Behrenfeld, M.J. Climate–mediated dance of the plankton. Nat. Clim. Chang. 2014, 4, 880–887. [Google Scholar] [CrossRef]
- Siegel, V. Introducing Antarctic Krill Euphausia superba Dana, 1850. In Biology and Ecology of Antarctic Krill; Siegel, V., Ed.; Advances in Polar Ecology; Springer: Cham, Switzerland, 2016; pp. 1–19. [Google Scholar] [CrossRef]
- McBride, M.M.; Dalpadado, P.; Drinkwater, K.F.; Godø, O.R.; Hobday, A.J.; Hollowed, A.B.; Kristiansen, T.; Murphy, E.J.; Ressler, P.H.; Subbey, S.; et al. Krill, climate, and contrasting future scenarios for Arctic and Antarctic fisheries. ICES J. Mar. Sci. 2014, 71, 1934–1955. [Google Scholar] [CrossRef]
- Yu, L.; Zhong, S.; Vihma, T.; Sui, C.; Sun, B. Sea ice changes in the Pacific sector of the Southern Ocean in austral autumn closely associated with the negative polarity of the South Pacific Oscillation. Geophys. Res. Lett. 2021, 48, e2021GL092409. [Google Scholar] [CrossRef]
- Pakhomov, E.; Pshenichnov, L.; Krot, A.; Paramonov, V.; Slypko, I.; Zabroda, P. Zooplankton distribution and community structure in the Pacific and Atlantic Sectors of the Southern Ocean during austral summer 2017–2018: A Pilot Study Conducted from Ukrainian Long–Liners. J. Mar. Sci. Eng. 2020, 8, 488. [Google Scholar] [CrossRef]
- Atkinson, A.; Siegel, V.; Pakhomov, E.; Rothery, P. Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature 2004, 432, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.J.; Watkins, J.L.; Trathan, P.; Reid, K.; Meredith, M.P.; Thorpe, S.E.; Fleming, A.H. Spatial and temporal operation of the Scotia Sea ecosystem: A review of large-scale links in a krill centred food web. J. R. Stat. Soc. Ser. B 2007, 362, 113–148. [Google Scholar] [CrossRef]
- Henschke, N.; Pakhomov, E.; Groeneveld, J.; Meyer, B. Modelling the life cycle of Salpa thompsoni. Ecol. Model. 2018, 387, 17–26. [Google Scholar] [CrossRef]
- Loeb, V.; Siegel, V.; Holm-Hansen, O.; Hewitt, R.; Fraser, W.; Trivelpiece, W.; Trivelpiece, S. Effects of sea–ice extent and krill or salp dominance on the Antarctic food web. Nature 1997, 387, 897–900. [Google Scholar] [CrossRef]
- Loeb, V.J.; Santora, J.A. Climate variability and spatio temporal dynamics of five Southern Ocean krill species. Prog. Oceanogr. 2015, 134, 93–122. [Google Scholar] [CrossRef]
- Steinberg, D.K.; Ruck, K.E.; Gleiber, M.R.; Garzio, L.M.; Cope, J.S.; Bernard, K.S.; Stammerjohn, S.; Schofield, O.M.E.; Quetin, L.B.; Ross, R.M. Long-term (1993–2013) changes in macrozooplankton off the Western Antarctic Peninsula. Deep-Sea Res. I 2015, 101, 54–70. [Google Scholar] [CrossRef]
- Murphy, E.J.; Thorpe, S.E.; Tarling, G.A.; Watkins, J.L.; Fielding, S.; Underwood, P. Restricted regions of enhanced growth of Antarctic krill in the circumpolar Southern Ocean. Sci. Rep. 2017, 7, 6963. [Google Scholar] [CrossRef]
- McBride, M.M.; Stokke, O.S.; Renner, A.H.; Krafft, B.A.; Bergstad, O.A.; Biuw, M.; Lowther, A.D.; Stiansen, J.E. Antarctic krill Euphausia superba: Spatial distribution, abundance, and management of fisheries in a changing climate. Mar. Ecol. Prog. Ser. 2021, 668, 185–214. [Google Scholar] [CrossRef]
- Ross, R.M.; Quetin, L.B.; Martinson, D.G.; Iannuzzi, R.A.; Stammerjohn, S.E.; Smith, R.C. Palmer LTER: Patterns of distribution of five dominant zooplankton species in the epipelagic zone west of the Antarctic Peninsula, 1993–2004. Deep-Sea Res. II 2008, 55, 2086–2105. [Google Scholar] [CrossRef]
- Hereu, C.M.; Suarez-Morales, E.; Lavaniegos, B.E. Record of the rare oceanic salp Helicosalpa komaii (Tunicata: Thaliacea: Salpida) in the Northeast Pacific. Rev. Mex. Biodivers. 2014, 85, 624–629. [Google Scholar] [CrossRef]
- Nicol, S.; Pauly, T.; Bindoff, N.L.; Wright, S.; Thiele, D.; Hosie, G.W.; Strutton, P.G.; Woehler, E. Ocean circulation off east Antarctica affects ecosystem structure and sea–ice extent. Nature 2000, 406, 504–507. [Google Scholar] [CrossRef]
- Martin, B.; Koppelmann, R.; Kassatov, P. Ecological relevance of salps and doliolids in the northern Benguela Upwelling System. J. Plankton Res. 2017, 39, 290–304. [Google Scholar] [CrossRef]
- Atkinson, A.; Hill, S.L.; Pakhomov, E.A.; Siegel, V.; Reiss, C.S.; Loeb, V.J.; Steinberg, D.K.; Schmidt, K.; Tarling, G.A.; Gerrish, L.; et al. Krill (Euphausia superba) distribution contracts southward during rapid regional warming. Nat. Clim. Chang. 2019, 9, 142–147. [Google Scholar] [CrossRef]
- Sylvester, Z.T.; Brooks, C.M. Protecting Antarctica through co–production of actionable science: Lessons from the CCAMLR marine protected area process. Mar. Policy 2020, 111, 103720. [Google Scholar] [CrossRef]
- Teschke, K.; Brtnik, P.; Hain, S.; Herata, H.; Liebschner, A.; Pehlke, H.; Brey, T. Planning marine protected areas under the CCAMLR regime–The case of the Weddell Sea (Antarctica). Mar. Policy 2021, 124, 104370. [Google Scholar] [CrossRef]
