Abstract
The fatty acid profile of seven different organs (brain, gills, heart, muscles, gonads, liver, and gall bladder) of the lipid-rich Antarctic silverfish, Pleuragramma antarcticum, from the Antarctic Sound was studied. Qualitative and quantitative analyses of fatty acids were performed using gas chromatography with mass-selective and flame ionization detectors. It was seen that the muscles and gills differed from the other organs with the prevalence of saturated fatty acids, mainly due to 16:0 and 14:0 that may reveal their additional metabolic role in these organs for adaptation of the fish to the cold Antarctic waters. Monounsaturated fatty acids 18:1(n − 9) and 16:1(n − 7) were prevalent in the other organs of silverfish, especially their high content in the liver, reflecting the carnivorous feeding type (mainly adult ice krill) of the species and their dietary preferences in the austral summer. The high content of essential fatty acids, 22:6(n − 3) (14% of total fatty acids) found in the brain and 20:5(n − 3) (12% of total fatty acids) found in the muscles, highlights the functional significance of these lipid components in the studied tissues of fish. The results will improve the understanding of the functional value of individual fatty acids in terms of their organ specificity and the adaptation of the Antarctic silverfish to life in the frigid ice waters of the Antarctic pelagic ecosystem.
1. Introduction
The Antarctic silverfish, Pleuragramma antarcticum (Boulenger 1902), is the only truly pelagic fish in the Antarctic waters [1] representing the Nototheniidae family; 97% of these species are endemic to the Southern Ocean and are characterized by a benthic or bentho–pelagic way of life [2,3]. It lives in the epipelagic and mesopelagic zones up to a depth 700 m and is abundant in both numbers and biomass in the shelf zone of most coastal regions of Antarctica, where nototheniid fishes comprise about 92% of the local ichthyofauna [2,4,5,6,7]. In the process of evolution, the Antarctic silverfish returned at least 40 m.y. ago to a secondary pelagic way of life [2] from a benthic ancestor without a swim bladder [8,9] and is considered the only species among nototheniids that retains a pelagic lifestyle through all its life stages [6,10,11,12,13]. This species is well adapted to life in the Antarctic seas [14], including through the lipid composition.
Pronounced lipid (fat) storage mainly in the form of triacylglycerols (TAGs) is a peculiar characteristic of the P. antarcticum as well as the lipid-rich Aethotaxis mitopteryx, distinguishing these shelf species among nototheniid fishes from the high-Antarctic ones [15,16,17]. Lipid accumulation in the silverfish body occurs during the long-lasting postlarval development (7–9 years) [18,19], reaching the maximum content in adults (30–57% of dry weight) [16,20,21]. Significant reserves of lipids and their fatty acid (FA) composition define the ability of silverfish to grow and develop in the shelf zone in ice-laden seawater (temperature about − 1.9 °C). In contrast, fishes from the Myctophidae family do not exhibit such adaptation at the lipid level and therefore are not competitive for living in the shelf zone, being dominant in the far offshore Antarctica [22,23].
Pleuragramma antarcticum has key adaptation in the form of subcutaneous (0.2–1.5 mm) and large intermuscular (0.5–3.0 mm diameter) lipid sacs of a unique structure, the wall of which consists of several white adipocytes arranged circumferentially around a large lipid droplet [15]. The high lipid content in the body (also due to the presence of lipid sacs) together with high water in the body (80%) [24] and reduced bone density [25,26] are considered an adaptation that strongly reduced the density of silverfish and thus achieved almost neutral buoyancy in seawater [27,28,29]. At the same time, the use of the lipids as sources of energy is not excluded [14,15,16,20,27]. The lipid sac walls are cellular, and the lipids may be available for metabolism in addition to their obvious role in buoyancy [15]. The high lipid content in the body (about 40% of dry weight), as a similar adaptation, can be seen in the polar fish Leptoclinus maculatus from the Arctic. L. maculatus postlarvae accumulates TAG in special large lipid sacs, the main functions of which are first to store energy and then to maintain buoyancy that allow the young fish to successfully develop in the pelagic waters (3–5 years) and survive during periods when food is insufficient in the Arctic [30,31,32,33].
The lipid composition of Antarctic silverfish during embryonic and especially during early postembryonic development, including its lipid sacs, has been well studied, revealing its dietary preferences at different developmental stages, using the FA composition as trophic biomarkers [16,20,28,34,35,36]. However, the changes in the lipid composition of silverfish during puberty and ageing are still poorly studied. The majority of research is about lipid and FA content of P. antarcticum in comparison with other Antarctic fish species, especially nototheniids [16,17,21,28,37,38]. There are only some studies on the lipid content in a few organs or tissues of adult nototheniids fishes from the Antarctic [38,39,40].
This study aims to present for the first time a detailed analysis of the FA profile in the key functional organs of the body–brain, heart, liver, gall bladder, gonads, gills, and muscles–of lipid-rich silverfish P. antarcticum from the Antarctic Sound (Southern Ocean) collected during the austral summer season. We propose to highlight the tissue specificity of FA content in the body of this silverfish, which will allow a better understanding of the role of lipids in fish as an organism and the mechanism by which the lipid level is used to adapt the species to the extreme Antarctic environment. The results of this study will complement the data on the lipids in P. antarcticum as a keystone irreplaceable species in the Antarctic coastal food web [41,42,43,44], being a highly energetic prey item for some fish [45], marine mammals and birds (e.g., toothed whales, seals, and Emperor penguins) [46,47].
2. Materials and Methods
The fishes were collected in the Antarctic Sound (Atlantic sector of Antarctic) separating the Joinville Island group from the northeast tip of the Antarctic Peninsula (63°28.6′ S; 56°31.2′ W). Sampling was carried out during the austral summer scientific expedition on board the research vessel “Akademik Mstislav Keldysh” in January 2022.
Adults of P. antarcticum (n = 4) were caught using Isaacs–Kidd midwater trawl as modified by Samyshev and Aseev, comprising a non-closing fishing gear with a mouth area of 6 m2, a net part length of 25 m from a knotless net with a mesh of 6 mm and an insert in the cod from a nylon sieve No. 15 (0.67 mm) [48]. The chosen depth range for trawling was aimed at collecting fish in the well-inhabited water column. It is known that generally, Antarctic silverfish inhabits a depth range of 300 to 500 m in daytime, within normal shelf depths [49]. In this paper, vertical migration to depths greater than 400 m is discussed from the position of escape behavior of silverfish from deep-diving predators [50,51,52]. Species identification was carried out as per the work in [53]. Length of the studied fishes (mean 15.43 ± 0.48 cm; range 14.1–16.4 cm) and weight (mean 40.95 ± 2.81 g; range 36.1–48.9 g) were measured before dissection. Organ and tissue samples (muscles, liver, heart, gonads, brain, gills, and gall bladder) were quickly removed using sterile scalpel and forceps with the fishes kept on frozen blocks and were then fixed at − 80 °C in an Eppendorf CryoCube freezer (Eppendorf, Stevenage, UK). The white muscle tissue for lipid analysis was biopsied from the posteriodorsal part of the body; the brain and the heart were considered and biopsied entirely; the results regarding the gall bladder are reported on the FA content of the bile. Only adult mature females were chosen for the present study. Hydrophysical parameters of the environment were measured using the CTD-rosette equipment package.
2.1. Total Lipids Extraction Procedure and Fatty Acids’ Analysis
Total lipids were extracted with chloroform/methanol (2:1 by volume) following the method by Folch et al. (1957) [54], using a Hei-VAP Advantage ML/G3 rotary evaporator (Heidolph Instruments, Schwabach, Germany). Qualitative and quantitative analyses of total FAs were conducted by gas chromatography (GC) with a mass selective detector (MSD) and a flame ionization detector (FID), after subjecting the total lipids mixture to FA methylation [55]. All analysis parameters for GC-MSD and GC-FID were identical except for the gas mobile phase (helium and nitrogen). To determine the qualitative content of the resulting mixture of FA methyl esters (FAMEs), they were separated by GC with a Maestro-αMS mono-quadrupole mass-selective detector (Scietegra, Moscow, Russia). Fractionation was performed for 35 min in a temperature gradient mode (140 °C for 5 min, raising the temperature to 240 °C at a rate of 4 °C/min) on an HP-88 capillary column (Agilent Technologies, Santa Clara, CA, USA), using helium as the mobile phase. FAMEs were detected in the SIM/SCAN mode: the SIM mode was used to detect FAs included in the analytical standards Supelco 37, Bacterial Acid Methyl Ester (BAME) Mix, and PUFA (polyunsaturated FAs) No. 1 Marine source (Sigma-Aldrich, Saint Louis, MO, USA); SCAN mode was used for searching and identifying unique FA components in the range of 50–400 m/z. The obtained data were analyzed using the Maestro Analyst v. 1.025 and the NIST library. Further, after qualitative identification on GC-MS, a quantitative analysis was performed on a GC-FID Chromatec-Cristall-5000.2 (Chromatec, Yoshkar-Ola, Russia). Fractionation was performed for 35 min in a temperature gradient mode (140 °C for 5 min, raising the temperature to 240 °C at a rate of 4 °C/min) on an HP-88 capillary column (Agilent Technologies, Santa Clara, CA, USA), using nitrogen as the mobile phase. The procedure for processing the results obtained in the software Chromatec-Analytic v. 3.0.298.1 (Chromatec, Yoshkar-Ola, Russia) is described in a previous study conducted by Murzina et al. (2020) [31]. Biochemical analysis was performed at the Laboratory of Ecological Biochemistry, using the equipment of the Core Facility of the Karelian Research Centre of the Russian Academy of Sciences.
2.2. Statistical Analysis
Statistical analysis of the results was conducted in the open R programming environment. The results are shown in Table 1 as mean M ± SE (standard error). The significance of differences between lipid components in different organs/tissues was analyzed using the nonparametric Wilcoxon–Mann–Whitney rank-sum test [56]. The differences were considered statistically significant at p-value ≤ 0.05. To visualize the differences in the FA content in the studied organs of P. antarcticum, the non-metric multidimensional scaling (NMDS) method was used. The calculation of the correlation coefficients of each of the biochemical parameters with the NMDS ordination axes and the assessment of the statistical significance of these coefficients based on the permutation test (with 999 permutations) showed the p-value ≤ 0.001 for all studied FAs, except 20:0 (p-value ≤ 0.012).

