
Identification and Characterization of the Detoxification Genes Based on the Transcriptome of Tomicus yunnanensis

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect and Tissue Collection
2.2. Total RNA Extraction and cDNA Synthesis
2.3. Library Construction, Sequencing, and Functional Annotation
2.4. Gene Identification and Sequence Analysis
2.5. Quantitative Real-Time PCR
3. Results
3.1. Transcriptome Assembly
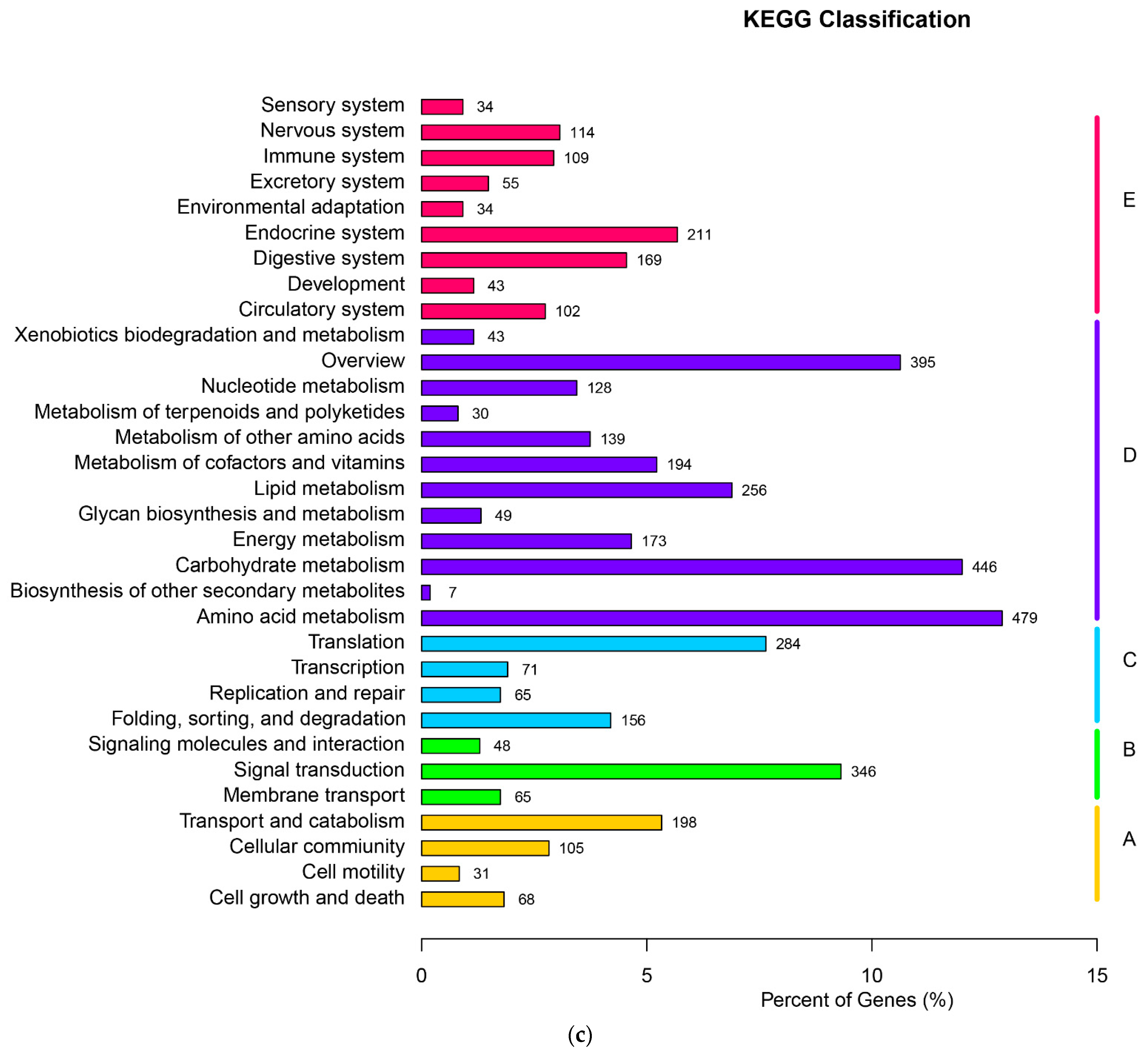
3.2. Functional Annotation of the Unigenes in T. yunnanensis
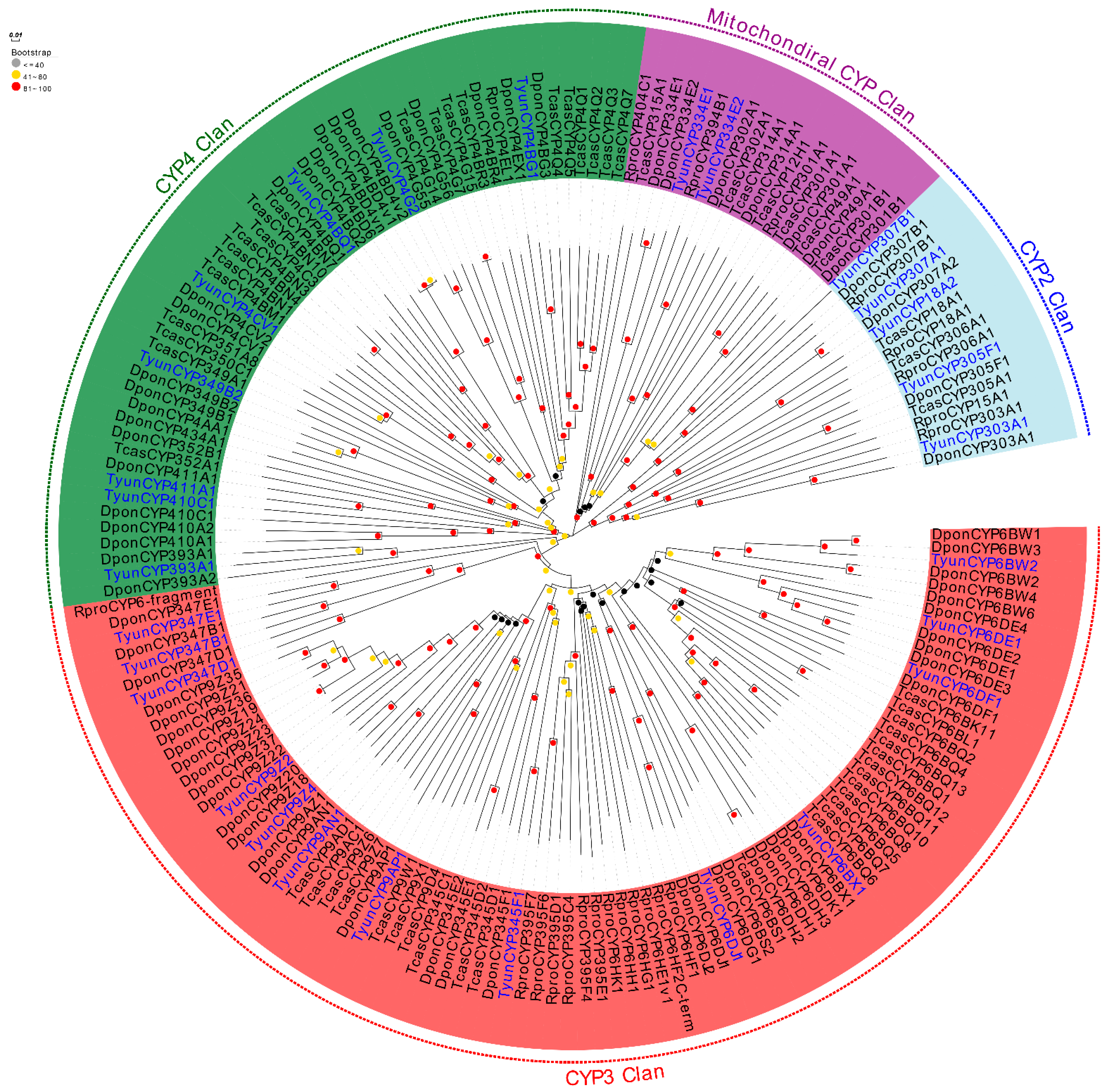
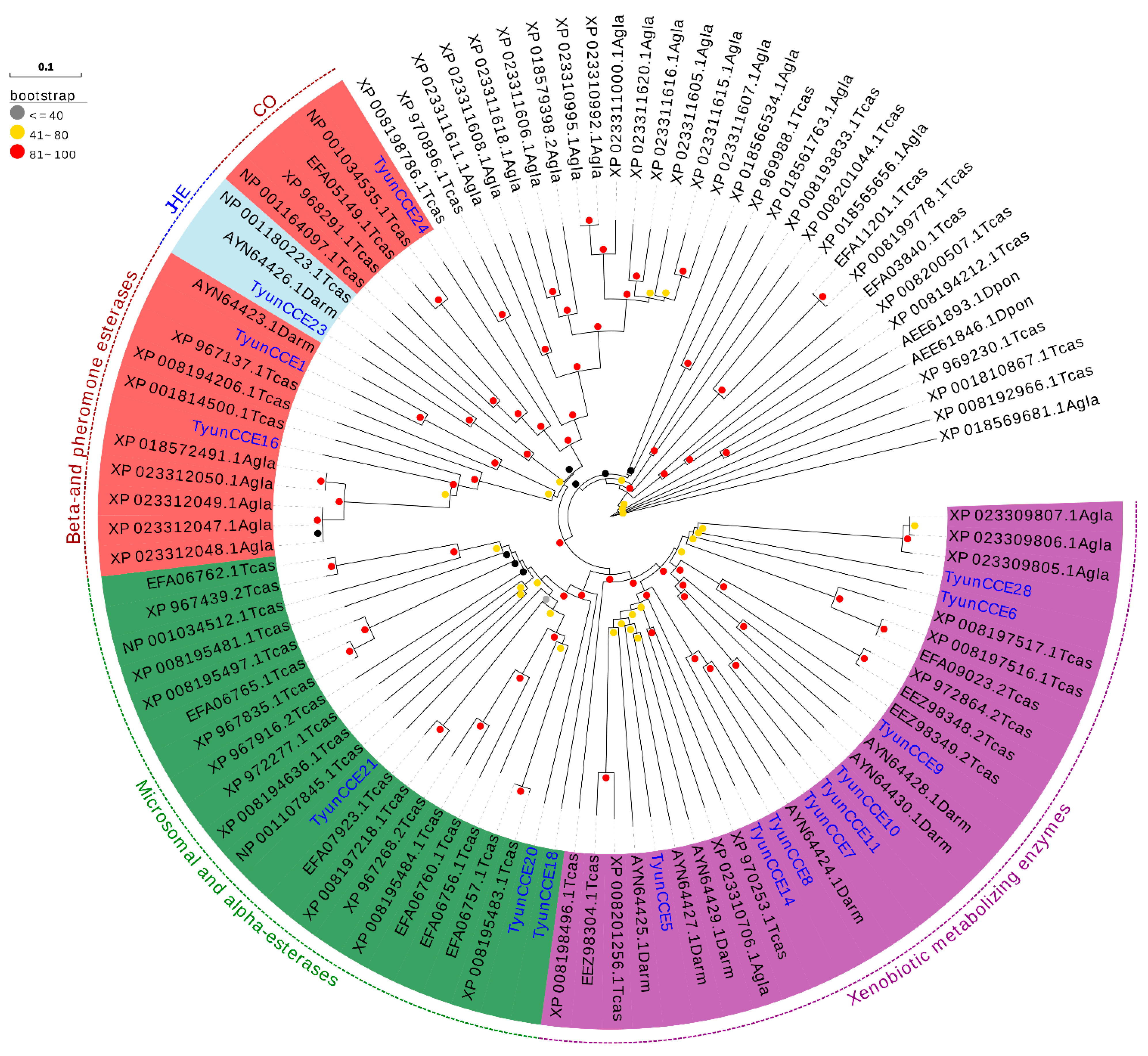
3.3. Identification of Candidate CYPs, GSTs, and CCEs
3.4. Tissue Expression Profile of the CYP, GST, and CCE Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.; Villarroel, C.A.; Ataíde, L.; Dermauw, W.; Glas, J.J.; et al. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef]
- Richard, K.; Anurag, A.A.; Marc, M. The benefits of induced defenses against herbivores. Ecology 1997, 78, 1351–1355. [Google Scholar] [CrossRef]
- Walling, L.L. The Myriad Plant Responses to Herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Hermsmeier, D.; Schittko, U.; Baldwin, I.T. Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera.; Sphingidae) and its natural host nicotiana attenuata. I. Large-Scale Changes in the accumulation of growth-and defense-related plant mRNAs. Plant Physiol. 2001, 125, 683–700. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Kang, Z.J.; Shi, X.Y.; Gao, X.W. Metabolic adaptation mechanisms of insects to plant secondary metabolites and their implications for insecticide resistance of insects. Acta Entomol. Sin. 2015, 58, 1126–1139. [Google Scholar]
- Ingelman-Sundberg, M. Genetic variability in susceptibility and response to toxicants. Toxicol. Lett. 2001, 120, 259–268. [Google Scholar] [CrossRef]
- Jiang, G.; Zhang, Y.; Chen, F.; Li, J.; Li, X.; Yue, J.; Liu, H.; Li, H.; Ran, C. Differential Analysis of the Cytochrome p450 Acaricide-Resistance Genes in Panonychus citri (Trombidiformes: Tetranychidae) Strains. Fla. Entomol. 2015, 98, 318–329. [Google Scholar] [CrossRef]
- Yi, X.Q.; Di, Y.D.; Zhang, Q. Preliminary Study on Genetic Diversity of Esox reicherti in Amur River Basin Based on Cytochrome b Gene. J. Anhui Agric. Sci. 2009, 37, 5403–5405. [Google Scholar] [CrossRef]
- Itokawa, K.; Komagata, O.; Kasai, S.; Kawada, H.; Mwatele, C.; Dida, G.O.; Njenga, S.; Mwandawiro, C.; Tomita, T. Global spread and genetic variants of the two CYP9M10 haplotype forms associated with insecticide resistance in Culex quinquefasciatus Say. Heredity 2013, 111, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Schuler, M.A.; Berenbaum, M.R. Structure and Function of Cytochrome P450S in Insect Adaptation to Natural and Synthetic Toxins: Insights Gained from Molecular Modeling. J. Chem. Ecol. 2013, 39, 1232–1245. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect CYP Genes and P450 Enzymes☆. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Mao, Y.B.; Cai, W.J.; Wang, J.W. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Liu, S.; Shi, X.-X.; Jiang, Y.-D.; Zhu, Z.-J.; Qian, P.; Zhang, M.-J.; Yu, H.; Zhu, Q.-Z.; Gong, Z.-J.; Zhu, Z.-R. De novo analysis of the Tenebrio molitor (Coleoptera: Tenebrionidae) transcriptome and identification of putative glutathione S-transferase genes. Appl. Entomol. Zool. 2015, 50, 63–71. [Google Scholar] [CrossRef]