- Brooks, C.M.; Crowder, L.B.; Curran, L.M.; Dunbar, R.B.; Ainley, D.G.; Dodds, K.J.; Gjerde, K.M.; Sumaila, U.R. Science-based management in decline in the Southern Ocean. Science 2016, 354, 185–187. [Google Scholar] [CrossRef]
- LaRue, M.; Brooks, C.; Wege, M.; Salas, L.; Gardiner, N. High-Resolution Satellite Imagery Meets the Challenge of Monitoring Remote Marine Protected Areas in the Antarctic and Beyond. Conserv. Lett. 2022, e12884, in press. [Google Scholar] [CrossRef]
- Heywood, K.J.; Garabato, A.C.N.; Stevens, D.P.; Muench, R.D. On the fate of the Antarctic Slope Front and the origin of the Weddell Front. J. Geophys. Res. 2004, 109, C06021. [Google Scholar] [CrossRef]
- Thompson, A.F.; Stewart, A.L.; Spence, P.; Heywood, K.J. The Antarctic slope current in a changing climate. Rev. Geophys. 2018, 56, 741–770. [Google Scholar] [CrossRef]
- Visbeck, M. Deep velocity profiling using Lowered Acoustic Doppler Current Profiler: Bottom track and inverse solution. J. Atmos. Ocean. Technol. 2002, 19, 794–807. [Google Scholar] [CrossRef]
- Egbert, G.D.; Erofeeva, S.Y. Efficient inverse modeling of barotropic ocean tides. J. Atmos. Ocean. Technol. 2002, 19, 183–204. [Google Scholar] [CrossRef]
- Weikert, H.; John, H.-C. Experiences with a modified Be multiple opening–closing plankton net. J. Plankton Res. 1981, 3, 167–176. [Google Scholar] [CrossRef]
- Tranter, D.J. Reviews on zooplankton sampling methods. In Zooplankton Sampling; Tranter, D.J., Ed.; Part I. 2; UNESCO: Paris, France, 1968; pp. 11–144. [Google Scholar]
- Harris, R.; Wiebe, P.; Lenz, J.; Skjoldal, H.; Huntley, M. (Eds.) Zooplankton Methodology Manual; Academic Press: London, UK; San Diego, CA, USA, 2000; p. 684. ISBN 978-0-080495-33-0. [Google Scholar]
- Bouchard, S.; Cote, S.; St-Jacques, J.; Robillard, G.; Renaud, P. Effectiveness of virtual reality exposure in the treatment of arachnophobia using 3D games. Technol. Health Care 2006, 14, 19–27. [Google Scholar] [CrossRef]
- World Register of Marine Species. 2022. Available online: https://www.marinespecies.org (accessed on 29 August 2022). [CrossRef]
- Hasle, G.R.; De Mendiola, B.R.E. The fine structure of some Thalassionema and Thalassiothrix species. J. Phycol. 1967, 6, 107–126. [Google Scholar] [CrossRef]
- Dodge, J.D.; Hermes, H.B. A revision of the Diplopsalis group of dinoflagellates (Dinophyceae) based on material from the British Isles. Bot. J. Linn. Soc. 1981, 83, 15–26. [Google Scholar] [CrossRef]
- Hasle, G.R.; Lange, C.B. Morphology and distribution of Coscinodiscus species from the Oslofjord, Norway, and the Skagerrak, North Atlantic. Diatom Res. 1992, 7, 37–68. [Google Scholar] [CrossRef]
- Hasle, G.R.; Syvertsen, E.E. Marine Diatoms. Identifying Marine Phytoplankton; Tomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1996; p. 385. [Google Scholar] [CrossRef]
- Zingone, A.; Chrtiennot-Dinet, M.-J.; Lange, M.; Medlin, L. Morphological and genetic characterization of Phaeocystis cordata and P. jahnii (Prymnesiophyceae), two new species from the Mediterranean Sea. J. Phycol. 1999, 35, 1322–1337. [Google Scholar] [CrossRef]
- Cefarelli, A.O.; Ferrario, M.E.; Almandoz, G.O.; Atencio, A.G.; Akselman, R.; Vernet, M. Diversity of the diatom genus Fragilariopsis in the Argentine Sea and Antarctic waters: Morphology, distribution and abundance. Polar Biol. 2010, 33, 1463–1484. [Google Scholar] [CrossRef]
- Chamnansinp, A.; Li, Y.; Lundholm, N.; Moestrup, O. Global diversity of two widespread, colony–forming diatoms of the marine plankton Chaetoceros socialis (syn. C. radians) and Chaetoceros gelidus sp. nov. J. Phycol. 2013, 49, 1128–1141. [Google Scholar] [PubMed]
- Zidarova, R.; Ivanova, P.; Dzhembekova, N. Diatom colonization and community development in Antarctic marine waters—A short–term experiment. Pol. Polar Res. 2020, 41, 187–212. [Google Scholar] [CrossRef]
- Stop-Bowitz, C. Polychaeta from the Michael Sars north Atlantic D–S Expedition 1910. Rep. Sci. Results Michael Sars North Atl. Deep-Sea Exped. 1948, 5, 1–91. [Google Scholar]
- Dales, R.P. Pelagic polychaetes of the Pacific Ocean. Bull. Scripps Inst. Oceanogr. 1957, 7, 99–168. [Google Scholar]
- Day, J.H. Polychaeta of Southern Africa. Bull. Br. Mus. Zool. 1967, 656, 1–878. [Google Scholar]
- Ushakov, P.V. Polychaete Worms; Fauna SSSR, Ed.; Izdatelstvo AN SSSR: Moscow, Russia, 1972; p. 446. [Google Scholar]
- Pertsova, K.N. Larvae of euphausiids of the Antarctic. Ecol. Biogeogr. Plankton Proc. Inst. Oceanol. RAS 1976, 105, 147–170. [Google Scholar]
- Fauchald, K. The Polychaete Worms: Definitions and Keys to the Orders, Families and Genera; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1977; p. 188. [Google Scholar]
- Boltovskoy, D. South Atlantic Zooplankton; Backhuys Publishers: Leiden, The Netherlands, 1999; p. 868. [Google Scholar]
- Razouls, C.; De Bovee, F.; Kouwenberg, J.; Desreumaux, N. Diversity and Geographic Distribution of Marine Planktonic Copepods; CNRS—Sorbonne University: Paris, France, 2005; Available online: http://copepodes.obs–banyuls.fr/en/ (accessed on 3 February 2022).