Table 1.
The content of fatty acids (% of total fatty acids) in different tissues of adult Antarctic silverfish Pleuragramma antarcticum from the Antarctic Sound, January 2022.
3. Results
Data were analyzed for the hydrophysical parameters of the environment, measured using the CTD-rosette equipment package [57]. The average value of the parameters in the sampling place are presented: temperature: − 0.54 °C; salinity: 34.49‰; oxygen: 10.51 mg/L; and chlorophyll A: 0.47 µg/L. The detailed dynamics of the environmental factors is shown in Figure 1.
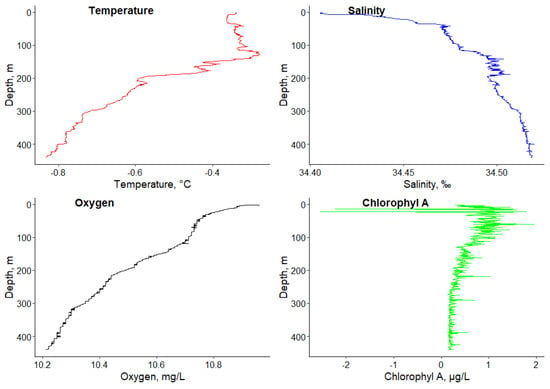
Figure 1.
Vertical profiles of temperature, salinity, dissolved oxygen, and chlorophyl A at the depth gradient (0–440 m) in the sampling area of Pleuragramma antarcticum (the Antarctic Sound, Atlantic sector of Antarctic).
The fatty acid composition of total lipids in different tissues of the Antarctic silverfish are presented in the Table 1.
3.1. Muscles
The dominant FAs of the muscles were 16:0, 18:1(n − 9), 20:5(n − 3), 16:1(n − 7), 14:0, 18:1(n − 7), 20:0, and 22:6(n − 3) (listed in the order of decreasing content), comprising more than 80% of total FAs (Table 1). Among the FA groups, saturated FAs (SFAs) were dominant (Figure 2) and accounted for 41.3% due to mainly 14:0 (10.2%) and 16:0 (19.7%) FAs. Monounsaturated FAs (MUFAs) were 35.1% due to three dominant FAs: 16:1(n − 7) (10.2%), 18:1(n − 9) (16.2%), and 18:1(n − 7) (6.3%). Polyunsaturated FAs (PUFA) accounted for 23.7% of which 21.1% belonged to the FA of n − 3 family (Figure 2). Muscles also differed from other organs by high content of such major FAs as 14:0, 16:0, 20:5(n − 3), and minor FAs such as 18:2(n − 6) (1.4%), 18:3(n − 3) (0.7%), and 18:4(n − 3) (3.1%). The content of eicosapentaenoic FA 20:5(n − 3) (12%) was the highest in the muscles compared with the other organs, whereas the level of docosahexaenoic FA 22:6(n-3) was only 4.9% of total FAs. The ratio of long-chain unsaturated essential 20:5(n − 3)/22:6(n − 3) PUFA was 2.5 and the ratio of 16:0/18:1(n − 9) was 1.1 pointed on lipogenesis rates.
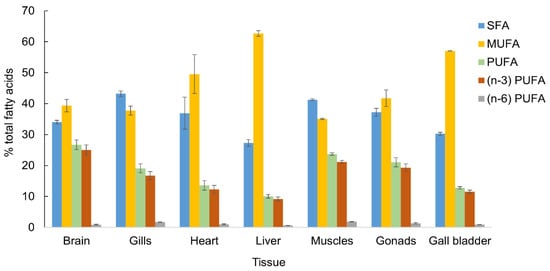
Figure 2.
The content of fatty acid groups (% of total fatty acids) in different tissues of adult Antarctic silverfish Pleuragramma antarcticum from the Antarctic Sound, January 2022. Abbreviations: SFA—saturated fatty acids, MUFA—monounsaturated fatty acids, PUFA—polyunsaturated fatty acids, (n − 3) and (n − 6)—families of PUFA.
3.2. Gills
The principal FAs of the gills were 16:0, 18:1(n − 9), 16:1(n − 7), 14:0, 20:5(n − 3), 18:1(n − 7), 20:0, and 22:6(n − 3) (listed in the order of decreasing content). SFAs also dominated in the gills and accounted for 43.2% (Figure 2) due to mainly 16:0 (19.7% of total FAs) and 14:0 (9.3%) FAs (Table 1). MUFA were 37.7% due to FAs 16:1(n − 7) (10.8%), 18:1(n − 9) (18.2%), and 18:1(n − 7) (6.5%). The major difference between the gills and muscles was in the content of PUFA (19.1%) in which the level of 20:5(n − 3) FA (9.3%) in the gills was significantly lower compared with that in the muscles. The content of essential FAs in the gills was higher than that in the other organs (except for muscles) and performed as 18:2(n − 6) (1.3%), 18:3(n − 3) (0.6%), and 18:4(n − 3) (2.5%). The ratio of 20:5(n − 3)/22:6(n − 3) was 2.3. The content of n − 3 PUFA was 16.1%. The gills in comparison to the muscles were distinguished by the higher content of 18:2(n − 6) FA (1.26 and 1.43, respectively). The ratio 16:0/18:1(n − 9) was 1.1. It is notable that this ratio was the highest for muscles and gills.
3.3. Heart
The dominant FAs in the heart were 18:1(n − 9), 16:1(n − 7), 16:0, 20:0, 20:5(n − 3), 18:0, 14:0, 18:1(n − 7), 18:0, and 22:6(n − 3). MUFA were the prevalent FAs in the heart and accounted for 49.5% of total FAs due to 18:1(n − 9) FA, which was 25.9% of total FAs and significantly lower than that in the liver. SFAs followed MUFAs and comprised 36.9%. Among them, 20:0 FA differed in the highest content of 8.6%. The PUFA content was 13.6%, where were 12.3% FA of n-3 family due to eicosapentaenoic FA (EPA) (6.3%) and docosahexaenoic FA (DHA) (3.6%). The ratio of essential FA 20:5(n − 3)/22:6(n − 3) was 1.7 and the ratio 16:0/18:1(n − 9) was 0.6.
3.4. Liver
The main FAs in the liver were 18:1(n − 9), 16:1(n − 7), 16:0, 20:0, 20:5(n − 3), 18:0, 14:0, 18:1(n − 7), 18:0, and 22:6(n − 3). MUFA were the prevalent FAs and accounted 62.7% (Figure 2). Among the MUFAs, the content of 16:1(n − 7) (23.4%) and 18:1(n − 9) (33.9% of total FAs) was the highest. SFAs followed the MUFAs and were 27.3%. The content of 14:0 (2.7%) and 16:0 (11.6%) FAs were significantly lower than that in the muscles and gills. The PUFA content only accounted for 10.1%; 9.2% was detected for n − 3 PUFAs due to EPA (4.8%) and DHA (2.7%). The ratio of essential FA 20:5(n − 3)/22:6(n − 3) was 1.8, and the ratio of 16:0/18:1(n − 9) was 0.3.
3.5. Gall Bladder
The major FAs in the gall bladder were 18:1(n − 9), 16:1(n − 7), 16:0, 20:5(n − 3), 20:0, 18:1(n − 7), 18:0, 14:0, 18:0 and 22:6(n − 3). MUFA were the prevalent FAs and accounted for 57.0% due to 16:1(n − 7) (19.7%) and especially 18:1(n − 9) FA (30.9%) (Table 1). MUFA were followed by SFAs and accounted for 30.3%. The PUFA content was 12.8% due to n-3 PUFA (11.5%). The 20:5(n − 3)/22:6(n − 3) ratio was 2.0, and the ratio of 16:0/18:1(n − 9) was 0.4.
3.6. Gonads
The major FAs in the gonads were 18:1(n − 9), 16:0, 16:1(n − 7), 20:5(n − 3), 20:0, 22:6(n − 3), 14:0, 18:1(n − 7) and 18:0. MUFA were the prevalent FAs and accounted for 41.7% of total FAs due to 18:1(n − 9) FA (20.8%). MUFA were followed by SFAs and accounted for 37.2%. The PUFA content was 21.1% due to n − 3 PUFAs (19.3%). The gonads differed in their high level of EPA after muscles and DHA (6.5%) after the brain. The ratio of 20:5(n − 3)/22:6(n − 3) was 1.5, and the ratio 16:0/18:1(n − 9) was 0.8.
3.7. Brain
The major FAs in the brain were 18:1(n − 9), 22:6(n − 3), 16:0, 16:1(n − 7), 20:5(n − 3), 20:0, 18:0, 18:1(n − 7) and 14:0. MUFA were the prevalent FAs and accounted for 39.3% due to 16:1(n − 7) (9.4%) and especially due to 18:1(n − 9), the content of the latter being two-fold higher (20%). The content of 24:1(n − 9) FA was dominant (3.2%) compared with the other organs (≤0.3%). MUFA were followed by SFAs and accounted for 34.0%. The PUFA content was 26.7% and was mostly contributed by n − 3 PUFAs (25.0%). The content of 22:6(n − 3) FA in the brain was the highest (14.2%) among all the studied organs. The ratio of essential FA 20:5(n − 3)/22:6(n − 3) was 0.6, and the ratio of 16:0/18:1(n − 9) was 0.7.
To visualize the differences in the FA content in the studied organs of P. antarcticum, the non-metric multidimensional scaling (NMDS) method was used as an indirect gradient analysis approach, which produces an ordination based on a distance or dissimilarity matrix (Figure 3). The NMDS method is used to facilitate recognition and the interpretation of patterns and differences among groups. The highest content of the major FA 22:6(n − 3) and minor FA 24:1(n − 9) was in the brain (p-value = 0.001) (Figure 3). Fields of gills and muscles are presented in the same part of the graph and differed in the higher content of minor food-derived FAs: 18:2(n − 6), 18:3(n − 3), 18:4(n − 3) and 14:0 FA. The liver differed in the content of 16:1(n − 7) and 18:1(n − 9) FAs. By calculating the Spearman correlation coefficient between the two matrices (with the original data and the chi-square matrix from the original data) for all options, the best metric was set at “man” (Manhattan distances). The measure of discrepancies between the original and simulated distance matrices (value “Stress”) was high at 0.061.
