
Application of eDNA Metabarcoding for Detecting Anura on a Tropical Island

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
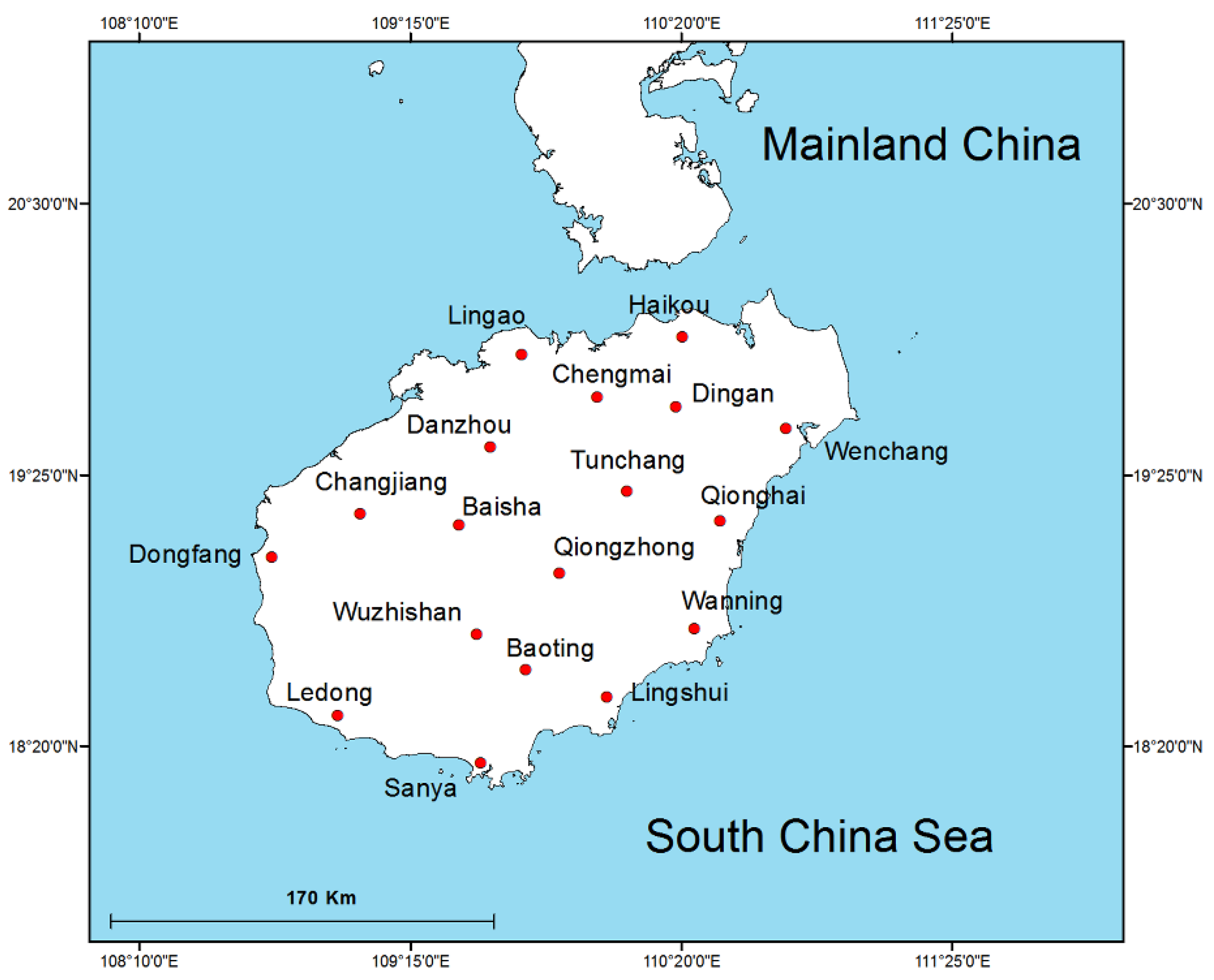
2.1. Study Area and Sample Location
2.2. Traditional Methods
2.3. Reference DNA Database
2.4. eDNA Metabarcoding Sampling
2.5. Statistical Analyses
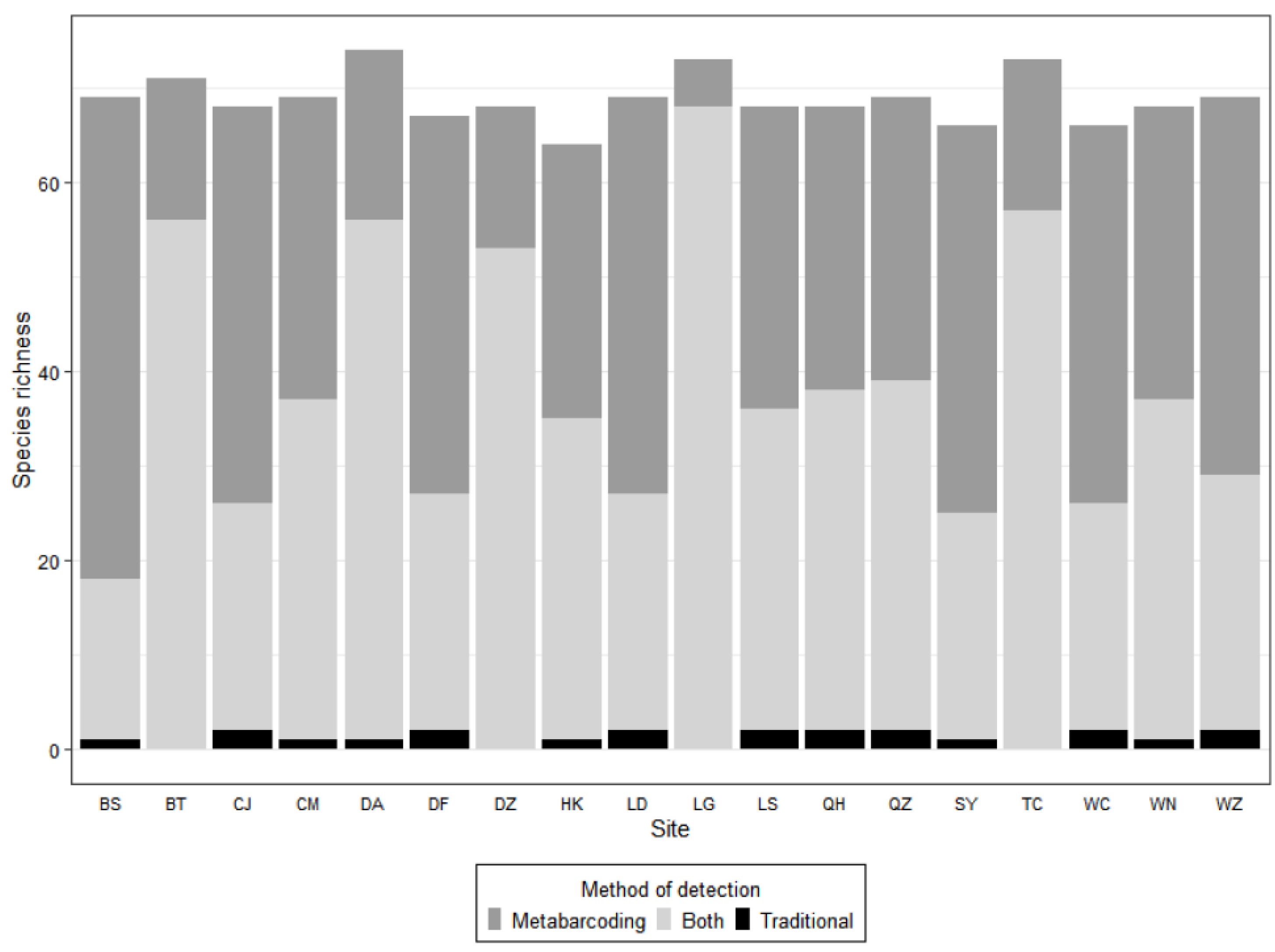
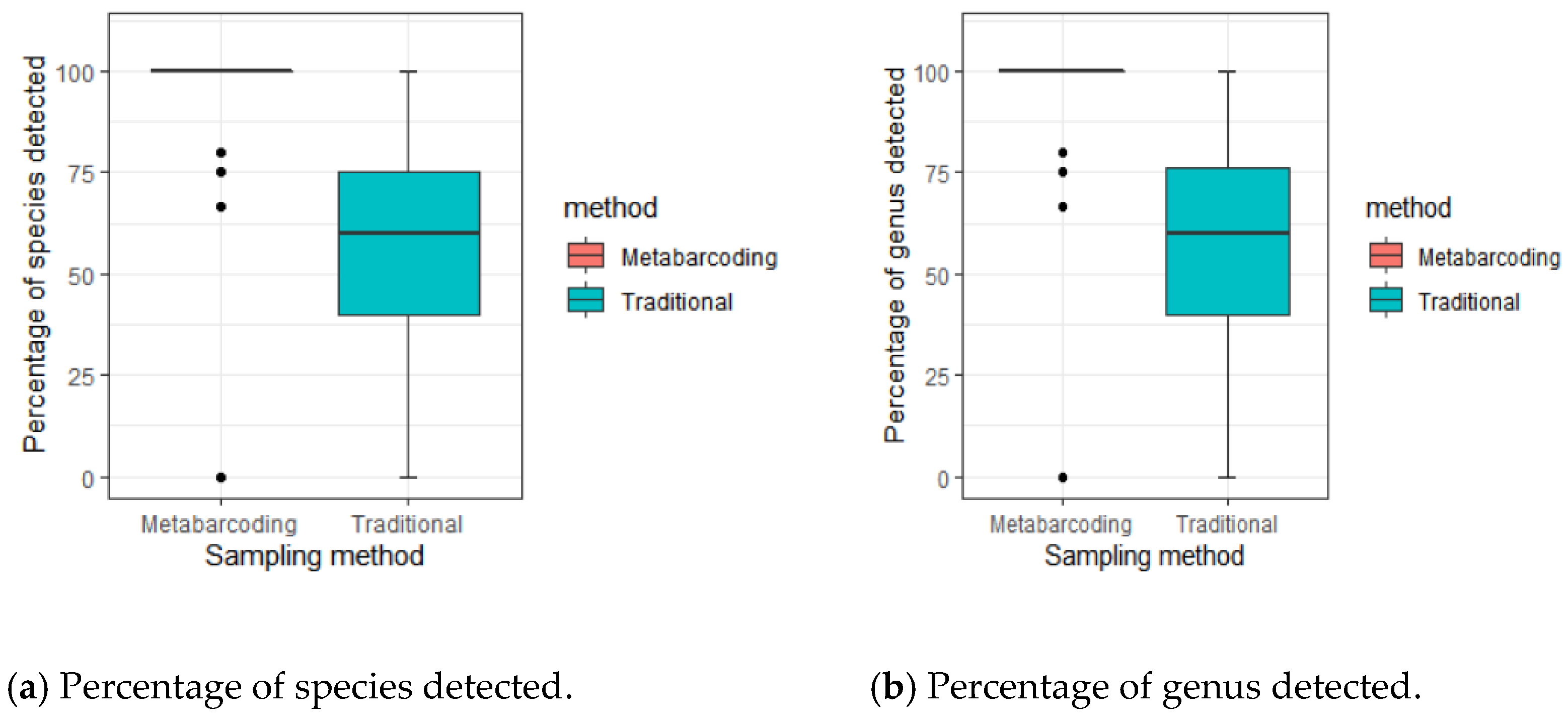
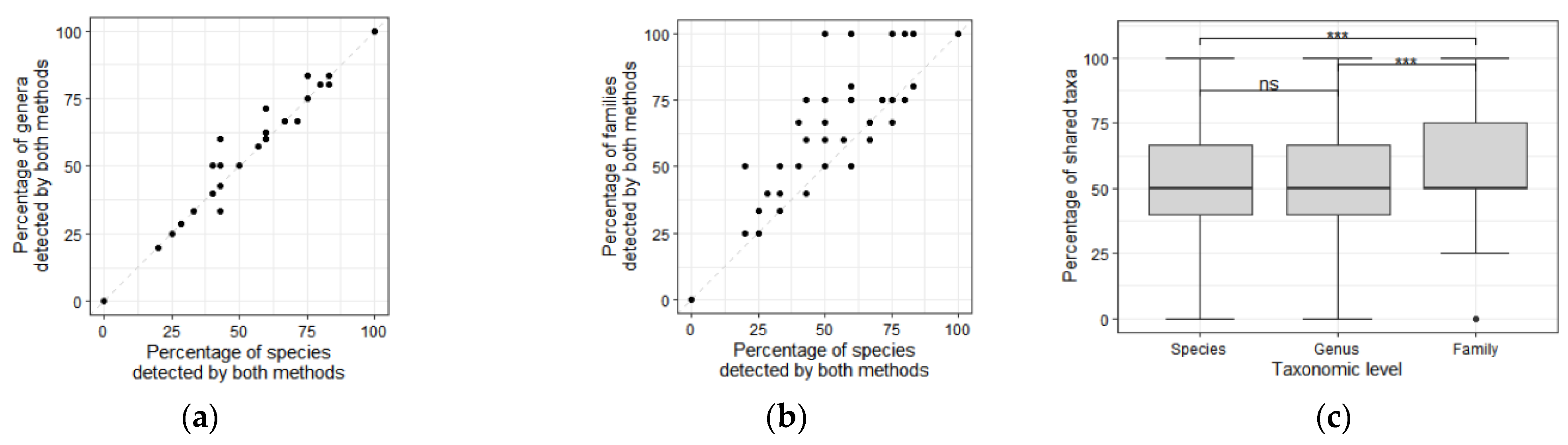
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [Green Version]
- Scheele, B.C.; Pasmans, F.; Skerratt, L.F.; Berger, L.; Martel, A.N.; Beukema, W.; Acevedo, A.A.; Burrowes, P.A.; Carvalho, T.; Catenazzi, A.; et al. Amphibian fungal panzootic causes catastrophic and ongoing loss of biodiversity. Science 2019, 363, 1459–1463. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.G.; Fonseca, C.R.; Haddad, C.F.B.; Batista, R.F.; Prado, P.I. Habitat split and the global decline of amphibians. Science 2007, 318, 1775–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wake, D.B.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 105, 11466–11473. [Google Scholar] [CrossRef] [Green Version]
- Murphy, B.R.; Willis, D.W. Fisheries Techniques; American Fisheries Society: Bethesda, MD, USA, 1996. [Google Scholar]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; De Vere, N.; et al. Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Yates, M.C.; Fraser, D.J.; Derry, A.M. Meta-analysis supports further refinement of eDNA for monitoring aquatic species-specific abundance in nature. Environ. DNA 2019, 1, 5–13. [Google Scholar] [CrossRef]