- Watkins, J.L.; Morris, D.J.; Ricketts, C. Nocturnal changes in the mean length of a euphausiid population: Vertical migration, net avoidance, or experimental error? Mar. Biol. 1985, 86, 123–127. [Google Scholar] [CrossRef]
- Morris, D.J.; Watkins, J.L.; Ricketts, C.; Buchholz, F.; Priddle, J. An assessment of the merits of length and weight measurements of Antarctic krill Euphausia superba. Br. Antarct. Surv. Bull. 1988, 79, 27–50. [Google Scholar]
- Lomakina, N.B. Euphausiids of the Oceans (Euphausiacea); Nauka: Leningrad, Russia, 1978; p. 222. [Google Scholar]
- Makarov, R.R.; Denys, C.J. Stages of sexual maturity of Euphausia superba Dana. BIOMASS Handb. 1980, 11, 1–13. [Google Scholar]
- North, A.W.; White, M.G. Key to fish postlarvae from the Scotia Sea, Antarctica. Cybium 1982, 6, 13–32. [Google Scholar]
- Efrementko, V.N. Atlas of fish larvae of the Southern Ocean. Cybium 1983, 7, 1–74. [Google Scholar]
- North, A.W.; Kellermann, A. Key to the early stages of Antarctic fish. Ber. Polarforsch. 1990, 67, 1–44. [Google Scholar]
- Iwami, T.; Naganobu, M. A Guide to Identification of Fishes Caught Along with Antarctic Krill; Working Document WG-EMM-07/32; CCAMLR: Hobart, Australia, 2007; pp. 1–27. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER–E Ltd.: Plymouth, UK, 2006; p. 192. [Google Scholar]
- Anderson, M.J.; Gorley, A.M.; Clarke, K.R. PERMANOVA + for PRIMER: Guide to Software and Statistical Methods; PRIMER–E Ltd.: Plymouth, UK, 2008; p. 214. [Google Scholar]
- Clark, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; PRIMER–E Ltd.: Plymouth, UK, 2001; p. 144. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Clarke, K.R.; Sommerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero–adjusted Bray–Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Clarke, K.R. Non–parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Siegel, V.; Kawaguchi, S.; Ward, P.; Litvinov, F.; Sushin, V.; Loeb, V.; Watkins, J. Krill demography and large–scale distribution in the southwest Atlantic during January/February 2000. Deep-Sea Res. II 2004, 51, 1253–1273. [Google Scholar] [CrossRef]
- Smith, P.E.; Richardson, S. Standard techniques for pelagic fish egg and larva surveys. FAO Fish. Aquac. Tech. Pap. 1977, 175, 27–73. [Google Scholar]
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Lee, W.; Kim, S.; Kang, S.; Bang, H.; Lee, K.; Kwak, I. Distribution and abundance of zooplankton in the Bransfield Strait and the western Weddell Sea during austral summer. Ocean Polar Res. 2004, 26, 607–618. [Google Scholar] [CrossRef]
- Atkinson, A.; Ward, P.; Hunt, B.P.V.; Pakhomov, E.A.; Hosie, G.W. An overview of Southern Ocean zooplankton data: Abundance, biomass, feeding and functional relationships. CCAMLR Sci. 2012, 19, 171–218. [Google Scholar]
- Tokarczyk, R. Classification of water masses in the Bransfield Strait and Southern part of the Drake Passage using a method of statistical multidimensional analysis. Pol. Polar Res. 1987, 8, 333–336. [Google Scholar]
- Orsi, A.H.; Nowlin, W.D.; Whitworth, T. On the circulation and stratification of the Weddell Gyre. Deep-Sea Res. I 1993, 40, 169–203. [Google Scholar] [CrossRef]
- Huneke, W.G.; Huhn, O.; Schroeder, M. Water masses in the Bransfield Strait and adjacent seas, austral summer 2013. Polar Biol. 2016, 39, 789–798. [Google Scholar] [CrossRef]
- Krechik, V.A.; Frey, D.I.; Morozov, E.G. Peculiarities of water circulation in the central part of the Bransfield Strait in January 2020. Dokl. Earth Sci. 2021, 496, 92–95. [Google Scholar] [CrossRef]
- Morozov, E.G.; Flint, M.V.; Spiridonov, V.A. Antarctic Peninsula region of the Southern Ocean. In Advances in Polar Ecology; Springer: Cham, Switzerland, 2021; Volume 6, p. 433. [Google Scholar] [CrossRef]
- Youngs, M.K.; Thompson, A.F.; Flexas, M.M.; Heywood, K.G. Weddell Sea export pathways from surface drifters. J. Phys. Oceanogr. 2015, 45, 1068–1085. [Google Scholar] [CrossRef]
- Morozov, E.G.; Krechik, V.A.; Frey, D.I.; Polukhin, A.A.; Artemiev, V.A.; Kasyan, V.V.; Sapozhnikov, P.V.; Mukhametianov, R.Z. Frontal Zone between Relatively Warm and Cold Waters in the Northern Weddell Sea. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 31–54. [Google Scholar] [CrossRef]
