
Genetic Diversity of the Surubim-Do-Iguaçu, a Giant Catfish Species Threatened with Extinction: Recommendations for Species Conservation

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
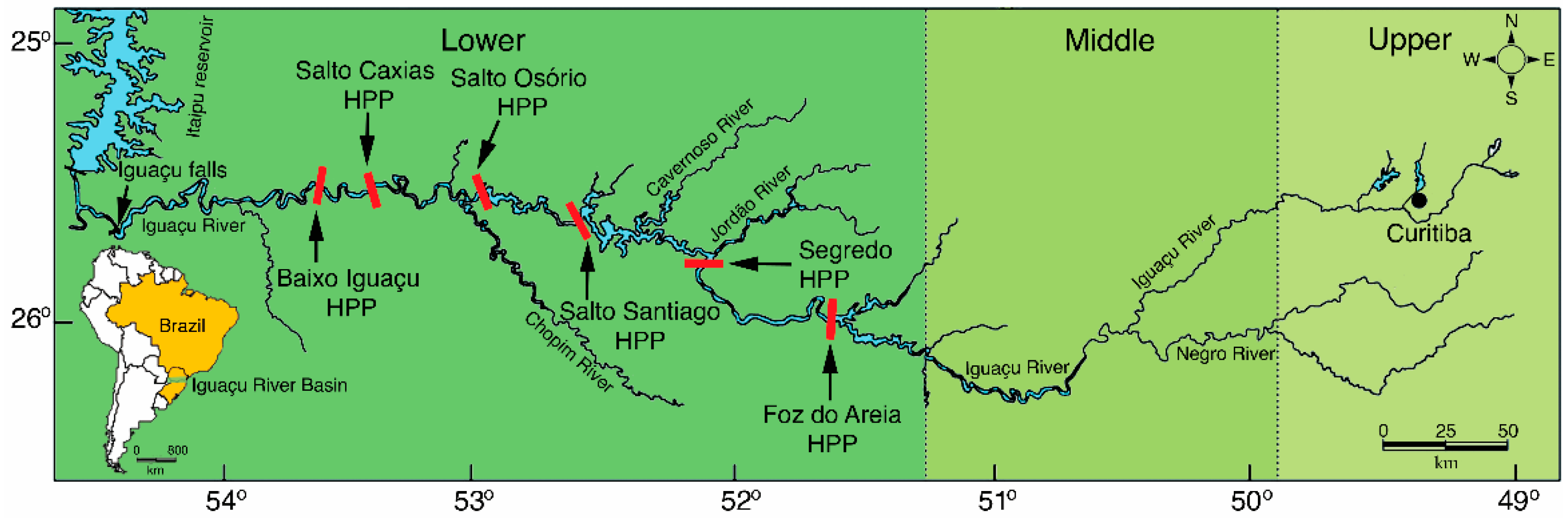
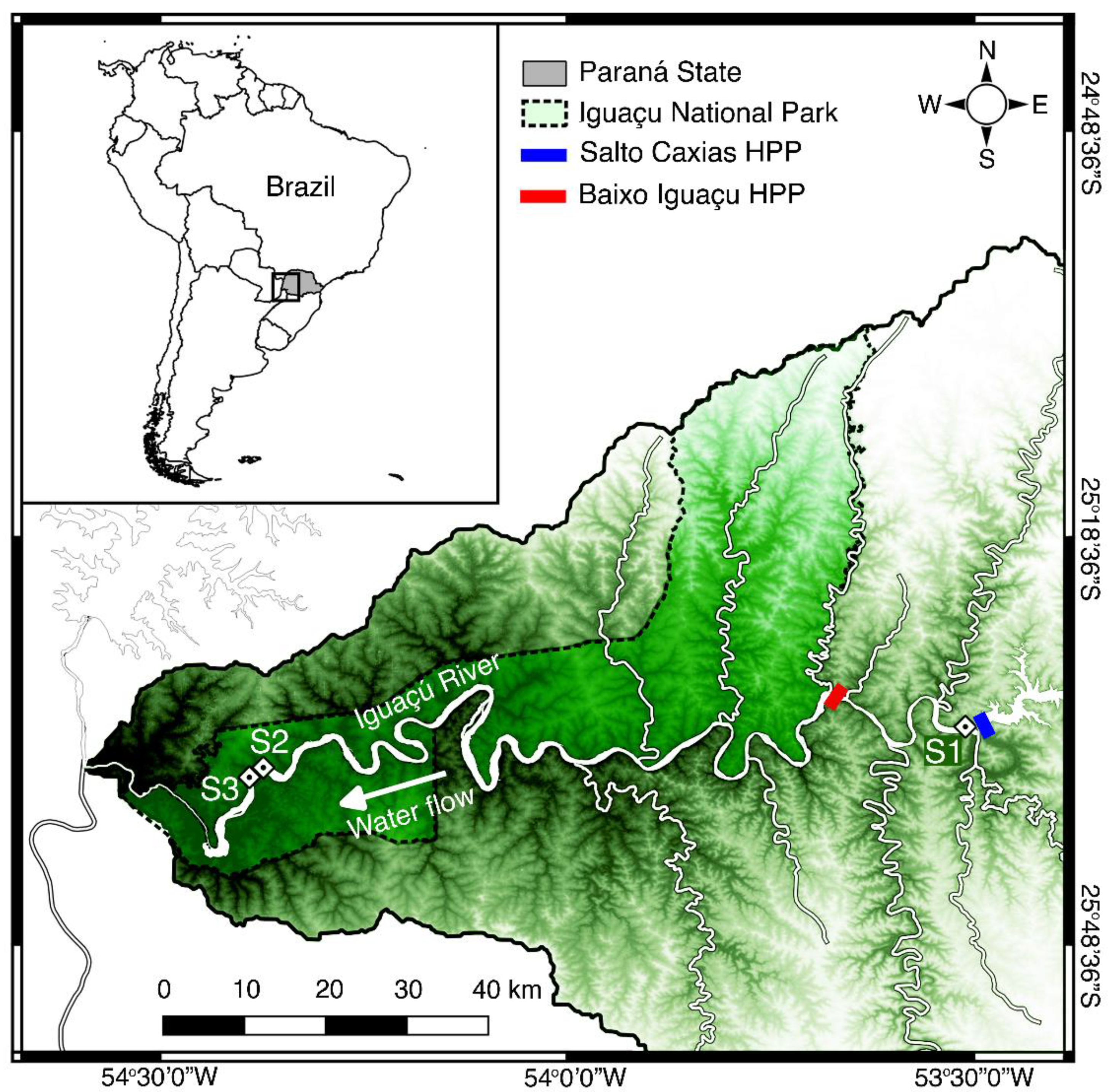
2.1. Sampling
2.2. DNA Extraction and Quantification
2.3. Microsatellite Markers (SSR)
2.4. mtDNA (D-Loop) Haplotypes
2.5. Genetic Diversity Analyses
2.6. Population Structure Analyses
2.7. Demographic Analyses
3. Results
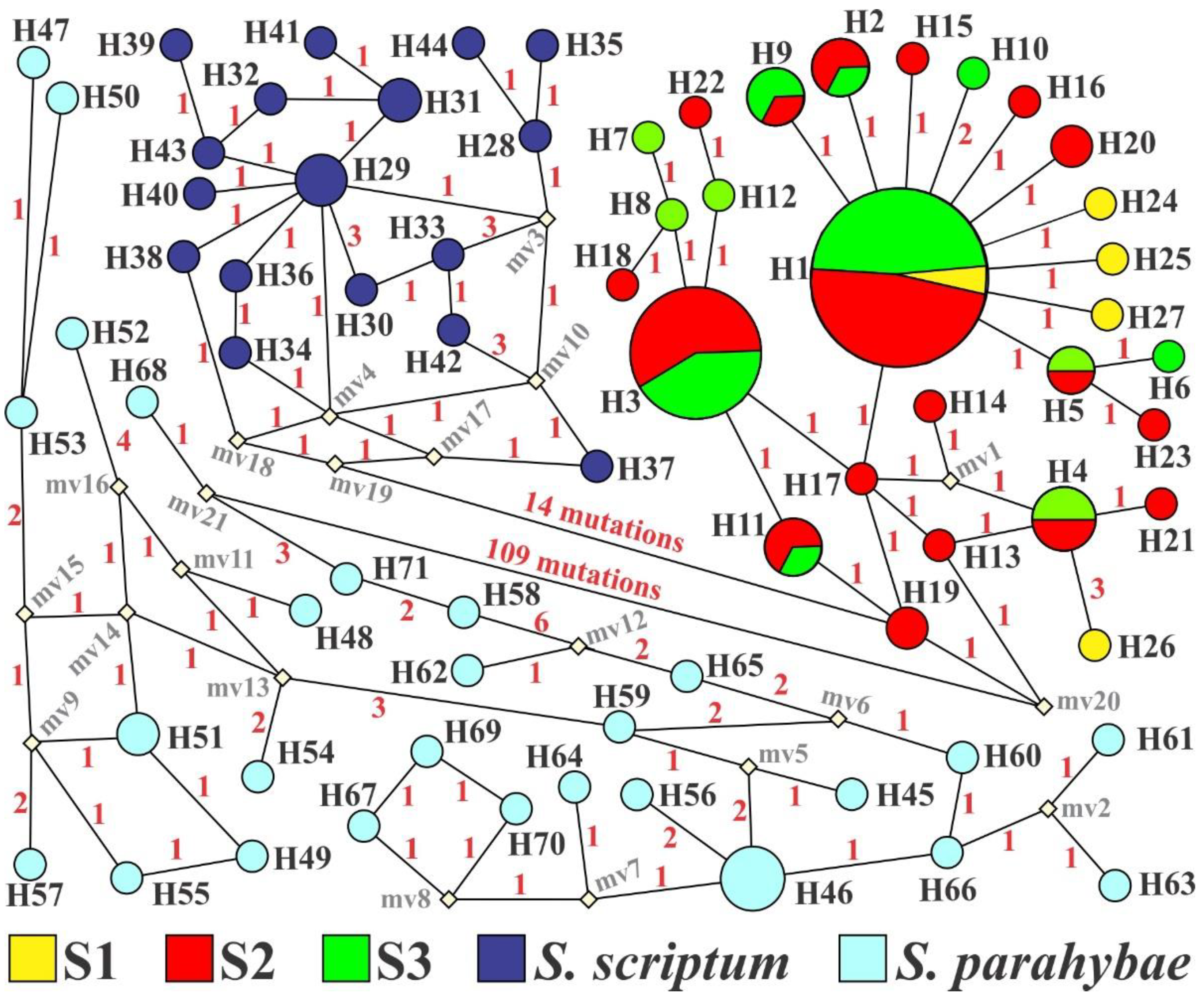
3.1. Genetic Diversity
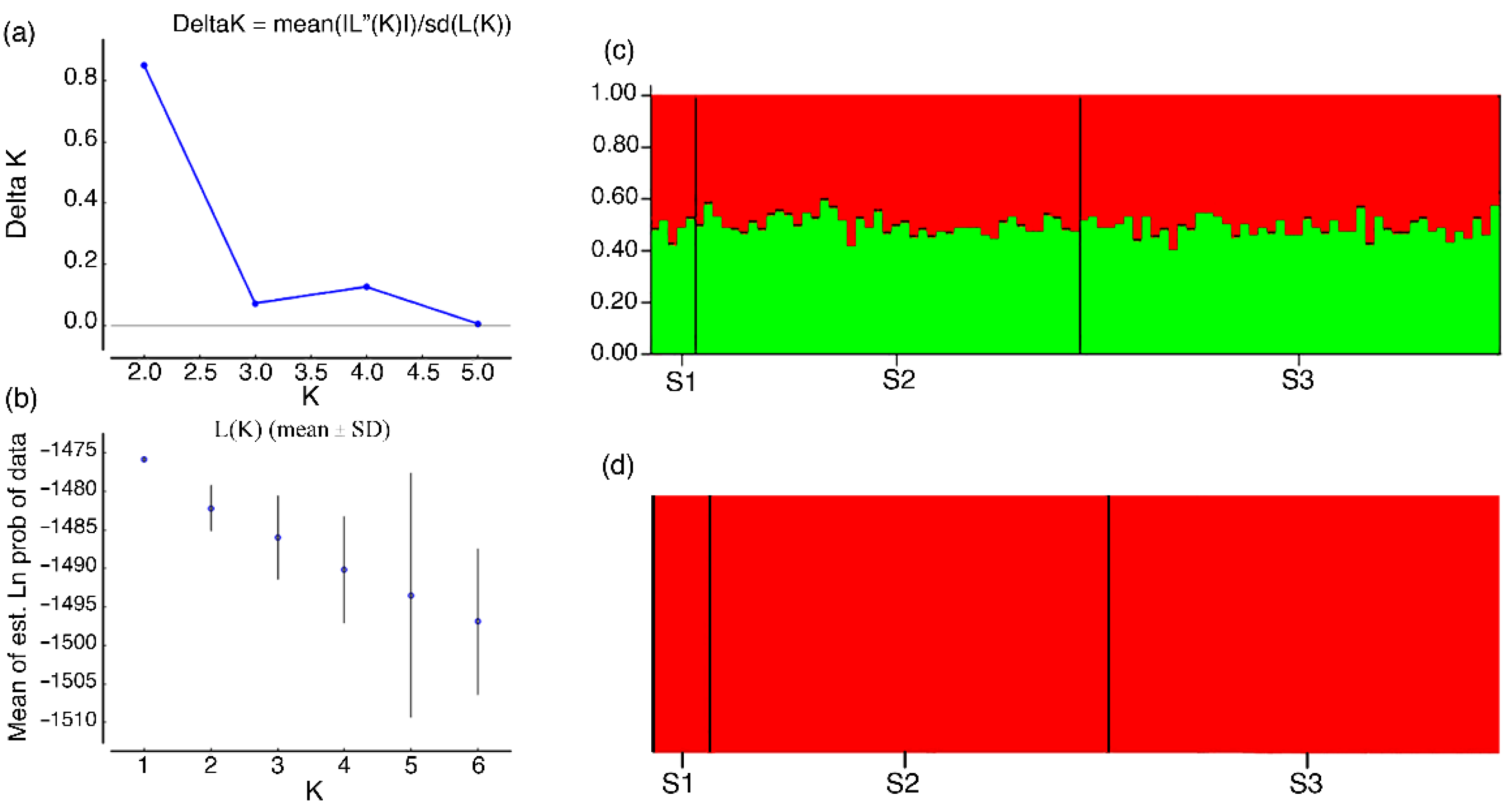
3.2. Population Structure
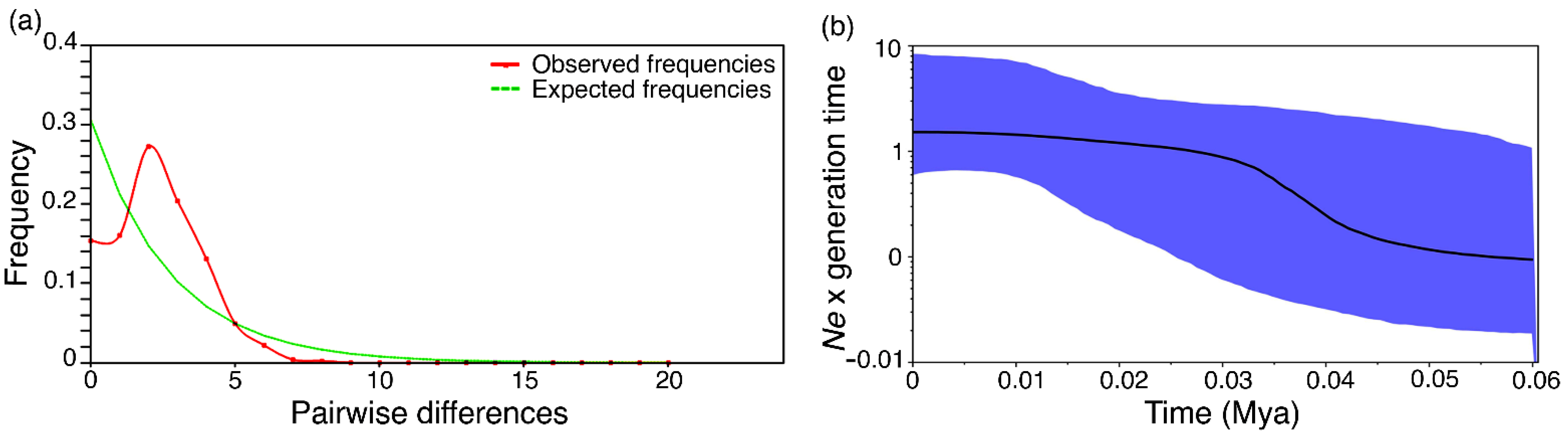
3.3. Demographic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collen, B.; Whitton, F.; Dyer, E.E.; Baillie, J.E.; Cumberlidge, N.; Darwall, W.R.; Pollock, C.; Richman, N.I.; Soulsby, A.M.; Bohm, M. Global patterns of freshwater species diversity, threat and endemism. Glob. Ecol. Biogeogr. 