- Lawson Handley, L. How will the ‘molecular revolution’ contribute to biological recording? Biol. J. Linn. Soc. 2015, 115, 750–766. [Google Scholar] [CrossRef] [Green Version]
- Hänfling, B.; Lawson Handley, L.; Read, D.S.; Hahn, C.; Li, J.; Nichols, P.; Blackman, R.C.; Oliver, A.; Winfield, I.J. Environmental DNA metabarcoding of lake fish communities reflects long-term data from established survey methods. Mol. Ecol. 2016, 25, 3101–3119. [Google Scholar] [CrossRef] [Green Version]
- Lim, N.K.; Tay, Y.C.; Srivathsan, A.; Tan, J.W.; Kwik, J.T.; Baloğlu, B.; Meier, R.; Yeo, D.C. Next-generation freshwater bioassessment: eDNA metabarcoding with a conserved metazoan primer reveals species-rich and reservoir-specific communities. R. Soc. Open Sci. 2016, 3, 160635. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, P.F.; Moller, P.R.; Sigsgaard, E.E.; Knudsen, S.W.; Jorgensen, O.A.; Willerslev, E. Environmental DNA from Seawater Samples Correlate with Trawl Catches of Subarctic, Deepwater Fishes. PLoS ONE 2016, 11, e0165252. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Hatton-Ellis, T.W.; Lawson Handley, L.J.; Kimbell, H.S.; Benucci, M.; Peirson, G.; Hänfling, B. Ground-truthing of a fish-based environmental DNA metabarcoding method for assessing the quality of lakes. J. Appl. Ecol. 2019, 56, 1232–1244. [Google Scholar] [CrossRef]