- Criales-Hernandez, M.I.; Jerez-Guerrero, M.; Latandret-Solana, S.A.; Gomez-Sanchez, M.D. Spatial distribution of meso- and macrozooplankton in the Bransfield Strait and around Elephant Island, Antarctic Peninsula, during the 2019–2020 austral summer. Polar Sci. 2022, 32, 100821. [Google Scholar] [CrossRef]
- Krek, A.V.; Krek, E.V.; Krechik, V.A. The Circulation and Mixing Zone in the Antarctic Sound in February 2020. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 83–100. [Google Scholar] [CrossRef]
- Dorschel, B.; Gutt, J.; Huhn, O.; Bracher, A.; Huntemann, M.; Huneke, W.; Gebhardt, C.; Schroder, M.; Herr, H. Environmental information for a marine ecosystem research approach for the northern Antarctic Peninsula (RV Polarstern expedition PS81, ANT–XXIX/3). Polar Biol. 2016, 39, 765–787. [Google Scholar] [CrossRef]
- Van Caspel, M.; Hellmer, H.H.; Mata, M.M. On the ventilation of Bransfield Strait deep basins. Deep-Sea Res. II 2018, 149, 25–30. [Google Scholar] [CrossRef]
- Sapozhnikov, P.V.; Kalinina, O.Y.; Morozova, T.V. Phytopelagic communities of the Powell Basin in the summer of. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 279–306. [Google Scholar] [CrossRef]
- Voronina, N.M. Pelagic Ecosystems of the Southern Ocean; Nauka: Moscow, Russia, 1984; p. 206. [Google Scholar]
- Pinkerton, M.H.; Decima, M.; Kitchener, J.A.; Takahashi, K.T.; Robinson, K.V.; Stewart, R.; Hosie, G.W. Zooplankton in the Southern Ocean from the continuous plankton recorder: Distributions and long–term change. Deep-Sea Res. I 2020, 162, 103303. [Google Scholar] [CrossRef]
- Tarling, G.A.; Ward, P.; Thorpe, S.E. Spatial distributions of Southern Ocean mesozooplankton communities have been re-silient to long-term surface warming. Glob. Change Biol. 2018, 24, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Tarling, G.A.; Peck, V.; Ward, P.; Ensor, N.; Achterberg, E.; Tynan, E.; Poulton, A.J.; Mitchell, E.; Zubkov, M.V. Effects of acute ocean acidification on spatially–diverse polar pelagic foodwebs: Insights from on–deck microcosms. Deep-Sea Res. II 2016, 127, 75–92. [Google Scholar] [CrossRef]
- Siegel, V.; Watkins, J.L. Distribution, Biomass and Demography of Antarctic Krill, Euphausia superba. In Biology and Ecology of Antarctic Krill; Siegel, V., Ed.; Advances in Polar Ecology; Springer: Cham, Switzerland, 2016; pp. 21–100. [Google Scholar] [CrossRef]
- Spiridonov, V.A.; Zalota, A.K.; Yakovenko, V.A.; Gorbatenko, K.M. Composition of population and transport of juveniles of Antarctic krill in Powell Basin region (northwestern Weddell Sea) in January 2020. Tr. VNIRO. 2020, 181, 33–51. [Google Scholar] [CrossRef]
- Ross, R.M.; Quetin, L.B.; Baker, K.S.; Vernet, M.; Smith, R.C. Growth limitation in young Euphausia superba under field conditions. Limnol. Oceanogr. 2000, 45, 31–43. [Google Scholar] [CrossRef]
- Meyer, B.; Atkinson, A.; Blume, B.; Bathmann, U.V. Feeding and energy budgets of larval Antarctic krill Euphausia superba in summer. Mar. Ecol. Prog. Ser. 2003, 57, 167–177. [Google Scholar] [CrossRef]
- Hempel, I.; Hempel, G. Field observations on the developmental ascent of larval Euphausia superba (Crustacea). Polar Biol. 1986, 6, 123–126. [Google Scholar] [CrossRef]
- Spiridonov, V.A. Spatial and temporal variability in reproductive timing of Antarctic krill (Euphausia superba Dana). Polar Biol. 1995, 15, 161–174. [Google Scholar] [CrossRef]
- Schmidt, K.; Atkinson, A.; Venables, H.J.; Pond, D.W. Early spawning of Antarctic krill in the Scotia Sea is fuelled by “superfluous” feeding on non–ice associated phytoplankton blooms. Deep-Sea Res. II 2012, 59–60, 159–172. [Google Scholar] [CrossRef]
- Murphy, E.J.; Thorpe, S.E.; Watkins, J.L.; Hewitt, R. Modeling the krill transport pathways in the Scotia Sea: Spatial and environmental connections generating the seasonal distribution of krill. Deep-Sea Res. II 2004, 51, 1435–1456. [Google Scholar] [CrossRef]
- Gao, Q.; Xu, Z.; Huang, H.; Chen, X.; Feng, C.; Li, L. Geographical distribution and age composition of Euphausia superba larvae (Crustacea: Euphausiacea) in the South Shetland Islands region and southern Scotia Sea in relation to environmental conditions. Acta Oceanol. Sin. 2013, 32, 59–67. [Google Scholar] [CrossRef]
