Abstract
This study analyzed the flora, life form, and vegetation of the Nakdong River wetland. Vegetation analysis was performed on 37 plots using the phytosociological method of the Zürich-Montpellier School. PCA analysis was conducted by using the vegetation data (ground cover of class; 1~9) of 37 plots surveyed by phytosociological method. PCA (Principal Component Analysis) was used to statistically analyze the objectivity of the community classification and the character species. The traditional classification and mathematical statistic methods were used. A total of 82 taxa belonging to 28 families, 65 genera, 72 species, 2 subspecies, and 8 varieties were present in the vegetation of the survey area. The life form was analyzed to be the Th-R5-D4-e type. The communities were classified into seven communities: Miscanthus sacchariflorus community, Phragmites communis community, Phragmites communis–Carex dispalata community, Ulmus parvifolia community, Zizania latifolia community, Setaria viridis community, and Salix koriyanagi–Salix chaenomeloides community. As a result of PCA analysis, it was classified into seven communities. Seven communities were analyzed, where the most dominant species (M. sacchariflorus, P. communis, C. dispalata, U. parvifolia, Z. latifolia, S. viridis, S. koriyanagi, S. chaenomeloides) of each community were examined as character species. Another species is analyzed as Salix koreensis. Of the sixteen M. sacchariflorus communities, Pterygopleurum neurophyllum was present in six plots (A-2 group) but not in ten plots (A-1 group). These two groups showed differences in coverage and the number of occurring species. As for the relative net contribution degree (r-NCD) in the A-2 group, most species showed low r-NCD except for M. sacchariflorus, which showed an r-NCD of 100. The r-NCDs in the A-1 group were as follows: Miscanthus sacchariflorus (100), P. neurophyllum (21.74), and Persicaria perfoliata (10.14). Therefore, P. neurophyllum is difficult to grow in the A-1 group. As a result, it is thought that the high density of M. sacchariflorus affects the growth and distribution of P. neurophyllum. In order to expand and maintain P. neurophyllum, the habitat environment needs to be altered by adjusting the density of M. sacchariflorus.
1. Introduction
The importance of the ecological and economic value of a country’s biological resources has been increasing since the Convention on Biological Diversity concluded in 1992, and sovereignty over biological resources began to be systematized [1]. However, species extinction continues to increase on Earth as the number of direct and indirect threats to species has increased [2,3]. It has been suggested that various factors such as habitat destruction, environmental pollution, and climate change have endangered plant species [4,5,6,7,8]. Ultimately, changes in the physical environment and climate owing to increased anthropogenic activities place plants at the risk of extinction [9]. These changes can be fatal for plant species with narrow distribution and low adaptability [1,10]. The International Union for Conservation of Nature (IUCN) reported that the number of endangered plants worldwide increased from 16,000 in 2019 to 20,000 in 2020, which is 4000 more species per year [11]. Moreover, the IUCN forecasted that approximately 20% of 250,000 vascular plants growing on the Earth will become extinct within the next 50 years [12,13,14]. Under these circumstances, each country designates, protects, and manages endangered species separate from the Red List [15]. South Korea also legally designates and manages 267 endangered species according to the “Wildlife Protection and Management Act”. Therefore, efforts to prevent this global trend, habitat loss of plant species, and ecological imbalance are necessary.
Pterygopleurum neurophyllum (Maxim.) Kitag (Figure 1) is a perennial plant belonging to the Apiaceae family and is limitedly distributed in the temperate climate regions of Korea, Japan, and China, particularly as one species in one genus in the Korean Peninsula [16]. This species is designated and managed as a Class II Endangered Species and Critically Endangered (CR) Class by the Ministry of Environment in South Korea and as an Endangered (EN) Class in Japan [16]. A few individuals of this species are naturally grown in the Nakdong River backswamp in Yangsan-si, South Gyeongsang-do. It has been reported in Dongjak-dong wetland in Seoul [17], and the specimens have been collected and recorded from wetlands and grasslands in Oryu-dong, Seoul, in 1967; Taereung, Seoul, in 1940; and Cheongnyangni, Seoul, in 1902. It has also been reported to grow naturally in Jeonju in Jeollabuk-do, Jeollanam-do, and Gyeongsangnam-do [16,18,19], but currently it has not been confirmed anywhere else, except in Yangsan, Gyeongsangnam-do.

Figure 1.
Photos of Pterygopleurum neurophyllum and wild habitat in research site.
Very little is known about the ecology of P. neurophyllum. Kwon et al. [20] recently evaluated the germination characteristics of P. neurophyllum seeds [20]. They showed that the seed embryos of P. neurophyllum began to develop after 4 weeks of low-temperature treatment (4 °C) and developed rapidly after 8 weeks. The seeds were germinated after 12 weeks, and the germination rate was 23.1%. When the seeds were treated for 12 weeks with low-temperature treatment and were submerged in 50% sulfuric acid (H2SO4) for 30 min, the germination rate was 29.4%. When the seeds were treated for 12 weeks with low-temperature treatment and GA3 (0.5 g L−1), the germination rate was 37.2%, which was the highest. The germination rate was the highest (79.8%) when both methods were applied. Low-temperature treatment and GA3 treatment after sulfuric acid pre-treatment broke dormancy and improved the germination rate of seeds. Although Son et al. [21] examined the vegetation and soil characteristics of P. neurophyllum in the remaining natural habitat in Yangsan-si, Gyeongsangnam-do, they did not record the distribution of P. neurophyllum [21]. As a result, the detailed characteristics of the native habitats of P. neurophyllum, including the vegetation of the native habitats, cannot be deduced. However, the results of this study showed that willows, common ditch reeds, reeds, and Zizania latifolia were growing in the Wondong marsh where P. neurophyllum was growing naturally. The soil analysis revealed no special findings, except that the sand content was high. It is assumed that, unlike general wetlands, this wetland has high sand content because it is behind the river; hence, the sand is transported to the marsh owing to flooding during the rainy season from the mainstream of the Nakdong River. Meanwhile, some studies have been conducted in Japan as well. Suzuki and Kokufuta examined the endangered plant in Japan and also recorded the habitat status of P. neurophyllum [22]. It was reported that the overall P. neurophyllum population has declined and is on the path of extinction owing to river development in its habitats (Ibaraki, Chiba, Tochigi, Gunma, Saitama, Oita, Kumamoto, and Kagoshima Prefectures). Moreover, Obata et al. and Sato et al. [23,24] included the emergence frequency of endangered species P. neurophyllum as a part of the Watarase Reservoir Restoration Project, which is distributed across Tochigi, Gunma, Saitama, and Ibaraki prefectures in Japan [23,24]. However, only a few studies have reported the vegetation and companion species of P. neurophyllum by investigating its actual natural habitat from plant and ecological perspectives. No study has analyzed the habitat characteristics of native P. neurophyllum in Japan as well as in South Korea.
