
Annual Cycle of the Synechococcus spp. and Picoeukaryotic Growth and Loss Rates in a Subtropical Coastal Ecosystem




Abstract
1. Introduction
2. Materials and Methods
2.1. Water Sampling
2.2. Incubation Experimental Design
3. Results
3.1. Environmental Parameters
3.2. Seasonal Variations of Picophytoplankton Abundance
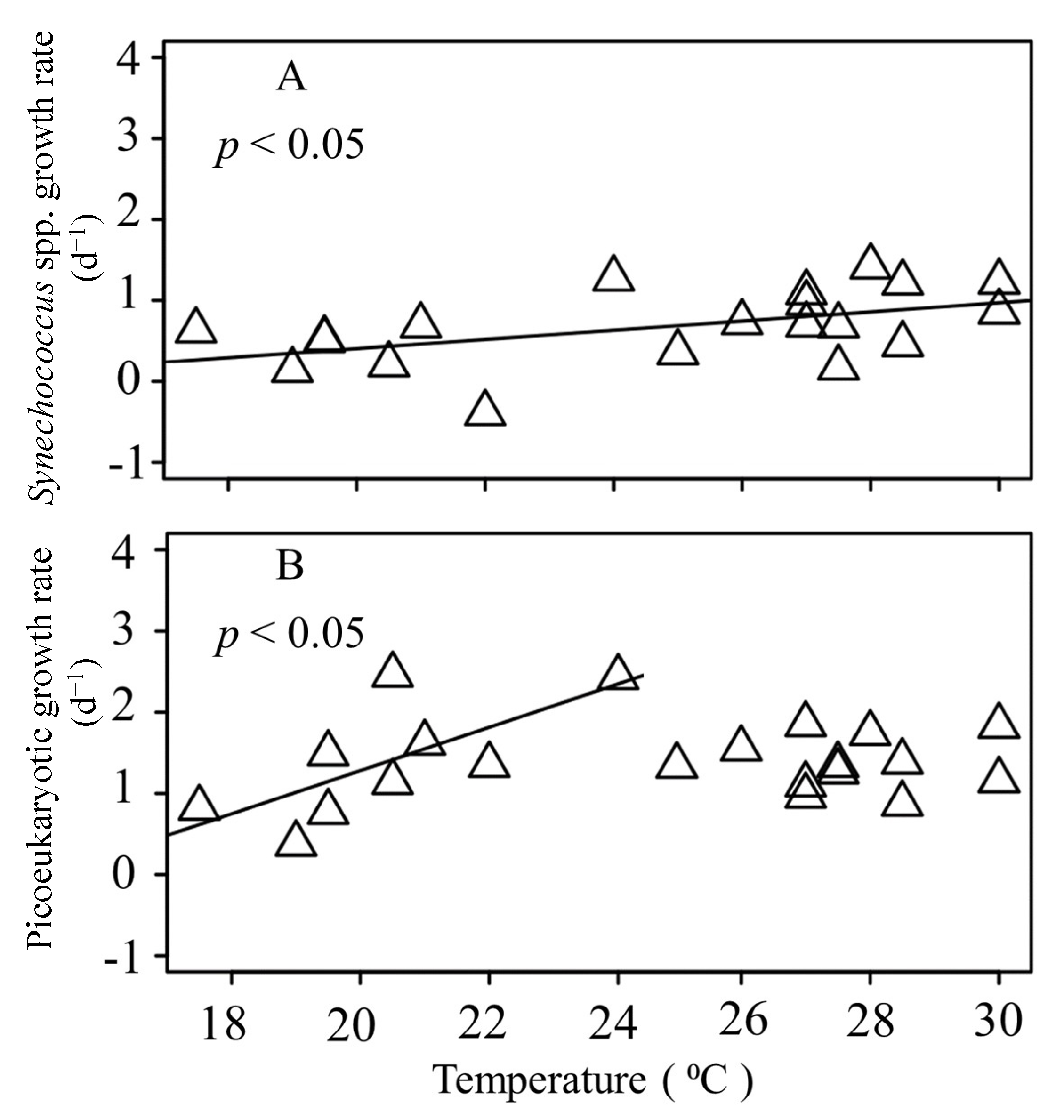
3.3. Seasonal Variations of Picophytoplankton Growth Rates
3.4. Seasonal Variations of Picophytoplankton Mortality
4. Discussion
4.1. Seasonal Dynamics of Synechococcus spp. and Picoeukaryotic Abundance
4.2. Seasonal Dynamics of Synechococcus spp. and Picoeukaryotic Growth Rates
4.3. Seasonal Dynamics of Synechococcus spp. and Picoeukaryotic Mortality Rate
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raven, J.A. The twelfth Tansley Lecture. Small is beautiful: The picophytoplankton. Funct. Ecol. 1998, 12, 503–513. [Google Scholar] [CrossRef]
- DuRand, M.D.; Olson, R.J.; Chisholm, S.W. Phytoplankton population dynamics at the Bermuda Atlantic Time-series station in the Sargasso Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 48, 1983–2003. [Google Scholar] [CrossRef]
- Mann, N.H. Phages of the marine cyanobacterial picophytoplankton. FEMS Microbiol. Rev. 2003, 27, 17–34. [Google Scholar] [CrossRef]
- Medlin, L.K.; Metfies, K.; Mehl, H.; Wiltshire, K.; Valentin, K. Picoeukaryotic plankton diversity at the Helgoland time series site as assessed by three molecular methods. Microb. Ecol. 2006, 52, 53–71. [Google Scholar] [CrossRef]
- Kirkham, A.R.; Lepère, C.; Jardillier, L.E.; Not, F.; Bouman, H.; Mead, A.; Scanlan, D.J. A global perspective on marine photosynthetic picoeukaryote community structure. ISME J. 2013, 7, 922–936. [Google Scholar] [CrossRef]
- Morán, X.A.G.; Fernández, E.; Pérez, V. Size-fractionated primary production, bacterial production and net community production in subtropical and tropical domains of the oligotrophic NE Atlantic in autumn. Mar. Ecol. Prog. Ser. 2004, 274, 17–29. [Google Scholar] [CrossRef]
- Li, W.K.W. Primary production of prochlorophytes, cyanobacteria, and eucaryotic ultraphytoplankton: Measurements from flow cytometric sorting. Limnol. Oceanogr. 1994, 39, 169–175. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Duarte, C.M.; Agustí, S. Nutrient and temperature control of the contribution of picoplankton to phytoplankton biomass and production. Limnol. Oceanogr. 2000, 45, 591–600. [Google Scholar] [CrossRef]