- Lopes, C.M.; Baêta, D.; Valentini, A.; Lyra, M.L.; Sabbag, A.F.; Gasparini, J.L.; Dejean, T.; Haddad, C.F.B.; Zamudio, K.R. Lost and found: Frogs in a biodiversity hotspot rediscovered with environmental DNA. Mol. Ecol. 2021, 30, 3289–3298. [Google Scholar] [CrossRef]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. Environmental DNA for Biodiversity Research and Monitoring; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Nichols, R.V.; Vollmers, C.; Newsom, L.A.; Wang, Y.; Heintzman, P.D.; Leighton, M.; Green, R.E.; Shapiro, B. Minimizing polymerase biases in metabarcoding. Mol. Ecol. Resour. 2018, 8, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Hou, X.; Xu, C.; Qin, M.; Wang, S.; Wei, L.; Wang, Y.; Liu, X.; Li, Y. Validating eDNA measurements of the richness and abundance of anurans at a large scale. J. Anim. Ecol. 2021, 90, 1466–1479. [Google Scholar] [CrossRef]
- Strickler, K.M.; Fremier, A.K.; Goldberg, C.S. Quantifying effects of UV-B, temperature, and pH on eDNA degradation in aquatic microcosms. Biol. Conserv. 2015, 183, 85–92. [Google Scholar] [CrossRef]
- Lindahl, T. Instability and decay of the primary structure of DNA. Nature 1993, 362, 709–715. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Goldberg, C.S.; Arkle, R.S.; Waits, L.P. Factors influencing detection of eDNA from a stream-dwelling amphibian. Mol. Ecol. Resour. 2014, 14, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Heyer, R.; Donnelly, M.A.; Foster, M.; Mcdiarmid, R. Measuring and Monitoring Biological Diversity: Standard Methods for Amphibibas; Smithsonian Institution Press: Washington, DC, USA; London, UK, 1994; pp. 103–107. [Google Scholar]