This study analyzed and evaluated the habitat environment characteristics of P. neurophyllum by focusing on the vegetation for the first time. This study aimed to analyze the flora and vegetation characteristics of the Nakdong River backswamp in Gyeongsangnam-do, the only native habitat of P. neurophyllum, and the distribution and vegetation structure characteristics of natural habitats. The elucidation of vegetation structure characteristics of P. neurophyllum, an endangered species, carries great academic importance, and it is expected that the analysis results can be used as baseline data for establishing restoration and conservation strategies for P. neurophyllum.
2. Materials and Methods
2.1. Study Site Overview
In this study, a flora and vegetation survey was conducted from July to September 2020 in the Wondong wetland in Yangsan-si, Gyeongsangnam-do, known as the native habitat of Pterygopleurum neurophyllum (Figure 2). This wetland was formed by the interactions between natural factors (e.g., the overflow of Nakdong River) and anthropogenic factors (e.g., agriculture). Roads and trails are built north of the study site, and it was affected by artificial disturbance because some areas were being covered by soil. The stream passing through the study site is the Singok-cheon Stream, which originates from Mt. Cheontae-san (Samrangjin-eup, Miryang-si). The Nakdong River flows at the south, and Wondong-cheon Stream, which originates from the valleys of Mt. Gaji-San at Milyang and Unyang, flows at the east. The water system affecting the study site is the Nakdong River on a large scale and Singok-cheon Stream and Wondong-cheon Stream on a small scale. During the rainy season in summer, the river overflowed and the entire study site was flooded. As it is a wetland located downstream of the river, the altitude of the study site is at the sea level, and it had almost no slope. At the study site, the mean annual temperature over the past 10 years was 14.9 °C, and the mean annual precipitation over the past 10 years was 1473 mm (Yangsan-si Meteorological Observatory). These values were approximately 1 °C higher than the mean temperature (13.8 °C) and 75 mm higher than the mean annual precipitation (1398 mm) of South Korea, respectively [25].
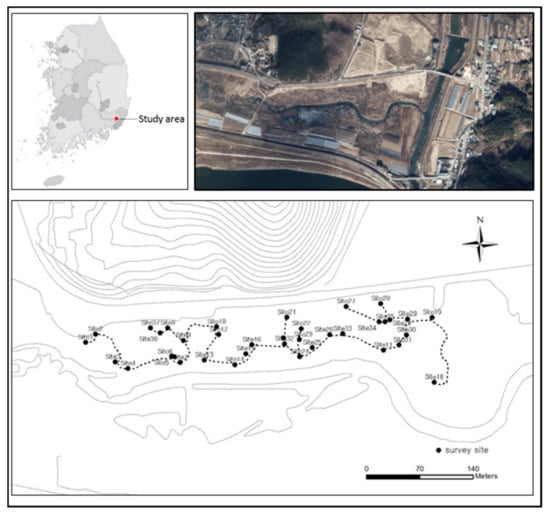
Figure 2.
Survey site of study area.
2.2. Survey and Analysis
For the flora survey, based on the route (Figure 2) traveled during the vegetation survey, plants that appeared and were observed during the survey were recorded. The observed plants were identified and classified according to illustrated books of the flora [18,19,26,27,28]. The arrangement of the botanical list followed the classification system of Cronquist [29], and the scientific and Korean names in this study followed Kim et al.’s study [30]. The endemic species of the Korean Peninsula [31], Floristic Regional Indicator Species [32], the Red List [16], and Ecosystem Disturbance and Exotic Plants [33] were analyzed based on the list of the vascular plants. The life form of the observed plant species was classified (i.e., dormancy, radicoid, disseminule, and growth) and presented by using the method of Raunkiaer and Numata after the modification of Lee’s method for adaptation to plant species in South Korea [19,34,35].
The vegetation survey was conducted at 37 plots in the study site including the area where Pterygopleurum neurophyllum was growing (Figure 2). Vegetation survey was conducted on 6 plots where P. neurophyllum was distributed, and the rest 31 plots were randomly selected. It was conducted according to the phytosociological method of the Zürich-Montpellier School [36]. The size of the quadrat was 4 m2 (2 m × 2 m) for herbaceous communities and 25 m2 (5 m × 5 m) for arborescent communities considering the characteristics of the communities including topography, upper canopy, and height of the plant species. The ground cover class of the observed plant species was measured by using the combined cover degree (9 degrees) (Figure 3) [37].
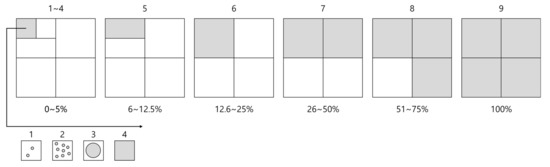
Figure 3.
Criteria for determining the cover degree of plant species [37]. White and gray areas indicate the quadrat areas without and with vegetation cover, while values above and below the squares represent scores (9 degrees) and proportions of the vegetated areas (up to 100%) for each ground cover class, respectively.
Moreover, this study applied the traditional classification method and a classification method transformed by natural logarithm, which is advantageous for mathematical analysis, for the extraction of vegetation units [38]. The relative net contribution degree (r-NCD), a quantitative relative value, was also calculated, which converted the contribution of each observed plant species to the plant community into a percentage for easy comparative analysis [39]. The absolute contribution (NCD) value and the relative contribution value were derived using the following formula:
where ∑Ci, N, ni, and NCDmax indicate the accumulated ground cover of species i in a community, the number of sampling plots for a target plant community, the number of sampling plots in which species i appeared, and the maximum contribution value within a target plant community, respectively. PCA analysis was used to analyze the statistical classification of community and indicator species of the community. For PCA analysis, the PC-ORD (Ver. 5.17) program was used, and vegetation survey data of 37 plots (ground cover of class; 1~9) (Appendix A Table A1) were used.
NCDi = ∑Ci/N × ni/N (Cmin ≤ NCD ≤ Cmax)
r-NCDi = ∑Ci/NCDmax × 100
3. Results and Discussion
3.1. Flora
The flora survey in the natural habitat of Pterygopleurum neurophyllum confirmed 82 taxa: 28 families, 65 genera, 72 species, 2 subspecies, and 8 varieties. Among them, Asteraceae plants appeared the most (11 taxa), followed by Polygonaceae (10 taxa), Cyperaceae (9 taxa), and Poaceae (9 taxa) plants. Pteridophytes and gymnosperms were not found, but 22 taxa of monocotyledons and 60 taxa of dicotyledons were observed (Table 1).