- Calbet, A.; Landry, M.R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnol. Oceanogr. 2004, 49, 51–57. [Google Scholar] [CrossRef]
- Chen, B.; Laws, E.A. Is there a difference of temperature sensitivity between marine phytoplankton and heterotrophs? Limnol. Oceanogr. 2017, 62, 806–817. [Google Scholar] [CrossRef]
- Evans, C.; Archer, S.D.; Jacquet, S.; Wilson, W.H. Direct estimates of the contribution of viral lysis and microzooplankton grazing to the decline of a Micromonas spp. population. Aquat. Microb. Ecol. 2003, 30, 207–219. [Google Scholar] [CrossRef]
- Tsai, A.-Y.; Chiang, K.-P.; Chang, J.; Gong, G.-C. Seasonal diel variations of picoplankton and nanoplankton in a subtropical western Pacific coastal ecosystem. Limnol. Oceanogr. 2005, 50, 1221–1231. [Google Scholar] [CrossRef]
- Worden, A.Z.; Binder, B.J. Application of dilution experiments for measuring growth and mortality rates among Prochlorococcus and Synechococcus populations in oligotrophic environments. Aquat. Microb. Ecol. 2003, 30, 159–174. [Google Scholar] [CrossRef]
- Worden, A.Z.; Nolan, J.K.; Palenik, B. Assessing the dynamics and ecology of marine picophytoplankton: The importance of the eukaryotic component. Limnol. Oceanogr. 2004, 49, 168–179. [Google Scholar] [CrossRef]
- Tsai, A.-Y.; Chiang, K.-P.; Chan, Y.-F.; Lin, Y.-C.; Chang, J. Pigmented nanoflagellates in the coastal western subtropical Pacific are important grazers on Synechococcus populations. J. Plankton Res. 2007, 29, 71–77. [Google Scholar] [CrossRef]
- Tsai, A.-Y.; Gong, G.-C.; Sanders, R.W.; Chiang, K.-P.; Huang, J.-K.; Chan, Y.-F. Viral lysis and nanoflagellate grazing as factors controlling diel variations of Synechococcus spp. summer abundance in coastal waters of Taiwan. Aquat. Microb. Ecol. 2012, 66, 159–167. [Google Scholar] [CrossRef]
- Guo, C.; Liu, H.; Zheng, L.; Song, S.; Chen, B.; Huang, B. Seasonal and spatial patterns of picophytoplankton growth, grazing and distribution in the East China Sea. Biogeosciences 2014, 11, 1847–1862. [Google Scholar] [CrossRef]
- Tsai, A.-Y.; Chiang, K.-P.; Chang, J.; Gong, G.-C. Seasonal variations in trophic dynamics of nanoflagellates and picoplankton in coastal waters of the western subtropical Pacific Ocean. Aquat. Microb. Ecol. 2008, 51, 263–274. [Google Scholar] [CrossRef]