2014, 23, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Grill, G.; Lehner, B.; Lumsdon, A.E.; MacDonald, G.K.; Zarfl, C.; Liermann, C.R. An index-based framework for assessing patterns and trends in river fragmentation and flow regulation by global dams at multiple scales. Environ. Res. Lett. 2015, 10, 015001. [Google Scholar] [CrossRef]
- Grill, G.; Lehner, B.; Thieme, M.; Geenen, B.; Tickner, D.; Antonelli, F.; Babu, S.; Borrelli, P.; Cheng, L.; Crochetiere, H.; et al. Mapping the world’s free-flowing rivers. Nature 2019, 569, 215–221. [Google Scholar] [CrossRef]
- Barbarossa, V.; Schmitt, R.J.P.; Huijbregts, M.A.J.; Zarfl, C.; King, H.; Schipper, A.M. Impacts of current and future large dams on the geographic range connectivity of freshwater fish worldwide. Proc. Natl. Acad. Sci. USA 2020, 117, 3648–3655. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.G.; Souza-Shibatta, L.; Shibatta, O.A.; Sofia, S.H.; Carlsson, J.; Dias, J.H.P.; Makrakis, S.; Makrakis, M.C. Genetic structure and diversity of migratory freshwater fish in a fragmented Neotropical river system. Rev. Fish Biol. Fisher. 2017, 27, 209–231. [Google Scholar] [CrossRef]
- Lira, N.A.; Pompeu, P.S.; Agostinho, C.S.; Agostinho, A.A.; Arcifa, M.S.; Pelicice, F.M. Fish passages in South America: An overview of studied facilities and research effort. Neotrop. Ichthyol. 2017, 15, e160139. [Google Scholar] [CrossRef]
- Makrakis, S.; Bertão, A.P.S.; Silva, J.F.M.; Makrakis, M.C.; Sanz-Ronda, L.F.; Celestino, L.F. Hydropower development and fishways: A need for connectivity in rivers of the Upper Paraná Basin. Sustainability 2019, 11, 3749. [Google Scholar] [CrossRef]
- Nilson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and flow regulation of the world’s large river system. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Instituto Brasileiro de Geografia e Estatística. Indicadores de Desenvolvimento Sustentável, 1st ed.; IBGE/Coordenação de Recursos Naturais e Estudos Ambientais e Coordenação de Geografia: Rio de Janeiro, Brazil, 2015.