- Hofmann, E.E.; Klinck, J.M.; Locarnini, R.A.; Fach, B. Krill transport in the Scotia Sea and environs. Antarct. Sci. 1998, 10, 406–415, hdl:10013/epic.21917.d001. [Google Scholar] [CrossRef]
- Makarov, R.R.; Menshenina, L.L. On the study of distribution and phenology of euphausiid larvae in the Scotia Sea. Antarctic 1988, 27, 166–174. [Google Scholar]
- Tebble, N. Distribution of pelagic polychaetes in the South Atlantic Ocean. Nature 1958, 182, 166–167. [Google Scholar] [CrossRef]
- Tebble, N. The Distribution of pelagic polychaetes in the South Atlantic Ocean. Discov. Rep. 1960, 30, 163–284. [Google Scholar] [CrossRef]
- Lana, P.C.; Blankensteyn, A. Distribution patterns of pelagic polychaetes in the southern Drake Passage and Bransfield Strait (January–February 1984). Neritica 1987, 2, 37–64. [Google Scholar]
- Sicinski, J. Pelagic Polychaeta in the Scotia Front west of Elephant Island (BIOMASS III, October–November 1986). Pol. Polar Res. 1988, 9, 277–282. [Google Scholar]
- Siegel, V.; Skibowski, A.; Harm, U. Community structure of the epipelagic zooplankton community under the sea–ice of the northern Weddell Sea. Polar Biol. 1992, 12, 15–24. [Google Scholar] [CrossRef]
- Kittel, W.; Sicinski, J.; Zmijewska, M.I.; Bielecka, L.; Błachowiak-Samolyk, K. Antarctic neritic zooplankton community (Admiralty Bay, King George Island, South Shetland Islands). Pol. Polar Res. 2001, 22, 11–33. [Google Scholar]
- Oresland, V.; Bray, R.A. Parasites and headless chaetognaths in the Indian Ocean. Mar. Biol. 2005, 147, 725–734. [Google Scholar] [CrossRef]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of worms emended: An update of polychaete feeding guilds. Annu. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef]
- Bluhm, B.A.; Janout, M.A.; Danielson, S.L.; Ellingsen, I.; Gavrilo, M.; Grebmeier, J.M.; Hopcroft, R.R.; Iken, K.B.; Ingvaldsen, R.B.; Jørgensen, L.L.; et al. The Pan-Arctic continental slope: Sharp gradients of physical processes affect pelagic and benthic ecosystems. Front. Mar. Sci. 2020, 7, 544386. [Google Scholar] [CrossRef]
- Fauchald, K.; Jumars, P.A. The diet of worms: A study of polychaete feeding guilds. Oceanogr. Mar. Biol. Annu. Rev. 1979, 17, 193–284. [Google Scholar]
- Amei, K.; Jimi, N.; Kitamura, M.; Yokoi, N.; Yamaguchi, A. Community structure and seasonal changes in population structure of pelagic polychaetes collected by sediment traps moored in the subarctic and subtropical western North Pacific Ocean. Zoosympozia 2020, 19, 41–50. [Google Scholar] [CrossRef]
- Hopkins, T.L. Food web of an Antarctic midwater ecosystem. Mar. Biol. 1985, 89, 197–212. [Google Scholar] [CrossRef]
- Hopkins, T.L. Midwater food web in McMurdo Sound, Ross Sea, Antarctica. Mar. Biol. 1987, 96, 93–106. [Google Scholar] [CrossRef]
- Hopkins, T.L.; Torres, J.J. Midwater food web in the vicinity of a marginal ice zone in the western Weddell Sea. Deep-Sea Res. II 1989, 36, 543–560. [Google Scholar] [CrossRef]
- Lee, S.H.; Joo, H.M.; Joo, H.; Kim, B.K.; Song, H.J.; Jeon, M.; Kang, S.H. Large contribution of small phytoplankton at Marian Cove, King George Island, Antarctica, based on long–term monitoring from 1996 to 2008. Polar Biol. 2015, 38, 207–220. [Google Scholar] [CrossRef]
- Deppeler, S.L.; Davidson, A.T. Southern Ocean phytoplankton in a changing climate. Front. Mar. Sci. 2017, 4, 40. [Google Scholar] [CrossRef]
- Biggs, T.E.; Alvarez-Fernandez, S.; Evans, C.; Mojica, K.D.; Rozema, P.D.; Venables, H.J.; Pond, D.W.; Brussaard, C.P.D. Antarctic phytoplankton community composition and size structure: Importance of ice type and temperature as regulatory factors. Polar Biol. 2019, 42, 1997–2015. [Google Scholar] [CrossRef]
- Johnston, N.M.; Murphy, E.J.; Atkinson, A.; Constable, A.J.; Cotte, C.; Cox, M.; Daly, K.L.; Driscoll, R.; Flores, H.; Halfter, S. Status, change, and futures of zooplankton in the Southern Ocean. Front. Ecol. Evol. 2022, 9, 624692. [Google Scholar] [CrossRef]
- Yang, G.; Atkinson, A.; Hill, S.L.; Guglielmo, L.; Granata, A.; Li, C. Changing circumpolar distributions and isoscapes of Antarctic krill: Indo–Pacific habitat refuges counter long–term degradation of the Atlantic sector. Limnol. Oceanogr. 2020, 66, 272–287. [Google Scholar] [CrossRef]