- Otero-Ferrer, J.L.; Cermeño, P.; Bode, A.; Fernández-Castro, B.; Gasol, J.M.; Morán, X.A.G.; Marañon, E.; Moreira-Coello, V.; Varela, M.M.; Villamaña, M.; et al. Factors controlling the community structure of picoplankton in contrasting marine environments. Biogeosciences 2018, 15, 6199–6220. [Google Scholar] [CrossRef]
- Christaki, U.; Dolan, J.R.; Pelegri, S.; Rassoulzadegan, F. Consumption of picoplankton-size particles by marine ciliates: Effects of physiological state of the ciliate and particle quality. Limnol. Oceanogr. 1998, 43, 458–464. [Google Scholar] [CrossRef]
- Monger, B.C.; Landry, M.R.; Brown, S.L. Feeding selection of heterotrophic marine nanoflagellates based on the surface hydrophobicity of their picoplankton prey. Limnol. Oceanogr. 1999, 44, 1917–1927. [Google Scholar] [CrossRef]
- Gong, G.-C.; Shiah, F.-K.; Liu, K.-K.; Wen, Y.-H.; Liang, M.-H. Spatial and temporal variation of chlorophyll a, primary productivity and chemical hydrography in the southern East China Sea. Cont. Shelf Res. 2000, 20, 411–436. [Google Scholar] [CrossRef]
- Anderson, S.R.; Harvey, E.L. Seasonal variability and drivers of microzooplankton grazing and phytoplankton growth in a subtropical estuary. Front. Mar. Sci. 2019, 6, 174. [Google Scholar] [CrossRef]
- Landry, M.R.; Hassett, R.P. Estimating the grazing impact of marine micro-zooplankton. Mar. Biol. 1982, 67, 283–288. [Google Scholar] [CrossRef]
- Rii, Y.M.; Karl, D.M.; Church, M.J. Temporal and vertical variability in picophytoplankton primary productivity in the North Pacific Subtropical Gyre. Mar. Ecol. Prog. Ser. 2016, 562, 1–18. [Google Scholar] [CrossRef]
- Li, W.K.W. Annual average abundance of heterotrophic bacteria and Synechococcus in surface ocean waters. Limnol. Oceanogr. 1998, 43, 1746–1753. [Google Scholar] [CrossRef]
- Kuosa, H. Picoplanktonic algae in the northern Baltic Sea: Seasonal dynamics and flagellate grazing. Mar. Ecol. Prog. Ser. 1991, 73, 269–276. [Google Scholar] [CrossRef]
- Alonso-Laita, P.; Navarro, N.; Duarte, C.M.; Agusti, S. Seasonality of pico-phytoplankton abundance and cell death in a Mediterranean Bay (Bay of Palma, Majorca Island). Vie Milieu 2005, 55, 177–184. [Google Scholar]
- Glibert, P.M.; Wilkerson, F.P.; Dugdale, R.C.; Raven, J.A.; Dupont, C.L.; Leavitt, P.R.; Parker, A.E.; Burkholder, J.M.; Kana, T.M. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr. 2016, 61, 165–197. [Google Scholar] [CrossRef]
- Moore, L.R.; Post, A.F.; Rocap, G.; Chisholm, S.W. Utilization of different nitrogen sources by the marine cyanobacteria Prochlorococcus and Synechococcus. Limnol. Oceanogr. 2002, 47, 989–996. [Google Scholar] [CrossRef]