- Bueno-Krawczyk, A.C.D.; Guiloski, I.C.; Piancini, L.D.S.; Azevedo, J.C.; Ramsdorf, W.A.; Ide, A.H.; Guimarães, A.T.B.; Cestari, M.M.; Assis, H.C.S. Multibiomarker in fish to evaluate a river used to water public supply. Chemosphere 2015, 135, 247–264. [Google Scholar] [CrossRef]
- Ghisi, N.C.; Larentis, C.; Oliveira, E.C.; Neves, M.P.; Zavaski, A.G.; Roque, A.A.; Wachtel, C.C.; Silva, A.P.; Lima, E.B.S.; Costa, G.O.N.; et al. Environmental assessment of Neotropical streams using fish as bioindicators: A multibiomarker and integrated approach. Hydrobiologia 2020. [Google Scholar] [CrossRef]
- Nimet, J.; Guimarães, A.T.B.; Delariva, R.L. Use of Muscular Cholinesterase of Astyanax bifasciatus (Teleostei, Characidae) as a Biomarker in Biomonitoring of Rural Streams. Bull. Environ. Contam. Toxicol. 2017, 99, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Nimet, J.; Neves, M.P.; Viana, N.P.; Amorim, J.P.A.; Delariva, R.L. Histopathological alterations in gills of a fish (Astyanax bifasciatus) in neotropical streams: Negative effects of riparian forest reduction and presence of pesticides. Environ. Monit. Assess. 2020, 192, 58. [Google Scholar] [CrossRef] [PubMed]
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater Ecoregions of the World: A New Map of Biogeographic Units for Freshwater Biodiversity Conservation. BioScience 2008, 58, 403–414. [Google Scholar] [CrossRef]
- Garavello, J.C.; Pavanelli, C.S.; Suzuki, H.I. Caracterização da ictiofauna do rio Iguaçu. In Reservatório de Segredo: Bases Ecológicas Para o Manejo; Agostinho, A.A., Gomes, L.C., Eds.; Eduem: Maringá, Brazil, 1997; pp. 61–84. [Google Scholar]
- Reis, R.B.; Frota, A.; Depra, G.D.C.; Ota, R.R.; Graca, W.J. Freshwater fishes from Paraná State, Brazil: An annotated list, with comments on biogeographic patterns, threats, and future perspectives. Zootaxa 2020, 4868, 451–494. [Google Scholar] [CrossRef]
- Baumgartner, G.; Pavanelli, C.S.; Baumgartner, D.; Bifi, A.G.; Debona, T.; Frana, V.A. Peixes do Baixo Rio Iguaçu; Eduem: Maringá, Brazil, 2012. [Google Scholar]
- Pini, S.F.R.; Makrakis, M.C.; Neves, M.P.; Makrakis, S.; Shibatta, O.A.; Kashiwaqui, E.A.L. Ichthyofauna in the last free-flowing river of the Lower Iguaçu basin: The importance of tributaries for conservation of endemic species. ZooKeys 2021, 1041, 183–203. [Google Scholar] [CrossRef]
- Assumpção, L.D.; Makrakis, M.C.; Silva, J.F.M.; Moraes, K.A.S.; Pini, S.F.R.; Silva, P.S.; Kashiwaqui, E.A.L.; Gentil, E.; Souza-Shibatta, L.; Shibatta, O.A.; et al. Deep Pools: Ecological Sanctuaries for Steindachneridion melanodermatum, a Large Endemic and Endangered Pimelodid of the Iguaçu River. Water 2021, 13, 1700. [Google Scholar] [CrossRef]
- Brehm, M.; Filippin, R.F.; Moura, R.R. O impacto ambiental causado à ictiofauna do rio Iguaçu pela exploração do potencial hidrelétrico: O caso do surubim do iguaçu (Steindachneridion melanodermatum). Rev. Bras. Ener. 2016, 22, 30–47. [Google Scholar]
- UNESCO [United Nations Educational, Scientific and Cultural Organization]; World Heritage Committee. Convention Concerning the Protection of the World Cultural and Natural Heritage. Report: 36th Session. WHC-12/36. COM/7B. Add. Saint-Petersburg. 2012. Available online: http://whc.unesco.org/archive/2012/whc12-36com-7BAdd-en.pdf (accessed on 23 June 2021).
- Assumpção, L.; Makrakis, S.; Silva, P.S.; Makrakis, M.C. Espécies de peixes ameaçadas de extinção no Parque Nacional do Iguaçu. Biodivers. Bras.-BioBrasil 2017, 7, 4–17. [Google Scholar]
- Delariva, R.L.; Neves, M.P.; Larentis, C.; Kliemann, B.C.K.; Baldasso, M.C.; Wolff, L.L. Fish fauna in forested and rural streams from an ecoregion of high endemism, lower Iguaçu river basin, Brazil. Biota Neotrop. 2018, 18. [Google Scholar] [CrossRef]
- Garavello, J.C. Revision of genus Steindachneridion (Siluriformes: Pimelodidae). Neotrop. Ichthyol. 2005, 3, 607–623. [Google Scholar] [CrossRef]
- Ludwig, L.A.M.; Gomes, E.; Artoni, R.F. Um método de reprodução induzida para o surubim Steindachneridion melanodermatum (Siluriformes: Pimelodidae) do Rio Iguaçu. Ciênc Biol. Saúde 2005, 11, 23–27. [Google Scholar]
- Akama, A.; Netto-Ferreira, A.L.; Zanata, A.M.; Calegari, B.B.; de Figueiredo, C.A.A.; Alves, C.B.M.; Cramer, C.A.; Zawadzki, C.H.; Röpke, C.P.; Moreira, C.R.; et al. Steindachneridion melanodermatum Garavello, 2005. In Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume 6 (Peixes); Instituto Chico Mendes de Conservação da Biodiversidade (Org.), ICMBio: Brasília, DF, Brazil, 2018; pp. 214–217. [Google Scholar]
- Agostinho, A.A.; Pavanelli, C.S.; Suzuki, H.I.; Latini, J.D.; Gomes, L.C.; Hahn, N.S.; Fugi, R.; Domingues, W.M. Reservatório de Salto Caxias-Bases Ecológicas Para o Manejo; Copel: Maringá, Brazil, 2002. [Google Scholar]
- Instituto Chico Mendes de Conservação da Biodiversidade. Lista de Espécies Ameaçadas [ICMBio 2014]. Available online: https://www.icmbio.gov.br/portal/especies-ameacadas-destaque (accessed on 10 September 2021).