Table 1.
The number of vascular plants in study area.
Two taxa of endangered species were found in the study site (Pterygopleurum neurophyllum and Viola raddeana). One taxon of endemic species in the Korean Peninsula was observed (Salix koriyanagi). P. neurophyllum and V. raddeana are endangered (CR) species listed in the Korean Red List, designated by the Ministry of Environment. The analysis results of floristic regional indicator plants (classifying floras into phytogeographic categories according to the degree of uniqueness in each region) revealed six taxa. One taxon (P. neurophyllum) was observed in Class V and is considered to have high phytogeographical value owing to its limited spatial distribution in South Korea. In addition, one taxon (Veronica peregrina) was observed in Class III, one taxon (V. raddeana) was observed in Class II, and three taxa (Actinostemma lobatum, Salix chaenomeloides, and Ulmus parvifolia.) were observed in Class I. Furthermore, 13 taxa (5 families) of alien plants, including Rumex nipponicus, Amaranthus patulus, Bidens frondosa, and Sonchus oleraceus, were confirmed (Table 2). Two taxa (one family) of invasive alien plants, Ambrosia artemisiifolia and A. trifida, were also identified (Table 2).
Table 2.
The list of vascular plants in study area.
The flora results of the native habitat of Pterygopleurum neurophyllum revealed that the overall rarity or specificity of the ecosystem was not high, as there were no species with a high floristic regional class or there were not a lot of endemic species in the Korean Peninsula. However, the study area was considered to be very important and to have very high protection value because P. neurophyllum and Viola raddeana, which are Class II endangered species that are distributed in a very limited area in South Korea, were found at the study site. In terms of invasive alien plants, species such as Ambrosia artemisiifolia and A. trifida, which have excellent environmental adaptability, can reproduce anywhere, and can form a large community, were observed. Due to the fact that these species can suppress existing ecosystem constituents or alter the ecosystem structure, active measures such as the physical removal of A. artemisiifolia and A. trifida. must be implemented to maintain the existing ecosystem [40].
3.2. Life Form of Observed Plants
The life form of an observed plant is determined to some extent by the climatic, geographical, and ecological factors of the site and the degree of disturbance. Hence, analyzing the growing environment by investigating the life form of the observed plant is an important analysis method for evaluating the environmental and ecological characteristics of study sites [41]. This study analyzed the dormancy form of the 82 taxa that appeared in the study site according to the method of Raunkiaer (Table 3). It was found that most taxa (29 taxa; 35.37%) were annual plants (Th), 12 taxa (14.63%) were hemicryptophytes (H), 10 taxa (12.20%) were Helophytes (HH), and 9 taxa (10.98%) were annual Helophytes (10.98%). It seems that because the study site was a backswamp of a river, which has frequent changes in the growing environment such as soil moisture, soil characteristics (nitrogen and phosphoric acid), and flooding, various annual plants, pioneer species which are advantageous in invasion and settlement [41], hemicryptophytes, and aquatic plants were present during the rainy season. It also seems that a few aquatic plants appeared despite the study site being a wetland because emerged plants such as Miscanthus sacchariflorus and Phragmites communis showed extensive ground cover, and the water quantity was not properly maintained.
Table 3.
Life form spectra of study area.
Among propagation forms, the most common radicoid form of the plants was R5 (55 taxa: 67.07%), which did not make any underground or ground connections. Ten taxa (12.20%) were R3, which had short-branched rhizomes and narrow-range connections. Seven taxa (8.54%) were classified as R2,3, which had both R2, which made lateral rhizome branches and connections in a slightly wider area, and R3, which formed short-branched rhizomes and the narrowest range of connections. Most species observed in the study site did not develop rhizomes, and they were short-branched and formed narrow-range connections even if rhizomes were developed. In terms of disseminule forms, the gravity dispersion form (D4) appeared most frequently (40 taxa: 48.78%), followed by the wind and water dispersion form (D1; 14 taxa: 15.85%), and D1,4 showed the characteristics of D1 and D4 (10 taxa: 12.20%). In terms of growth forms, the erect (e) form was observed most frequently (25 taxa: 30.49%), followed by the thick (t) form (15 taxa: 18.29%), and the pseudorosette (ps) form (7 taxa: 8.54%). The reproduction and growth of plants are closely related to adaptation to the environment. The fact that R5 and R3 were more common than other underground organ types and that gravity dispersion and wind and water dispersion forms appeared at a higher rate may be a result of the fast life cycle providing a competitive advantage in an environment where reproduction and adaptation were difficult [42]. In other words, the dormant form of the study site had a high percentage of annuals and a low percentage of perennials. Among propagation forms, R5 plants without connections were dominant, and the ratio of wind and water dispersion and gravity dispersion forms was high, indicating that there was continuous invasion and disturbance of pioneer species. However, as the perennial hemicryptophytes Miscanthus sacchariflorus and Phragmites communis dominated at a high rate, it is expected that pioneers species cannot easily enter the site as they are outcompeted. Even if pioneer species invade, it is assumed that their stable status would be maintained after being pushed behind in the competition.
The life form of the species found in the study site was Th-R5-D4-e type, which was a different dormancy form from H-R5-D4-e type, a typical type in the estuary of the central region of South Korea [43]. It seems that Th, more favorable to settlement than Hemicryptophyytes (H), appears more than H because the study site is adjacent to the surrounding road; is exposed to an environment that can be easily invaded owing to human access by road and building construction throughout the year; and undergoes alterations and disturbances in the existing growing environment owing to heavy precipitation in the rainy season. The life form of Pterygopleurum neurophyllum is HH-R3(s)-D4-ps. As an aquatic plant with advantages for growing in the wetlands, its rhizome develops in a narrow range, has a succulent type that stores water in the plant body, and has the property of R3(s), which was the only species in the study site possessed that property. It seems that although P. neurophyllum is a wetland plant that grows in humid environments, it can withstand water shortage or an arid environment for a certain period of time. For propagation, a gravity dispersion type refers to the dispersion in which the seeds are dropped by gravity and dispersed by the flow of the surrounding water, followed by reproduction when the water quantity is high. Although it is necessary to conduct studies on P. neurophyllum in various fields, considering the life form characteristics of this study including those of another existing seed study [20], it does not seem that P. neurophyllum is a species that has difficult seed germination conditions or is difficult to grow or propagate. Instead, it is expected that site characteristics such as the disturbance, vegetation, and surrounding environment of the location affect the growth and adaptation of P. neurophyllum.