- Carlsson, P.; Caron, D.A. Seasonal variation of phosphorus limitation of bacterial growth in a small lake. Limnol. Oceanogr. 2001, 46, 108–120. [Google Scholar] [CrossRef]
- Ayukai, T. Possible limitation of the dilution technique for estimating growth and grazing mortality rates of picoplanktonic cyanobacteria in oligotrophic tropical waters. J. Exp. Mar. Bio. Ecol. 1996, 198, 101–111. [Google Scholar] [CrossRef]
- Tsai, A.Y.; Gong, G.C.; Huang, Y.W. Importance of the viral shunt in nitrogen cycling in Synechococcus spp. growth in subtropical Western Pacific coastal waters. Terr. Atmos. Ocean. Sci. 2014, 25, 839–846. [Google Scholar] [CrossRef]
- Bec, B.; Husseini-Ratrema, J.; Collos, Y.; Souchu, P.; Vaquer, A. Phytoplankton seasonal dynamics in a Mediterranean coastal lagoon: Emphasis on the picoeukaryote community. J. Plankton Res. 2005, 27, 881–894. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Duarte, C.M. Growth and abundance of Synechococcus sp. in a Mediterranean Bay: Seasonality and relationship with temperature. Mar. Ecol. Prog. Ser. 1998, 170, 45–53. [Google Scholar] [CrossRef]
- Behrenfeld, M.; Hardy, J.; Gucinski, H.; Hanneman, A.; Lee, H.; Wones, A. Effects of ultraviolet-B radiation on primary production along latitudinal transects in the South Pacific ocean. Mar. Environ. Res. 1993, 35, 349–363. [Google Scholar] [CrossRef]
- Helbling, E.W.; Buma, A.G.J.; Boer, M.K. de In situ impact of solar ultraviolet radiation on photosynthesis and DNA in temperate marine phytoplankton. Mar. Ecol. Prog. Ser. 2001, 211, 43–49. [Google Scholar] [CrossRef]
- Llabrés, M.; Agustí, S. Picophytoplankton cell death induced by UV radiation: Evidence for oceanic Atlantic communities. Limnol. Oceanogr. 2006, 51, 21–29. [Google Scholar] [CrossRef]
- Zhou, L.; Tan, Y.; Huang, L.; Huang, J.; Liu, H.; Lian, X. Phytoplankton growth and microzooplankton grazing in the continental shelf area of northeastern South China Sea after Typhoon Fengshen. Cont. Shelf Res. 2011, 31, 1663–1671. [Google Scholar] [CrossRef]
- Poulin, F.J.; Franks, P.J. Size-structured planktonic ecosystems: Constraints, controls and assembly instructions. J. Plankton Res. 2010, 32, 1121–1130. [Google Scholar] [CrossRef]
- Dolan, J.R.; Gallegos, C.L. Dilution effects on microzooplankton in dilution grazing experiments. Mar. Ecol. Prog. Ser. 2000, 200, 127–139. [Google Scholar] [CrossRef]