- Assumpção, L.; Fávaro, L.F.; Makrakis, S.; Silva, P.S.; Pini, S.F.R.; Kashiwaqui, E.A.L.; Makrakis, M.C. Population structure and reproduction of Steindachneridion melanodermatum (Siluriformes: Pimelodidae), a large catfish endemic to Neotropical ecoregion. Mar. Fresh. Res. 2021, 1, 1–15. [Google Scholar] [CrossRef]
- Souza-Shibatta, L.; Kotelok-Diniz, T.; Ferreira, D.G.; Shibatta, O.A.; Sofia, S.H.; Assumpção, L.; Pini, S.F.R.; Makrakis, S.; Makrakis, M.C. Genetic Diversity of the Endangered Neotropical Cichlid Fish (Gymnogeophagus setequedas) in Brazil. Front. Genet. 2018, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.Q.; Shen, S.K.; Zhang, X.J.; Wang, Y.H.; Sun, W.B. Genetic diversity and population structure of an extremely endangered species: The world’s largest Rhododendron. AoB Plants 2015, 7, plu082. [Google Scholar] [CrossRef]
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.J.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R.; Ballou, J.J.D.; Briscoe, D.D.A. Introduction to Conservation Genetics; Cambridge University Press: New York, NY, USA, 2010. [Google Scholar]
- Allendorf, F.W.; Luikart, G.H.; Aitken, S.N. Conservation and the Genetics of Populations; Wiley Blackwell Press: Oxford, UK, 2012. [Google Scholar]
- Palstra, F.P.; Ruzzante, D.E. Genetic estimates of contemporary effective population size: What can they tell us about the importance of genetic stochasticity for wild population persistence? Mol. Ecol. 2008, 17, 3428–3447. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Hohenlohe, P.A.; Luikart, G. Genomics and the future of conservation genetics. Nat. Rev. Genet. 2010, 11, 698–709. [Google Scholar] [CrossRef]
- Chauhan, T.; Rajiv, K. Molecular markers and their applications in fisheries and aquaculture. Adv. Biosci. Biotechnol. 2010, 1, 281–291. [Google Scholar] [CrossRef]
- Fonseca, F.S.; Domingues, R.R.; Hallerman, E.M.; Hilsdorf, A.W.S. Genetic Diversity of an Imperiled Neotropical Catfish and Recommendations for Its Restoration. Front. Genet. 2017, 8, 196. [Google Scholar] [CrossRef]
- Frantine-Silva, W.; Sofia, S.H.; Orsi, M.L.; Almeida, F.S. DNA barcoding of freshwater ichthyoplankton in the Neotropics as a tool for ecological monitoring. Mol. Ecol. Resour. 2015, 15, 1226–1237. [Google Scholar] [CrossRef]
- Ojeda, A.P.; Hilsdorf, A.W.S.; Leite, A.C.; Yang, A.; Izuno, A.; He, C.; Zhou, C.; Kyogoku, D.; Caneppele, D.; Zhu, D.; et al. Microsatellite records for Volume 8, Issue 4. Conserv. Genet. Resour. 2016, 8, 587–594. [Google Scholar] [CrossRef]
- Paiva, A.L.B.; Kalapothakis, E. Isolation and characterization of microsatellite loci in Pimelodus maculatus (Siluriformes: Pimelodidae). Mol. Ecol. Resour. 2008, 8, 1078–1080. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 5, 35–38. [Google Scholar] [CrossRef]
- Iervolino, F.; Resende, E.K.; Hilsdorf, A.W.S. The lack of genetic differentiation of pacu (Piaractus mesopotamicus) populations in the Upper-Paraguay Basin revealed by the mitochondrial DNA D-loop region: Implications for fishery management. Fish. Res. 2010, 101, 27–31. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Marshall, T.C.; Slate, J.; Kruuk, L.E.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Yang, R.; Boyle, T.J.; Xiyan, J.M. Pop Gene 32: Microsoft Window-Based Freeware for Population Genetic Analysis; University of Alberta: Edmonton, AB, USA, 2000. [Google Scholar]
- Goudet, J. FSTAT, a Program to Estimate and Test Gene Diversities and Fixation Indices (Version 2.9.3). 2001. Available online: http://www.unil.ch/izea/softwares/fstat.html (accessed on 13 November 2020).