3.3. Classification of Plant Communities
Vegetation of 37 plots within the study site was classified into seven communities: Miscanthus sacchariflorus community, Phragmites communis community, Phragmites communis–Carex dispalata community, Ulmus parvifolia community, Zizania latifolia community, Setaria viridis community, and Salix koriyanagi–Salix chaenomeloides community. Plant species constituting the classified plant communities were of 58 taxa (Table 4) (Appendix A Table A1).
Table 4.
Synopsis of plant communities in study area by r-NCD value.
The relative contributions of each community, mean ground cover, mean number of observed species, and mean height are presented in Table 4. Plant species of 34 taxa appeared in 16 plots of the Miscanthus sacchariflorus community. M. sacchariflorus appeared with the highest relative contribution (100), followed by Persicaria perfoliata (7.64) and Phragmites communis (6.25) (Table 4). In the P. communis community, eighteen taxa appeared in five plots, and P. communis appeared with the highest relative contribution (100), followed by P. perfoliata (9.77), and Actinostemma lobatum (7.44). In the P. communis–Carex dispalata community, fifteen taxa appeared in three plots, and P. communis showed the highest relative contribution (100), followed by C. dispalata (95.45) and P. perfoliata (18.18). In the Ulmus parvifolia community, fifteen taxa appeared in three plots. There were three layers in the canopy: an arborescent layer, a shrub layer, and an herbaceous layer. U. parvifolia appeared with the highest relative contribution (100), followed by M. sacchariflorus (25) and P. communis (16.67) in the herbaceous layer. Rosa multiflora (16.67) had the highest relative contribution in the shrub layer. In the Zizania latifolia community, eight taxa appeared in three plots, and the number of observed species was the lowest among the examined communities. Z. latifolia showed the highest relative contribution (100), followed by P. communis (16), P. perfoliata (5.33), and Actinostemma lobatum (5.33). In the Setaria viridis community, thirty taxa appeared in three plots. S. viridis appeared with the highest relative contribution (100), followed by Commelina communis, Kummerowia striata, and P. perfoliata (17.39). In the Salix koriyanagi–Salix chaenomeloides community, nineteen taxa appeared in four plots. The vertical structure comprised an arborescent layer, a shrub layer, and an herbaceous layer. S. koriyanagi appeared with the highest relative contribution (100) in the arborescent layer, followed by S. chaenomeloides (96.55) and S. koreensis (31.03) (Table 4).
The PCA analysis results are presented in Figure 4. PCA analysis was performed in order to secure the objectivity of community classification. The PCA analysis results are presented in Figure 4. The results were analyzed by one axis and two axis. Similarly to the community classification analyzed using the traditional classification method, the statistical classification using the PCA analysis was also classified into seven communities (Figure 4). In addition, the character species (M. sacchariflorus, P. communis, C. dispalata, U. parvifolia, Z. latifolia, S. viridis, S. koriyanagi, and S. chaenomeloides) of each seven communities analyzed in the traditional classification method were analyzed as character species contributing to the classification of each community in PCA analysis. Another species is the Salix koreensis, which showed rather high contributions as a result of the PCA analysis. The reason for this is that the Salix koreensis was not distributed in other communities other than the S. koriyanagi–S. chaenomeloides community or showed a very low degree of dominance. Through these results, the significance of a plant phytosociological community classification was secured. It is judged that the physiognomy, which is an important factor in the field survey, has properly selected a survey plot.
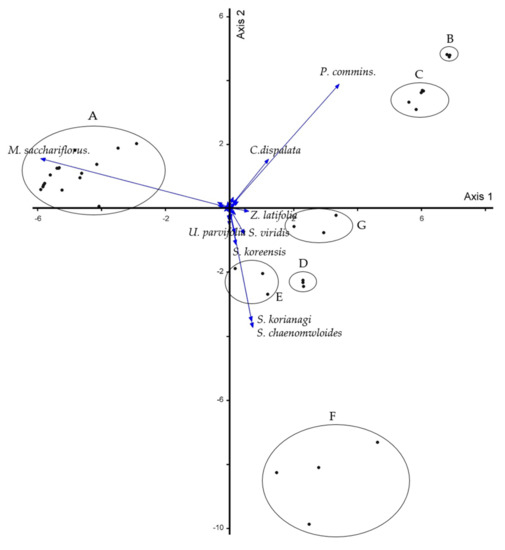
Figure 4.
PCA plot of vegetation of the survey area (A: M. sacchariflorus community; B: P. communis-C. dispalata community; C: P. communis community; D: S. viridis community; E: U. parvifolia community; F: S. koriyanagi-S. chaenomeloides community; G: Z. latifolia community).
The Phragmites communis and communities accounted for the majority of the vegetation in the study site. The Zizania latifolia, Setaria viridis, and Ulmus parvifolia communities dominated at the boundary of the plots. The Salix koriyanagi–Salix chaenomeloides community was distributed partially on a small scale. The Miscanthus sacchariflorus community, which distributed over the widest area, was observed on mounds that were relatively high owing to the sedimentation of sand and debris of M. sacchariflorus in the wetland. The P. communis community was not observed on mounds. It was different from the distribution of the P. communis community and M. sacchariflorus. Moreover, accompanying species hardly appeared in the P. communis and M. sacchariflorus communities due to high dominance. In addition, these two communities are emerged plant communities in the riverside wetland and grow in emerged areas where the water barely flows or is stagnant [44]. It seems that they can create a community easily by forming rhizomes, and they are very advantageous for maintaining the community owing to its strong vitality [44]. The U. parvifolia community showed the characteristic of being distributed at the edge of the wetland without stagnant water but with a humid environment. Although it is a wetland plant, it can grow in a rather humid environment and has strong resistance to wet environments. It is believed that the U. parvifolia community will be maintained continuously unless rapid environmental changes and disturbances such as urban development occur. The Z. latifolia community grows in an environment with shallow water depth throughout the year and forms a colony at the boundary between water and land. Floating leaved plants and emerged plants should be concurrently present with it owing to the site characteristics of the Z. latifolia community, but these species were not observed in this study. The Setaria viridis community can be commonly found in fields and riversides across South Korea. The S. viridis community in the study site was present in a rather arid area away from the wetland, which had the characteristic of temporarily supplying soil moisture during the period of intermittent increased precipitation. Moreover, due to the site of the community, tall plant species were not found and the cover of the community was low. This community had higher biodiversity than other communities because it was easy for annual plants such as Kummerowia striata, Acalypha australis L., and Lysimachia vulgaris var. davurica to invade.