- Vontas, J.G.; Small, G.J.; Nikou, D.C.; Ranson, H.; Hemingway, J. Purification, molecular cloning and heterologous expression of a glutathione S-transferase involved in insecticide resistance from the rice brown planthopper, Nilaparvata lugens. Biochem. J. 2002, 362, 329–337. [Google Scholar] [CrossRef]
- Friedman, R. Genomic organization of the glutathione S-transferase family in insects. Mol. Phylogenetics Evol. 2011, 61, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.N.; Dai, L.L.; Zhang, R.R.; Chen, H. Cloning and expression of glutathione S-transferase gene DaGSTe1 of Dendroctonus armandi. J. Northwest A F Univ. 2015, 43, 117–122. [Google Scholar]
- Durand, N.; Carot-Sans, G.; Chertemps, T.; Bozzolan, F.; Party, V.; Renou, M.; Debernard, S.; Rosell, G.; Maïbèche-Coisné, M. Characterization of an antenna carboxylesterase from the pest moth Spodoptera littoralis degrading a host Plant Odorant. PLoS ONE 2010, 5, e15026. [Google Scholar] [CrossRef]
- Yu, L.F.; Huang, H.G.; Ze, S.Z.; Ren, L.L.; Zong, S.X.; Lu, W.J.; Luo, Y.Q. Research on the spatial distribution patterns of Tomicus sp. in Pinus yunnanensis during the shoot feeding period. Chin. J. Appl. Entomol. 2017, 54, 940–946. [Google Scholar]
- Chen, P.; Lu, J.; Haack, R.A.; Ye, H. Attack pattern and reproductive ecology of Tomicus brevipilosus (Coleoptera: Curculionidae) on Pinus yunnanensis in Southwestern China. J. Insect Sci. 2015, 15, 43. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Chen, P.; Wang, Y.X.; Yuan, R.L.; Feng, D.; Li, L.S.; Ye, H.; Pan, Y.; Lv, J.; Zhou, Y.F.; et al. Population Structure and Succession Law of Tomicus Species in Yunnan. For. Res. 2018, 31, 167–172. [Google Scholar]
- Lü, J.; Hu, S.J.; Ma, X.Y.; Chen, J.M.; Li, Q.Q.; Ye, H.; Arthofer, W. Origin and Expansion of the Yunnan Shoot Borer, Tomicus yunnanensis (Coleoptera: Scolytinae): A Mixture of Historical Natural Expansion and Contemporary Human-Mediated Relocation. PLoS ONE 2014, 9, e111940. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Z.; Kong, X.; Wang, H.; Zhang, S. Intraspecific and interspecific attraction of three Tomicus beetle species during the shoot-feeding phase. Bull. Entomol. Res. 2015, 105, 225–233. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Cock, P.J.A.; Christopher, J.F.; Naohisa, G.; Michael, L.H.; Peter, M.R. The Sanger FASTQ file format for sequences with quality scores, and the Solexa/Illumina FASTQ variants. Nucleic Acids Res. 2010, 38, 1767–1771. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. 8—Insect CYP Genes and P450 Enzymes. In Insect Molecular Biology and Biochemistry; Academic Press: San Diego, CA, USA, 2012; pp. 236–316. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Balakrishnan, S.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Singh, V.K.; Mangalam, A.K.; Dwivedi, S.; Naik, S. Primer Premier: Program for Design of Degenerate Primers from a Protein Sequence. BioTechniques 1998, 24, 318–319. [Google Scholar] [CrossRef]
- Simon, P. Q-Gene: Processing quantitative real-time RT–PCR data. Bioinformatics 2003, 19, 1439–1440. [Google Scholar] [CrossRef]
- Shi, H.; Pei, L.; Gu, S.; Zhu, S.; Wang, Y.; Zhang, Y.; Li, B. Glutathione S-transferase (GST) genes in the red flour beetle, Tribolium castaneum, and comparative analysis with five additional insects. Genomics 2012, 100, 327–335. [Google Scholar] [CrossRef]