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population Genetics Software for Exact Tests and Ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Paixão, R.V.; Ribolli, J.; Zaniboni-Filho, E. Genetic Variation of the Endangered Neotropical Catfish Steindachneridion scriptum (Siluriformes: Pimelodidae). Front. Genet. 2018, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Corander, J.; Marttinen, P.; Sirén, J.; Tang, J. BAPS: Bayesian Analysis of Population Structure; University of Helsinki: Helsinki, Finland, 2013. [Google Scholar]
- Beaumont, M.A.; Rannala, B. The Bayesian revolution in genetics. Nat. Rev. Genet. 2004, 5, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, D.J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 10, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Cons. Gen. Res. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.M. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Luikart, G.; Allendorf, F.W.; Cornuet, J.M.; Sherwin, W.B. Distortion of allele frequency distributions provides a test for recent population bottlenecks. J. Hered. 1998, 89, 238–247. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical test of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [PubMed]
- Drummond, A.J.; Rambaut, A.; Shapiro, B.; Pybus, O.G. Bayesian coalescent inference of past population dynamics from molecular sequences. Mol. Biol Evol. 2005, 22, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef]
- Donaldson, K.A.; Wilson, R.R., Jr. Amphi-panamic geminates of snook (Percoidei: Centropomidae) provide a calibration of the divergence rate in the mitochondrial DNA control region of fishes. Mol. Phylogenet. Evol. 1999, 13, 208–213. [Google Scholar] [CrossRef]
- Waples, R.S.; Do, C. LdNe: A program for estimating effective population size from data on linkage disequilibrium. Mol. Ecol. Resour. 2008, 8, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator v2: Re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour 2014, 14, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 3, 314–331. [Google Scholar]
- Revaldaves, E.; Pereira, L.H.G.; Foresti, F.; Oliveira, C. Isolation and characterization of microsatellite locos in Pseudoplatystoma corruscans (Siluriformes: Pimelodidae) and cross-species amplification. Mol. Ecol. Notes 2005, 5, 463–465. [Google Scholar] [CrossRef]
- Bignotto, T.S.; Prioli, A.J.; Prioli, S.M.A.P.; Maniglia, T.C.; Boni, T.A.; Lucio, L.C.; Gomes, V.N.; Prioli, R.A.; Oliveira, A.V.; Júlio-Junior, H.F.; et al. Genetic divergence between Pseudoplatystoma corruscans and Pseudoplatystoma reticulatum (Siluriformes: Pimelodidae) in the Paraná River Basin. Braz. J. Biol. 2009, 69, 681–689. [Google Scholar] [CrossRef]
- Batista, J.S.; Alves-Gomes, J.A. Phylogeography of Brachyplatystoma rousseauxii (Siluriformes–Pimelodidae) in the Amazon Basin offers preliminary evidence for the first case of “homing” for an Amazonian migratory catfish. Genet. Mol. Res. 2006, 5, 723–740. [Google Scholar]
- Huergo, G.M.; Filgueiras-Souza, R.J.; Batista, J.D.S.; Formiga-Aquino, K.; Alves-Gomes, J.A. Molecular genetics as a tool for fisheries management in the Brazilian Amazon: Piraíba (Brachyplatystoma filamentosum and Brachyplatystoma capapretum) (Siluriformes: Pimelodidae) in white-water rivers. Pan-Am. J. Aquat. Sci. 2011, 6, 280–289. [Google Scholar]
- Ochoa, L.E.; Pereira, L.H.G.; Costa-Silva, G.J.; Roxo, F.F.; Batista, J.S.; Formiga, K.; Foresti, F.; Oliveira, C. Genetic structure and historical diversification of catfish Brachyplatystoma platynemum (Siluriformes: Pimelodidae) in the Amazon basin with implications for its conservation. Ecol. Evol. 2015, 5, 2005–2020. [Google Scholar] [CrossRef]
- So, N.; Maes, G.E.; Volckaert, F.A.M. High genetic diversity in cryptic populations of the migratory sutchi catfish Pangasianodon hypophthalmus in the Mekong River. Heredity 2006, 96, 166–174. [Google Scholar] [CrossRef]
- Batista, J.S.; Farias, I.P.; Formiga-Aquino, K.; Sousa, A.C.B.; Alves-Gomes, J.A. DNA microsatellite markers for "dourada" (Brachyplatystoma rousseauxii, Siluriformes: Pimelodidae), a migratory catfish of utmost importance for fisheries in the Amazon: Development, characterization and inter-specific amplification. Conserv. Genet. Resour. 2010, 2, 5–10. [Google Scholar] [CrossRef]
- Telles, M.P.C.; Collevatti, R.G.; Braga, R.S.; Guedes, L.B.S.; Castro, T.G.; Costa, M.C.; Silva Júnior, N.J.; Barthem, R.B.; Diniz Filho, J.A.F. Geographical genetics of Pseudoplatystoma punctifer (Castelnau, 1855) (Siluriformes, Pimelodidae) in the Amazon basin. Genet. Mol. Res. 2014, 13, 3656–3666. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, M.T.; Hirvonen, H.; Landry, P.A.; Primmer, C.R. The benefits of increasing the number of microsatellites utilized in genetic population studies: An empirical perspective. Hereditas 2004, 141, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Petit, R.; El Mousadik, A.; Pons, O. Identifying populations for conservation on the basis of genetic markers. Conserv. Biol. 1998, 12, 844–855. [Google Scholar] [CrossRef]
- Matoso, D.A.; Silva, M.; Cortinhas, M.C.S.; Cestari, M.M.; Almeida, M.C.; Vicari, M.R.; Artoni, R.F. Two genetic stocks of Steindachneridion melanodermatum living in sympatry in nature and genetic variability of wild parents and F1 generation. Genet. Mol. Res. 2011, 10, 2606–2612. [Google Scholar] [CrossRef] [PubMed]