3.4. Distribution Characteristics of Pterygopleurum neurophyllum
Pterygopleurum neurophyllum, an endangered species, did not appear in communities other than the Miscanthus sacchariflorus community (Table 5). P. neurophyllum appeared in only 6 plots out of 16 M. sacchariflorus plots. The habitat of P. neurophyllum had the same site as that of M. sacchariflorus, and it is considered that they share the same habitat. Moreover, the distribution of P. neurophyllum was relatively located at the edge in the M. sacchariflorus communities. Although P. neurophyllum and M. sacchariflorus share the same habitat, P. neurophyllum appeared in locations where the soil was more compacted and dried owing to visitors.
Table 5.
Miscanthus sacchariflorus community vegetation table of study area.
The results showed that the Phragmites communis community where Pterygopleurum neurophyllum was not found was adjacent to the Miscanthus sacchariflorus community. Although P. neurophyllum was not observed in the P. communis community, it is assumed that it could grow in it. This finding seems to be related to the previously mentioned slightly raised mounds and the necromass of M. sacchariflorus and sand sediment. Unlike herbaceous plant communities, woody plant communities such as the Ulmus parvifolia community and the Salix koriyanagi–Salix chaenomeloides community form a canopy. Therefore, the light is blocked, and it is difficult for P. neurophyllum to grow. The Zizania latifolia community grows in shallow water, and the distribution of P. neurophyllum indicates that it is not able to grow in deep water habitats. In the case of the Setaria viridis community, since it grows in a somewhat dry wetland, it is not suitable for the growth of P. neurophyllum.
The vegetation table that summarizes the ground cover class (Classes 1 to 9) using the combined cover degree showed that the cover and ground cover rate of Miscanthus sacchariflorus, the dominant species of the M. sacchariflorus community(A-1) where Pterygopleurum neurophyllum did not appear, were 8–9 and 98.5% (±2.6%), respectively. The cover and ground cover rate of the M. sacchariflorus community(A-2) where P. neurophyllum appeared were 7–8 and 77.5% (±8.2%), respectively, which were relatively low (Table 5). The low ground cover rate of the M. sacchariflorus community where P. neurophyllum appeared provided conditions for the growth and maintenance of P. neurophyllum and increased the appearance probability. P. neurophyllum did not grow in a community with dense M. sacchariflorus, and it seemed that P. neurophyllum was distributed as a refugee in areas with low density of M. sacchariflorus. Moreover, due to this, more sunlight entered into the community, increasing the number of emerged species, and the species compositions of the communities were different as a result. In fact, the number of emerged species in the M. sacchariflorus community without P. neurophyllum(A-1) was 5.7 (±2.45) and that in the M. sacchariflorus community with P. neurophyllum(A-2) was 10 (±1.78), implying that the latter had five more species and different species composition. This suggests that the growth of species in the M. sacchariflorus community was difficult and the composition of the community was simplified.
According to the analysis of the relative contribution of the Miscanthus sacchariflorus community with or without Pterygopleurum neurophyllum, M. sacchariflorus showed the highest relative contribution (100), followed by Persicaria perfoliata (12.13), and Phragmites communis (4.49) in the M. sacchariflorus community without P. neurophyllum. Other emerged species showed a relative contribution of ≤1. By contrast, the relative contribution of M. sacchariflorus (100) was the highest, followed by P. neurophyllum (21.74), P. communis (10.14), and Artemisia princeps (8.70) in the M. sacchariflorus community with P. neurophyllum. In addition, Hemarthria sibirica, Rosa multiflora, and Potentilla anemonefolia showed relative contributions of ≥5. It appeared that it was difficult for other species to grow in the M. sacchariflorus community without P. neurophyllum owing to the particularly high dominance of M. sacchariflorus. Conversely, as the dominance of M. sacchariflorus was relatively lower in the M. sacchariflorus community with P. neurophyllum, other species such as P. neurophyllum and H. sibirica could grow. According to the result of this study, the influence of M. sacchariflorus was very strong, and its dominance was very high. P. neurophyllum was distributed in rather weak M. sacchariflorus community edge. It distributed sporadically in individual units. Overall, the influence of P. neurophyllum was very weak, and it will likely be forced out in the future. Moreover, if the current situation continues, the remaining P. neurophyllum will be simpler, and it will be difficult to maintain P. neurophyllum individuals as the density of M. sacchariflorus increases over time.
3.5. Conservation and Restoration Direction of Pterygopleurum neurophyllum
Conservation and restoration of endangered species’ habitats are very important tasks in the modern society from the viewpoint of improving biodiversity, securing biological sovereignty, and providing ecosystem services. The conservation of endangered species is an essential task in the modern age when living organisms are national property. The current threat to the native habitat of Pterygopleurum neurophyllum is the extremely high dominance of the dominant species. This study conducted vegetation analysis in the Wondong wetland, the native habitat of P. neurophyllum, and found seven vegetation communities. Among them, P. neurophyllum was scattered or distributed at the individual level in the Miscanthus sacchariflorus community. The distribution of P. neurophyllum was determined by the difference in the dominance of the dominant species within the community: The difference between the community with and without P. neurophyllum. First, P. neurophyllum was not found where M. sacchariflorus was highly dominant. Second, owing to the geographical limitations of the Nakdong River backswamp, the site was submerged for several days due to flooding during the rainy season. Therefore, disturbance occurs in the Wondong wetland, which may affect the distribution and sustainability of P. neurophyllum. Third, anthropogenic threats were inherent, such as visitors and human interference due to development. After eliminating and resolving these three threat factors, the direction of conservation and restoration can be established. In order to solve the first problem, the density of M. sacchariflorus must be controlled. It is necessary to remove M. sacchariflorus so that P. neurophyllum can proliferate. This is because P. neurophyllum does not grow in communities with high dominance of M. sacchariflorus. As the second threat is a natural problem, it is difficult to prevent flooding through additional development or other means. Preventing it is not appropriate because it may collapse the entire wetland ecosystem, including changes in the waterways of the Wondong wetland. Instead, ex situ conservation should be promoted propagation and restoration utilizing seeds and individuals of P. neurophyllum. Third, conservation policies should be established by using an active administrative approach. The original habitat of P. neurophyllum should be protected from human disturbance by designating endangered wildlife conservation areas and installing fences and information boards. In summary, for the conservation of the study site, it is necessary to establish physical conservation measures such as habitat restoration by controlling the density of M. sacchariflorus and biological conservation measures such as ex situ conservation through propagation.