- Schama, R.; Pedrini, N.; Juárez, M.P.; Nelson, D.R.; Torres, A.Q.; Valle, D.; Mesquita, R.D. Rhodnius prolixus supergene families of enzymes potentially associated with insecticide resistance. Insect. Biochem. Mol. Biol. 2016, 69, 91–104. [Google Scholar] [CrossRef]
- Feyereisen, R. Evolution of insect P450. Biochem. Soc. Trans. 2006, 34, 1252–1255. [Google Scholar] [CrossRef]
- Liu, N.Y.; Li, Z.B.; Zhao, N.; Song, Q.S.; Zhu, J.Y.; Yang, B. Identification and characterization of chemosensory gene families in the bark beetle, Tomicus yunnanensis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 25, 73–85. [Google Scholar] [CrossRef]
- Liu, N.N.; Li, M.; Gong, Y.H.; Liu, F.; Li, T. Cytochrome P450s-Their expression, regulation, and role in insecticide resistance. Pestic. Biochem. Physiol. 2015, 120, 77–81. [Google Scholar] [CrossRef]
- Bozzolan, F.; Siaussat, D.; Maria, A.; Durand, N.; Pottier, M.-A.; Chertemps, T.; Maïbèche-Coisne, M. Antennal uridine diphosphate (UDP)-glycosyltransferases in a pest insect: Diversity and putative function in odorant and xenobiotics clearance. Insect. Mol. Biol. 2014, 23, 539–549. [Google Scholar] [CrossRef]
- Claudia, C.R.; María, F.L.; Ana, K.C.A.; Verónica, P.M.; Brian, T.S.; Gerardo, Z. Isolation and expression of cytochrome P450 genes in the antennae and gut of pine beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae) following exposure to host monoterpenes. Gene 2013, 520, 47–63. [Google Scholar] [CrossRef]
- Pottier, M.A.; Bozzolan, F.; Chertemps, T.; Jacquin-Joly, E.; Lalouette, L.; Siaussat, D.; Maïbèche-Coisne, M. Cytochrome P450s and cytochrome P450 reductase in the olfactory organ of the cotton leafworm Spodoptera littoralis. Insect Mol. Biol. 2012, 21, 568–580. [Google Scholar] [CrossRef]
- He, P.; Zhang, Y.-F.; Hong, D.-Y.; Wang, J.; Wang, X.-L.; Zuo, L.-H.; Tang, X.-F.; Xu, W.-M.; He, M. A reference gene set for sex pheromone biosynthesis and degradation genes from the diamondback moth, Plutella xylostella, based on genome and transcriptome digital gene expression analyses. BMC Genom. 2017, 18, 219. [Google Scholar] [CrossRef]
- You, Y.; Xie, M.; Ren, N.; Cheng, X.; Li, J.; Ma, X.; Zou, M.; Vasseur, L.; Gurr, G.M.; You, M. Characterization and expression profiling of glutathione S-transferasesin the diamondback moth, Plutella xylostella (L.). BMC Genom. 2015, 16, 152. [Google Scholar] [CrossRef]
- Zou, X.P.; Xu, Z.B.; Zou, H.W.; Liu, J.S.; Chen, S.N.; Feng, Q.L.; Zheng, S.C. Glutathione S-transferase SlGSTE1 in Spodoptera litura may be associated with feeding adaptation of host plants. Insect. Biochem. Mol. Biol. 2016, 70, 32–43. [Google Scholar] [CrossRef]
- Rogers, M.E.; Jani, M.K.; Vogt, R.G. An olfactory-specific glutathione-S-transferase in the sphinx moth Manduca sexta. J. Exp. Biol. 1999, 202, 1625–1637. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.L.; Ma, J.N.; Ma, M.Y.; Zhang, H.Q.; Shi, Q.; Zhang, R.; Chen, H. Characterisation of GST genes from the Chinese white pine beetle Dendroctonus armandi (Curculionidae: Scolytinae) and their response to host chemical defence. Pest Manag. Sci. 2016, 72, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Oakeshott, J.G.; Claudianos, C.; Campbell, P.M.; Newcomb, R.D.; Russell, R.J. Biochemical Genetics and Genomics of Insect Esterases. Sci. Ref. Modul. Life Sci. 2019, 5, 309–381. [Google Scholar] [CrossRef]