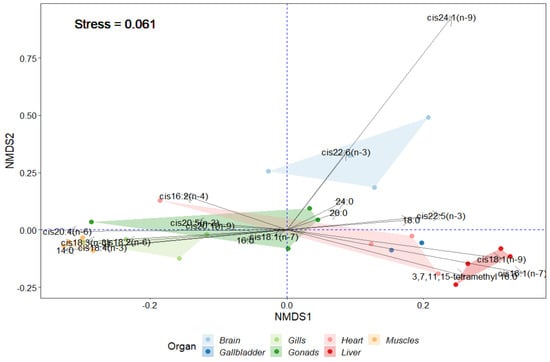
Figure 3.
Visualization of the differences in the fatty acid content in different tissues of Pleuragramma antarcticum using non-metric multidimensional scaling (NMDS).
4. Discussion
It was found that 14:0, 16:0, 16:1(n − 7), 18:1(n − 9), 18:1(n − 7), 20:5(n − 3), and 22:6(n − 3) FAs were the most dominant in P. antarcticum in all the studied tissues. A similar profile of these dominant FAs was shown in the composition of total lipids in the whole body of silverfish and other studied nototheniid fishes from the Weddell and Lazarev seas of the Antarctic [17]. It is known that these FAs are included in the main groups of FAs in fish, accounting for about 75–85% of total FAs [58,59]. The tissue specificity of FA content in silverfish was reflected in the present study due to the content of some major and minor FAs.
The muscles and gills of the silverfish were different from the other organs in the prevalence of SFAs (41% and 43%, respectively) due to the high level of 16:0 and 14:0. Palmitic acid, 16:0, is an essential component of FA metabolism in any organism. It has recently been suggested that myristic acid, 14:0, may be an activator of the conversion of α-linolenic acid to docosahexaenoic acid (DHA) [60]. According to the content of FA groups in the muscles and gills, it can be arranged as follows: SFA (41–43%) > MUFA (35–38%) > PUFA (19–23%). MUFA dominated and was followed by SFAs in the other studied organs of silverfish. A similar content of FA groups as in the gills and muscles in this study was shown in the whole body of adult P. antarcticum, with SFA (31–33%) > MUFA (54%) > PUFA (14–15%), and the lipid-rich fish, Aethotaxis mitopteryx, collected from the Weddell and Lazarev seas in another study by Hagen et al. (2000) [17] that is supported by our results. In contrast, the content of SFA was below that of all other FA groups: MUFA (43–48%) > PUFA (28–32%) > SFA (19–27%) in the body of other benthic lipid-poor, Antarctic nototheniids such as Dolloidraco longedorsalis, Bathydraco marry and bentho–pelagic Trematomus lepidorhinus [17]. The prevalence of SFA in the body of the lipid-rich P. antarcticum (specifically in the gills and muscles) distinguishes this species from others and may point to the special role of SFA in the adaptation of the P. antarcticum to live in the frigid, ice-shelf pelagic waters. SFAs, due to their saturation, are less oxidized in the highly oxygenated Antarctic waters, and these FAs are the “starting point” of FA metabolism or lipogenesis in general, maintaining any demand of an organism.
It is known that saturated and unsaturated FAs are indispensable components in the structure of membranes, with their ratio and positional redistribution in membrane lipids maintaining the definite phase states and the fluidity for work of membrane enzymes and ion channels at low temperatures [61,62,63,64,65]. Generally, low environmental temperatures serve as a trigger to activate several cold-inducible genes, increasing the extent of FA unsaturation, changes in the protein composition, etc. [66,67,68]. Despite the predominance of SFAs in the muscles and gills of silverfish, the necessary content of unsaturation is provided by vital PUFAs. Among the PUFAs in all the studied organs, FAs of n-3 PUFAs were prevalent due to DHA and EPA, which is characteristic of marine fish living in extreme environments [69,70,71,72]. The PUFA content reaches the highest values of 24% and 26% only in the muscles and brain, respectively. The content of PUFA in silverfish muscle was the same as that of some Arctic polar fish, for example the adults of the Arctic fish, the daubed shanny Leptoclinus maculatus (21–22%) [73,74]. At the same time, the ratio n-3/n-6 PUFA in the muscles of silverfish was 12.0, whereas in fishes from the Pacific Ocean, it ranges from 4.9 to 8.1 [75]. This ratio reaches 7.4 in L. maculatus adults from the Arctic [74,76]. Thus, we see that there is an exclusive accumulation of high amounts of PUFAs that are known as essential and uniform due to their physical and chemical components that help silverfish maintain normal physiology in extremely cold waters and high environmental fluctuations.
The high content of SFAs in silverfish gills and muscles may indicate their special role in the adaptation of these fish to life in the shelf ice zone of the Antarctic. It is known that apart from the main function of SFAs as components of phospholipid or sphingolipid domains, contributing to the densification of the biomembrane, SFAs have other roles no less important in the body [77]. SFAs are involved, for instance, in lipogenesis, hormone production, fat deposition, PUFA bioavailability and apoptosis, elucidated biochemical mechanisms such as protein acylation (N-myristoylation, S-palmitoylation), and regulation of gene transcription [78]. SFAs are part of the myelin sheath of neurons in significant amounts, which indicates the common functional activity in the brain and nervous system. The coordination of muscle contraction and the ability to learn also require the intake of SFAs [78]. It was shown that the heart of silverfish was distinguished by a higher content of saturated 20:0 FA (Table 1), which may indicate its importance for the functioning of the fish heart. It is known that dietary fats in rats directly affect the properties of the myocardium [79].
Silverfish has been observed to be feeding inshore in large schools [80,81] under the ice or at the edge of the ice. It can also be assumed that the high level of SFAs probably also turn on certain defense mechanisms in fish. In the study by Sampaio et al. (2011) [82], it was established that the sphingolipids during polarization became longer, more saturated, and more hydroxylated–as required, to generate an apical membrane domain that serves as a protective barrier for the epithelial sheet. In marine mammals, saturated fats are often found in adult cell membranes due to their lower chemical ability to be damaged from reactive oxygen species [83], and thus play a critical role in protecting the animal from the effects of hypoxia. Despite a generally slow lifestyle of adult P. antarcticum, its individuals are able to perform diel and seasonal vertical migrations under the ice down to 600 m and deeper in inshore troughs and fjords [50,51,52] and also seasonal vertical migrations towards the bottom prior to winter [84]. The high level of SFAs in the gills suggests that these acids may be involved in the maintenance of fish gas exchange, especially during periods of ice feeding and during migration to great depths. Its migratory pattern allows P. antarcticum to avoid visual predators such as penguins, seals, and seabirds during the day and to capitalize on abundant prey such as Antarctic krill Euphausia superba nearer the surface at night [51,85,86]. These vertical migrations require saving energy or additional energy costs. Saturation of the membrane leads to a decrease in membrane fluidity and leakiness to both protons and ions and a decrease in significant energy expenditure of the cell [87]. Some studied shown high level of saturation of membrane phospholipids in insulin resistant skeletal muscle [88,89,90], while there is a decrease in the glycogen synthesis in muscle and glucose uptake.
MUFAs were prevalent in all the studied organs with the exception only of gills and muscles. They are the direct components of storage lipids such as TAGs, which are used first as energy sources unlike wax esters [91,92,93]. The accumulation of lipids in P. antarcticum mainly in form of TAGs increases from 60% of total lipids in first-year juveniles to 80% of total lipids in adults [16,20,21]. MUFAs in the lipid sacs of silverfish do not exclude their being used as an energy source [14,15,16,27]. It is well known that SFAs are synthesized in the organism and used as a source of energy [94], while endogenous biosynthesis requires additional energy of the organism as compared with dietary sources. Triacylglycerol lipase preferentially mobilizes unsaturated fats relative to saturated fats, for example, in Antarctic nototheniids Trematomus newnesi and Notothenia gibber [95], and carnitine palmitoyl transferase activities are higher with 16:1 as a substrate than with 16:0, 18:1, 18:2, or 20:4 [96]. The high level of SFAs in the gills and muscles could be construed as the additional energy sources in gas change processes or during vertical migrations of silverfish, maintaining its lipid reserves and thereby contributing to the neutral buoyancy of this fish. The lipid and FA content of P. antarcticum during the winter period is poorly studied when food is probably scarce; this could clarify the main sources of energy of silverfish more accurately.
Among the MUFAs, as components of energetic TAG, 18:1(n − 9) is by far the most dominant FA in the adults of P. antarcticum, followed by 16:1(n − 7) and 16:0. Oleic FA, 18:1(n − 9), was dominant in the whole body of adults (25% of total FAs) of Antarctic silverfish from the Weddell and Lazarev seas, followed by similar amounts of 16:0 and 14:0 (15% TFA) [16,17]. Fatty acid analysis of Antarctic fish species reveals the dominance of oleic FA, 18:1(n − 9), in TAG in species such as Dissostichus mawsoni, Trematomus newnesi and Notothenia gibberifrons [40]. The FAs of the structure 18:1 are known to be the most important in the adaptation of coldwater fishes to deep waters [71,91,97,98,99]. The maximum amount of the dominant MUFAs 18:1(n − 9) (34%) and 16:1(n − 7) (24%) was shown in the liver of silverfish in the present study. Among the seven groups visualized using the NMDS and representing the seven studied organs, it was shown that the group of liver is not overlapped with other organ areas and is defined due to 18:1(n − 9), 16:1(n − 7), and 16:0 FAs in tandem. MUFA are mainly derived from food deposited into the liver as the main organ of lipid and FA metabolism. Gall bladder also had a high level of these FAs (30% and 20%, respectively), showing their significance in supporting ordinary physiological and functional conditions of this organ of the digestive system. It is known that the FAs of a bile in the gall bladder are involved in mechanisms that regulate gall bladder and extrahepatic bile duct motility [100,101]. Pancreatic and gall bladder responses were augmented by increasing FA chain length and FA loads [102]. The dominance of these FAs in other tissues and organs indicates further distribution of this FA within the blood system.