4. Conclusions
This study analyzed the flora, life form, and vegetation of the emerged species in the backswamp of the Nakdong River in Yangsan-si (Gyeongsangnam-do) the native habitat of Pterygopleurum neurophyllum, an endangered Class II plant. Vegetation analysis was performed using the phytosociological method of the Zürich-Montpellier School along with a traditional classification method and a mathematical statistical classification method. Regarding the flora of this study site, 82 taxa were identified: 28 families, 65 genera, 72 species, 2 subspecies, and 8 varieties. The life form was of Th-R5-D4-e type, and many annual plants appeared owing to the characteristics of the site. There were seven communities by traditional classification method and PCA anaylsis: Miscanthus sacchariflorus community, Phragmites communis community, Ulmus parvifolia community, Phragmites communis–Carex dispalata community, Zizania latifolia community, Setaria viridis community, and Salix koriyanagi–Salix chaenomeloides community. The character species was analyzed as eight species (M. sacchariflorus, P. communis, C. dispalata, U. parvifolia, Z. latifolia, S. viridis, S. koriyanagi, S. chaenomeloides, and S. koreensis). In particular, M. sacchariflorus showed a high relative contribution (100) and dominated at a very high rate in the M. sacchariflorus community. P. neurophyllum was distributed in a part of the M. sacchariflorus community. It seemed that its distribution was related to the density, dominance, and ground cover rate of M. sacchariflorus. In fact, the dominance and ground cover rate of M. sacchariflorus, the dominant species in the M. sacchariflorus community without P. neurophyllum, were 8–9 and 98.8% (±2.8%), respectively, which were relatively higher than those of M. sacchariflorus, the dominant species in the M. sacchariflorus community with P. neurophyllum, in which the dominance and ground cover rate were 7–8 and 77.5% (±8.2%), respectively. P. neurophyllum appeared in the M. sacchariflorus communities with relatively low dominance of M. sacchariflorus.
In this study, we analyzed vegetation, flora, and vegetation characteristics among many factors in the native habitat of Pterygopleurum neurophyllum. The distribution of species is determined by many factors. As mentioned above, it is necessary to investigate environmental factors such as soil, light quantity, and moisture conditions that may affect the distribution characteristics of P. neurophyllum, in addition to the results of this study. Nevertheless, the results of this study are meaningful in that they reveal the vegetation characteristics of P. neurophyllum. Further studies are warranted by utilizing continuous monitoring.
Author Contributions
Conceptualization, H.J.P. and S.K.; Data curation, H.B.P. and P.B.K.; Formal analysis, S.K. and J.E.H.; Funding acquisition, N.Y.K.; Investigation, C.W.L.; Methodology, J.A.; Supervision, N.Y.K.; Validation, J.A. and B.-D.L.; Visualization, H.B.P.; Writing—original draft, H.J.P.; Writing—review & editing, H.J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by National Institute of Ecology (NIE), grant number NIE-C-2021-46.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
The vegetation table of study area.
Table A1.
The vegetation table of study area.
| A | B | C | D | E | F | G | ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plot no. | 7 | 8 | 12 | 15 | 26 | 29 | 30 | 22 | 10 | 33 | 18 | 19 | 13 | 14 | 5 | 6 | 16 | 20 | 21 | 24 | 27 | 23 | 3 | 32 | 9 | 11 | 34 | 28 | 31 | 11 | 1 | 2 | 37 | 4 | 17 | 31 | 25 | |
| Survey area (m2) | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 5 × 5 | 5 × 5 | 5 × 5 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 2 × 2 | 5 × 5 | 5 × 5 | 5 × 5 | 5 × 5 | |
| Height (m) | 1.5 | 2 | 2 | 2 | 2.5 | 2.3 | 1.8 | 1.6 | 2 | 2 | 1.2 | 2 | 2 | 2 | 2.3 | 2 | 1.8 | 1.8 | 1.8 | 2.5 | 2.5 | 1.8 | 2 | 2 | 5.5 | 6 | 5 | 1.5 | 1.8 | 1.7 | 0.8 | 1 | 1 | 4.5 | 4.5 | 3.5 | 3.5 | |
| Coverage (%) | 95 | 100 | 100 | 100 | 100 | 100 | 95 | 95 | 100 | 100 | 85 | 65 | 80 | 80 | 70 | 85 | 95 | 95 | 95 | 100 | 95 | 95 | 95 | 90 | 75 | 70 | 75 | 90 | 90 | 95 | 65 | 90 | 80 | 70 | 95 | 65 | 65 | |
| Character species of community | ||||||||||||||||||||||||||||||||||||||
| Miscanthus sacchariflorus | H | 8 | 9 | 9 | 9 | 9 | 9 | 9 | 9 | 9 | 9 | 8 | 7 | 8 | 8 | 7 | 8 | 3 | 1 | 2 | 1 | 1 | 1 | 2 | ||||||||||||||
| Phragmites communis | H | 3 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 3 | 8 | 8 | 9 | 9 | 9 | 8 | 8 | 8 | 2 | 1 | 1 | 2 | 1 | 1 | 3 | 1 | ||||||||||||
| Carex dispalata | H | 1 | 7 | 7 | 7 | |||||||||||||||||||||||||||||||||
| Ulmus parvifolia | T2 | 8 | 8 | 8 | ||||||||||||||||||||||||||||||||||
| Zizania latifolia | H | 3 | 2 | 8 | 8 | 9 | ||||||||||||||||||||||||||||||||
| Setaria viridis | H | 7 | 8 | 8 | ||||||||||||||||||||||||||||||||||
| Salix koriyanagi | T2 | 1 | 7 | 8 | 7 | 7 | ||||||||||||||||||||||||||||||||
| Salix chaenomeloides | T2 | 7 | 7 | 7 | 7 | |||||||||||||||||||||||||||||||||
| Companions | ||||||||||||||||||||||||||||||||||||||
| Pterygopleurum neurophyllum | H | 1 | 1 | 1 | 1 | 3 | 3 | |||||||||||||||||||||||||||||||
| Persicaria perfoliata | H | 2 | 1 | 1 | 1 | 1 | 2 | 1 | 2 | 1 | 1 | 1 | 2 | 5 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | ||||||||
| Lespedeza cuneata | H | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||
| Humulus japonicus | H | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| Rosa multiflora | H | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 2 | 2 | 1 | ||||||||||||||||||||||||
| Paederia scandens | H | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||
| Persicaria lapathifolia | H | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 1 | ||||||||||||||||||||||||||
| Actinostemma lobatum | H | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| Persicaria nodosa | H | 1 | 3 | 2 | ||||||||||||||||||||||||||||||||||