- Tamaki, Y.; Kawasaki, K.; Yamada, H.; Koshihara, T.; Osaki, N.; Ando, T.; Yoshida, S.; Kakinohana, H. Z-11-hexadecenal and Z-11-hexadecenyl acteact: Sex pheromone components of the Diamonback moth (Lepideptera: Plutellidae). Appl. Entomol. Zool. 1997, 12, 208–210. [Google Scholar] [CrossRef]
- Dai, L.L.; Gao, H.M.; Ye, J.Q.; Fu, D.Y.; Sun, Y.Y.; Chen, H. Isolation of CarE genes from the Chinese white pine beetle Dendroctonus armandi (Curculionidae: Scolytinae) and their response to host chemical defense. Pest Manag. Sci. 2019, 75, 986–997. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.J.; Wang, Z.Q.; Zhu, J.Y.; Liu, N.Y. Identification and characterization of detoxification genes in two cerambycid beetles, Rhaphuma horsfieldi and Xylotrechus quadripes (Coleoptera: Cerambycidae: Clytini). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2020, 243–244, 110431. [Google Scholar] [CrossRef]
- Durand, N.; Carot-Sans, G.; Chertemps, T.; Montagné, N.; Jacquin-Joly, E.; Debernard, S.; Maïbèche-Coisne, M. A diversity of putative carboxylesterases are expressed in the antennae of the noctuid moth Spodoptera littoralis. Insect Mol. Biol. 2010, 19, 87–97. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Zhang, Y.N.; Yang, K.; Li, Z.Q.; Dong, S.L. An antenna-biased carboxylesterase is specifically active to plant volatiles in Spodoptera exigua. Pestic. Biochem. Physiol. 2015, 123, 93–100. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Q20 (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|
| MA_1 | 615,114,60 | 582,887,10 | 97.95 | 94.15 | 47.57 |
| MA_2 | 474,005,30 | 462,665,14 | 97.90 | 93.91 | 42.76 |
| MA_3 | 529,246,96 | 516,082,96 | 98.06 | 94.27 | 42.81 |
| MH_1 | 581,529,36 | 558,021,48 | 97.75 | 93.46 | 38.56 |
| MH_2 | 506,735,06 | 493,021,74 | 97.96 | 94.00 | 38.74 |
| MH_3 | 610,987,72 | 590,914,14 | 98.04 | 94.16 | 40.09 |
| ML_1 | 582,201,60 | 559,355,36 | 98.04 | 94.29 | 45.54 |
| ML_2 | 561,873,52 | 535,354,48 | 98.05 | 94.30 | 43.28 |
| ML_3 | 587,793,32 | 564,878,02 | 97.88 | 93.93 | 42.65 |
| MC_1 | 640,881,58 | 628,616,66 | 97.63 | 93.21 | 38.55 |
| MC_2 | 651,352,06 | 633,407,70 | 97.87 | 93.79 | 39.15 |
| MC_3 | 591,512,94 | 566,883,04 | 97.10 | 92.16 | 41.95 |
| FA_1 | 523,288,02 | 510,649,40 | 98.11 | 94.44 | 46.17 |
| FA_2 | 568,975,58 | 533,953,70 | 98.00 | 94.23 | 46.38 |
| FA_3 | 535,300,88 | 500,120,04 | 98.00 | 94.24 | 46.14 |
| FH_1 | 513,240,72 | 502,592,66 | 97.95 | 93.94 | 39.67 |
| FH_2 | 558,425,54 | 547,795,96 | 97.81 | 93.65 | 39.48 |
| FH_3 | 640,915,78 | 629,613,86 | 97.62 | 93.18 | 39.24 |
| FL_1 | 586,049,54 | 572,336,12 | 97.83 | 93.72 | 43.91 |
| FL_2 | 503,729,98 | 491,791,08 | 97.82 | 93.72 | 42.40 |
| FL_3 | 435,955,04 | 420,544,90 | 97.94 | 94.19 | 52.60 |
| FC_1 | 572,708,34 | 560,824,28 | 97.67 | 93.30 | 40.48 |
| FC_2 | 533,416,24 | 517,611,84 | 97.87 | 93.78 | 41.24 |
| FC_3 | 505,727,10 | 495,473,28 | 97.77 | 93.55 | 39.88 |
| Min Length | Mean Length | Max Length | N50 | Total Nucleotides | |
|---|---|---|---|---|---|
| Transcripts | 154 | 1005 | 59,000 | 1465 | 184,171,785 |
| Genes | 301 | 877 | 59,000 | 1153 | 880,711,34 |
| Number of Genes | Percentage (%) | |
|---|---|---|
| Annotated in NR | 60,067 | 59.79 |
| Annotated in NT | 54,782 | 54.53 |
| Annotated in SwissProt | 50,102 | 49.87 |
| Annotated in PFAM | 53,205 | 52.96 |
| Annotated in GO | 53,205 | 52.96 |
