
Effect of Saline Water for Drip Irrigation on Microbial Diversity and on Fertility of Aeolian Sandy Soils

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sampling
2.2. Sampling
2.3. Determination Methods
2.3.1. Diversity of the Soil Bacterial Community
2.3.2. Physical and Chemical Analysis
2.3.3. Soil Bacterial Mass
2.3.4. Soil Enzyme Activity
2.4. Data Analysis
2.5. Evaluation of Soil Fertility
3. Results and Analysis
3.1. Soil Bacterial Diversity
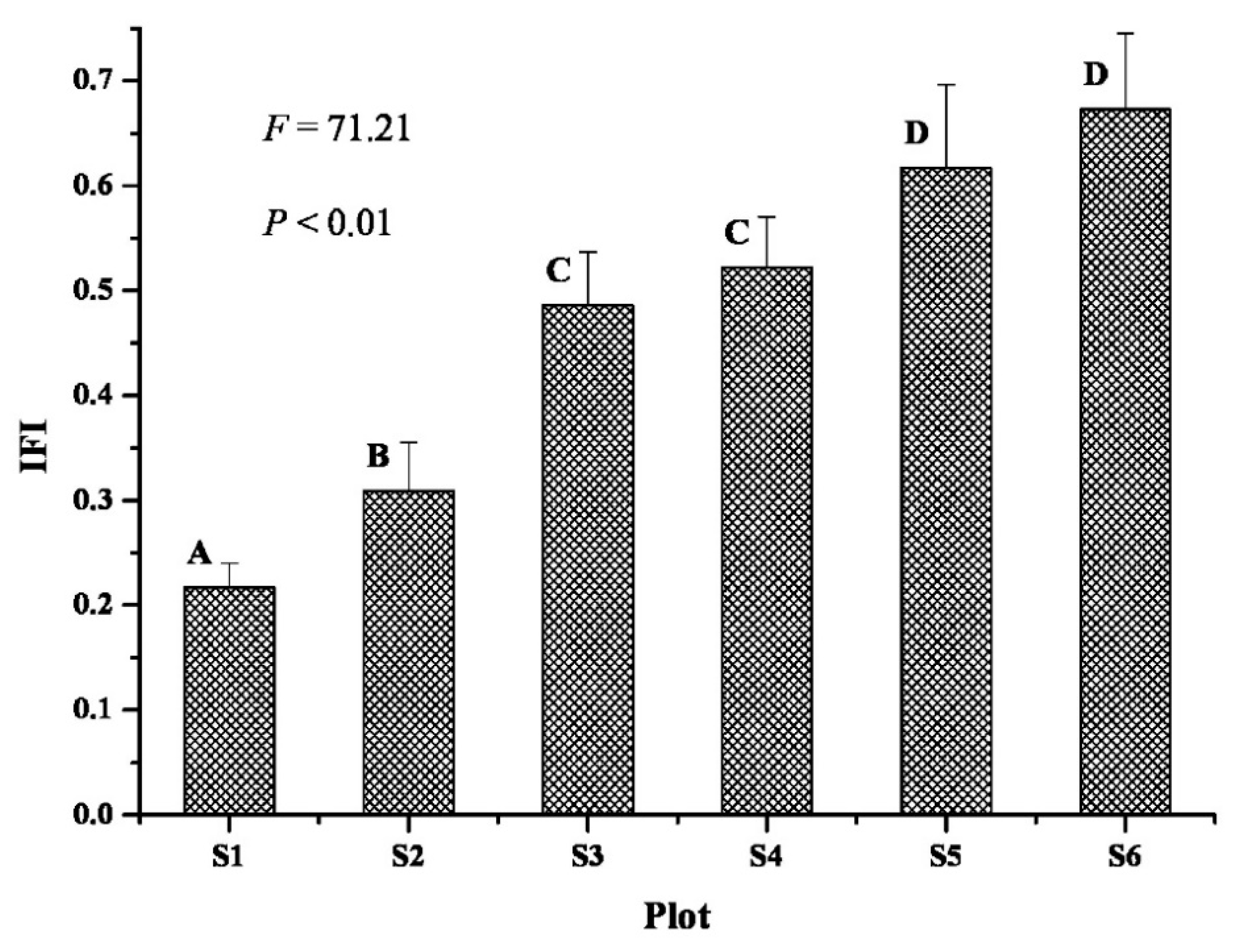
3.2. Soil Fertility
3.3. Relationship between Soil Bacterial Diversity and Soil Fertility
3.4. Composition Characters of Soil Salinity
4. Discussion and Conclusions
4.1. Effect of Salinity of Irrigation Water on Soil Bacterial Diversity
4.2. Changes in Soil Fertility under Drip Irrigation with Saline Water
4.3. The Relationship between Soil Microorganisms and Soil Fertility
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Breure, A.M.; Mulder, C.; Römbke, J.; Ruf, A. Ecological classification and assessment concepts in soil protection. Ecotoxicol. Environ. Saf. 2005, 62, 211–229. [Google Scholar] [CrossRef]
- Doran, J.W.; Coleman, D.C.; Bezdicek, D.F. Defining Soil Quality for a Sustainable Environment; SSSA Special Publication: Madison, WI, USA, 1994. [Google Scholar]
- Sun, B.; Zhou, S.; Zhao, Q. Evaluation of spatial and temporal changes of soil quality based on geostatistical analysis in the hill region of subtropical China. Geoderma 2003, 115, 85–99. [Google Scholar] [CrossRef]
- Harris, J.A. Measurements of the soil microbial community for estimating the success of restoration. Eur. J. Soil Sci. 2003, 54, 801–808. [Google Scholar] [CrossRef]
- Bonanomi, G.; D’Ascoli, R.; Antignani, V.; Capodilupo, M.; Cozzolino, L.; Marzaioli, R.; Puopolo, G.; Rutigliano, F.A.; Scelza, R.; Scotti, R.; et al. Assessing soil quality under intensive cultivation and tree orchards in Southern Italy. Appl. Soil Ecol. 2011, 47, 184–194. [Google Scholar] [CrossRef]
- Liu, Z.F.; Fu, B.J.; Liu, G.H.; Zhu, Y.G. Soil quality: Concept, indicators and its assessment. Acta Ecol. Sin. 2006, 26, 901–910. [Google Scholar]
- Schaeffer, S.M.; Billings, S.A.; Evans, R.D. Responses of soil nitrogen dynamics in a Mojave Desert ecosystem to manipulations in soil carbon and nitrogen availability. Oecologia 2003, 134, 547–553. [Google Scholar] [CrossRef]