| Artemisia princeps | H | 1 | 1 | 2 | 1 | 1 | 2 | |||||||||||||||||||||||||||||||
| Ambrosia trifida | H | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||
| Galium dahuricum var. tokyoense | H | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||
| Carex dimorpholepis | H | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | |||||||||||||||||||||||||||||
| Hemarthria sibirica | H | 2 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||
| Kummerowia striata | H | 1 | 1 | 2 | ||||||||||||||||||||||||||||||||||
| Ambrosia artemisiifolia | H | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||
| Echinochloa crusgalli | H | 1 | 1 | |||||||||||||||||||||||||||||||||||
| Achyranthes japonica | H | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||
| Mosla dianthera | H | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||
| Persicaria maackiana | H | 2 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||
| Rumex nipponicus | H | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||
| Conyza canadensis | H | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||
| Carex thunbergii var. appendiculata | H | 2 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||
| Commelina communis | H | 1 | 1 | 2 | ||||||||||||||||||||||||||||||||||
| Phyllanthus ussuriensis | H | 1 | ||||||||||||||||||||||||||||||||||||
| Potentilla anemonefolia | H | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||
| Acalypha australis | H | 1 | 1 | |||||||||||||||||||||||||||||||||||
| Bidens bipinnata | H | 1 | 1 | |||||||||||||||||||||||||||||||||||
| Amaranthus patulus | H | 1 | ||||||||||||||||||||||||||||||||||||
| Chenopodium album | H | 1 | 1 | |||||||||||||||||||||||||||||||||||
| Salix koreensis | T2 | 1 | 2 | 4 | 2 | 1 | ||||||||||||||||||||||||||||||||
| Carex neurocarpa | H | 1 | ||||||||||||||||||||||||||||||||||||
| Bromus japonicus | H | 1 | ||||||||||||||||||||||||||||||||||||
| Agropyron tsukushiense var. transiens | H | 1 | 1 | |||||||||||||||||||||||||||||||||||
| Conyza sumatrensis | H | 1 | ||||||||||||||||||||||||||||||||||||
| Amorpha fruticosa | S | 1 | ||||||||||||||||||||||||||||||||||||
| Viola lactiflora | H | 1 | ||||||||||||||||||||||||||||||||||||
| Lysimachia vulgaris var. davurica | H | 1 | ||||||||||||||||||||||||||||||||||||
| Acer tataricum subsp. ginnala | H | 1 | 1 | |||||||||||||||||||||||||||||||||||
| Glycine soja | H | 1 | ||||||||||||||||||||||||||||||||||||
| Persicaria hydropiper | H | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||
| Galium spurium var. echinospermum | H | 1 | ||||||||||||||||||||||||||||||||||||
| Scirpus radicans | H | 1 | 2 | 1 | ||||||||||||||||||||||||||||||||||
| Rubus parvifolius | H | 1 | ||||||||||||||||||||||||||||||||||||
| Persicaria sagittata | H | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||
| Eclipta prostrata | H | 1 | ||||||||||||||||||||||||||||||||||||
| Juncus effusus var. decipiens | H | 1 | ||||||||||||||||||||||||||||||||||||
| Rorippa palustris | H | 1 | ||||||||||||||||||||||||||||||||||||
| Xanthium canadense | H | 1 | ||||||||||||||||||||||||||||||||||||
| Dioscorea batatas | H | 1 | ||||||||||||||||||||||||||||||||||||
| Cyperus microiria | H | |||||||||||||||||||||||||||||||||||||
| Cosmos bipinnatus | H | 1 | ||||||||||||||||||||||||||||||||||||
A: Miscanthus sacchariflorus community; B: Phragmites communis community; C: Phragmites communis-Carex dispalata community; D: Ulmus parvifolia community; E: Zizania latifolia community; F: Setaria viridis community; G: Salix koriyanagi-Salix chaenomeloides community. T2: subtree; S: shrub; H: herb.
References
- Yoon, J.W.; Yi, M.H.; Kim, Y.S. Growth environment and vegetation structure of native habitats of Wikstroemia ganpi (Sieb. Et Zucc.) maxim. Korean J. Environ. Ecol. 2014, 28, 331–341. (In Korean) [Google Scholar] [CrossRef]
- Sodhi, N.S.; Enrlich, P.R. Conservation Biology for All; Oxford University Press: Oxford, UK, 2010; 341p. [Google Scholar]
- Lim, B.S.; Kim, D.K.; Kim, A.R.; Seal, J.W.; Lee, C.S. Analysis of Ecodiversity as the Foundation for Conserving Biodiversity and Its Restoration Strategy. Korean Soc. Limnol. 2020, 53, 408–426. (In Korean) [Google Scholar]
- Schemske, D.W.; Husband, B.C.; Ruckelshaus, M.H.; Goodwillie, C.; Parker, I.M.; Bishop, J.G. Evaluating approached to the conservation of rare and endangered plants. Ecology 1994, 75, 584–606. [Google Scholar] [CrossRef] [Green Version]
- Keith, D.A. An evaluation and modification of World Conservation Union Red List criteria for classification of extinction risk in vascular plants. Conserv. Biol. 1998, 12, 1076–1090. [Google Scholar] [CrossRef]
- Naeem, S.; Knops, J.M.H.; Tilman, D.; Haddad, N.M.; Mitchell, C.E.; Haarstad, J.; Ritchie, M.E.; Howe, K.M.; Reich, P.B.; Siemann, E. Effects of plant species richness on invasion dynamics, disease outbreaks, insect abundances and diversity. Ecol. Lett. 1999, 2, 286–293. [Google Scholar]
- Mace, G.M.; Collar, N.J.; Gaston, K.J.; Hilton-Taylor, C.; Akçakaya, H.R.; Leader-Williams, N.; Milner-Gulland, E.J.; Stuart, S.N. Quantification of extinction risk: IUCN’s system for classifying threatened species. Conserv. Biol. 2008, 22, 1424–1442. [Google Scholar] [CrossRef] [PubMed]
- Primack, R.B. A Primer of Conservation Biology, 3rd ed.; Sinaurer Associates: Sunderland, MA, USA, 2008; p. 292. [Google Scholar]
- Lee, S.H.; Yeon, M.H.; Shim, J.K. Habitat and Distribution Feature of Endangered Species Leontice microrhyncha S. Moore. Korea J. Environ. Ecol. 2011, 25, 819–2011. (In Korean) [Google Scholar]
- Hampe, A.; Petit, R.J. Conservation biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [Green Version]
- IUCN. Summary Statistics of the IUCN Red List of Threatened Species from 1996 to 2020. Available online: http://www.iucnredlist.org (accessed on 25 March 2020).