| Annotated in KOG | 19,808 | 19.71 |
| Annotated in all databases | 1666 | 1.65 |
| Annotated in at least one database | 79,508 | 79.14 |
| Total unigenes | 100,455 | 100 |
| Gene Name | Gene ID | aa | E-Value | Identity | Accession | Species Name |
|---|---|---|---|---|---|---|
| TyunCYP349A1 | i2_LQ_TYUN_c39805/f1p0/2021 | 60 | 1 × 10−20 | 65.00% | AEL88544.1 | Dendroctonus rhizophagus |
| TyunCYP9Z1 | i2_LQ_TYUN_c22584/f1p0/2717 | 145 | 5 × 10−82 | 82.64% | AEL88550.1 | Dendroctonus rhizophagus |
| TyunCYP314A1 | i1_LQ_TYUN_c152368/f1p0/1050 | 147 | 6 × 10−78 | 83.33% | AFI45004.1 | Dendroctonus ponderosae |
| TyunCYP347B1 | i4_LQ_TYUN_c2131/f1p0/4666 | 447 | 0 | 81.94% | AFI45010.1 | Dendroctonus ponderosae |
| TyunCYP410C1 | Cluster-27834.11928 | 456 | 1 × 10−107 | 39.43% | AFI45013.1 | Dendroctonus ponderosae |
| TyunCYP4CV1 | Cluster-27834.3883 | 507 | 0 | 77.47% | AFI45020.1 | Dendroctonus ponderosae |
| TyunCYP6BW1 | i3_LQ_TYUN_c10935/f1p0/4006 | 283 | 9 × 10−180 | 85.16% | AFI45024.1 | Dendroctonus ponderosae |
| TyunCYP6BW2 | i1_HQ_TYUN_c416/f4p1/1768 | 506 | 0 | 86.76% | AFI45026.1 | Dendroctonus ponderosae |
| TyunCYP6BX1 | Cluster-27834.4019 | 478 | 0 | 71.43% | AFI45028.1 | Dendroctonus ponderosae |
| TyunCYP6DJ1 | i1_LQ_TYUN_c33388/f1p0/1763 | 507 | 0 | 70.71% | AFI45041.1 | Dendroctonus ponderosae |
| TyunCYP9Z4 | i1_LQ_TYUN_c78340/f1p0/1911 | 526 | 0 | 84.22% | AFI45045.1 | Dendroctonus ponderosae |
| TyunCYP6DJ2 | i2_LQ_TYUN_c23405/f1p0/2443 | 66 | 6 × 10−7 | 67.65% | AGF69211.1 | Dendroctonus valens |
| TyunCYP4G1 | i3_LQ_TYUN_c4999/f1p0/3248 | 242 | 5 × 10−158 | 93.80% | ALD15896.1 | Dendroctonus armandi |
| TyunCYP305F1 | Cluster-27834.3827 | 491 | 0 | 87.53% | ALD15904.1 | Dendroctonus armandi |
| TyunCYP352B1 | i2_LQ_TYUN_c27719/f1p0/2254 | 225 | 4 × 10−74 | 69.36% | ALD15909.1 | Dendroctonus armandi |
| TyunCYP4BD4V | i1_LQ_TYUN_c15852/f1p0/1569 | 166 | 8 × 10−68 | 67.31% | ALD15912.1 | Dendroctonus ponderosae |
| TyunCYP6DF1 | Cluster-27834.8537 | 505 | 0 | 72.31% | ALD15922.1 | Dendroctonus armandi |
| TyunCYP9Z2 | i2_LQ_TYUN_c15205/f1p0/2172 | 575 | 0 | 81.30% | ALD15924.1 | Dendroctonus armandi |
| TyunCYP434A1 | Cluster-27834.7270 | 371 | 7 × 10−147 | 71.53% | XP_019754109.1 | Dendroctonus ponderosae |
| TyunCYP4BQ1 | Cluster-27834.7070 | 397 | 0 | 67.84% | XP_019754731.1 | Dendroctonus ponderosae |
| TyunCYP6DE1 | i1_HQ_TYUN_c29325/f3p0/1840 | 509 | 0 | 76.23% | XP_019755300.1 | Dendroctonus ponderosae |
| TyunCYP315A1 | Cluster-27834.2194 | 292 | 8 × 10−139 | 72.26% | XP_019755328.1 | Dendroctonus ponderosae |
| TyunCYP315A2 | Cluster-27834.2193 | 208 | 2 × 10−20 | 70.15% | XP_019755336.1 | Dendroctonus ponderosae |
| TyunCYP4G2 | i1_HQ_TYUN_c24270/f27p4/1844 | 563 | 0 | 92.11% | XP_019755432.1 | Dendroctonus ponderosae |
| TyunCYP4BG1 | Cluster-27834.11078 | 501 | 0 | 78.09% | XP_019756327.1 | Dendroctonus ponderosae |
| TyunCYP410A1 | i1_LQ_TYUN_c6617/f1p1/1851 | 273 | 5 × 10−82 | 53.52% | XP_019756499.1 | Dendroctonus ponderosae |
| TyunCYP411A1 | i2_LQ_TYUN_c22094/f1p0/2786 | 490 | 0 | 76.22% | XP_019758316.1 | Dendroctonus ponderosae |
| TyunCYP411A2 | i2_LQ_TYUN_c11742/f1p1/2726 | 185 | 6 × 10−77 | 71.43% | XP_019758318.1 | Dendroctonus ponderosae |
| TyunCYP345F1 | Cluster-26110.0 | 500 | 0 | 82.20% | XP_019759785.1 | Dendroctonus ponderosae |