- Yakovenko, V.A.; Spiridonov, V.A.; Gorbatenko, K.M.; Shadrin, N.V.; Samyshev, E.Z.; Minkina, N.I. Macro- and mesozooplankton in the Powell Basin (Antarctica): Species composition and distribution of abundance and biomass in February 2020. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 131–141. [Google Scholar] [CrossRef]
- Ross, R.M.; Quetin, L.B.; Kirsch, E. Effect of temperature on developmental times and survival of early larval stages of Euphausia superba Dana. J. Exp. Mar. Biol. Ecol. 1988, 121, 55–71. [Google Scholar] [CrossRef]
- Warwick-Evans, V.; Fielding, S.; Reiss, C.; Watters, G.; Trathan, P. Estimating the average distribution of Antarctic krill Euphausia superba at the northern Antarctic Peninsula during austral summer and winter. Polar Biol. 2022, 45, 857–871. [Google Scholar] [CrossRef]
- Atkinson, A.; Shreeve, R.; Hirst, A.; Rothery, P.; Tarling, G.; Pond, D.; Korb, R.; Murphy, E.; Watkins, J.L. Natural growth rates in Antarctic krill (Euphausia superba): II. Predictive models based on food, temperature, body length, sex, and maturity stage. Limnol. Oceanogr. 2006, 51, 973–987. [Google Scholar] [CrossRef]
- Tarling, G.A.; Shreeve, R.S.; Hirst, A.G.; Atkinson, A.; Pond, D.W.; Murphy, E.J. Natural growth rates in Antarctic krill (Euphausia superba): I. Improving methodology and predicting intermolt period. Limnol. Oceanogr. 2006, 51, 959–972. [Google Scholar] [CrossRef]
- Tarling, G.A.; Cuzin-Roudy, J.; Thorpe, S.E.; Shreeve, R.S.; Ward, P.; Murphy, E.J. Recruitment of Antarctic krill Euphausia superba in the South Georgia region: Adult fecundityand the fate of larvae. Mar. Ecol. Prog. Ser. 2007, 331, 161–179. [Google Scholar] [CrossRef]
- Murzina, S.A.; Voronin, V.P.; Frey, D.I.; Orlov, A.M. Features of the distribution of juveniles and mature Antarctic krill Euphausia superba depending on some environmental factors. In Proceedings of the Actual Problems of Crustacea Studies Conference, Borok, Russia, 23–25 May 2022; p. 44. [Google Scholar]
- Wiedenmann, J.; Cresswell, K.; Mangel, M. Temperature–dependent growth of Antarctic krill: Predictions for a changing climate from a cohort model. Mar. Ecol. Prog. Ser. 2008, 358, 191–202. [Google Scholar] [CrossRef]
- Perry, F.A.; Atkinson, A.; Sailley, S.F.; Tarling, G.A.; Hill, S.L.; Lucas, C.H.; Mayor, D.J. Habitat partitioning in Antarctic krill: Spawning hotspots and nursery areas. PLoS ONE 2019, 14, e0219325. [Google Scholar] [CrossRef]
- Nishikawa, J.; Naganobu, M.; Ichii, T.; Ishii, H.; Terazaki, M.; Kawaguchi, K. Distribution of salps near the South Shetland Islands during austral summer, 1990–1991 with special reference to krill distribution. Polar Biol. 1995, 15, 31–39. [Google Scholar]
- Brotz, L.; Cheung, W.W.L.; Kleisner, K.; Pakhomov, E.; Pauly, D. Increasing jellyfish populations: Trends in large marine ecosystems. Hydrobiologia 2012, 690, 3–20. [Google Scholar] [CrossRef]
- Constable, A.J.; Melbourne-Thomas, J.; Corney, S.P.; Arrigo, K.R.; Barbraud, C.; Barnes, D.K.A.; Bindoff, N.L.; Boyd, P.W.; Brandt, A.; Costa, D.P.; et al. Climate change and Southern Ocean ecosystems I: How changes in physical habitats directly affect marine biota. Glob. Chang. Biol. 2014, 20, 3004–3025. [Google Scholar] [CrossRef]
- Henschke, N.; Everett, J.D.; Richardson, A.J.; Suthers, I.M. Rethinking the role of salps in the ocean. Trends Ecol. Evol. 2016, 31, 720–733. [Google Scholar] [CrossRef]
- Pakhomov, E.A.; Dubischar, C.; Strass, V.; Brichta, M.; Bathmann, U. The tunicate Salpa thompsoni ecology in the Southern Ocean.—I. Distribution, biomass, demography andfeeding ecophysiology. Mar. Biol. 2006, 149, 609–623. [Google Scholar] [CrossRef]
- Everett, J.; Baird, M.; Suthers, I. Three–dimensional structure of a swarm of the salp Thalia democratica within a cold–core eddy off southeast Australia. J. Geophys. Res. 2011, 116, C12046. [Google Scholar] [CrossRef]
- Kohlbach, D.; Graeve, M.; Lange, B.A.; David, C.; Schaafsma, F.L.; van Franeker, J.A.; Vortkamp, M.; Brandt, A.; Flores, H. Dependency of Antarctic zooplankton species on ice algae–produced carbon suggests a sea ice–driven pelagic system during winter. Glob. Change Biol. 2018, 24, 4667–4681. [Google Scholar] [CrossRef]