- Rödel, M.O.; Ernst, R. Measuring and Monitoring Amphibian Diversity in Tropical Forests. I. An Evaluation of Methods with Recommendations for Standardization. Soc. Trop. Ecol. 2004, 10, 1–14. [Google Scholar]
- Li, Y.; Xu, F.; Guo, Z.; Liu, X.; Jin, C.; Wang, Y.; Wang, S. Reduced predator species richness drives the body gigantism of a frog species on the Zhoushan Archipelago in China. J. Anim. Ecol. 2011, 80, 171–182. [Google Scholar] [CrossRef]
- Ribeiro-Júnior, M.A.; Gardner, T.A.; Ávila-Pires, T.C.S. Evaluating the Effectiveness of Herpetofaunal Sampling Techniques across a Gradient of Habitat Change in a Tropical Forest Landscape. J. Herpetol. 2008, 42, 733–749. [Google Scholar] [CrossRef]
- Yiming, L.; Zhengjun, W.; Duncan, R.P. Why islands are easier to invade: Human influences on bullfrog invasion in the Zhoushan archipelago and neighboring mainland China. Oecologia 2006, 148, 129–136. [Google Scholar] [CrossRef]
- Yiming, L.I.; Niemelä, J.; Dianmo, L.I. Nested Distribution of Amphibians in the Zhoushan archipelago, China: Can Selective Extinction Cause Nested Subsets of Species? Oecologia 1998, 113, 557–564. [Google Scholar] [CrossRef]
- Liu, X.; Wang, S.; Ke, Z.; Cheng, C.; Wang, Y.; Zhang, F.; Xu, F.; Li, X.; Gao, X.; Jin, C.; et al. More invaders do not result in heavier impacts: The effects of non-native bullfrogs on native anurans are mitigated by high densities of non-native crayfish. J. Anim. Ecol. 2018, 87, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Haitao, S.; Ermi, Z.; Lijun, W. Amphibians and Reptiles in Hainan; Science Press: Beijing, China, 2011. [Google Scholar]
