
Eranthis Salisb. (Ranunculaceae) in South Siberia: Insights into Phylogeography and Taxonomy



Abstract
:1. Introduction
2. Materials and Methods
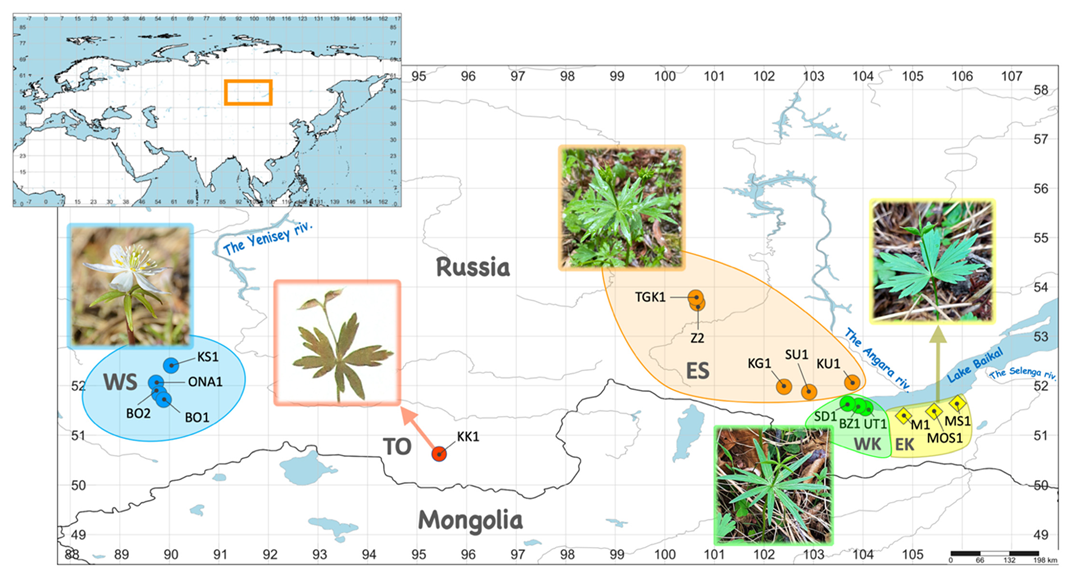
2.1. Plant Material Collection
2.2. DNA Isolation
2.3. PCR, Cloning, and Sequencing
2.4. Sequence Alignment and Phylogenetic Analysis
3. Results
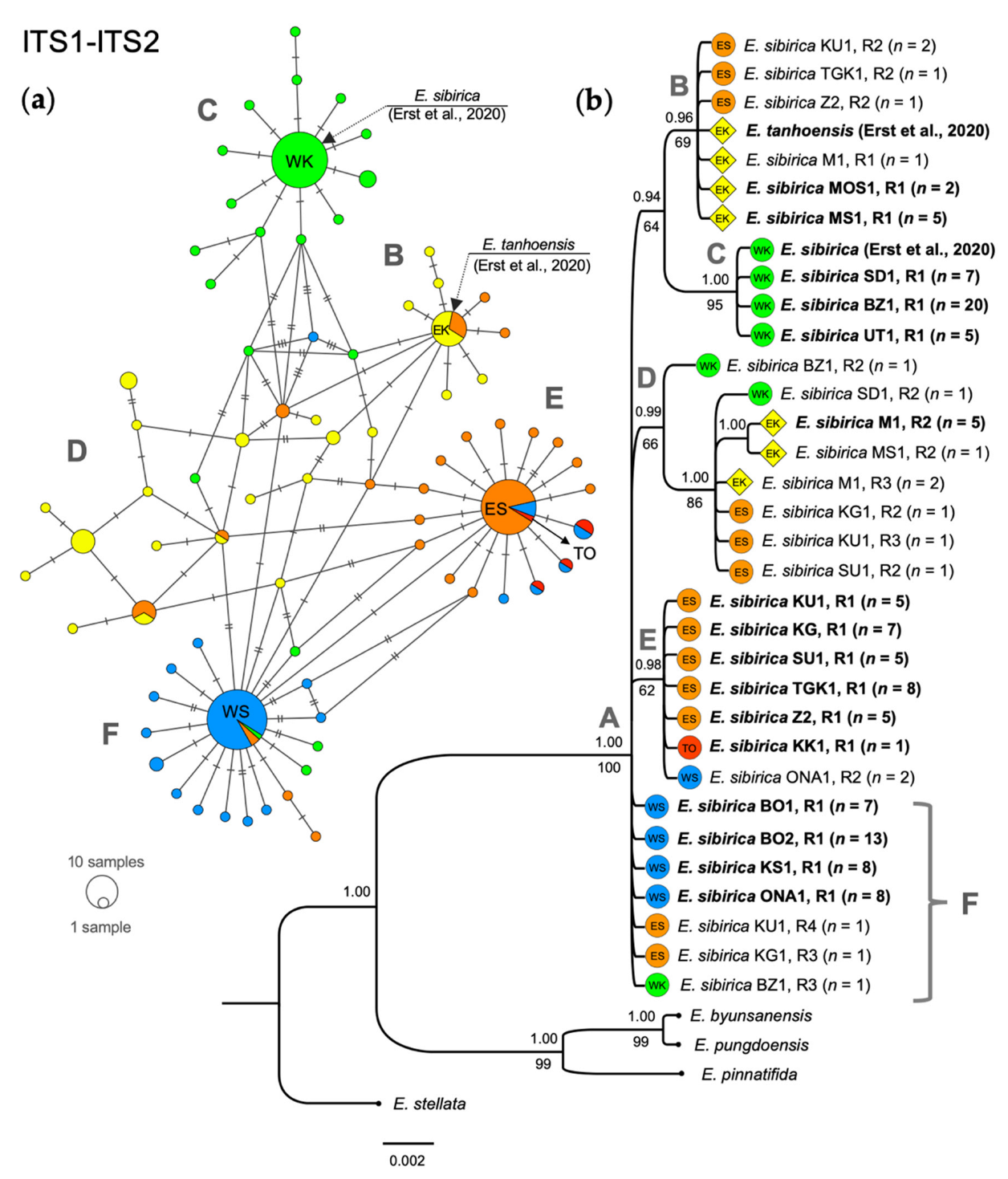
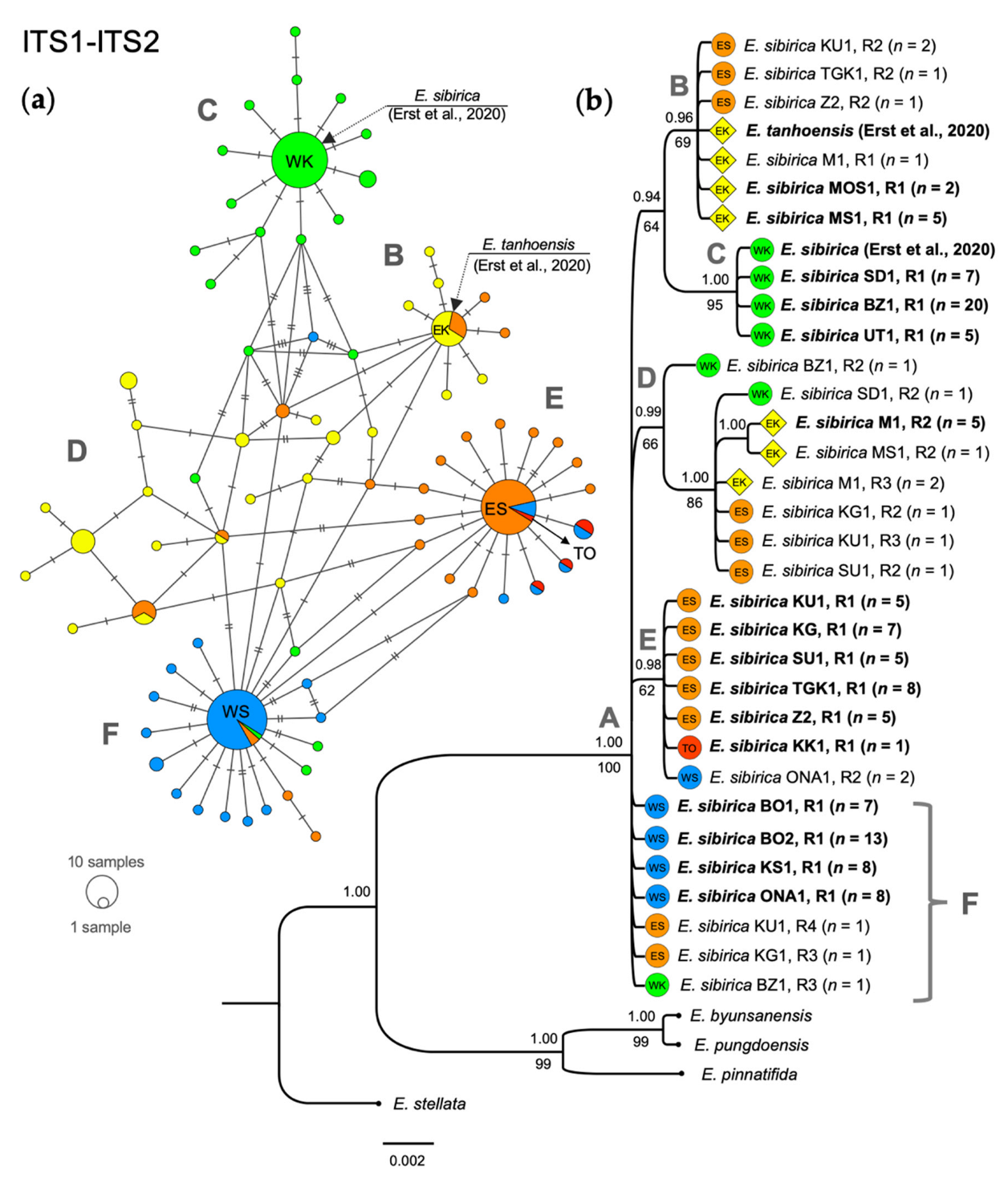
3.1. Phylogenetic Analysis Based on Nuclear DNA
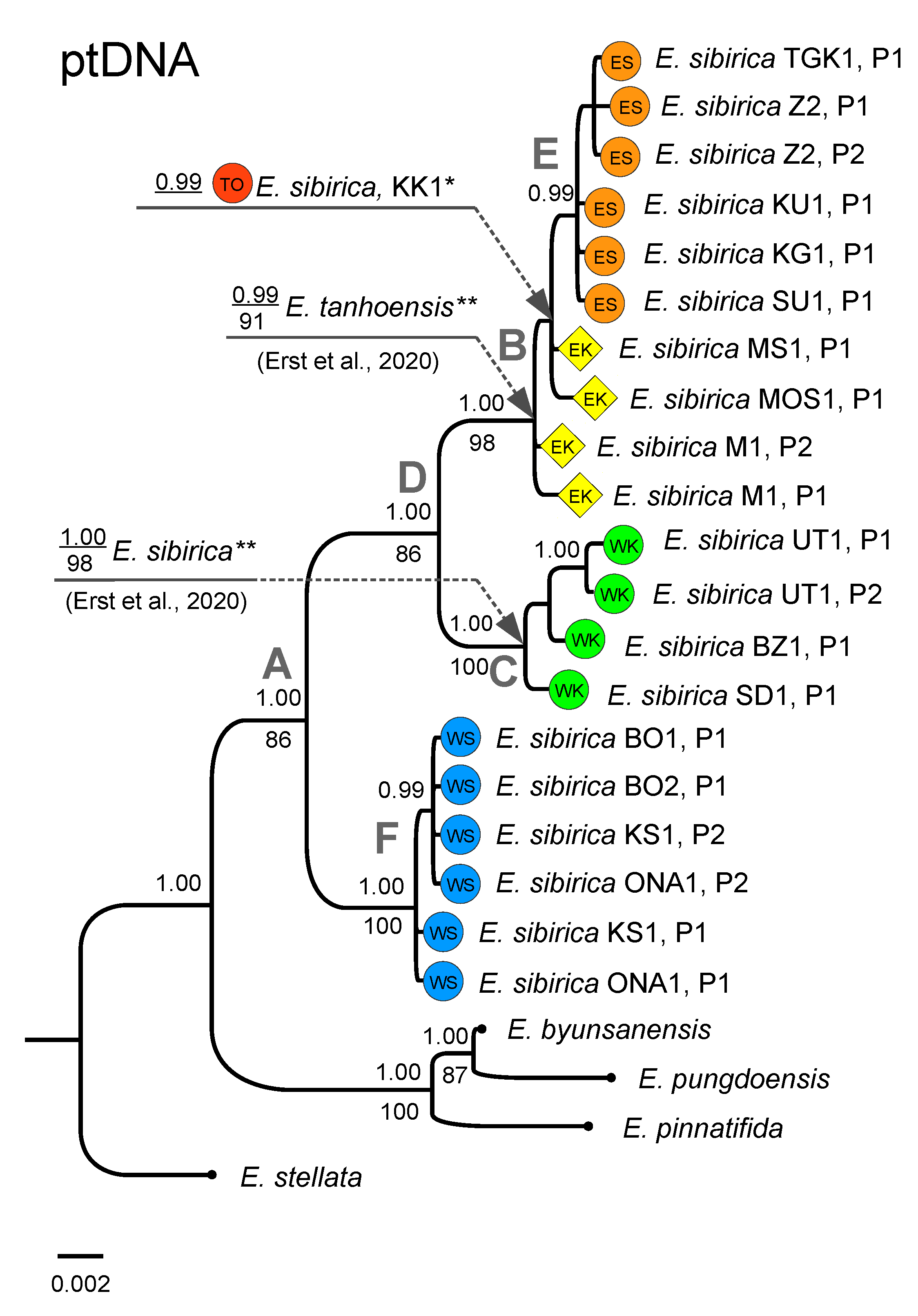
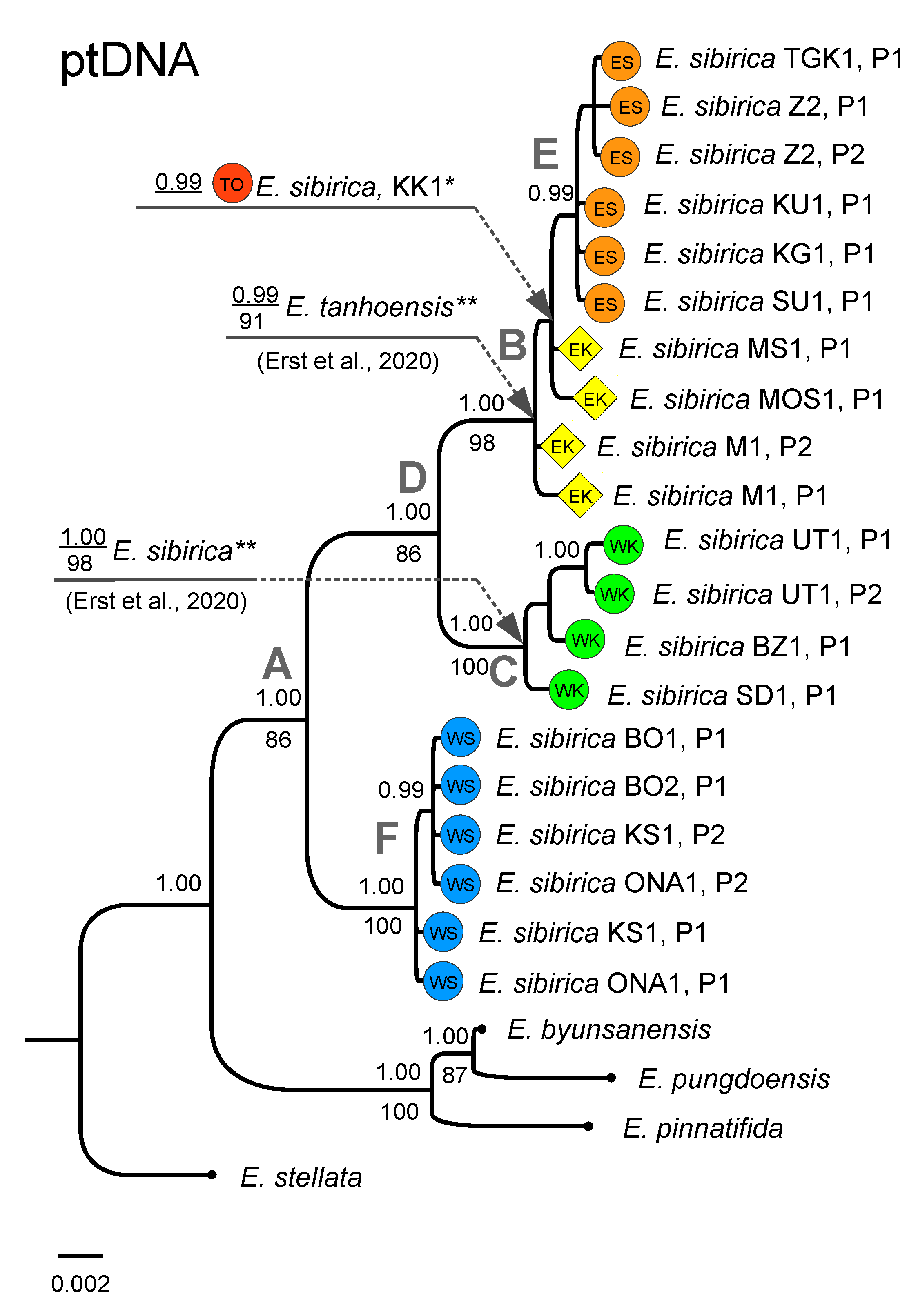
3.2. Phylogenetic Analysis Based on Plastid DNA
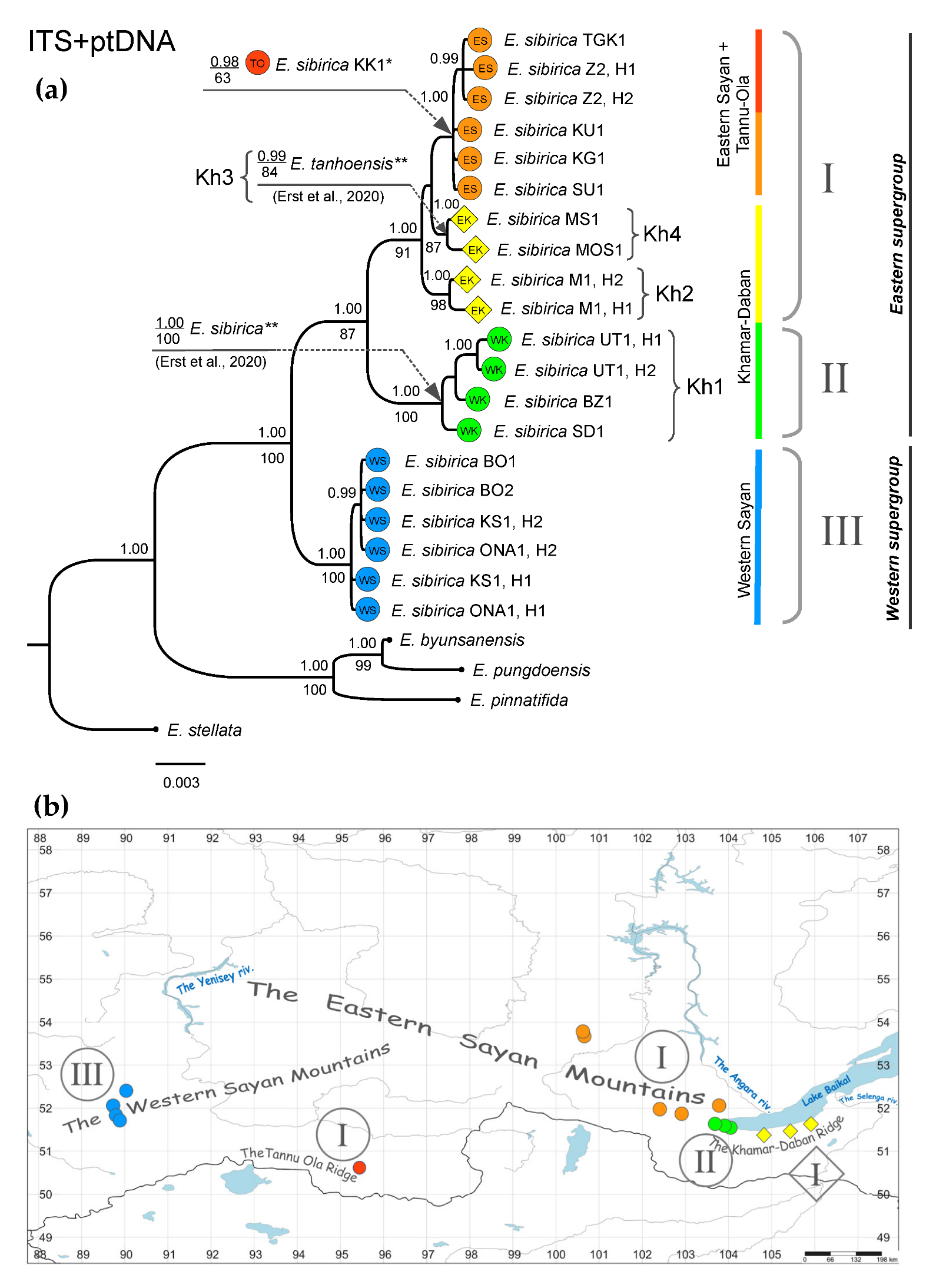
3.3. Combined Phylogenetic Analysis
4. Discussion
4.1. Phylogeographic Patterns of Eranthis in South Siberia
4.2. Phylogeny of Siberian Eranthis and Taxonomic Status of E. tanhoensis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, C.S.; Yeau, S.H.; Lee, N.S. Taxonomic Status and Genetic Variation of Korean Endemic Plants, Eranthis byunsanensis and Eranthis pungdoensis (Ranunculaceae) based on nrDNA ITS and cpDNA Sequences. J. Plant Biol. 2012, 55, 165–177. [Google Scholar] [CrossRef]