- Slosarczyk, W.; Cielniaszek, Z. Postlarval and juvenile fish (Pisces, Perciformes and Myctophiformes) in the Antarctic Peninsula region during BIOMASS–SIBEX 1983/84. Polish. Polar Res. 1985, 6, 159–165. [Google Scholar]
- Kellermann, A. On the biology of early life stages of notothenioid fishes (Pisces) off the Antarctic Peninsula. Ber. Zur Polarforsch. 1986, 31, 1–149. [Google Scholar]
- Kellermann, A. The larval fish community in the zone of seasonal pack-ice cover and its seasonal and interannual variability. Arch. Fisch. 1989, 39, 81–109. [Google Scholar]
- Kellermann, A.; Kock, K.H. Patterns of spatial and temporal distribution and their variation in early life stages of Antarctic fish in the Antarctic Peninsula region. In Antarctic Ocean and Resources Variability; Springer: Berlin/Heidelberg, Germany, 1989; pp. 147–159. [Google Scholar]
- Kellermann, A.; Schadwinkel, S. Winter aspects of the ichthyoplankton community in Antarctic Peninsula waters. Polar Biol. 1991, 11, 117–127. [Google Scholar] [CrossRef]
- Morales-Nin, B.; Palomera, I.; Schadwinkel, S. Larval fish distribution and abundance in the Antarctic Peninsula region and adjacent waters. Polar Biol. 1995, 15, 143–154. [Google Scholar] [CrossRef]
- Catalan, I.A.; Morales-Nin, B.; Rotllant, G.; Palomera, I.; Emelianov, M. Environmental influences on zooplankton and micronekton distribution in the Bransfield Strait and adjacent waters. Polar Biol. 2008, 31, 691–707. [Google Scholar] [CrossRef]
- La Mesa, M.; La Mesa, G.; Catalano, B.; Jones, C.D. Spatial distribution pattern and physical–biological interactions in the larval notothenioid fish assemblages from the Bransfield Strait and adjacent waters. Fish. Oceanogr. 2016, 25, 624–636. [Google Scholar]
- Lancraft, T.M.; Torres, J.J.; Hopkins, T.L. Micronekton and macrozooplankton in the open waters near Antarctic ice edge zones (AMERIEZ 1983 and 1986). Polar Biol. 1989, 9, 225–233. [Google Scholar] [CrossRef]
- Skora, K.E. Ichthyoplankton near ice edge between King George Island and the South Orkney Islands. Pol. Polar Res. 1991, 12, 4–9. [Google Scholar]
- Fisher, E.C.; Kaufmann, R.S.; Smith, K.L. Variability of epipelagic macrozooplankton/micronekton community structure in the NW Weddell Sea, Antarctica (1995–1996). Mar. Biol. 2004, 144, 345–360. [Google Scholar] [CrossRef]
- White, M.G.; Piatkowski, U. Abundance, horizontal and vertical distribution of fish in eastern Weddell Sea micronekton. Polar Biol. 1993, 13, 41–53. [Google Scholar] [CrossRef]
- Loeb, V.J.; Kellermann, A.K.; Koubbi, P.; North, A.W.; White, M.G. Antarctic larval fish assemblages: A review. Bull. Mar. Sci. 1993, 53, 416–449. [Google Scholar]
- Santora, J.A.; Sydeman, W.J.; Schroeder, I.D.; Reiss, C.S.; Wells, B.K.; Field, J.C.; Cossio, A.M.; Loeb, V.J. Krill space: A comparative assessment of mesoscale structuring in polar and temperate marine ecosystems. ICES J. Mar. Sci. 2012, 69, 1317–1327. [Google Scholar] [CrossRef]
- Hamner, W.M. Biomechanics of filter feeding in the Antarctic krill Euphausia superba: Review of past work and new observations. J. Crustac. Biol. 1988, 8, 149–163. [Google Scholar] [CrossRef]
- Cleary, A.C.; Durbin, E.G.; Casas, M.C. Feeding by Antarctic krill Euphausia superba in the West Antarctic Peninsula: Differences between fjords and open waters. Mar. Ecol. Prog. Ser. 2018, 595, 39–54. [Google Scholar] [CrossRef]
- Haberman, K.L.; Ross, R.M.; Quetin, L.B. Diet of the Antarctic krill (Euphausia superba): I selective grazing in mixed phytoplankton assemblages. J. Exp. Mar. Biol. Ecol. 2003, 283, 97–113. [Google Scholar] [CrossRef]
- Pauli, N.-C.; Metfies, K.; Pakhomov, E.A.; Neuhaus, S.; Graeve, M.; Wenta, P.; Flintrop, C.M.; Badewien, T.H.; Iversen, M.H.; Meyer, B. Selective feeding in Southern Ocean key grazers—Diet composition of krill and salps. Commun. Biol. 2021, 4, 1061. [Google Scholar] [CrossRef]
- Paffenhofer, G.A.; Koster, M. Digestion of diatoms by planktonic copepods and doliolids. Mar. Ecol. Prog. Ser. 2005, 297, 303–310. [Google Scholar] [CrossRef]
- Von Harbou, L.; Dubischar, C.D.; Pakhomov, E.A.; Hunt, B.P.V.; Hagen, W. Salps in the Lazarev Sea, Southern Ocean: I. Feeding dynamics. Mar. Biol. 2011, 158, 2009–2026. [Google Scholar] [CrossRef]