- Bai, C.; Liu, X.; Fisher, M.C.; Garner, T.W.J.; Li, Y. Global and endemic Asian lineages of the emerging pathogenic fungus Batrachochytrium dendrobatidis widely infect amphibians in China. Divers. Distrib. 2012, 18, 307–318. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, W.; Gao, X.U.; Li, X.; Yan, S.; Liu, X.; Yang, J.; Gao, Z.; Li, Y. Population size and time since island isolation determine genetic diversity loss in insular frog populations. Mol. Ecol. 2014, 23, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Ficetola, G.F.; Coissac, E.; Zundel, S.; Riaz, T.; Shehzad, W.; Bessière, J.; Taberlet, P.; Pompanon, F. An in silico approach for the evaluation of DNA barcodes. BMC Genom. 2010, 11, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, F.; Mercier, C.; Bonin, A.; Le Bras, Y.; Taberlet, P.; Coissac, E. Obitools: A unix-inspired software package for DNA metabarcoding. Mol. Ecol. Resour. 2016, 16, 176–182. [Google Scholar] [CrossRef]
- He, L.; Miao, X.; Lv, G.; Yang, P.; Wu, W.; Jia, L. Nutritional rehabilitation of mitochondrial aberrations in aplastic anaemia. Br. J. Nutr. 2011, 105, 1180–1187. [Google Scholar] [CrossRef] [Green Version]
- Valentin, R.E.; Maslo, B.; Lockwood, J.L.; Pote, J.; Fonseca, D.M. Real-time PCR assay to detect brown marmorated stink bug, Halyomorpha halys (Stal), in environmental DNA. Pest Manag. Sci. 2016, 72, 1854–1861. [Google Scholar] [CrossRef] [PubMed]