It is known that a significant contribution to the level of FAs in the body is the food intake [103,104,105,106]. However, considerable biosynthetic production of lipids in adult P. antarcticum de novo from the elongation and desaturation of 14:0 and 16:0 dietary precursors to 18:1(n − 9) cannot be ruled out, considering it has been found in other marine fishes [93,104]. P. antarcticum are highly flexible in their feeding. It is known that adults of the Antarctic silverfish are primarily carnivorous and prey mainly on pelagic and bentho–pelagic invertebrates, with dominance of euphausiids (E. superba and E. crystallorophias) and copepods (especially Calanus acutus, C. propinquus, Metridia gerlachei, Paraeuchaeta sp., and Rhincalanus gigas [42,43,107,108,109,110,111]. In addition, the diet of P. antarcticum may also include amphipods, mysids, pelagic polychaetes, pteropods, ostracods, and other benthic invertebrates as well as fish [42,80,107,108,109,110,112,113]. No studies have found any benthic prey being consumed by adult silverfish, even when they were caught near the seabed [42,47,113].
The use of FAs as trophic biomarkers becomes impractical due to the possibility of obtaining the same acids from different sources. However, the composition and the content of FAs in some Arctic invertebrates may allow emphasis on the main sources of nutrition of silverfish. A high ratio of 18:1n − 9/18:1n − 7 FA is a signature of carnivorousness in aquatic organisms [103,105,106,114]. The value of this ratio in the liver of silverfish was 7.9, which is twice that in other carnivorous marine (5.0) and freshwater (5.6) fishes [115], supporting the carnivorous type of feeding of silverfish. The main prey of silverfish during the austral summer is considered to be ice krill, Euphausia crystallorophias and Thysanoessa macrura, which are especially rich in the 18:1(n − 9) FA [116,117]. Indeed, E. superba, which is dominant near the continental slope or over deeper waters, was sometimes the only prey item found in silverfish stomachs [41,80]. P. antarcticum were able to seize and hold prey individually [41], reflected in the structure of the jaw apparatus (conical, curved, and mostly small teeth, but with usually 3–4 larger teeth, including prominent canines) [53], which confirms the possibility of selective feeding. Moreover, silverfish able to switch feeding mode according to the abundance and size of the available prey have both types of feeding, similar to a ram and a tow-net filter [118]. The tow-net filter feeders surround the prey with their open mouths while swimming rapidly [119]. In ram feeding, the predator ingests free-swimming prey by forward movement of the body and protruding jaws [120], which is more conducive to selective feeding. E. superba mainly accumulates TAG apart from smaller amounts of PL [17,121]. E. crystallorophias and T. macrura primarily store wax esters [99,121,122]. The FA composition of younger stages of E. superba was dominated by 20:5(n − 3), 22:6(n − 3), and 16:0 FAs, whereas 18:1(n − 9), 14:0, and 16:0 FAs were prevalent in the FAs of adults, comprising about 70% of the total TAG FAs [123]. The predominance of 18:1(n − 9), 14:0, and 16:0 FAs in silverfish may also indicate their feeding during the austral summer mainly on adult Antarctic krill rich in lipids.
The 18:1(n − 9) FA may also come from the other principal prey items of silverfish: cyclopoid copepods of the genus Oncaea [124]. They are similarly rich in wax esters as Calanoides acutus, but the former’s FAs are strongly dominated by 18:1(n − 9) (33–79% of total FA), while 14:0 and 16:0 prevail in the fatty alcohol of wax esters [125]. Calanoid copepods Rhincalanus gigas and Euchaeta spp. [124] are the other relevant food items accumulating large amounts of wax esters with a dominance of 18:1(n − 9) FA; hence, an analysis of FAs cannot be used to trace the consumption of these copepods [17]. Nevertheless, the high percentage of 16:1(n − 7) in those copepods, up to 25% of total FAs in Euchaeta spp., may reflect the feeding on the copepods by adult silverfish [124].
It is known that the content of 20:1(n − 9) and 22:1(n − 11) FAs is considered as trophic markers of Arctic dominant copepods (Calanus spp.); 20:1 and 22:1 FAs are biomarkers for such invertebrates living in the Antarctic region as C. acutus and Calanus propinquus [104,126,127], which also are food sources for silverfish. The trophic marker 20:1(n − 9) comprises about a quarter of total FAs in C. acutus [126,128]. C. propinquus is the only dominant Antarctic copepod that biosynthesizes very high amounts of the FA 22:1(n − 9), with up to 26% of total FAs [129]. In our work, the content of 20:1(n − 9) was identified only in trace amounts (less than 2% in all the organs). The level of 20:1(n − 9) and 22:1(n − 11) FAs in P. antarcticum from the Weddell and Lazarev seas were higher: 5.7% and 4.5% of total FAs, respectively [17]. Elevated proportions of these two FAs were also found in the tissues of adult silverfish in the Dumont d’Urville Sea in East Antarctic [36]. It is known that the content of 20:1(n − 9) reached up to 17% of total FAs in juvenile silverfish but decreased to 6% of total FAs in the adults. A similar trend occurred for 22:1(n − 11) and 22:1(n − 9), with maxima of 12% and 10% in juveniles, respectively, decreasing to about 3–4% in adults [16]. This result implies the smaller contribution of C. acutus and C. propinquus to the diet of silverfish in the Antarctic Sound during the austral summer.
The study of the distribution of PUFAs in the organs and tissues of silverfish showed that the gills and muscles differed from the other organs in the higher content of EPA and of minor (no more than 1.5%) essential 18:2(n − 6), 18:3(n − 3), and 18:4(n − 3) FAs, which are of dietary origin. The NMDS analyses showed that the groups of gills and muscles are located nearby and removed to a certain distance from other organ groups (Figure 3) that pointed to a direct impact on the environmental factors and the role of these FAs in the formation of a compensatory response to their influence. These FAs may be accidentally ingested with ice algae [104] or introduced in the diet via herbivorous species or aquatic invertebrates. Recently, it has been shown that they can also be biosynthesized in many aquatic invertebrates, whose desaturases have activities of Δ12 and Δ15 [129,130,131]. The highest level of n-3 PUFA due to EPA and DHA was in the brain, muscles, and gonads. It is known that these PUFAs are important for the development of the gonads and further in the process of embryonic and postembryonic development of fish [104,132]. Lipids have an important function in some nototheniids as energy reserves for gonad development [133]. The overlap of NMDS groups of gonads and heart could be an indication of the importance of certain FAs to maintain functional activity of such energy demand organs. It is interesting that among the PUFAs in the muscle, EPA (12%) was prevalent, while in the brain it was DHA (14%), which are outstanding proportions compared with the content in the other organs (3.5–5%). The prevalence of EPA in the muscles may reflect its role in functionality of this tissue. EPA plays not only the role of maintaining the necessary fluidity of biomembranes for the functioning of membrane-bound enzymes, but it can also be used for the synthesis of eicosanoids [70,97,134]. In addition, endothelial cell migration ability of EPA is known, which is important in wound-healing processes [135].
The significant content of DHA in the silverfish brain clearly highlights the role of this FA in this vital organ as an important structural and functional component of the nervous system [136,137,138]. DHA is the most abundant FA of the n − 3 family in the brain and retina of vertebrates [136,138]. The very high concentration of DHA neural tissues was established in other fishes [136,137,139,140]. Recent studies highlight the importance of DHA in maintaining cellular homeostasis and brain cell survival and recovery [141]. The results of some studies [137] indicate that DHA can be synthesized in the fish brain and eye in vivo, not excluding its role for liver in the biosynthesis and provision of DHA for developing neural tissues in fish. The ratio of n − 3/n − 6 PUFA in the brain of silverfish was 33%, which is extremely high. Marine fish invariably obtain EPA and DHA from their natural food, which is rich in these FAs [134,136]. Nutritional depletion of DHA in the brain and retinal membranes is accompanied by functional defects such as reduced visual acuity and impaired learning abilities [136]. Some brain lipids may support brain function at low temperatures. Highly polar brain gangliosides suggest wide-ranging neurophysiological adaptations to ensure proper functioning of the nervous system in icy waters [14]. We can only assume the contribution of DHA in the performance of similar functions in P. antarcticum. The brain of silverfish was different due to the higher content of minor FA 24:1(n − 9) compared with that in the other organs. Using NMDS, the brain is clearly distinguished among other organs due to DHA and 24:1(n − 9) FAs. The area of the brain is not overlapped with the area of other groups, indicating a very specific and important role of these FAs in this organ compared to the others (Figure 3). Nervonic acid 24:1 is the dominant FA of sphingomyelin of the brain and can be half of all FAs in sphingomyelin in teleosts [58]. It is known that DHA levels in fish may also reflect the level of their motor activity [142]. It was shown that among the marine fish, the highest content of DHA in their body was found in tunas and flying fishes (20.8%), which have the highest motor activity [142]. In nototheniids, with average locomotor activity, the value was 8.9%, and in inactive ones, for example, flounders and halibuts, it was 5.8% [142]. The content of DHA in silverfish muscles was significantly lower (4.9% of total fatty acids) compared to the brain. This is the reason that the high content of this FA in the brain mainly reflects its functional value in this organ. Marine fish have higher levels of DHA and EPA in the brain (from 6.66 to 29.47% of total lipids for cod, saithe, redfish, salmon, and trout [143]) than freshwater fish (0.1–1.9% of total lipids for carp [144]), suggesting the special role for these FAs in adapting the fish brain for functioning in saltwater conditions. Data from cell and animal studies justify the indication of DHA in relation to brain function for neuronal cell growth and differentiation through the modulation of the physical properties of neuronal membranes, as well as in relation to signal transduction associated with G proteins, and gene expression [136,145].