| TyunCYP307B1 | Cluster-27834.15214 | 399 | 1 × 10−153 | 77.06% | XP_019760073.1 | Dendroctonus ponderosae |
| TyunCYP307A1 | i3_LQ_TYUN_c19449/f1p0/3461 | 492 | 0 | 96.54% | XP_019760634.1 | Dendroctonus ponderosae |
| TyunCYP393A2 | i0_LQ_TYUN_c4806/f1p0/966 | 112 | 5 × 10−17 | 71.93% | XP_019760717.1 | Dendroctonus ponderosae |
| TyunCYP9AP1 | i1_LQ_TYUN_c81629/f1p2/1918 | 483 | 0 | 72.57% | XP_019760793.1 | Dendroctonus ponderosae |
| TyunCYP9AN1 | i2_LQ_TYUN_c15846/f1p0/2129 | 523 | 0 | 79.31% | XP_019761014.1 | Dendroctonus ponderosae |
| TyunCYP433A1 | i1_LQ_TYUN_c80202/f1p0/1820 | 274 | 2 × 10−128 | 70.45% | XP_019761671.1 | Dendroctonus ponderosae |
| TyunCYP9Z3 | i2_LQ_TYUN_c27749/f1p0/2479 | 151 | 2 × 10−44 | 73.33% | XP_019764934.1 | Dendroctonus ponderosae |
| TyunCYP345E1 | i1_HQ_TYUN_c62660/f2p0/1795 | 304 | 4 × 10−159 | 72.33% | XP_019765603.1 | Dendroctonus ponderosae |
| TyunCYP347D1 | Cluster-20450.0 | 498 | 0 | 78.93% | XP_019767652.1 | Dendroctonus ponderosae |
| TyunCYP303A1 | Cluster-27834.21448 | 484 | 0 | 76.80% | XP_019768066.1 | Dendroctonus ponderosae |
| TyunCYP18A1 | i2_LQ_TYUN_c27030/f1p0/2265 | 159 | 8 × 10−82 | 79.87% | XP_019768143.1 | Dendroctonus ponderosae |
| TyunCYP18A2 | i2_HQ_TYUN_c25448/f2p0/2291 | 525 | 0 | 90.50% | XP_019768147.1 | Dendroctonus ponderosae |
| TyunCYP393A1 | Cluster-27834.3112 | 461 | 0 | 69.78% | XP_019768810.1 | Dendroctonus ponderosae |
| TyunCYP334E1 | Cluster-27834.2023 | 583 | 0 | 85.74% | XP_019770320.1 | Dendroctonus ponderosae |
| TyunCYP347E1 | i2_LQ_TYUN_c3851/f1p0/2279 | 500 | 0 | 79.00% | XP_019771168.1 | Dendroctonus armandi |
| TyunCYP6DG3 | i3_LQ_TYUN_c17722/f1p0/3197 | 354 | 0 | 77.68% | XP_019771861.1 | Dendroctonus ponderosae |
| TyunCYP9AZ1 | i2_LQ_TYUN_c28708/f1p0/2957 | 134 | 2 × 10−70 | 79.85% | XP_019773084.1 | Dendroctonus ponderosae |
| TyunCYP334E2 | i2_LQ_TYUN_c27105/f1p0/2563 | 498 | 0 | 83.77% | XP_019773300.1 | Dendroctonus ponderosae |
| TyunCYP349B2 | Cluster-40684.0 | 420 | 0 | 82.86% | XP_019773560.1 | Dendroctonus ponderosae |
| TyunCYP434A2 | i1_HQ_TYUN_c939/f2p0/1483 | 360 | 5 × 10−124 | 50.00% | XP_019869691.1 | Aethina tumida |
| TyunCYP6BW3 | i2_LQ_TYUN_c30164/f1p0/2137 | 382 | 2 × 10−121 | 48.28% | XP_023014703.1 | Leptinotarsa decemlineata |
| TyunCYP410C1 | i0_LQ_TYUN_c9810/f1p0/524 | 150 | 8 × 10−12 | 32.69% | XP_030755100.1 | Sitophilus oryzae |
| Gene Name | Gene ID | aa | E-Value | Identity | Accession | Species Name |
|---|---|---|---|---|---|---|
| TyunGST1 | i2_LQ_TYUN_c36781/f1p0/2077 | 217 | 2 × 10−145 | 91.71% | AIC76455.1 | Dendroctonus armandi |
| TyunGST2 | Cluster-27834.9472 | 70 | 6 × 10−41 | 97.14% | AJE61306.1 | Dendroctonus armandi |
| TyunGST3 | i0_LQ_TYUN_c12133/f1p0/748 | 141 | 3 × 10−48 | 57.25% | AJE61307.1 | Dendroctonus armandi |
| TyunGST4 | i2_LQ_TYUN_c18657/f1p0/2385 | 112 | 3 × 1062 | 76.74% | AJE61308.1 | Dendroctonus armandi |
| TyunGST5 | i3_LQ_TYUN_c11135/f1p0/3582 | 79 | 5 × 10−33 | 82.35% | AJE61309.1 | Dendroctonus armandi |
| TyunGST6 | Cluster-27834.6624 | 230 | 2 × 10−146 | 83.48% | AJE61311.1 | Dendroctonus armandi |
| TyunGST7 | Cluster-27834.8595 | 244 | 8 × 10−160 | 88.52% | AJE61312.1 | Dendroctonus armandi |
| TyunGST8 | i1_LQ_TYUN_c149780/f1p1/1152 | 245 | 2 × 10−150 | 82.04% | AVR54955.1 | Sitophilus oryzae |
| TyunGST9 | i3_LQ_TYUN_c12086/f1p0/3529 | 218 | 2 × 10−142 | 88.58% | AVR54957.1 | Sitophilus oryzae |