- Kamiyama, T.; Arima, S. Feeding characteristics of two tintinnid ciliate species on phytoplankton including harmful species: Effects of prey size on ingestion rates and selectivity. J. Exp. Mar. Bio. Ecol. 2001, 257, 281–296. [Google Scholar] [CrossRef]
- Calbet, A.; Trepat, I.; Almeda, R.; Saló, V.; Saiz, E.; Movilla, J.I.; Alcaraz, M.; Yebra, L.; Simó, R. Impact of micro- And nanograzers on phytoplankton assessed by standard and size-fractionated dilution grazing experiments. Aquat. Microb. Ecol. 2008, 50, 145–156. [Google Scholar] [CrossRef]
- Arenovski, A.L.; Lim, E.L.; Caron, D.A. Mixotrophic nanoplankton in oligotrophic surface waters of the Sargasso Sea may employ phagotrophy to obtain major nutrients. J. Plankton Res. 1995, 17, 801–820. [Google Scholar] [CrossRef]
- Christaki, U.; Van Wambeke, F.; Dolan, J.R. Nanoflagellates (mixotrophs, heterotrophs and autotrophs) in the oligotrophic eastern Mediterranean: Standing stocks, bacterivory and relationships with bacterial production. Mar. Ecol. Prog. Ser. 1999, 181, 297–307. [Google Scholar] [CrossRef][Green Version]
- Suttle, C.A.; Chan, A.M. Dynamics and distribution of cyanophages and their effect on marine Synechococcus spp. Appl. Environ. Microbiol. 1994, 60, 3167–3174. [Google Scholar] [CrossRef] [PubMed]
- Baudoux, A.; Veldhuis, M.; Noordeloos, A.; van Noort, G.; Brussaard, C.P.D. Estimates of virus- vs. grazing induced mortality of picophytoplankton in the North Sea during summer. Aquat. Microb. Ecol. 2008, 52, 69–82. [Google Scholar] [CrossRef]
- Baudoux, A.-C.; Veldhuis, M.J.W.; Witte, H.J.; Brussaard, C.P.D. Viruses as mortality agents of picophytoplankton in the deep chlorophyll maximum layer during IRONAGES III. Limnol. Oceanogr. 2007, 52, 2519–2529. [Google Scholar] [CrossRef]
- Long, A.; McDaniel, L.D.; Mobberley, J.; Paul, J.H. Comparison of lysogeny (prophage induction) in heterotrophic bacterial and Synechococcus populations in the Gulf of Mexico and Mississippi river plume. ISME J. 2008, 2, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Kimmance, S.A.; Wilson, W.H.; Archer, S.D. Modified dilution technique to estimate viral versus grazing mortality of phytoplankton: Limitations associated with method sensitivity in natural waters. Aquat. Microb. Ecol. 2007, 49, 207–222. [Google Scholar] [CrossRef]
- Tsai, A.-Y.; Gong, G.-C.; Chung, C.-C.; Huang, Y.-T. Different impact of nanoflagellate grazing and viral lysis on Synechococcus spp. and picoeukaryotic mortality in coastal waters. Estuar. Coast. Shelf Sci. 2018, 209, 1–6. [Google Scholar] [CrossRef]