5. Conclusions
P. antarcticum is a key species in the Antarctic shelf ecosystem providing an efficient energy flow through the lipid-based high Antarctic food web. The detailed FA variations in seven organs, namely the brain, gills, heart, muscles, liver, gonads, and gall bladder, were studied for the first time. FAs are the most labile components of lipids, diverse in their structure and physicochemical properties, which play a structural and energetic role in the metabolism at all stages of this species’ life cycle and are of key importance in biochemical adaptations [70,93,134].
The data obtained indicate the differences in the content of FAs in the studied tissues of silverfish. Muscles and gills differ from the other organs in the prevalence of the content of SFAs among other FA groups due to 16:0 and an especially high level of 14:0 FA. The higher levels of minor essential 18:2(n − 6) and 18:3(n − 3) FAs and 18:4(n − 3) in these organs, which are of dietary origin, were noted. The prevalence of SFAs in the body of the lipid-rich P. antarcticum (gills and muscles) distinguishes this species from the others and may point to the significant role of SFAs in fish as an organism that may reveal their additional metabolic role in these organs for adapting it to live in the icy Antarctic waters. Liver and gall bladder differed from the organs by the highest content of MUFAs due to 16:1(n − 7) and especially 18:1(n − 9), that reflects the dietary preferences of silverfish. It supports the carnivorous mode of life of silverfish and may reflect their trophic origin from adult krill Euphausia spp. as its main prey during the austral summer. The content of calanoid copepods biomarker FA (20:1 and 22:1) was minor and pointed to the small contribution of these preys to the silverfish diet. A very high content of DHA in the brain and EPA in the muscles has been established, which shows the specificity of their functions in these organs and may reflect their role in the adaptation to life in the icy Antarctic shelf waters.
Thus, silverfish is a unique, very lipid rich and ancient and endemic species, which inhabits the ice-covered shelf zones of the Antarctic seas. It is well adapted to life in the Antarctic region, which is clearly reflected in the composition and content of FAs in the studied organs. The results obtained will allow a better understanding of the role of lipids and the mechanism of adaptation at the lipid level in the body of fish living in frigid ice-shelf pelagic waters.
Author Contributions
Conceptualization, S.A.M. and S.N.P.; methodology, S.A.M., S.N.P., V.P.V., A.V.M. and A.M.O.; software, V.P.V.; validation, S.A.M. and S.N.P.; formal analysis, S.A.M., S.N.P., and V.P.V.; investigation, V.P.V. and S.A.M.; resources, V.P.V. and S.A.M.; data curation, S.A.M.; writing—original draft preparation, S.N.P., S.A.M.; writing—review and editing, S.A.M., A.M.O., and A.V.M.; visualization, V.P.V.; supervision, S.A.M.; project administration, S.A.M. and A.M.O.; funding acquisition, S.A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by State Task FMWE-2022-0001 by the Ministry of Science and Higher Education of the Russian Federation and the State Task for KarRC RAS FMEN-2022-0006.
Institutional Review Board Statement
Approved. The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of the Institute of Biology KarRC RAS (protocol code 27, 12 September 2022).
Data Availability Statement
All data are presented in the paper.
Acknowledgments
We are deeply thankful to the administration of the expedition and the crew of the research vessel “Academic Mstislav Keldysh” for their invaluable and professional help during the expedition and biomaterial collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kailola, P.J.; Williams, M.J.; Stewart, P.C.; Reichelt, R.E.; McNee, A.; Grieve, C. Australian Fisheries Resources; Bureau of Resource Sciences: Canberra, Australia, 1993; 422p.
- DeWitt, H.H. Coastal and Deep-Water Fishes of the Antarctic; Antarctic Map Folio Series; Folio 15; Bushnell, V.C., Ed.; American Geophysical Society: New York, NY, USA, 1971; pp. 1–18. [Google Scholar]
- Andriashev, A.P. A general review of the Antarctic fish fauna. In Biogeography and Ecology in Antarctica; Part of the Monographiae Biologicae book series; van Mieghem, J., van Oye, P., Eds.; Springer Science: Dordrecht, The Netherlands, 1965; Volume 15, pp. 491–550. [Google Scholar] [CrossRef]
- Hubold, G. Spatial distribution of Pleuragramma antarcticum (Pisces: Nototheniidae) near the Filchner-and Larsen ice shelves (Weddell Sea/Antarctica). Polar Biol. 1984, 3, 231–236. [Google Scholar] [CrossRef]
- Hubold, G. Ecology of notothenioid fish in the Weddell Sea. In Biology of Antarctic Fish; Springer: Berlin/Heidelberg, Germany, 1991; pp. 3–22. [Google Scholar]
- Donnelly, J.; Torres, J.J.; Sutton, T.T.; Simoniello, C. Fishes of the eastern Ross Sea, Antarctica. Polar Biol. 2004, 27, 637–650. [Google Scholar] [CrossRef]
- Eastman, J.T. The nature of the diversity of Antarctic fishes. Polar Biol. 2005, 28, 93–107. [Google Scholar] [CrossRef]
- Clarke, A.; Johnston, I.A. Evolution and adaptive radiation of Antarctic fishes. Trends Ecol. Evol. 1996, 11, 212–218. [Google Scholar] [CrossRef]
- Eastman, J.T. Evolution and diversification of Antarctic notothenioid fishes. Am. Zool. 1991, 31, 93–109. [Google Scholar] [CrossRef]
- Hubold, G.; Ekau, W. Midwater fish fauna of the Weddell Sea, Antarctica. In Proceedings of the Fifth Congress of European Ichthyologists, Stockholm, Sweden; Kullander, S.O., Fernholm, B., Eds.; Swedish Museum of Natural History: Stockholm, Sweden, 1987; pp. 391–396. [Google Scholar]
- Eastman, J.T.; DeVries, A.L. Buoyancy studies of notothenioid fishes in McMurdo Sound, Antarctica. Copeia 1982, 2, 385–393. [Google Scholar] [CrossRef]
- La Mesa, M.; Eastman, J.T. Antarctic silverfish: Life strategies of a key species in the high-Antarctic ecosystem. Fish Fish. 2012, 13, 241–266. [Google Scholar] [CrossRef]
- Orlov, A.M.; Mishin, A.V.; Artemenkov, D.V.; Murzina, S.A. Length-weight characteristics of some pelagic fishes in the high latitudes of Atlantic sector of the Southern Ocean. J. Ichthyol. 2022, 62, 237–243. [Google Scholar]
- Wöhrmann, A.P.; Hagen, W.; Kunzmann, A. Adaptations of the Antarctic silverfish Pleuragramma antarcticum (Pisces: Nototheniidae) to pelagic life in high-Antarctic waters. Mar. Ecol. Prog. Ser. 1997, 151, 205–218. [Google Scholar] [CrossRef][Green Version]
- Eastman, J.T.; DeVries, A.L. Ultrastructure of the lipid sac wall in the Antarctic notothenioid fish Pleuragramma antarcticum. Polar Biol. 1989, 9, 333–335. [Google Scholar] [CrossRef]
- Hagen, W.; Kattner, G. The role of lipids in the life history of the Antarctic silverfish Pleuragramma antarctica. In The Antarctic Silverfish: A Keystone Species in a Changing Ecosystem; Springer: Cham, Switzerland, 2017; pp. 131–148. [Google Scholar]
- Hagen, W.; Kattner, G.; Friedrich, C. The lipid compositions of High-Antarctic notothenioid fish species with different life strategies. Polar Biol. 2000, 23, 785–791. [Google Scholar] [CrossRef]
- Faleyeva, T.I.; Gerasimchuk, V.V. Features of reproduction in the Antarctic sidestripe, Pleuragramma antarcticum Nototheniidae. J. Ichthyol. 1990, 30, 67–79. [Google Scholar]
- Ferrando, S.; Hanchet, S.; Angiolillo, M.; Gambardella, C.; Pisano, E.; Tagliaferro, G.; Vacchi, M. Insights into the life cycle of the Antarctic Silverfish. In Reproduction Features of the Ross Sea Population. In Proceedings of the Abstracts of the International Polar Year Oslo Science Conference, Oslo, Norway, 8 June 2010; p. 1. [Google Scholar]
- Reinhardt, S.B.; Van Vleet, E.S. Lipid composition of twenty-two species of Antarctic midwater zooplankton and fish. Mar. Biol. 1986, 91, 149–159. [Google Scholar] [CrossRef]
- Friedrich, C.; Hagen, W. Lipid contents of five species of notothenioid fish from high-Antarctic waters and ecological implications. Polar Biol. 1994, 14, 359–369. [Google Scholar] [CrossRef]
- DeWitt, H.H. The character of the midwater fish fauna of the Ross Sea, Antarctica. In Antarctic Ecology I; Holdgate, M.W., Ed.; Academic Press: London, UK, 1970; pp. 305–314. [Google Scholar]
- Donnelly, J.; Torres, J.J. Pelagic fishes in the Marguerite Bay region of the West Antarctic Peninsula continental shelf. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 523–539. [Google Scholar] [CrossRef]
- Reisenbichler, K.R. Growth and Chemical Composition in Two Populations of the Antarctic Silverfish, Pleuragramma antarcticum (Pisces, Notoheniidae). Ph.D. Dissertation, University of California, Santa Barbara, CA, USA, 1993. [Google Scholar]