- Ramella, M.S.; Kroth, M.A.; Meurer, S.; Nuñer, A.P.O.; Zaniboni-Filho, E.; Arisi, A.C.M. Genetic variability in four fish species (Pimelodus maculatus, Prochilodus lineatus, Salminus brasiliensis and Steindachneridion scripta) from Uruguay river basin. Braz. Arch. Biol. Technol. 2006, 49, 589–598. [Google Scholar] [CrossRef]
- Iwersen, L.H.L.; Melo, C.M.R.; Lazoski, C.; Zaniboni-Filho, E.; Ribolli, J. Genetic implications of restocking programs on wild populations of streaked prochilod Prochilodus lineatus. Bol. Inst. Pesca 2019, 45, e497. [Google Scholar] [CrossRef]
- Williamson, K.S.; Murdoch, A.R.; Pearsons, T.N.; Ward, E.J.; Ford, M.J. Factors influencing the relative fitness of hatchery and wild spring Chinook salmon (Oncorhynchus tshawytscha) in the Wenatchee River, Washington, USA. Can. J. Fish. Aquat Sci. 2010, 67, 1840–1851. [Google Scholar] [CrossRef]
- Christie, M.R.; Marine, M.L.; French, R.A.; Blouin, M.S. Genetic adaptation to captivity can occur in a single generation. Proc. Natl. Acad. Sci. USA 2012, 109, 238–242. [Google Scholar] [CrossRef]
- Piorski, N.M.; Sanches, A.; Carvalho-Costa, L.F.; Hatanaka, T.; Carrillo-Avila, M.; Freitas, P.D.; Galetti, P.M., Jr. Contribution of conservation genetics in assessing neotropical freshwater fish biodiversity. Braz. J. Biol. 2008, 68, 1039–1050. [Google Scholar] [CrossRef][Green Version]
- Clark, P.U.; Dyke, A.S.; Shakun, J.D.; Carlson, A.E.; Clark, J.; Wohlfarth, B.; McCabe, A.M. The Last Glacial Maximum. Science 2009, 325, 710–714. [Google Scholar] [CrossRef]
- Costa, C.L.N.; Ivan Perez, S.; Louvise, J.; Tonhatti, C.H.; Clemente-Carvalho, R.B.G.; Petry, A.C.; dos Reis, S.F. Demographic Expansion and Contraction in a Neotropical Fish during the Late Pleistocene-Holocene. Open J. Stat. 2019, 9, 470–483. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. ARLEQUIN v. 3.0: An integrated software/package for population genetics data analysis. Evol. Bioinform. Online 2005, 281, 47–50. [Google Scholar] [CrossRef]
- Grant, W.S.; Bowen, B.W. Shallow population histories in deep evolutionary lineages of marine fishes: Insights from sardines and anchovies and lessons for conservation. J. Hered. 1998, 89, 415–426. [Google Scholar] [CrossRef]
- Franklin, I.R. Evolutionary change in small populations. In Conservation Biology: An Evolutionary Ecological Perspective; Soulé, M.E., Wilcox, B.A., Eds.; Sinauer: Sunderland, MA, USA, 1980; pp. 135–149. [Google Scholar]
- Brito, D.; Fernandez, F.A.S. Dealing with extinction is forever: Understanding the risks faced by small populations. Ciênc. Cul. 2000, 52, 161–170. [Google Scholar]
- Clarke, G.M.; O’dvlyer, C. Genetic variability and population structure of the endangered golden sun moth, Synemon plana. Biol. Conserv. 2000, 92, 371–381. [Google Scholar] [CrossRef]
- Carolsfeld, J.; Harvey, B.; Ross, C.; Baer, A. Migratory Fishes of South America; World Fisheries Trust: Victoria, BC, Canada, 2003. [Google Scholar]
- Makrakis, M.C.; Miranda, L.E.; Makrakis, S.; Fontes, H.M., Jr.; Morlis, W.G.; Dias, J.H.P.; Garcia, J.O. diversity in migratory patterns among Neotropical fishes in a highly regulated river basin. J. Fish. Biol. 2012, 81, 866–881. [Google Scholar] [CrossRef] [PubMed]
- Petrere-Jr, M. Fisheries in large tropical reservoirs in South America. Lakes Reserv. Res. Manag. 1996, 2, 111–133. [Google Scholar] [CrossRef]
- Hilsdorf, A.W.S.; Petrere-Jr, M. Conservação de peixes da Bacia do rio Paraíba do Sul. Ciência Hoje 2002, 30, 62–67. [Google Scholar]
- Vrijenhoek, R.C. Conservation genetics of freshwater fish. J. Fish. Biol. 1998, 53, 394–412. [Google Scholar] [CrossRef]
- Esguícero, A.L.H.; Arcifa, M.S. Fragmentation of a Neotropical migratory fish population by a century-old dam. Hydrobiologia 2010, 638, 41–53. [Google Scholar] [CrossRef]
- Clay, C.H. Design of Fishways and Other Fish. Facilities, 2nd ed.; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Larinier, M. Environmental Issues, Dams and Fish Migration. In Dams, Fish and Fisheries: Opportunities, Challenges and Conflict Resolution; Marmulla, G., Ed.; Fisheries Technical Paper; FAO Food and Agriculture Organization: Rome, Italy, 2001; pp. 45–90. ISBN 92-5-104694-8. [Google Scholar]
- Celestino, L.F.; Sanz-Ronda, F.J.; Miranda, L.E.; Makrakis, M.C.; Dias, J.H.P.; Makrakis, S. Bidirectional connectivity via fish ladders in a large Neotropical river. River Res. Appl. 2019, 35, 236–246. [Google Scholar] [CrossRef]
- Makrakis, S.; Miranda, L.E.; Gomes, L.C.; Makrakis, M.C.; Junior, H.M.F. Ascent of neotropical fish in the Itaipu reservoir fish pass. River Res. Appl. 2011, 27, 511–519. [Google Scholar] [CrossRef]
- Hahn, L.; Martins, E.G.; Nunes, L.D.; Machado, L.S.; Lopes, T.M.; Câmara, L.F. Semi-natural fishway efficiency for goliath catfish (Brachyplatystoma spp.) in a large dam in the Amazon Basin. Hydrobiologia 2020, 849, 323–338. [Google Scholar] [CrossRef]
- Makrakis, S.; Makrakis, M.C.; Wagner, R.L.; Dias, J.H.P.; Gomes, L.C. Utilization of the fish ladder at the Engenheiro Sergio Motta Dam, Brazil, by long distance migrating potamodromous species. Neotrop. Ichthyol. 2007, 5, 197–204. [Google Scholar] [CrossRef]
- Godinho, A.L.; Kynard, B. Migratory fishes of Brazil: Life history and fish passage needs. River Res. Appl. 2008, 25, 702–712. [Google Scholar] [CrossRef]