- Oh, A.; Oh, B.-U. The speciation history of northern- and southern-sourced Eranthis (Ranunculaceae) species on the Korean peninsula and surrounding areas. Ecol. Evol. 2019, 9, 2907–2919. [Google Scholar] [CrossRef]
- Park, S.Y.; Jeon, M.J.; Ma, S.H.; Wahlsteen, E.; Amundsen, K.; Kim, J.H.; Suh, J.K.; Chang, J.S.; Joung, Y.H. Phylogeny and genetic variation in the genus Eranthis using NrITS and CpIS single nucleotide polymorphisms. Hortic. Environ. Biotechnol. 2019, 60, 239–252. [Google Scholar] [CrossRef]
- Xiang, K.-L.; Erst, A.S.; Yang, J.; Peng, H.-W.; Ortiz, R.D.C.; Jabbour, F.; Erst, T.V.; Wang, W. Biogeographic diversification of Eranthis (Ranunculaceae) reflects the geological history of the three great Asian plateaus. Proc. R. Soc. B Boil. Sci. 2021, 288, 20210281. [Google Scholar] [CrossRef] [PubMed]
- Erst, A.S.; Sukhorukov, A.P.; Mitrenina, E.Y.; Skaptsov, M.V.; Kostikova, V.A.; Chernisheva, O.A.; Troshkina, V.; Kushunina, M.; Krivenko, D.A.; Ikeda, H.; et al. An integrative taxonomic approach reveals a new species of Eranthis (Ranunculaceae) in North Asia. PhytoKeys 2020, 140, 75–100. [Google Scholar] [CrossRef] [PubMed]
- Chepinoga, V.V.; Mishina, A.V.; Protopopova, M.V.; Pavlichenko, V.V.; Bystrov, S.O.; Vilor, M.A. New Data on Distribution of Several Nemoral Relict Plant Species on the Foothills of the Khamar-Daban Ridge (Southern Baikal). Bot. Zhurnal 2015, 100, 478–489. (In Russian) [Google Scholar]
- Timokhina, S.A.; Frizen, N.V.; Vlasova, N.V.; Zuev, V.V.; Kovtonyuk, N.K.; Baykov, K.S. Flora Sibiriae. Portulacaceae—Ranunculaceae; Malyshev, L.I., Peshkova, G.A., Eds.; Nauka: Novosibirsk, Russia, 1993; Volume 6, ISBN 5-02-030132-9. (In Russian) [Google Scholar]
- Chepinoga, V.V.; Protopopova, M.V.; Pavlichenko, V.V.; Dudov, S.V. Habitat Distribution Patterns of Nemoral Relict Plant Species on the Khamar-Daban Ridge (the South of Eastern Siberia) According to Grid Mapping Data. Russ. J. Ecol. 2021, 52, 212–222. [Google Scholar] [CrossRef]