- Eastman, J.T.; Witmer, L.M.; Ridgely, R.C.; Kuhn, K.L. Divergence in skeletal mass and bone morphology in Antarctic notothenioid fishes. J. Morphol. 2014, 275, 841–861. [Google Scholar] [CrossRef]
- Voskoboinikova, O.; Detrich, H.W.; Albertson, R.C.; Postlethwait, J.H.; Ghigliotti, L.; Pisano, E. Evolution reshaped life for the water column: The skeleton of the Antarctic silverfish Pleuragramma antarctica Boulenger, 1902. In The Antarctic Silverfish: A Keystone Species in a Changing Ecosystem; Springer: Cham, Switzerland, 2017; pp. 3–26. [Google Scholar]
- Devries, A.L.; Eastman, J.T. Lipid sacs as a buoyancy adaptation in an Antarctic fish. Nature 1978, 271, 352–353. [Google Scholar] [CrossRef]
- Eastman, J.T. Lipid storage systems and the biology of two neutrally buoyant Antarctic notothenioid fishes. Comp. Biochem. Physiol. 1988, 90, 529–537. [Google Scholar] [CrossRef]
- Near, T.J.; Jones, C.D.; Eastman, J.T. Geographic intraspecific variation in buoyancy within Antarctic notothenioid fishes. Antarct. Sci. 2009, 21, 123–129. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Falk-Petersen, I.B.; Sargent, J.R. Structure and function of an unusual lipid storage organ in the Arctic fish Lumpenus maculatus Fries. Sarsia 1986, 71, 1–6. [Google Scholar] [CrossRef]
- Murzina, S.A.; Pekkoeva, S.N.; Kondakova, E.A.; Nefedova, Z.A.; Filippova, K.A.; Nemova, N.N.; Orlov, A.M.; Berge, J.; Falk-Petersen, S. Tiny but Fatty: Lipids and Fatty Acids in the Daubed Shanny (Leptoclinus maculatus), a Small Fish in Svalbard Waters. Biomolecules 2020, 10, 368. [Google Scholar] [CrossRef]
- Pekkoeva, S.N.; Murzina, S.A.; Nefedova, Z.A.; Ripatti, P.O.; Falk-Petersen, S.; Berge, J.; Lonne, O.J.; Nemova, N.N. Ecological role of lipids and fatty acids in the early postembryonic development of the daubed shanny, Leptoclinus maculatus (Fries, 1838) from Kongsfjorden, West Spitsbergen in winter. Russ. J. Ecol. 2017, 48, 240–244. [Google Scholar] [CrossRef]
- Pekkoeva, S.N.; Murzina, S.A.; Nefedova, Z.A.; Falk-Petersen, S.; Berge, J.; Lønne, O.J.; Nemova, N.N. Fatty acid composition of the postlarval daubed shanny (Leptoclinus maculatus) during the polar night. Polar Biol. 2020, 43, 657–664. [Google Scholar] [CrossRef]
- Mayzaud, P.; Chevallier, J.; Tavernier, E.; Moteki, M.; Koubbi, P. Lipid composition of the Antarctic fish Pleuragramma antarcticum. Influence of age class. Polar Sci. 2011, 5, 264–271. [Google Scholar] [CrossRef]
- Giraldo, C.; Mayzaud, P.; Tavernier, E.; Irisson, J.O.; Penot, F.; Becciu, J.; Chartier, A.; Boutoute, M.; Koubbi, P. Lipid components as a measure of nutritional condition in fish larvae (Pleuragramma antarcticum) in East Antarctica. Mar. Biol. 2013, 160, 877–887. [Google Scholar] [CrossRef]
- Giraldo, C.; Mayzaud, P.; Tavernier, E.; Boutoute, M.; Penot, F.; Koubbi, P. Lipid dynamics and trophic patterns in Pleuragramma antarctica life stages. Antarct. Sci. 2015, 27, 429–438. [Google Scholar] [CrossRef]
- Nachman, R.J. Unusual predominance of even-carbon hydrocarbons in an antarctic food chain. Lipids 1985, 20, 629–633. [Google Scholar] [CrossRef]
- Phleger, C.F.; Nichols, P.D.; Erb, E.; Williams, R. Lipids of the notothenioid fishes Trematomus spp. and Pagothenia borchgrevinki from East Antarctica. Polar Biol. 1999, 22, 241–247. [Google Scholar]
- Kamler, E.; Krasicka, B.; Rakusa-Suszczewski, S. Comparison of lipid content and fatty acid composition in muscle and liver of two notothenioid fishes from Admiralty Bay (Antarctica): An eco-physiological perspective. Polar Biol. 2001, 24, 735–743. [Google Scholar]
- Lund, E.D.; Sidell, B.D. Neutral lipid compositions of Antarctic fish tissues may reflect use of fatty acyl substrates by catabolic systems. Mar. Biol. 1992, 112, 377–382. [Google Scholar] [CrossRef]
- DeWitt, H.H.; Hopkins, T.L. Aspects of the diet of the Antarctic silverfish, Pleuragramma antarcticum. In Adaptations within Antarctic Ecosystems: Proceedings of the Third SCAR Symposium on Antarctic Biology; Llano, G.A., Ed.; Smithsonian Institution: Washington, DC, USA, 1977; pp. 557–568. [Google Scholar]
- Hubold, G. Stomach contents of the Antarctic silverfish Pleuragramma antarcticum from the southern and eastern Weddell Sea (Antarctica). Polar Biol. 1985, 5, 43–48. [Google Scholar] [CrossRef]
- Kellermann, A. Food and feeding ecology of postlarval and juvenile Pleuragramma antarcticum (Pisces; Notothenioidei) in the seasonal pack ice zone off the Antarctic Peninsula. Polar Biol. 1987, 7, 307–315. [Google Scholar] [CrossRef]
- Vacchi, M.; Pisano, E.; Ghigliotti, L. (Eds.) The Antarctic silverfish: A keystone species in a changing ecosystem. Advances in Polar Ecology; Springer: Berlin/Heidelberg, Germany, 2017; 314p. [Google Scholar]
- La Mesa, M.; Eastman, J.T.; Vacchi, M. The role of notothenioid fish in the food web of the Ross Sea shelf waters: A review. Polar Biol. 2004, 27, 321–338. [Google Scholar] [CrossRef]
- Eastman, J.T. Pleuragramma antarcticum (Pisces, Nototheniidae) as food for other fishes in McMurdo Sound, Antarctica. Polar Biol. 1985, 4, 155–160. [Google Scholar] [CrossRef]
- Takahashi, M.; Nemoto, T. The food of some Antarctic fish in the western Ross Sea in summer 1979. Polar Biol. 1984, 3, 237–239. [Google Scholar] [CrossRef]
- Kobyliansky, S.G.; Orlov, A.M.; Gordeeva, N.V. Composition of deepsea pelagic ichthyocenes of the Southern Atlantic, from waters of the range of the Mid-Atlantic and Walvis Ridges. J. Ichthyol. 2010, 50, 932–949. [Google Scholar] [CrossRef]
- Parker, M.L.; Fraser, W.R.; Ashford, J.; Patarnello, T.; Zane, L.; Torres, J.J. Assemblages of micronektonic fishes and invertebrates in a gradient of regional warming along the western Antarctic peninsula. J. Mar. Syst. 2015, 152, 18–41. [Google Scholar] [CrossRef]
- Ainley, D.G.; DeMaster, D.P. The upper trophic levels in polar marine ecosystems. In Polar Oceanography; Smith, I., Walker, O., Eds.; Academic Press: London, UK, 1990; pp. 599–630. [Google Scholar]
- Robison, B.H. What drives the diel vertical migrations of Antarctic midwater fish? J. Mar. Biol. Assoc. UK 2003, 83, 639–642. [Google Scholar] [CrossRef]
- Williams, T.D. The Penguins; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Gon, O.; Heemstra, P.C. (Eds.) Fishes of the Southern Ocean; JBL Smith Institute of Ichthyology: Grahamstown, South Africa, 1990. [Google Scholar]
- Folch, J.; Lees, M.; Sloan-Stanley, G.H. A simple method for the isolation and purification of total lipids animal tissue (for brain, liver and muscle). J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Tsyganov, E.P. Method for direct lipid methylation after TLC without elution with silica gel. Lab. Delo 1971, 8, 490–493. [Google Scholar]
- Kabakoff, R. R in Action: Data Analysis and Graphics with R; Volkova, P.A., Ed.; DMK Press: Moscow, Russia, 2014; 588p. [Google Scholar]
- Morozov, E.G.; Flint, M.V.; Orlov, A.M.; Frey, D.I.; Molodtsova, T.N.; Krechik, V.A.; Latushkin, A.A.; Salyuk, P.A.; Murzina, S.A.; Minin, K.V.; et al. Oceanographic and Ecosystem Studies in the Atlantic Sector of Antarctica (Cruise 87 of the Research Vessel Akademik Mstislav Keldysh). Oceanology 2022, 62, 825–827. [Google Scholar]
- Kreps, E.M. Lipids of Cellular Membranes. The Evolution of Lipids of the Brain. Adaptive Function of Lipids; Nauka: Leningrad, Russia, 1981. [Google Scholar]
- Sidorov, V.S. Environmental biochemistry of fishes. In Lipids; Shatunovskyi, M.I., Ed.; Nauka: Moscow, Russia, 1983; pp. 154–178. [Google Scholar]
- Rioux, V.; Catheline, D.; Beauchamp, E.; Le Bloc’h, J.; Pédrono, F.; Legrand, P. Substitution of dietary oleic acid for myristic acid increases the tissue storage of α-linolenic acid and the concentration of docosahexaenoic acid in the brain, red blood cells and plasma in the rat. Animal 2008, 2, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Hazel, J.R. Effects of temperature on the structure and metabolism of cell membranes in fish. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1984, 246, R460–R470. [Google Scholar] [CrossRef] [PubMed]
- Stillwell, W.; Wassall, S.R. Docosahexaenoic acid: Membrane properties of a unique fatty acid. Chem. Phys. Lipids 2003, 126, 1–27. [Google Scholar] [CrossRef]
- Escribá, P.V.; González-Ros, J.M.; Goñi, F.M.; Kinnunen, P.K.; Vigh, L.; Sánchez-Magraner, L.; Fernández, A.M.; Busquets, X.; Horváth, I.; Barceló-Coblijn, G. Membranes: A meeting point for lipids, proteins and therapies. J. Cell. Mol. Med. 2008, 12, 829–875. [Google Scholar] [CrossRef] [PubMed]