- Dejean, T. Improved detection of an alien invasive species through environmental DNA barcoding: The example of the Ameriacan bullfrog Lithobates catesbeianus. J. Appl. Ecol. 2012, 49, 953–959. [Google Scholar] [CrossRef]
- Goldberg, C.S. Critical considerations for the application of environmental DNA methods to detect aquatic species. Methods Ecol. Evol. 2016, 7, 1299–1307. [Google Scholar] [CrossRef]
- McKee, A.M. The effect of dilution and the use of a post-extraction nucleic acid purification column on the accuracy, precision, and inhibition of environmental DNA samples. Biol. Conserv. 2015, 183, 70–76. [Google Scholar] [CrossRef]
- Bálint, M.; Nowak, C.; Márton, O.; Pauls, S.U.; Wittwer, C.; Aramayo, J.L.; Jansen, M. Accuracy, limitations and cost efficiency of eDNA-based community survey in tropical frogs. Mol. Ecol. Resour. 2018, 18, 1415–1426. [Google Scholar] [CrossRef]
- Chao, A. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef]
- Chiu, C.H.; Wang, Y.T.; Walther, B.A.; Chao, A. An improved nonparametric lower bound of species richness via a modified good-turing frequency formula. Biometrics 2014, 70, 671–682. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org/ (accessed on 3 May 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Stevens, M.H.H. VEGAN: Community Ecology Package. R Package Version 2.4-1. 2016. Available online: https://CRAN.R-project.org/package=vegan (accessed on 3 May 2021).