- Da Silva, P.S.; Makrakis, M.C.; Miranda, L.E.; Makrakis, S.; Assumpção, L.; Paula, S.; Dias, J.H.P.; Marques, H. Importance of reservoir tributaries to spawning of migratory fish in the upper Paraná River. River Res. Appl. 2015, 31, 313–322. [Google Scholar] [CrossRef]
- Zaniboni-Filho, E.; Schulz, U.H. Migratory fishes of the Uruguay River. In Migratory Fishes of the South América: Biology, Fisheries and Conservation Status; Carosfeld, J., Harvey, B., Ross, C., Baer, A., Eds.; World Fisheries Trust: Victoria, BC, Canada, 2003; pp. 161–192. [Google Scholar]
- Chan, S.; Putrea, S.; Hortle, H.G. Using local knowledge to inventory deep pools, important fish habitats in Cambodia. In Proceedings of the 6th Technical Symposium on Mekong Fisheries, Pakse, Laos, 26–28 November 2003; Burnhill, T.J., Hewitt, M.M., Eds.; Mekong River Commission: Vientiane, Laos, 2003; pp. 57–76. [Google Scholar]
- Rakowitz, G.; Berger, B.; Schludermann, E.; Tritthart, M.; Habersack, H.; Keckeis, H. Deep pools of the Danube River: Ecological function or turbulent sink? Hydrobiologia 2014, 729, 143–159. [Google Scholar] [CrossRef]
- Baird, I.G.; Kisouvannalath, P.; Inthaphaisy, V.; Phylavanh, B. The Potential for Ecological Classification as a Tool for Establishing and Monitoring Fish Conservation Zones in the Mekong River. In Technical Report. Environmental Protection and Community Development in Siphandone Wetland Project; CESVI Cooperation and Development: Pakse, Laos, 1998. [Google Scholar]
- Baird, I.G.; Flaherty, M.S. Mekong river fish conservation zones in Southern Laos: Assessing effectiveness using local ecological knowledge. Environ. Manag. 2005, 36, 439–454. [Google Scholar] [CrossRef]
- Poulsen, A.F.; Poeu, O.; Virvong, S.; Suntornratana, U.; Tung, N.T. Deep Pools as Dry Season Fish Habitats in the Mekong Basin. In MRC Technical Paper No. 4; Mekong River Commission: Phnom Penh, Cambodia, 2002. [Google Scholar]
- Sard, N.M.; Johnson, M.A.; Jacobson, D.P.; Hogansen, M.J.; O’Malley, K.G.; Banks, M.A. Genetic monitoring guides adaptive management of a migratory fish reintroduction program. Anim. Conserv. 2016, 19, 570–577. [Google Scholar] [CrossRef]
- Lusardi, R.A.; Moyle, P.B. Two-way trap-and-haul as a conservation strategy for anadromous salmonids. Fisheries 2017, 42, 478–487. [Google Scholar] [CrossRef]
- Sajan, S.; Mercy, A.T.V.; Malika, V. Age, growth and population dynamics of an endangered fish Sahyadria denisonii (Day 1865) from the western ghats hotspot of India. Asian Fish. Sci. 2015, 28, 130–142. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Microsatellites | mtDNA | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | A | HO | HE | FIS | Ne | Nh | h | π | D | Fs | |||
| S1 | 5 | 19 | 3.167 | 2.797 | 0.667 | 0.567 | −0.066 | Infinite (CI 95% = 1.1–infinite) | 5 | 1.000 | 0.0048 | −0.807 | −0.845 |
| S2 | 45 | 36 | 6.000 | 3.754 | 0.693 | 0.626 | −0.094 | 39.6 (CI 95% = 22.7–88.1) | 18 | 0.858 | 0.0032 | −1.217 | −1.225 |
| S3 | 45 | 33 | 5.500 | 3.298 | 0.644 | 0.606 | −0.051 | 78 (CI 95% = 32.5–infinite) | 12 | 0.759 | 0.0029 | −1.059 | −1.013 |
| All samples | 95 | 38 | 6.333 | 3.588 | 0.668 | 0.619 | −0.073 | 148.4 (CI 95% = 70.2–997.6) | 27 | 0.846 | 0.0033 | −1.483 | −2.274 |
| Microsatellite | mtDNA | |||
|---|---|---|---|---|
| Pairwise Comparisons | FST | p-Value | ΦST | p-Value |
| S1 × S2 | −0.01181 | 0.591 | 0.092 | 0.063 |
| S1 × S3 | −0.01503 | 0.721 | 0.071 | 0.162 |
| S2 × S3 | −0.00012 | 0.375 | −0.016 | 0.936 |
| Samples | N | Teste Wilcoxon Sign-Rank | Allele Frequency Distribution | |||||
|---|---|---|---|---|---|---|---|---|
| IAM a | TPM b | SMM c | ||||||
| He/Hd | p-Value | He/Hd | p-Value | He/Hd | p-Value | |||
| S1 | 5 | 5/1 | 0.023 *e | 5/1 | 0.039 *e | 5/1 | 0.078 | L-shaped |
| S2 | 45 | 6/0 | 0.007 *e | 6/0 | 0.007 *e | 2/4 | 0.718 | L-shaped |
| S3 | 45 | 6/0 | 0.007 *e | 4/2 | 0.039 *e | 1/5 | 0.945 | L-shaped |
| All the samples | 95 | 6/0 | 0.007 *e | 4/2 | 0.039 *e | 2/4 | 0.921 | L-shaped |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza-Shibatta, L.; Ferreira, D.G.; Assumpção, L.d.; Shibatta, O.A.; Sofia, S.H.; Pini, S.F.R.; Silva, P.S.d.; Makrakis, S.; Makrakis, M.C. Genetic Diversity of the Surubim-Do-Iguaçu, a Giant Catfish Species Threatened with Extinction: Recommendations for Species Conservation. Diversity 2022, 14, 16. https://doi.org/10.3390/d14010016
Souza-Shibatta L, Ferreira DG, Assumpção Ld, Shibatta OA, Sofia SH, Pini SFR, Silva PSd, Makrakis S, Makrakis MC. Genetic Diversity of the Surubim-Do-Iguaçu, a Giant Catfish Species Threatened with Extinction: Recommendations for Species Conservation. Diversity. 2022; 14(1):16. https://doi.org/10.3390/d14010016
Chicago/Turabian StyleSouza-Shibatta, Lenice, Dhiego G. Ferreira, Lucileine de Assumpção, Oscar A. Shibatta, Silvia H. Sofia, Suelen F. R. Pini, Patricia S. da Silva, Sergio Makrakis, and Maristela C. Makrakis. 2022. "Genetic Diversity of the Surubim-Do-Iguaçu, a Giant Catfish Species Threatened with Extinction: Recommendations for Species Conservation" Diversity 14, no. 1: 16. https://doi.org/10.3390/d14010016
APA StyleSouza-Shibatta, L., Ferreira, D. G., Assumpção, L. d., Shibatta, O. A., Sofia, S. H., Pini, S. F. R., Silva, P. S. d., Makrakis, S., & Makrakis, M. C. (2022). Genetic Diversity of the Surubim-Do-Iguaçu, a Giant Catfish Species Threatened with Extinction: Recommendations for Species Conservation. Diversity, 14(1), 16. https://doi.org/10.3390/d14010016