- Luckey, M. Membrane Structural Biology: With Biochemical and Biophysical Foundations; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Jones, P.G.; Inouye, M. The cold-shock response—A hot topic. Mol. Microbiol. 1994, 11, 811–818. [Google Scholar] [CrossRef]
- Murata, N.; Los, D.A. Membrane fluidity and temperature perception. Plant Physiol. 1997, 115, 875. [Google Scholar] [CrossRef]
- Tiku, P.E.; Gracey, A.Y.; Macartney, A.I.; Beynon, R.J.; Cossins, A.R. Cold-induced expression of Δ9-desaturase in carp by transcriptional and posttranslational mechanisms. Science 1996, 271, 815–818. [Google Scholar] [CrossRef]
- Coutteau, P.; Geurden, I.; Camara, M.R.; Bergot, P.; Sorgeloos, P. Review on the dietary effects of phospholipids in fish and crustacean larviculture. Aquaculture 1997, 155, 149–164. [Google Scholar] [CrossRef]
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. The lipids. In Fish Nutrition, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 181–257. [Google Scholar]
- Velansky, P.V.; Kostetsky, E.Y. Lipids of marine cold-water fishes. Russ. J. Mar. Biol. 2008, 34, 51–56. [Google Scholar] [CrossRef]
- Visentainer, J.V.; Noffs, M.D.A.; de Oliveira Carvalho, P.; de Almeida, V.V.; de Oliveira, C.C.; de Souza, N.E. Lipid content and fatty acid composition of 15 marine fish species from the southeast coast of Brazil. J. Am. Oil Chem. Soc. 2007, 84, 543–547. [Google Scholar] [CrossRef]
- Murzina, S.A. The Role of Lipids and Their Fatty Acid Components in Biochemical Adaptations of the Spotted Lumpen Leptoclinus maculatus F. of the Northwestern Coast of the Island Svalbard. Ph.D. Dissertation, Russian Academy of Sciences, Petrozavodsk, Russia, 2010; 184p. [Google Scholar]
- Pekkoeva, S.N. Changes in the Lipid Composition of the Muscles and the Lipid Sac of the Representative of the Arctic Ichthyofauna, Lumpen Spotted Leptoclinus maculatus in Postembryonic Development. Ph.D. Dissertation, Russian Academy of Sciences, Sankt-Peterburg, Russia, 2018; 164p. [Google Scholar]
- Murillo, E.; Rao, K.S.; Armando, A.D. The lipid content and fatty acid composition of four eastern central Pacific native fish species. J. Food Compos. Anal. 2014, 33, 1–5. [Google Scholar] [CrossRef]
- Murzina, S.A.; Nefedova, Z.A.; Falk-Petersen, S.; Ripatti, P.O.; Ruokolainen, T.R.; Pekkoeva, S.N.; Nemova, N.N. Lipid status of the two high latitude fish species, Leptoclinus maculatus and Lumpenus fabricii. Int. J. Mol. Sci. 2013, 14, 7048–7060. [Google Scholar] [CrossRef]
- Enig, M. The Importance of Saturated Fats for Biological Functions; Weston A. Prince Foundation: Washington, DC, USA, 2004. [Google Scholar]
- Legrand, P.; Rioux, V. The complex and important cellular and metabolic functions of saturated fatty acids. Lipids 2010, 45, 941–946. [Google Scholar] [CrossRef]
- Pepe, S.; McLennan, P.L. Dietary fish oil confers direct antiarrhythmic properties on the myocardium of rats. J. Nutr. 1996, 126, 34–42. [Google Scholar] [CrossRef]
- Daniels, R.A. Feeding ecology of some fishes of the Antarctic Peninsula. Fish Bull. 1982, 80, 575–588. [Google Scholar]
- Daniels, R.A.; Lipps, J.H. Distribution and ecology of fishes of the Antarctic Peninsula. J. Biogeogr. 1982, 9, 1–9. [Google Scholar] [CrossRef]
- Sampaio, J.L.; Gerl, M.J.; Klose, C.; Ejsing, C.S.; Beug, H.; Simons, K.; Shevchenko, A. Membrane lipidome of an epithelial cell line. Proc. Natl. Acad. Sci. USA 2011, 108, 1903–1907. [Google Scholar] [CrossRef]
- Trumble, S.J.; Kanatous, S.B. Fatty acid use in diving mammals: More than merely fuel. Front. Physiol. 2012, 3, 184. [Google Scholar] [CrossRef]
- Geoffroy, M.; Daase, M.; Cusa, M.; Darnis, G.; Graeve, M.; Santana Hernández, N.; Berge, J.; Renaud, P.E.; Cottier, F.; Falk-Petersen, S. Mesopelagic sound scattering layers of the high Arctic: Seasonal variations in biomass, species assemblages, and trophic relationships. Front. Mar. Sci. 2019, 6, 364. [Google Scholar] [CrossRef]
- Hopkins, T.L.; Torres, J.J. The zooplankton community in the vicinity of the ice edge, western Weddell Sea, March 1986. Polar Biol. 1988, 9, 79–87. [Google Scholar] [CrossRef]
- Lancraft, T.M.; Reisenbichler, K.R.; Robison, B.H.; Hopkins, T.L.; Torres, J.J. A krill-dominated micronekton and macrozooplankton community in Croker Passage, Antarctica with an estimate of fish predation. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2004, 51, 2247–2260. [Google Scholar] [CrossRef]
- Kelley, D.E.; Goodpaster, B.H.; Storlien, L. Muscle triglyceride and insulin resistance. Annu. Rev. Nutr. 2002, 22, 325. [Google Scholar] [CrossRef] [PubMed]
- Borkman, M.; Storlien, L.H.; Pan, D.A.; Jenkins, A.B.; Chisholm, D.J.; Campbell, L.V. The relation between insulin sensitivity and the fatty-acid composition of skeletal-muscle phospholipids. N. Engl. J. Med. 1993, 328, 238–244. [Google Scholar] [CrossRef]
- Vessby, B.; Tengblad, S.; Lithell, H. Insulin sensitivity is related to the fatty acid composition of serum lipids and skeletal muscle phospholipids in 70-year-old men. Diabetologia 1994, 37, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.A.; Lillioja, S.; Milner, M.R.; Kriketos, A.D.; Baur, L.A.; Bogardus, C.; Storlien, L.H. Skeletal muscle membrane lipid composition is related to adiposity and insulin action. J. Clin. Investig. 1995, 96, 2802–2808. [Google Scholar] [CrossRef]
- Murzina, S.A.; Dgebuadze, P.Y.; Pekkoeva, S.N.; Voronin, V.P.; Mekhova, E.S.; Thanh, N.T. Lipids and fatty acids of the gonads of Sea Urchin Diadema Setosum (Echinodermata) from the coastal area of the Nha Trang Bay, Central Vietnam. Eur. J. Lipid Sci. Technol. 2021, 123, 2000321. [Google Scholar] [CrossRef]
- Voronin, V.P.; Nemova, N.N.; Ruokolainen, T.R.; Artemenkov, D.V.; Rolskii, A.Y.; Orlov, A.M.; Murzina, S.A. Into the Deep: New Data on the Lipid and Fatty Acid Profile of Redfish Sebastes mentella Inhabiting Different Depths in the Irminger Sea. Biomolecules 2021, 11, 704. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 5th ed.; W.H. Freeman & Company: New York, NY, USA, 2008; 1294p. [Google Scholar]
- Sidell, B.D.; Hazel, J.R. Triacylglycerol lipase activities in tissues of Antarctic fishes. Polar Biol. 2002, 25, 517–522. [Google Scholar] [CrossRef]
- Crockett, E.L.; Sidell, B.D. Some pathways of energy metabolism are cold adapted in Antarctic fishes. Physiol. Zool. 1990, 63, 472–488. [Google Scholar] [CrossRef]
- Arts, M.T.; Kohler, C.C. Health and condition in fish: The influence of lipids on membrane competency and immune response. In Lipids in Aquatic Ecosystems; Springer: New York, NY, USA, 2009; pp. 237–256. [Google Scholar] [CrossRef]
- Lapin, V.I.; Shatunovsky, M.I. Features of composition, physiological and ecological value of fish lipids. Usp. Sovrem. Biol. 1981, 92, 380–394. [Google Scholar]
- Voronin, V.P.; Artemenkov, D.V.; Orlov, A.M.; Murzina, S.A. Lipids and Fatty Acids in Some Mesopelagic Fish Species: General Characteristics and Peculiarities of Adaptive Response to Deep-Water Habitat. J. Mar. Sci. Eng. 2022, 10, 949. [Google Scholar] [CrossRef]
- Pasternak, A.; Bugajska, J.; Szura, M.; Walocha, J.A.; Matyja, A.; Gajda, M.; Sztefko, K.; Gil, K. Biliary polyunsaturated fatty acids and telocytes in gallstone disease. Cell Transplant. 2017, 26, 125–133. [Google Scholar] [CrossRef]
- Tancharoenrat, P.; Zaefarian, F.; Ravindran, V. Composition of chicken gallbladder bile. Br. Poult. Sci. 2022, 63, 548–551. [Google Scholar] [CrossRef]
- Malagelada, J.R.; Longstreth, G.F.; Summerskill, W.H.J.; Go, V.L.W. Measurement of gastric functions during digestion of ordinary solid meals in man. Gastroenterology 1976, 70, 203–210. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Hagen, W.; Kattner, G.; Clarke, A.; Sargent, J. Lipids, trophic relationships, and biodiversity in Arctic and Antarctic krill. Can. J. Fish. Aquat. Sci. 2000, 57, 178–191. [Google Scholar] [CrossRef]
- Dalsgaard, J.; John, M.S.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [CrossRef]
- Graeve, M.; Kattner, G.; Piepenburg, D. Lipids in Arctic benthos: Does the fatty acid and alcohol composition reflect feeding and trophic interactions? Polar Biol. 1997, 18, 53–61. [Google Scholar] [CrossRef]
- Parrish, C.C. Lipids in marine ecosystems. ISRN Oceanogr. 2013, 9, 604045. [Google Scholar] [CrossRef]
- Hopkins, T.L. Midwater food web in McMurdo Sound, Ross Sea, Antarctica. Mar. Biol. 1987, 96, 93–106. [Google Scholar] [CrossRef]
- Mintenbeck, K. Trophic Interactions within High Antarctic Shelf Communities—Food Web Structure and the Significance of Fish. Ph.D. Thesis, University of Bremen, Bremen, Germany, 2008; 246p. [Google Scholar]