- Jacquet, S.; Heldal, M.; Iglesias-Rodriguez, D.; Larsen, A.; Wilson, W.; Bratbak, G. Flow cytometric analysis of an Emiliana huxleyi bloom terminated by viral infection. Aquat. Microb. Ecol. 2002, 27, 111–124. [Google Scholar] [CrossRef]
- Sandaa, R.-A.; Larsen, A. Seasonal variations in virus-host populations in norwegian coastal waters: Focusing on the cyanophage community infecting marine Synechococcus spp. Appl. Environ. Microbiol. 2006, 72, 4610–4618. [Google Scholar] [CrossRef]
- Jacquet, S.; Domaizon, I.; Personnic, S.; Ram, A.S.P.; Hedal, M.; Duhamel, S.; Sime-Ngando, T. Estimates of protozoan- and viral-mediated mortality of bacterioplankton in Lake Bourget (France). Freshw. Biol. 2005, 50, 627–645. [Google Scholar] [CrossRef]
- Šimek, K.; Pernthaler, J.; Weinbauer, M.G.; Hornák, K.; Dolan, J.R.; Nedoma, J.; Mašín, M.; Amann, R. Changes in bacterial community composition and dynamics and viral mortality rates associated with enhanced flagellate grazing in a mesoeutrophic reservoir. Appl. Environ. Microbiol. 2001, 67, 2723–2733. [Google Scholar] [CrossRef] [PubMed]
- Weinbauer, M.G.; Christaki, U.; Nedoma, J. Comparing the effects of resource enrichment and grazing on viral production in a meso-eutrophic reservoir. Aquat. Microb. Ecol. 2003, 31, 137–144. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Synechococcus spp. | |||
|---|---|---|---|
| Date | k (200 μm Filtered) (d−1) | k (0.2 μm Diluted) (d−1) | k (30 kDa Diluted) (d−1) |
| 4 Oct | 0.69 ± 0.07 | −0.5 ± 0.12 | −4.2 ± 0.31 |
| 9 Oct | 0.73 ± 0.14 | 0.43 ± 0.17 | −1.92 ± 0.48 |
| 22 Oct | −2.21 ± 0.24 | −1.3 ± 0.14 | −2.01 ± 0.51 |
| 4 Nov | −0.59 ± 0.27 | 0.34 ± 0.21 | −0.13 ± 0.42 |
| 3 Dec | 0.14 ± 0.31 | −0.39 ± 0.33 | −0.91 ± 0.17 |
| 17 Dec | ND | ND | ND |
| 5 Jan | −0.77 ± 0.31 | −0.12 ± 0.11 | −0.52 ± 0.42 |
| 11 Feb | −0.39 ± 0.24 | 0.02 ± 0.04 | 0.12 ± 0.05 |
| 9 Mar | −0.01 ± 0.03 | 0.05 ± 0.02 | 0.1 ± 0.03 |
| 26 Mar | 0.69 ± 0.14 | 0.71 ± 0.22 | 0.71 ± 0.31 |
| 16 Apr | 0.37 ± 0.03 | 0.44 ± 0.02 | 0.45 ± 0.07 |
| 6 May | 0.28 ± 0.02 | 0.32 ± 0.01 | 0.32 ± 0.08 |
| 17 May | 0.66 ± 0.07 | 0.66 ± 0.01 | 0.68 ± 0.01 |
| 2 Jun | 0.75 ± 0.04 | 0.86 ± 0.02 | 0.86 ± 0.07 |
| 15 Jun | 0.17 ± 0.14 | 0.18 ± 0.07 | 0.19 ± 0.09 |
| 17 Jul | 1.04 ± 0.04 | 1.13 ± 0.03 | 1.14 ± 0.08 |
| 27 Jul | 0.7 ± 0.02 | 0.74 ± 0.01 | 0.78 ± 0.02 |