- Dinno, A. Dunn. Test: Dunn’s Test of Multiple Comparisons Using Rank Sums. R Package Version 1.3.4. 2017. Available online: https://CRAN.R-project.org/package=dunn.test (accessed on 3 May 2021).
- Ficetola, G.F.; Boyer, F.; Valentini, A.; Bonin, A.; Meyer, A.; Dejean, T.; Gaboriaud, C.; Usseglio-Polatera, P.; Taberlet, P. Comparison of markers for the monitoring of freshwater benthic biodiversity through DNA metabarcoding. Mol. Ecol. 2021, 30, 3189–3202. [Google Scholar] [CrossRef]
- Zinger, L.; Donald, J.; Brosse, S.; Gonzalez, M.A.; Iribar, A.; Leroy, C.; Lopes, C.M. Advances and Prospects of Environmental DNA in Neotropical Rainforests; Academic Press: Cambridge, MA, USA, 2020; Volume 62, pp. 331–373. [Google Scholar]
- Tsuji, S.; Ushio, M.; Sakurai, S.; Minamoto, T.; Yamanaka, H. Water temperature-dependent degradation of environmental DNA and its relation to bacterial abundance. PLoS ONE 2017, 12, e0176608. [Google Scholar] [CrossRef] [PubMed]
- Cilleros, K.; Valentini, A.; Allard, L.; Dejean, T.; Etienne, R.; Grenouillet, G.; Iribar, A.; Taberlet, P.; Vigouroux, R.; Brosse, S. Unlocking biodiversity and conservation studies in high-diversity environments using environmental DNA (eDNA): A test with Guianese freshwater fishes. Mol. Ecol. Resour. 2019, 19, 27–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasso, T.; Lopes, C.M.; Valentini, A.; Dejean, T.; Zamudio, K.R.; Haddad, C.F.; Martins, M. Environmental dna characterization of amphibian communities in the brazilian atlantic forest: Potential application for conservation of a rich and threatened fauna. Biol. Conserv. 2017, 215, 225–232. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region Number | Region Name | Latitude | Longitude | Date of TLTM Sampling | Date of eDNA Sampling | Sample Code |
|---|---|---|---|---|---|---|
| 1 | Haikou | 110.33462 | 19.97135 | 2018-4-20 | 2018 | HK |
| 2 | Wenchang | 110.74983 | 19.60477 | 2018-4-21 | 2018 | WC |
| 3 | Qionghai | 110.48429 | 19.23553 | 2018-4-22 | 2018 | QH |
| 4 | Wanning | 110.38378 | 18.80618 | 2018-4-23 | 2018 | WN |
| 5 | Lingshui | 110.03361 | 18.53303 | 2018-4-24 | 2018 | LS |
| 6 | Sanya | 109.52814 | 18.27064 | 2018-4-25 | 2018 | SY |
| 7 | Ledong | 108.95645 | 18.46054 | 2018-4-26 | 2018 | LD |
| 8 | Dongfang | 108.69555 | 19.09091 | 2018-4-27 | 2018 | DF |
| 9 | Changjiang | 109.0469 | 19.26384 | 2018-4-28 | 2018 | CJ |
| 10 | Baisha | 109.44084 | 19.21888 | 2018-4-29 | 2018 | BS |
| 11 | Danzhou | 109.56594 | 19.53215 | 2018-4-30 | 2018 | DZ |
| 12 | Lingao | 109.69121 | 19.90166 | 2018-5-1 | 2018 | LG |
| 13 | Chengmai | 109.99477 | 19.73185 | 2018-5-2 | 2018 | CM |
| 14 | Dingan | 110.30891 | 19.69298 | 2018-5-3 | 2018 | DA |
| 15 | Tunchang | 110.11322 | 19.35441 | 2018-5-4 | 2018 | TC |