- Moreno, C.A.; Rueda, T.; Asencio, G. The trophic niche of Pleuragramma antarcticum in the Bransfield Strait, Antarctica: Quantitative comparison with other areas of the Southern Ocean. Ser. Cient. INACH 1986, 35, 101–117. [Google Scholar]
- Olaso, I.; Lombarte, A.; Velasco, F. Daily ration of antarctic silverfish (Pleuragramma antarcticum Boulenger, 1902) in the eastern Weddell Sea. Sci. Mar. 2004, 68, 419–424. [Google Scholar] [CrossRef][Green Version]
- Williams, R. Trophic relationship between pelagic fish and euphausiids in Antarctic waters. In Antarctic Nutrient Cycles and Food Webs; Siegfried, W.R., Condy, P.R., Laws, R.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 452–459. [Google Scholar] [CrossRef]
- Linkowski, T.B.; Presler, P.; Zukowski, C. Food habits of nototheniid fishes (Nototheniidae) in Admiralty Bay (King George Island, South Shetland Islands). Pol. Polar Res. 1983, 4, 79–95. [Google Scholar]
- Pinkerton, M.H. Diet and trophic ecology of adult Antarctic silverfish (Pleuragramma antarctica). In The Antarctic Silverfish: A Keystone Species in a Changing Ecosystem; Advances in Polar Ecology; Vacchi, M., Pisano, E., Ghigliotti, L., Eds.; Springer: Cham, Switzerland, 2017; Volume 3, pp. 93–111. [Google Scholar] [CrossRef]
- Graeve, M.; Dauby, P.; Scailteur, Y. Combined lipid, fatty acid and digestive tract content analyses: A penetrating approach to estimate feeding modes of Antarctic amphipods. Polar Biol. 2001, 24, 853–862. [Google Scholar] [CrossRef]
- Parzanini, C.; Colombo, S.M.; Kainz, M.J.; Wacker, A.; Parrish, C.C.; Arts, M.T. Discrimination between freshwater and marine fish using fatty acids: Ecological implications and future perspectives. Environ. Rev. 2020, 28, 546–559. [Google Scholar] [CrossRef]
- Bottino, N.R. Lipid composition of two species of Antarctic krill: Euphausia superba and E. crystallorophias. Comp. Biochem. Physiol. Part B Comp. Biochem. 1975, 50, 479–484. [Google Scholar] [CrossRef]
- Kattner, G.; Hagen, W. Lipid metabolism of the Antarctic euphausiid Euphausia crystallorophias and its ecological implications. Mar. Ecol. Prog. Ser. 1998, 170, 203–213. [Google Scholar] [CrossRef]
- Carlig, E.; Christiansen, J.S.; Di Blasi, D.; Ferrando, S.; Pisano, E.; Vacchi, M.; O’Driscoll, R.L.; Ghigliotti, L. Midtrophic fish feeding modes at the poles: An ecomorphological comparison of polar cod (Boreogadus saida) and Antarctic silverfish (Pleuragramma antarctica). Polar Biol. 2021, 44, 1629–1642. [Google Scholar] [CrossRef]
- Lazzaro, X.A. A review of planktivorous fishes: Their evolution, feeding behaviours, selectivities, and impacts. Hydrobiologia 1987, 146, 97–167. [Google Scholar] [CrossRef]
- Liem, K.F. Acquisition of energy by teleosts: Adaptive mechanisms and evolutionary patterns. In Environmental Physiology of Fishes; Ali, M.A., Ed.; Plenum Press: New York, NY, USA, 1980; pp. 299–334. [Google Scholar]
- Hagen, W.; Van Vleet, E.S.; Kattner, G. Seasonal lipid storage as overwintering strategy of Antarctic krill. Mar. Ecol. Prog. Ser. 1996, 134, 85–89. [Google Scholar] [CrossRef]
- Hagen, W.; Kattner, G. Lipid metabolism of the Antarctic euphausiid Thysanmssa macrura and its ecological implications. Limnol. Oceanogr. 1998, 43, 1894–1901. [Google Scholar] [CrossRef]
- Hagen, W.; Kattner, G.; Terbrüggen, A.; Van Vleet, E.S. Lipid metabolism of the Antarctic krill Euphausia superba and its ecological implications. Mar. Biol. 2001, 139, 95–104. [Google Scholar] [CrossRef]
- Hubold, G.; Hagen, W. Seasonality of feeding and lipid content of Pleuragramma antarcticum (Nototheniidae) in the southern Weddell Sea. In Antarctic Communities: Species, Structure and Survival; Battaglia, B., Valencia, J., Walton, D.W.H., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 277–283. [Google Scholar]
- Kattner, G.; Albers, C.; Graeve, M.; Schnack-Schiel, S.B. Fatty acid and alcohol composition of the small polar copepods, Oithona and Oncaea: Indication on feeding modes. Polar Biol. 2003, 26, 666–671. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Sargent, J.R.; Lønne, O.J.; Timofeev, S. Functional biodiversity of lipids in Antarctic zooplankton: Calanoides acutus, Calanus propinquus, Thysanoessa macrura and Euphausia crystallorophias. Polar Biol. 1999, 21, 37–47. [Google Scholar] [CrossRef]
- Graeve, M.; Hagen, W.; Kattner, G. Herbivorous or omnivorous? On the significance of lipid compositions as trophic markers in Antarctic copepods. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1994, 41, 915–924. [Google Scholar] [CrossRef]
- Kattner, G.; Graeve, M.; Hagen, W. Ontogenetic and seasonal changes in lipid and fatty acid/alcohol compositions of the dominant Antarctic copepods Calanus propinquus, Calanoides acutus and Rhincalanus gigas. Mar. Biol. 1994, 118, 637–644. [Google Scholar] [CrossRef]
- Kabeya, N.; Fonseca, M.M.; Ferrier, D.E.; Navarro, J.C.; Bay, L.K.; Francis, D.S.; Tocher, D.R.; Castro, L.F.C.; Monroig, Ó. Genes for de novo biosynthesis of omega-3 polyunsaturated fatty acids are widespread in animals. Sci. Adv. 2018, 4, eaar6849. [Google Scholar] [CrossRef]
- Monroig, Ó.; Kabeya, N. Desaturases and elongases involved in polyunsaturated fatty acid biosynthesis in aquatic invertebrates: A comprehensive review. Fish. Sci. 2018, 84, 911–928. [Google Scholar] [CrossRef]
- Monroig, Ó.; Shu-Chien, A.C.; Kabeya, N.; Tocher, D.R.; Castro, L.F.C. Desaturases and elongases involved in long-chain polyunsaturated fatty acid biosynthesis in aquatic animals: From genes to functions. Prog. Lipid Res. 2022, 86, 101157. [Google Scholar] [CrossRef]
- Watanabe, T. Importance of docosahexaenoic acid in marine larval fish. J. World Aquac. Soc. 1993, 24, 152–161. [Google Scholar] [CrossRef]
- Kozlov, A.N. Relationship between intensity and periods of protein increase and fat accumulation in the marbled Notothenia, Notothenia rossi marmorata (Nototheniidae). J. Ichthyol. 1981, 21, 99–102. [Google Scholar]
- Sargent, J.R.; Bell, J.G.; Bell, M.V.; Henderson, R.J.; Tocher, D.R. Requirement criteria for essential fatty acids. J. Appl. Ichthyol. 1995, 11, 183–198. [Google Scholar] [CrossRef]
- Kaur, G.; Cameron-Smith, D.; Garg, M.; Sinclair, A.J. Docosapentaenoic acid (22:5n-3): A review of its biological effects. Prog. Lipid Res. 2011, 50, 28–34. [Google Scholar] [CrossRef]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.; Ciappolino, V.; Agostoni, C. DHA effects in brain development and function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Mourente, G. Accumulation of DHA (docosahexaenoic acid; 22:6n-3) in larval and juvenile fish brain. In The Big Fish Bang; Institute of Marine Research: Bergen, Norway, 2003; pp. 239–248. [Google Scholar]
- Salem, N., Jr.; Litman, B.; Kim, H.Y.; Gawrisch, K. Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids 2001, 36, 945–959. [Google Scholar] [CrossRef]
- Bell, M.V.; Dick, J.R. Molecular species composition of the major diacyl glycerophospholipids from muscle, liver, retina and brain of cod (Gadus morhua). Lipids 1991, 26, 565–573. [Google Scholar] [CrossRef]
- Tocher, D.R.; Harvie, D.G. Fatty acid compositions of the major phosphoglycerides from fish neural tissues: (n-3) and (n-6) polyunsaturated fatty acids in rainbow trout (Salmo gairdneri) and cod (Gadus morhua) brains and retinas. Fish Physiol. Biochem. 1988, 5, 229–239. [Google Scholar] [CrossRef]
- Eady, T.L.; Khoutorova, A.; Obenaus, A.; Mohd-Yusof, A.; Bazan, N.G.; Belayev, L. Docosahexaenoic acid complexed to albumin provides neuroprotection after experimental stroke in aged rats. Neurobiol. Dis. 2014, 62, 1–7. [Google Scholar] [CrossRef]
- Shulman, G.E.; Yuneva, T.V. The role of docosahexaenoic acid in fish adaptations (review). Ecol. Physiol. Biochem. Aquat. Anim. 1990, 26, 43–51. [Google Scholar]
- Stoknes, I.S.; Okland, H.M.; Falch, E.; Synnes, M. Fatty acid andlipid class composition in eyes and brain from teleosts andelasmobranchs. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 138, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Zhou, Y.; Wu, H.; Luo, Y.; Shen, H. Lipid content and fatty acid profile of muscle, brain and eyes of seven freshwater fish: A comparative study. J. Am. Oil Chem. Soc. 2014, 91, 795–804. [Google Scholar] [CrossRef]
- Sambra, V.; Echeverria, F.; Valenzuela, A.; Chouinard-Watkins, R.; Valenzuela, R. Docosahexaenoic and arachidonic acids as neuroprotective nutrients throughout the life cycle. Nutrients 2021, 13, 986. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).