- Shaulo, D.N.; Zykova, E.Y.; Shmakov, A.I.; Tupitsyna, N.N.; Molokova, N.I.; Artemov, I.A.; An’kova, T.V.; Sonnikova, A.E.; Shanmak, R.B.; Saak, N.V.; et al. Floristic fndings in south of Central Siberia: Krasnoyarsk Territory, Republics of Khakassia and Tuva. Turczaninowia 2019, 22, 80–93. (In Russian) [Google Scholar] [CrossRef]
- Protopopova, M.V.; Pavlichenko, V.V.; Gnutikov, A.A.; Adelshin, R.V.; Chepinoga, V.V. Application of Genetic Markers for Ecological Status Assessment of the Relict Plant Species of Baikal Siberia. RUDN J. Ecol. Life Safety 2015, 4, 28–36. (In Russian) [Google Scholar]
- Protopopova, M.V.; Pavlichenko, V.V.; Chepinoga, V.V. Some Aspects of Genetic Polymorphism in Two Relict Plant Species from Baikal Siberia. In Proceedings of the Modern Achivements in Population, Evolutionary, and Ecological Genetics: International Symposium, Vladivostoc—Vostok Marine Biological Station, Vladivostok, Russia, 1–10 September 2015; p. 57. [Google Scholar]
- Protopopova, M.V.; Pavlichenko, V.V.; Konovalov, A.D.; Chepinoga, V.V. The Study of the Historical Dynamics of Some Relic Plant Species in Baikal Siberia during Global Climatic Change Using Molecular Genetic Markers. In Proceedings of the All-Russian Conference with International Participation and School for Young Scientists: Factors of Plant and Microorganism Resistance in Extremal Nature Conditions and Technogenic Environment, Irkutsk, Russia, 12–15 September 2016; V.B. Sochava Institute of Geography SB RAS Publishing House: Irkutsk, Russia, 2016; pp. 250–251. (In Russian). [Google Scholar]
- Bobo-Pinilla, J.; Salmerón-Sánchez, E.; Mendoza-Fernández, A.J.; Mota, J.F.; Peñas, J. Conservation and Phylogeography of Plants: From the Mediterranean to the Rest of the World. Diversity 2022, 14, 78. [Google Scholar] [CrossRef]
- Shorthouse, D.P. SimpleMappr, an Online Tool to Produce Publication-Quality Point Maps. Available online: https://www.simplemappr.net (accessed on 27 July 2022).