| 15 Aug | 1.17 ± 0.08 | 1.19 ± 0.04 | 1.22 ± 0.01 |
| 27 Aug | 0.46 ± 0.11 | 0.47 ± 0.09 | 0.47 ± 0.12 |
| 2 Sep | 1.1 ± 0.24 | 1.11 ± 0.14 | 1.12 ± 0.48 |
| 8 Sep | 1.42 ± 0.42 | 1.43 ± 0.17 | 1.43 ± 0.28 |
| Picoeukaryotes | |||
|---|---|---|---|
| Date | k (200 μm Filtered) (d−1) | k (0.2 μm Diluted) (d−1) | k (30 kDa Diluted) (d−1) |
| 4 Oct | 0.77 ± 0.52 | 0.62 ± 0.12 | 0.91 ± 0.14 |
| 9 Oct | 1.55 ± 0.71 | 1.54 ± 0.42 | −1.14 ± 0.78 |
| 22 Oct | −0.25 ± 0.04 | −1.1 ± 0.15 | −0.3 ± 0.11 |
| 4 Nov | −0.06 ± 0.07 | 1.19 ± 0.15 | 0.76 ± 0.32 |
| 3 Dec | 0.38 ± 0.14 | 0 ± 0.09 | 0 ± 0.48 |
| 17 Dec | 1.14 ± 0.42 | 1.39 ± 0.51 | 1.12 ± 0.34 |
| 5 Jan | −0.63 ± 0.62 | 0.43 ± 0.11 | −0.12 ± 0.09 |
| 11 Feb | 0.82 ± 0.47 | 0.82 ± 0.27 | 0.42 ± 0.14 |
| 9 Mar | 1.58 ± 0.03 | 1.9 ± 0.02 | 2.02 ± 0.01 |
| 26 Mar | 1.16 ± 0.04 | 1.26 ± 0.02 | 1.39 ± 0.03 |
| 16 Apr | 0.43 ± 0.04 | 0.57 ± 0.02 | 0.6 ± 0.01 |
| 6 May | 0.84 ± 0.03 | 0.96 ± 0.03 | 1.09 ± 0.04 |
| 17 May | 0.93 ± 0.12 | 0.94 ± 0.01 | 0.96 ± 0.01 |
| 2 Jun | 1.73 ± 0.01 | 1.76 ± 0.01 | 1.79 ± 0.02 |
| 15 Jun | 1.17 ± 0.11 | 1.18 ± 0.02 | 1.23 ± 0.01 |
| 17 Jul | 1.31 ± 0.01 | 1.33 ± 0.01 | 1.35 ± 0.01 |
| 27 Jul | 0.8 ± 0.03 | 0.88 ± 0.02 | 0.98 ± 0.04 |
| 15 Aug | 1.73 ± 0.02 | 1.78 ± 0.02 | 1.79 ± 0.07 |
| 27 Aug | 0.87 ± 0.09 | 0.88 ± 0.04 | 0.9 ± 0.03 |
| 2 Sep | 1.05 ± 0.08 | 1.07 ± 0.02 | 1.1 ± 0.01 |
| 8 Sep | 1.6 ± 0.02 | 1.67 ± 0.03 | 1.69 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, P.-C.; Gong, G.-C.; Mukhanov, V.; Zhu, Z.-Y.; Tsai, A.-Y. Annual Cycle of the Synechococcus spp. and Picoeukaryotic Growth and Loss Rates in a Subtropical Coastal Ecosystem. Diversity 2022, 14, 49. https://doi.org/10.3390/d14010049
Ho P-C, Gong G-C, Mukhanov V, Zhu Z-Y, Tsai A-Y. Annual Cycle of the Synechococcus spp. and Picoeukaryotic Growth and Loss Rates in a Subtropical Coastal Ecosystem. Diversity. 2022; 14(1):49. https://doi.org/10.3390/d14010049
Chicago/Turabian StyleHo, Pei-Chi, Gwo-Ching Gong, Vladimir Mukhanov, Zhi-Yu Zhu, and An-Yi Tsai. 2022. "Annual Cycle of the Synechococcus spp. and Picoeukaryotic Growth and Loss Rates in a Subtropical Coastal Ecosystem" Diversity 14, no. 1: 49. https://doi.org/10.3390/d14010049
APA StyleHo, P.-C., Gong, G.-C., Mukhanov, V., Zhu, Z.-Y., & Tsai, A.-Y. (2022). Annual Cycle of the Synechococcus spp. and Picoeukaryotic Growth and Loss Rates in a Subtropical Coastal Ecosystem. Diversity, 14(1), 49. https://doi.org/10.3390/d14010049