| TyunGST10 | i2_LQ_TYUN_c14505/f1p9/2537 | 52 | 6 × 10−15 | 71.43% | AVR54966.1 | Sitophilus oryzae |
| TyunGST11 | Cluster-27834.9551 | 105 | 9 × 10−41 | 64.42% | AVT42177.1 | Lissorhoptrus oryzophilus |
| TyunGST12 | i0_LQ_TYUN_c7185/f1p0/508 | 148 | 6 × 10−48 | 62.50% | AVT42178.1 | Lissorhoptrus oryzophilus |
| TyunGST13 | i2_LQ_TYUN_c7650/f1p0/2527 | 118 | 3 × 10−64 | 87.85% | AVT42182.1 | Lissorhoptrus oryzophilus |
| TyunGST14 | Cluster-27834.21190 | 219 | 3 × 10−69 | 51.96% | AVT42185.1 | Lissorhoptrus oryzophilus |
| TyunGST15 | i2_LQ_TYUN_c8057/f1p0/2856 | 294 | 7 × 10−150 | 67.12% | AVT42197.1 | Lissorhoptrus oryzophilus |
| TyunGST16 | Cluster-27834.9541 | 219 | 1 × 10−144 | 89.50% | QFU14637.1 | Dendroctonus armandi |
| TyunGST17 | i4_LQ_TYUN_c8880/f1p0/4497 | 137 | 3 × 10−69 | 93.69% | QFU14640.1 | Dendroctonus armandi |
| TyunGST18 | Cluster-1630.0 | 205 | 1 × 10−76 | 56.78% | QFU14643.1 | Dendroctonus armandi |
| TyunGST19 | i2_LQ_TYUN_c12200/f1p0/2343 | 231 | 3 × 10−137 | 77.92% | QFU14646.1 | Dendroctonus armandi |
| TyunGST20 | i3_LQ_TYUN_c2494/f1p0/3326 | 218 | 4 × 10−146 | 90.41% | XP_019753833.1 | Dendroctonus ponderosae |
| TyunGST21 | i2_LQ_TYUN_c34577/f1p0/2723 | 78 | 7 × 10−41 | 83.54% | XP_019753835.1 | Dendroctonus ponderosae |
| TyunGST22 | i2_LQ_TYUN_c23261/f1p0/2987 | 124 | 2 × 10−73 | 93.86% | XP_019755130.1 | Dendroctonus ponderosae |
| TyunGST23 | i2_LQ_TYUN_c24320/f1p0/2660 | 150 | 2 × 10−35 | 84.72% | XP_019755704.1 | Dendroctonus ponderosae |
| TyunGST24 | i7_LQ_TYUN_c267/f1p0/7646 | 72 | 7 × 10−22 | 67.21% | XP_019755792.1 | Dendroctonus ponderosae |
| TyunGST25 | i3_LQ_TYUN_c4401/f1p0/3564 | 111 | 4 × 10−53 | 86.27% | XP_019755793.1 | Dendroctonus ponderosae |
| TyunGST26 | i3_LQ_TYUN_c26609/f1p0/3037 | 203 | 3 × 10107 | 72.00% | XP_019755830.1 | Dendroctonus ponderosae |
| TyunGST27 | i0_LQ_TYUN_c24616/f1p0/906 | 205 | 1 × 10−129 | 85.37% | XP_019760728.1 | Dendroctonus ponderosae |
| TyunGST28 | i2_LQ_TYUN_c11257/f1p0/2424 | 559 | 0 | 75.62% | XP_019764595.1 | Dendroctonus ponderosae |
| TyunGST29 | Cluster-27834.9273 | 224 | 4 × 10−124 | 73.66% | XP_019766435.1 | Dendroctonus ponderosae |
| TyunGST30 | i1_LQ_TYUN_c146891/f1p0/1112 | 237 | 3 × 10−158 | 89.83% | XP_019768505.1 | Dendroctonus ponderosae |
| TyunGST31 | i0_LQ_TYUN_c13481/f1p0/385 | 70 | 4 × 10−32 | 82.86% | XP_019770972.1 | Dendroctonus ponderosae |
| TyunGST32 | i1_LQ_TYUN_c153362/f1p1/1072 | 206 | 3 × 10−77 | 57.28% | XP_023313068.1 | Anoplophora glabripennis |
| TyunGST33 | i1_LQ_TYUN_c150961/f1p1/1003 | 206 | 2 × 10−78 | 58.54% | XP_023313069.1 | Anoplophora glabripennis |
| Gene Name | aa | E-Value | Identity | Accession | Species Name | |
|---|---|---|---|---|---|---|
| TyunCCE1 | i2_LQ_TYUN_c19726/f1p0/2240 | 580 | 0 | 79.41% | AYN64423.1 | Dendroctonus armandi |
| TyunCCE2 | i1_LQ_TYUN_c11797/f1p0/1951 | 405 | 0 | 93.03% | AYN64424.1 | Dendroctonus armandi |
| TyunCCE3 | Cluster-27834.6014 | 133 | 3 × 10−43 | 61.42% | AYN64428.1 | Dendroctonus armandi |
| TyunCCE4 | Cluster-27834.16668 | 243 | 5 × 10−156 | 88.48% | AYN64429.1 | Dendroctonus armandi |
| TyunCCE5 | Cluster-27834.3878 | 550 | 5 × 10−164 | 45.34% | XP_019754206.1 | Dendroctonus ponderosae |
| TyunCCE6 | Cluster-27834.11182 | 570 | 0 | 81.05% | XP_019754592.1 | Dendroctonus ponderosae |
| TyunCCE7 | Cluster-27834.18917 | 577 | 0 | 87.19% | XP_019755320.1 | Dendroctonus ponderosae |