- Polozhii, A.V.; Krapivkina, E.D. Relikty Tretichnyh Shirokolistvennyh Lesov vo Flore Sibiri [Relics of Tertiary Deciduous Forests in the Flora of Siberia]; Tomsk University Press: Tomsk, Russia, 1985. (In Russian) [Google Scholar]
- Krestov, P.V.; Barkalov, V.Y.; Omelko, A.M.; Yakubov, V.V.; Nakamura, Y.; Sato, K. Relic Vegetation Complexes in the Modern Refugia of Northeast Asia. Komarovskie Chtenia [V. L. Komarov Memorial Lectures] 2009, 56, 5–63. (In Russian) [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Utelli, A.; Roy, B.; Baltisberger, M. Molecular and morphological analyses of European Aconitum species (Ranunculaceae). Plant Syst. Evol. 2000, 224, 195–212. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.A.; Simpson, B.B. Paraphyly of Tarasa (Malvaceae) and Diverse Origins of the Polyploid Species. Syst. Bot. 2003, 28, 723–737. [Google Scholar]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef]
- Lahr, D.J.G.; Katz, L.A. Reducing the impact of PCR-mediated recombination in molecular evolution and environmental studies using a new-generation high-fidelity DNA polymerase. BioTechniques 2009, 47, 857–866. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T.-A. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Jukes, T.H.; Cantor, C.R. Evolution of Protein Molecules; Academic Press: New York, NY, USA, 1969; Volume 3, pp. 21–132. [Google Scholar]
- Rambaut, A. FigTree: Tree Figure Drawing Tool, Version 1.4.3. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 15 July 2022).
- Leigh, J.W.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Ribosomal RNA genes in plants: Variability in copy number and in the intergenic spacer. Plant Mol. Biol. 1987, 9, 509–520. [Google Scholar] [CrossRef]
- Xu, B.; Zeng, X.-M.; Gao, X.-F.; Jin, D.-P.; Zhang, L.-B. ITS non-concerted evolution and rampant hybridization in the legume genus Lespedeza (Fabaceae). Sci. Rep. 2017, 7, 40057. [Google Scholar] [CrossRef]
- Belov, A.V.; Bezrukova, E.V.; Sokolova, L.P.; Abzayeva, A.A.; Letunova, P.P.; Fisher, E.E.; Orlova, L.A. Vegetation of the Baikal Region as an Indicator of Global and Regional Changes in Natural Conditions of North Asia in the Late Cainozoic. Geogr. Nat. Resour. 2006, 6, 5–18. (In Russian) [Google Scholar]
- Alter, S.P. K Istorii Formirovaniya Doliny Eniseya [to the History of the Yenisey Valley Formation]. Doklady instituta geografii Sibiri i Dal’nego Vostoka [Reports of the Institute of Geography of Siberia and the Far East]. 1965, 8, 38–44. (In Russian) [Google Scholar]
- Chepinoga, V.V.; Protopopova, M.V.; Pavlichenko, V.V. Detection of the most probable Pleistocene microrefugia on the northern macroslope of the Khamar-Daban Ridge (Southern Prebaikalia). Contemp. Probl. Ecol. 2017, 10, 38–42. [Google Scholar] [CrossRef]
- Protopopova, M.V.; Pavlichenko, V.V.; Orlova, D.A.; Chepinoga, V.V. Phylogeographic Structure of Anemone baicalensis (Ranunculaceae) Based on Plastid DNA Polymorphism (trnL-trnF) as an Evidence of Pleistocene Microrefugia Existence on the Khamar-Daban Ridge (Southern Baikal Region). Bull. Irkutsk State Univ. Ser. Biol. Ecol. 2019, 30, 3–15. [Google Scholar] [CrossRef]
- Malyschev, L.I. Flora Alpina Montium Sajanensium Orientalium; Nauka: Moscow, Russia; Leningrad, Russia, 1965. (In Russian) [Google Scholar]
- Krasnoborov, I.M. Flora Alpina Montium Sajanensium Occidentalium; Nauka: Moscow, Russia; Leningrad, Russia, 1976. (In Russian) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality Abbr. | Herbarium Voucher Information | Coordinates, Altitude * |
|---|---|---|
| BO1 | Russia, the Republic of Khakassia, Tashtypskiy Raion, the Western Sayan Mts., the Bol’shoy On Riv., 15 June 2018, V. Chepinoga, V. Pavlichenko, M. Protopopova (IRKU085077) | 51.715213° N, 089.885315° E, 2080 m alt. |
| BO2 | Russia, the Republic of Khakassia, Tashtypskiy Raion, the Western Sayan Mts., the Bol’shoy On Riv., 15 June 2018, V. Pavlichenko, M. Protopopova, V. Chepinoga (IRKU045394) | 51.837447° N, 089.794825° E, 1363 m alt. |