- Kasatkina, S.M.; Abramov, A.M.; Sokolov, M.Y. Biomass and distribution of Antarctic krill in the Antarctic Atlantic area in January–February 2020. Tr. AtlantNIRO 2021, 5, 49–61. [Google Scholar]
- Cox, M.J.; Borchers, D.L.; Demer, D.A.; Cutter, G.R.; Brierley, A.S. Estimating the density of Antarctic krill (Euphausia superba) from multibeam echo–sounder observations using distance sampling methods. J. R. Stat. Soc. Ser. C 2011, 60, 301–316. [Google Scholar] [CrossRef]
- Kasatkina, S.M. Methodical aspects of acoustic survey for Antarctic krill in the CCAMLR Convention. Tr. AtlantNIRO 2021, 5, 39–48. [Google Scholar]
- Krag, L.A.; Herrmann, B.; Iversen, S.A.; Engas, A.; Nordrum, S.; Krafft, B.A. Size Selection of Antarctic Krill (Euphausia superba) in trawls. PLoS ONE 2014, 9, e102168. [Google Scholar] [CrossRef]
- Wang, Z.; Tang, H.; Herrmann, B.; Xu, L. Catch pattern for Antarctic krill (Euphausia superba) of different commercial trawls in similar times and overlapping fishing grounds. Front. Mar. Sci. 2021, 8, 670663. [Google Scholar] [CrossRef]
- Samyshev, E.; Minkina, N.; Chudinovskih, E.; Melnikov, V.; Bitiutskii, D.; Usachev, S.; Saluk, P.; Zuev, O.; Serebrennikov, A.; Orlov, A. Distribution and demography of Antarctic krill and salps in the Atlantic Sector of the Antarctic during austral summer 2021–2022. Water 2022, 14. submitted. [Google Scholar]
- Hill, S.L.; Phillips, T.; Atkinson, A. Potential Climate Change Effects on the Habitat of Antarctic Krill in the Weddell Quadrant of the Southern Ocean. PLoS ONE 2013, 8, e72246. [Google Scholar] [CrossRef]
- Murphy, E.J.; Johnston, N.M.; Hofmann, E.E.; Phillips, R.A.; Jackson, J.A.; Constable, A.J.; Henley, S.F.; Melbourne-Thomas, J.; Trebilco, R.; Cavanagh, R.D.; et al. Global Connectivity of Southern Ocean Ecosystems. Front. Ecol. Evolution. 2021, 9, 624451. [Google Scholar] [CrossRef]
- Xiao, C. Changes in Antarctic Climate System: Past, Present and Future. Adv. Clim. Chang. Res. 2008, 4, 1–7. [Google Scholar]
- Beaugrand, G.; Luczak, C.; Edwards, M. Rapid biogeographical plankton shifts in the North Atlantic Ocean. Glob. Change Biol. 2009, 15, 1790–1803. [Google Scholar] [CrossRef]
- Henley, S.F.; Schofield, O.M.; Hendry, K.R.; Schloss, I.R.; Steinberg, D.K.; Moffat, C.; Peck, L.S.; Costa, D.P.; Bakker, D.C.E.; Hughes, C.; et al. Variability and change in the west Antarctic Peninsula marine system: Research priorities and opportunities. Prog. Oceanogr. 2019, 173, 208–237. [Google Scholar] [CrossRef]
- McCormack, S.A.; Melbourne-Thomas, J.; Trebilco, R.; Griffith, G.; Hill, S.L.; Hoover, C.; Johnston, N.M.; Marina, T.I.; Murphy, E.J.; Pakhomov, E.A.; et al. Southern Ocean Food Web Modelling: Progress, Prognoses, and Future Priorities for Research and Policy Makers. Front. Ecol. Evol. 2021, 9, 624763. [Google Scholar] [CrossRef]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasyan, V.V.; Bitiutskii, D.G.; Mishin, A.V.; Zuev, O.A.; Murzina, S.A.; Sapozhnikov, P.V.; Kalinina, O.Y.; Syomin, V.L.; Kolbasova, G.D.; Voronin, V.P.; et al. Composition and Distribution of Plankton Communities in the Atlantic Sector of the Southern Ocean. Diversity 2022, 14, 923. https://doi.org/10.3390/d14110923
Kasyan VV, Bitiutskii DG, Mishin AV, Zuev OA, Murzina SA, Sapozhnikov PV, Kalinina OY, Syomin VL, Kolbasova GD, Voronin VP, et al. Composition and Distribution of Plankton Communities in the Atlantic Sector of the Southern Ocean. Diversity. 2022; 14(11):923. https://doi.org/10.3390/d14110923
Chicago/Turabian StyleKasyan, Valentina V., Dmitrii G. Bitiutskii, Aleksej V. Mishin, Oleg A. Zuev, Svetlana A. Murzina, Philipp V. Sapozhnikov, Olga Yu. Kalinina, Vitaly L. Syomin, Glafira D. Kolbasova, Viktor P. Voronin, and et al. 2022. "Composition and Distribution of Plankton Communities in the Atlantic Sector of the Southern Ocean" Diversity 14, no. 11: 923. https://doi.org/10.3390/d14110923
APA StyleKasyan, V. V., Bitiutskii, D. G., Mishin, A. V., Zuev, O. A., Murzina, S. A., Sapozhnikov, P. V., Kalinina, O. Y., Syomin, V. L., Kolbasova, G. D., Voronin, V. P., Chudinovskikh, E. S., & Orlov, A. M. (2022). Composition and Distribution of Plankton Communities in the Atlantic Sector of the Southern Ocean. Diversity, 14(11), 923. https://doi.org/10.3390/d14110923