| TyunCCE8 | i1_LQ_TYUN_c11402/f1p0/1940 | 563 | 0 | 58.70% | XP_019755320.1 | Dendroctonus ponderosae |
| TyunCCE9 | Cluster-27834.6015 | 557 | 0 | 64.16% | XP_019755963.1 | Dendroctonus ponderosae |
| TyunCCE10 | i1_HQ_TYUN_c4678/f4p0/1715 | 558 | 0 | 82.08% | XP_019756055.1 | Dendroctonus ponderosae |
| TyunCCE11 | i1_LQ_TYUN_c2026/f1p0/1764 | 559 | 0 | 85.15% | XP_019756056.1 | Dendroctonus ponderosae |
| TyunCCE12 | Cluster-27834.3444 | 122 | 5 × 10−75 | 90.98% | XP_019756397.1 | Dendroctonus ponderosae |
| TyunCCE13 | Cluster-27834.5714 | 268 | 2 × 10−169 | 88.01% | XP_019756739.1 | Dendroctonus ponderosae |
| TyunCCE14 | i1_LQ_TYUN_c29743/f1p0/1703 | 545 | 0 | 50.54% | XP_019757154.1 | Dendroctonus ponderosae |
| TyunCCE15 | i1_LQ_TYUN_c35851/f1p0/1947 | 228 | 8 × 10−114 | 75.60% | XP_019758307.1 | Dendroctonus ponderosae |
| TyunCCE16 | i1_LQ_TYUN_c30292/f1p1/1995 | 566 | 0 | 88.18% | XP_019761960.1 | Dendroctonus ponderosae |
| TyunCCE17 | i1_LQ_TYUN_c16028/f1p0/1766 | 347 | 0 | 81.23% | XP_019764466.1 | Dendroctonus ponderosae |
| TyunCCE18 | i1_LQ_TYUN_c9615/f1p0/1928 | 532 | 0 | 76.15% | XP_019765893.1 | Dendroctonus ponderosae |
| TyunCCE19 | i1_LQ_TYUN_c37855/f1p0/1526 | 414 | 0 | 83.09% | XP_019766062.1 | Dendroctonus ponderosae |
| TyunCCE20 | Cluster-27834.3692 | 531 | 0 | 85.50% | XP_019766553.1 | Dendroctonus ponderosae |
| TyunCCE21 | Cluster-27834.10155 | 569 | 0 | 86.12% | XP_019769801.1 | Dendroctonus ponderosae |
| TyunCCE22 | i3_LQ_TYUN_c17625/f1p0/3579 | 266 | 2 × 10−136 | 89.52% | XP_019769801.1 | Dendroctonus ponderosae |
| TyunCCE23 | i1_LQ_TYUN_c34869/f1p0/1995 | 577 | 0 | 69.46% | XP_019769830.1 | Dendroctonus ponderosae |
| TyunCCE24 | Cluster-27834.16736 | 569 | 0 | 75.66% | XP_019772474.1 | Dendroctonus ponderosae |
| TyunCCE25 | i2_LQ_TYUN_c30644/f1p0/2041 | 337 | 0 | 78.34% | XP_019773718.1 | Dendroctonus ponderosae |
| TyunCCE26 | Cluster-27834.18773 | 65 | 9 × 10−7 | 42.86% | XP_023727217.2 | Cryptotermes secundus |
| TyunCCE27 | Cluster-27834.13066 | 308 | 7 × 10−127 | 60.06% | XP_030749177.1 | Sitophilus oryzae |
| TyunCCE28 | Cluster-27834.7029 | 572 | 0 | 47.51% | XP_030754488.1 | Sitophilus oryzae |
| TyunCCE29 | i2_LQ_TYUN_c24874/f1p0/2585 | 308 | 3 × 10−107 | 55.89% | XP_030768230.1 | Sitophilus oryzae |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Yang, B.; Liu, N.; Zhu, J.; Li, Z.; Ze, S.; Yu, J.; Zhao, N. Identification and Characterization of the Detoxification Genes Based on the Transcriptome of Tomicus yunnanensis. Diversity 2022, 14, 23. https://doi.org/10.3390/d14010023
Li W, Yang B, Liu N, Zhu J, Li Z, Ze S, Yu J, Zhao N. Identification and Characterization of the Detoxification Genes Based on the Transcriptome of Tomicus yunnanensis. Diversity. 2022; 14(1):23. https://doi.org/10.3390/d14010023
Chicago/Turabian StyleLi, Wen, Bin Yang, Naiyong Liu, Jiaying Zhu, Zongbo Li, Sangzi Ze, Jinde Yu, and Ning Zhao. 2022. "Identification and Characterization of the Detoxification Genes Based on the Transcriptome of Tomicus yunnanensis" Diversity 14, no. 1: 23. https://doi.org/10.3390/d14010023
APA StyleLi, W., Yang, B., Liu, N., Zhu, J., Li, Z., Ze, S., Yu, J., & Zhao, N. (2022). Identification and Characterization of the Detoxification Genes Based on the Transcriptome of Tomicus yunnanensis. Diversity, 14(1), 23. https://doi.org/10.3390/d14010023