| BZ1 | Russia, Irkutskaya Oblast’, Slyudyanskiy Raion, the Khamar-Daban Ridge, the Bezymyannaya Riv., 03 June 2022, M. Protopopova, V. Pavlichenko (IRKU085082) | 51.59373° N, 103.90829° E, 461 m alt. |
| KG1 | Russia, the Republic of Buryatia, Tunkinskiy Raion, the Eastern Sayan Mts, the Kyngyrga Riv., 25 May 2019, M. Protopopova, V. Pavlichenko, V. Chepinoga, V. Belova (IRKU045872) | 51.976379° N, 102.406746° E, 1390 m alt. |
| KK1 | Russia, the Republic of Tyva, Tes-Khemskiy kozhuun, the Tannu-Ola Ridge, 10 km E of Shuurmak setl., the Kuskunug-Khem Riv., 6 August 1979, M. Danilov, A. Kritsin (NSK0003650) | unknown, 1350 m alt. |
| KS1 | Russia, the Republic of Khakassia, Tashtypskii Raion, the Western Sayan Mts., the Karasibo Riv., 16 June 2018, V. Pavlichenko, M. Protopopova, V. Chepinoga (IRKU045389) | 52.404736° N, 090.034289° E, 603 m alt. |
| KU1 | Russia, Irkutskaya Oblast’, Shelekhovskiy Raion, the Eastern Sayan Mts. foothills, around Shamanka setl., the Kuitun Riv., 23 May 2022, M. Protopopova, V. Pavlichenko, P. Nelyubina (IRKU085078) | 52.060886° N, 103.782335° E, 459 m alt. |
| M1 ** | Russia, the Republic of Buryatia, Kabanskiy Raion, the Khamar-Daban Ridge, the Bol’shoy Mamay Riv., 2 June 2022, M. Protopopova, V. Pavlichenko, P. Nelyubina (IRKU085079) | 51.43050° N, 104.80330° E, 542 m alt. |
| MOS1 ** | Russia, the Republic of Buryatia, Kabanskiy Raion, the Khamar-Daban Ridge, the watershed of the Osinovka and Ushakovka rivers, 1 July 2016, M. Protopopova, V. Pavlichenko (IRKU045436) | 51.52742° N, 105.42017° E, 1507 m alt. |
| MS1 ** | Russia, the Republic of Buryatia, Kabanskiy Raion, the Khamar-Daban Ridge, around Babushkin town, the Mysovka Riv., 31 May 2022, M. Protopopova, V. Pavlichenko (IRKU085081) | 51.67841° N, 105.89572° E, 513 m alt. |
| ONA1 | Russia, the Republic of Khakassia, Tashtypskii Raion, the Western Sayan Mts., the Ona Riv., 16 June 2018, V. Pavlichenko, V. Chepinoga, M. Protopopova (IRKU085084) | 52.057791° N, 089.730494° E, 784 m alt. |
| SD1 | Russia, Irkutskaya Oblast’, Slyudyanskiy Raion, the Khamar-Daban Ridge, around Slyudyanka town, the Slyudyanka Riv., 5 June 2021, M. Protopopova, V. Pavlichenko (IRKU085083) | 51.63547° N, 103.68811° E, 540 m alt. |
| SU1 | Russia, the Republic of Buryatia, Tunkinskiy Raion, the Eastern Sayan Mts, the Sagan-Ugun Riv., 26 May 2019, M. Protopopova, V. Pavlichenko, V. Chepinoga, V. Belova (IRKU045867) | 51.870508° N, 102.913274° E, 818 m alt. |
| TGK1 | Russia, Irkutskaya Oblast’, Kuytunskiy Raion, the Eastern Sayan Mts. foothills, the Kireyskaya Tagna Riv., 14 June 2015, V. Chepinoga, E. Zolotovskaya, R. Fitz, M. Protopopova *** (IRKU045864) | 53.78218° N, 100.62312° E, 561 m alt. |
| UT1 | Russia, Irkutskaya Oblast’, Slyudyanskiy Raion, the Khamar-Daban Ridge, the Utulik Riv., 30 May 2022, M. Protopopova, V. Pavlichenko (IRKU085080) | 51.54594° N, 104.04675° E, 453 m alt. |
| Z2 | Russia, Irkutskaya Oblast’, Tulunskiy Raion, the Eastern Sayan Mts. foothills, the Zima Riv., 15 June 2015, M. Protopopova, V. Chepinoga (IRKU085085) | 53.67042° N, 100.65604° E 610 m alt. |
| Species Name Used in This Study | Species Name According to Erst et al. [5] | Locality | GenBank Accession Numbers | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Ribotype | Plastotype | Ref. | |||||||
| ITS | trnL | trnL-trnF | trnH-psbA | ||||||
| E. byunsanensis | – | – | – | JF505768.1 | – | JF505894.1 | JF505894.1 | JF505810.1 | [1] |
| E. sibirica | E. sibirica | BO1 | R1 | OP380051 | P1 | OP380801 | OP380822 | OP380843 | this study |
| E. sibirica | E. sibirica | BO2 | R1 | OP380052 | P1 | OP380802 | OP380823 | OP380844 | this study |
| E. sibirica | E. sibirica | BZ1 | R1 | OP380053 | P1 | OP380803 | OP380824 | OP380845 | this study |
| E. sibirica | E. sibirica | BZ1 | R2 | OP380054 | – | – | – | – | this study |
| E. sibirica | E. sibirica | BZ1 | R3 | OP380055 | – | – | – | – | this study |