| 16 | Qiongzhong | 109.84268 | 19.0279 | 2018-5-5 | 2018 | QZ |
| 17 | Wuzhishan | 109.51214 | 18.78224 | 2018-5-6 | 2018 | WZ |
| 18 | Baoting | 109.70767 | 18.64269 | 2018-5-8 | 2018 | BT |
| Species | Traditional Method | eDNA Metabarcoding | Historical Records |
|---|---|---|---|
| Duttaphrynus melanostictus | + | + | + |
| Fejervarya multistriata | + | + | + |
| Hoplobatrachus rugulosus | + | + | + |
| Hyla simplex | + | + | + |
| Kalophrynus pleurostigma | + | + | + |
| Microhyla ornata | + | + | + |
| Microhyla pulchra | + | + | + |
| Polypedates megacephalus | + | + | + |
| Sylvirana guentheri | + | + | + |
| Fejervarya kawamurai | + | + | |
| Kaloula pulchra | + | + | |
| Limnonectes fragilis | + | + | |
| Microhyla butleri | + | + | |
| Micryletta inornata | + | + | |
| Occidozyga martensii | + | + | |
| Amolops hainanensis | + | ||
| Amolops torrentis | + | ||
| Buergeria oxycephala | + | ||
| Chirixalus doriae | + | ||
| Fejervarya moodiei | + | ||
| Hylarana macrodactyla | + | ||
| Hylarana spinulosa | + | ||
| Hylarana taipehensis | + | ||
| Ingerophrynus ledongensis | + | ||
| Leptobrachium hainanense | + | ||
| Liuixalus hainanus | + | ||
| Liuixalus ocellatus | + | ||
| Liuixalus romeri | + | ||
| Microhyla heymonsi | + | ||
| Nidirana hainanensis | + | ||
| Occidozyga lima | + | ||
| Odorrana graminea | + | ||
| Odorrana hainanensis | + | ||
| Odorrana leporipes | + | ||
| Odorrana nasuta | + | ||
| Parapelophryne scalpta | + | ||
| Polypedates mutus | + | ||
| Rana johnsi | + | ||
| Rhacophorus rhodopus | + | ||
| Rohanixalus vittatus | + | ||
| Theloderma albopunctatum | + | ||
| Theloderma corticale | + | ||
| Tylototriton hainanensis | + | ||
| Zhangixalus dennysi | + | ||
| Zhangixalus yinggelingensis | + |
| Method | Species Richness | Chao II | Chao II.low | Chao II.up |
|---|---|---|---|---|
| eDNA | 15 | 15 | 15 | 15.057 |
| Traditional | 9 | 9 | 9 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Song, T.; Hou, X.; Qin, M.; Xu, C.; Li, Y. Application of eDNA Metabarcoding for Detecting Anura on a Tropical Island. Diversity 2021, 13, 440. https://doi.org/10.3390/d13090440
Li W, Song T, Hou X, Qin M, Xu C, Li Y. Application of eDNA Metabarcoding for Detecting Anura on a Tropical Island. Diversity. 2021; 13(9):440. https://doi.org/10.3390/d13090440
Chicago/Turabian StyleLi, Wenhao, Tianjian Song, Xianglei Hou, Mingshuo Qin, Chunxia Xu, and Yiming Li. 2021. "Application of eDNA Metabarcoding for Detecting Anura on a Tropical Island" Diversity 13, no. 9: 440. https://doi.org/10.3390/d13090440
APA StyleLi, W., Song, T., Hou, X., Qin, M., Xu, C., & Li, Y. (2021). Application of eDNA Metabarcoding for Detecting Anura on a Tropical Island. Diversity, 13(9), 440. https://doi.org/10.3390/d13090440