- Wilson, E.O. The Diversity of Life; The Belknap Press of Harvard University Press: Cambridge, MA, USA, 1992; p. 424. [Google Scholar]
- Falk, D.A.; Olwell, P. Scientific and policy conservations on reintroduction of endangered species. Rhodora 1992, 94, 287–315. [Google Scholar]
- Barnes, B. Landscape Ecosystem approach and conservation od endangered spaces. End. Species Update 1992, 10, 13–19. [Google Scholar]
- Lucas, G.; Synge, H. The IUCN Plant Red Data Book; IUCN: Morges, Switzerland, 1978. [Google Scholar]
- National Institute of Biological Resources. Red Data Book of Endangered Vascular Plants in Korea; National Institute of Biological Resources, Ministry of Environment: Incheon, Korea, 2012; p. 167. (In Korean)
- Rim, K.H.; Chung, B.S. Pharmacognostical study on the Pterygopleurum neurophyllum Kitagawa. J. Pharm. Soc. Korea 1966, 10, 1–4. (In Korean) [Google Scholar]
- Lee, W.T. Lineamenta Flora Korea; Academy Press: Seoul, Korea, 1996; p. 689. (In Korean) [Google Scholar]
- Lee, W.T. Standard Illustrations of Korean Plants; Academy Press: Seoul, Korea, 1996; p. 624. (In Korean) [Google Scholar]
- Kwon, H.J.; Kim, Y.R.; Kim, S.Y. First report on the germination requirement of Pterygopleurum neurophyllum seeds. Seed Sci. Technol. 2020, 48, 41–48. [Google Scholar] [CrossRef]
- Son, S.G.; Lee, P.H.; Kim, C.S.; Oh, K.H. Vegetation Structure and Sedimnet Properties of Wondong Wetland in Nakdong-gang. J. Wetl. Res. 2002, 4, 21–31. (In Korean) [Google Scholar]
- Suzuki, M.; Kokufuta, S. Endangered vascular plants in Ibaraki Prefecture, Japan. Bull. Inst. Educ. Fac. Educ. Ibaraki Univ. 1994, 26, 63–72. (In Japanese) [Google Scholar]
- Obata, T.; Ishii, J.; Kadoya, T.; Washitani, I. Effect of past topsoil removal on the current distribution of threatened plant species in a moist tall grassland of the Watarase wetland, Japan: Mapping of selected sites for wetland restoration by topsoil removal. Jpn. J. Conserv. Ecol. 2012, 17, 221–233. (In Japanese) [Google Scholar]
- Sato, R.; Saai, J.; Kodama, Y.; Ishii, M. Wetland conservation and restoration methods in the Watarase Flood Control Basin. Rep. Riverfront Res. Inst. 2009, 20, 95–102. (In Japanese) [Google Scholar]
- Korea Meteorological Administration. Climate Information Portal Home Page. Available online: http://www.climate.go.kr (accessed on 1 July 2021).
- Lee, T.B. Illustrated Flora of Korea; Hyangmun Publ. Co., Ltd.: Seoul, Korea, 1980; p. 990. (In Korean) [Google Scholar]
- Lee, T.B. Coloured Flora of Korea; Hyangmun Publ. Co., Ltd.: Seoul, Korea, 2003; Volumes I and II, p. 1928. (In Korean) [Google Scholar]
- Lee, Y.N. New Flora of Korea; Gyohak Publ. Co., Ltd.: Seoul, Korea, 2006; Volumes I and II, 1859p. (In Korean) [Google Scholar]
- Cronquist, A. An Integrated System of Classification of Flowering Plants; Columbia University Press: New York, NY, USA, 1981; 1262p. [Google Scholar]
- Kim, J.S.; Jang, H.D.; Kim, J.H.; Lee, B.Y. Division Pteridophyta, Division Pinophyta, Division Magnoliophyta. National Species List of Korea I: Plants, Fungi, Algae, Prokaryotes; National Institute of Biological Resources, Ed.; Designzip Publ. Co., Ltd.: Seoul, Korea, 2019; p. 188. [Google Scholar]
- Chung, G.Y.; Chang, K.S.; Chung, J.M.; Choi, H.J.; Paik, W.K.; Hyun, J.O. A checklist of endemic plants on the Korean peninsula. Korean J. Pl. Taxon. 2017, 47, 264–288. (In Korean) [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H.; Moon, M.O.; Ahn, J.K.; Hwang, I.C.; Lee, S.H.; Choi, S.S.; Lee, J.H.; Bum, H.M.; Kim, C.G.; Cha, J.Y. Floristic Target Species in Korea; National Institute of Ecology: Seocheon, Korea, 2018; 788p. (In Korean) [Google Scholar]
- National Institute of Ecology Home Page. Available online: http://kias.nie.re.kr (accessed on 1 July 2021).
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Oxford University Press: Oxford, UK, 1934; p. 632. [Google Scholar]
- Numata, M. Illustrated Plant Ecology; Ashakura Book Co.: Tokyo, Japan, 1970; pp. 33–43. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoniologie der Vegetationskunde, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 1964; p. 631. [Google Scholar]
- Westhoff, V.; Van Der Maarel, E. The braun-blanquet approach. In Classification of Plant Communities; Springer: Berlin/Heidelberg, Germany, 1978; pp. 287–399. [Google Scholar]
- Becking, R.W. The Zürich-Montpellier school of phytosociology. Bot. Rev. 1957, 23, 411–488. [Google Scholar] [CrossRef]
- Kim, J.W.; Manyko, Y.I. Syntaxonomical and synchorological characteristics of the cool-temperate mixed forest in the southern Sikhote Alin, Russian Far East. Korean J. Ecol. 1994, 17, 391–413. [Google Scholar]
- You, J.H.; Mun, S.J.; Leem, W.S. Management Plan and Vascular Plants of the Hwarang District in Gueongju National Park. J. Korean Environ. Res Technol. 2011, 14, 17–35. (In Korean) [Google Scholar]
- Kim, J.M. Plant Ecology of Korea; Hyundai Science Sinsu Publ. Co., Ltd.: Seoul, Korea, 1984; p. 226. (In Korean) [Google Scholar]
- Choi, Y.E.; Kim, C.H. A Study on the Plant Life Forms of Uninhabited Islands in Buan-gun and Gochang-gun of Jeollabukdo. J. Korean Isl. 2017, 29, 245–261. (In Korean) [Google Scholar] [CrossRef]
- Lee, W.T.; Yim, Y.J. Plant Geograpy; Kangwon National University Press: ChunCheon, Korea, 2002; p. 412. (In Korean) [Google Scholar]
- Kim, J.G. Wetland vegetation in Korea. Nat. Conserv. 2007, 138, 35–56. (In Korean) [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).