| E. sibirica | E. sibirica | KG1 | R1 | OP380056 | P1 | OP380804 | OP380825 | OP380846 | this study |
| E. sibirica | E. sibirica | KG1 | R2 | OP380057 | – | – | – | – | this study |
| E. sibirica | E. sibirica | KG1 | R3 | OP380058 | – | – | – | – | this study |
| E. sibirica | E. sibirica | KK1 | R1 | OP380059 | P1 | OP380805 | OP380826 | no product | this study |
| E. sibirica | E. sibirica | KS1 | R1 | OP380060 | P1 | OP380806 | OP380827 | OP380847 | this study |
| E. sibirica | E. sibirica | KS1 | – | – | P2 | OP380807 | OP380828 | OP380848 | this study |
| E. sibirica | E. sibirica | KU1 | R1 | OP380061 | P1 | OP380808 | OP380829 | OP380849 | this study |
| E. sibirica | E. sibirica | KU1 | R2 | OP380062 | – | – | – | – | this study |
| E. sibirica | E. sibirica | KU1 | R3 | OP380063 | – | – | – | – | this study |
| E. sibirica | E. sibirica | KU1 | R4 | OP380064 | – | – | – | – | this study |
| E. sibirica | E. tanhoensis | M1 | R1 | OP380065 | P1 | OP380809 | OP380830 | OP380850 | this study |
| E. sibirica | E. tanhoensis | M1 | R2 | OP380066 | P2 | OP380810 | OP380831 | OP380851 | this study |
| E. sibirica | E. tanhoensis | M1 | R3 | OP380067 | – | – | – | – | this study |
| E. sibirica | E. tanhoensis | MOS1 | R1 | OP380068 | P1 | OP380811 | OP380832 | OP380852 | this study |
| E. sibirica | E. tanhoensis | MS1 | R1 | OP380069 | P1 | OP380812 | OP380833 | OP380853 | this study |
| E. sibirica | E. tanhoensis | MS1 | R2 | OP380070 | – | – | – | – | this study |
| E. sibirica | E. sibirica | ONA1 | R1 | OP380071 | P1 | OP380813 | OP380834 | OP380854 | this study |
| E. sibirica | E. sibirica | ONA1 | R2 | OP380072 | P2 | OP380814 | OP380835 | OP380855 | this study |
| E. sibirica | E. sibirica | SD1 | R1 | OP380073 | P1 | OP380815 | OP380836 | OP380856 | this study |
| E. sibirica | E. sibirica | SD1 | R2 | OP380074 | – | – | – | – | this study |
| E. sibirica | E. sibirica | SU1 | R1 | OP380075 | P1 | OP380816 | OP380837 | OP380857 | this study |
| E. sibirica | E. sibirica | SU1 | R2 | OP380076 | – | – | – | – | this study |
| E. sibirica | E. sibirica | TGK1 | R1 | OP380077 | P1 | OP380817 | OP380838 | OP380858 | this study |
| E. sibirica | E. sibirica | TGK1 | R2 | OP380078 | – | – | – | – | this study |
| E. sibirica | E. sibirica | UT1 | R1 | OP380079 | P1 | OP380818 | OP380839 | OP380859 | this study |
| E. sibirica | E. sibirica | UT1 | – | – | P2 | OP380819 | OP380840 | OP380860 | this study |
| E. sibirica | E. sibirica | Z2 | R1 | OP380080 | P1 | OP380820 | OP380841 | OP380861 | this study |
| E. sibirica | E. sibirica | Z2 | R2 | OP380081 | P2 | OP380821 | OP380842 | OP380862 | this study |
| E. sibirica | E. sibirica | – | – | MW716491.1 | – | MW722268.1 | MW722268.1 | MW722293.1 | [4] |
| E. stellata | – | – | – | JF505766.1 | – | JF505892.1 | JF505892.1 | JF505808.1 | [1] |
| E. tanhoensis | E. tanhoensis | – | – | MW716498.1 | – | MW722274.1 | MW722274.1 | MW722300.1 | [4] |
| E. pinnatifida | – | – | – | JF505801.1 | – | JF505927.1 | JF505927.1 | JF505843.1 | [1] |
| E. pungdoensis | – | – | – | JF505793.1 | – | JF505919.1 | JF505919.1 | JF505835.1 | [1] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Protopopova, M.V.; Pavlichenko, V.V. Eranthis Salisb. (Ranunculaceae) in South Siberia: Insights into Phylogeography and Taxonomy. Diversity 2022, 14, 779. https://doi.org/10.3390/d14100779
Protopopova MV, Pavlichenko VV. Eranthis Salisb. (Ranunculaceae) in South Siberia: Insights into Phylogeography and Taxonomy. Diversity. 2022; 14(10):779. https://doi.org/10.3390/d14100779
Chicago/Turabian StyleProtopopova, Marina V., and Vasiliy V. Pavlichenko. 2022. "Eranthis Salisb. (Ranunculaceae) in South Siberia: Insights into Phylogeography and Taxonomy" Diversity 14, no. 10: 779. https://doi.org/10.3390/d14100779
APA StyleProtopopova, M. V., & Pavlichenko, V. V. (2022). Eranthis Salisb. (Ranunculaceae) in South Siberia: Insights into Phylogeography and Taxonomy. Diversity, 14(10), 779. https://doi.org/10.3390/d14100779