- Jin, Z.; Lei, J.; Xu, X.; Li, S.; Fan, J.; Zhao, S. Characteristics of the soil microbial population in forest land irrigated with saline water in the desert area. J. Arid. Land 2010, 2, 107–115. [Google Scholar] [CrossRef]
- Yuqin, S.; Ji, Z. Vertical distributions in number of soil microorganism in stabilized sand dune with Artemisia ordosica in the Hobq of Inter-mongol. Acta Scient. Natur. Univer. Neim. 2000, 31, 198–200. [Google Scholar]
- Wang, Z.; Peng, R.Y.; Wang, L.; Liu, L.Y. Studies on soil properties of aeolian sandy land improvement and utilization in south edge of Musu desert. J. Soil Water Conserv. 2006, 20, 14–17. [Google Scholar]
- Cao, C.Y.; Jiang, D.M.; Quan, G.J.; Geng, L.; Cui, Z.B.; Luo, Y.M. Soil physical and chemical characters changes of Caragana mi-crophylla plantation for sand fixation in Keerqin sandy land. J. Soil Water Conserv. 2004, 18, 108–112. [Google Scholar]
- Guo, K.; Dong, X.J.; Liu, Z.M. Characteristics of soil moisture content on sand dunes in Mu Us sandy grassland: Why Artemisia ordosica declines old fixed sand dunes. Acta Phyt. Sin. 2000, 24, 275–279. [Google Scholar]
- Liu, J.-J.; Fang, S.-Z.; Xie, B.-D.; Hao, J.-J. Effects of bio-mulching on rhizosphere soil microbial population, enzyme activity and tree growth in poplar plantation. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2008, 19, 1204–1210. [Google Scholar]
- Calbrix, R.; Laval, K.; Barray, S. Analysis of the potential functional diversity of the bacterial community in soil: A reproducible procedure using sole-carbon-source utilization profiles. Eur. J. Soil Biol. 2005, 41, 11–20. [Google Scholar] [CrossRef]
- Duan, X.-J.; Min, H. Diversity of microbial genes in paddy soil stressed by cadmium using DGGE. Huan Jing Ke Xue Huanjing Kexue 2004, 25, 122–126. [Google Scholar]
- Pennanen, T.; Paavolainen, L.; Hantula, J. Rapid PCR-based method for the direct analysis of fungal communities in complex environmental samples. Soil Biol. Biochem. 2001, 33, 697–699. [Google Scholar] [CrossRef]
- Larkin, R.P.; Honeycutt, C.W.; Griffin, T.S. Effect of swine and dairy manure amendments on microbial communities in three soils as influenced by environmental conditions. Biol. Fert. Soils 2006, 43, 51–61. [Google Scholar] [CrossRef]
- Institute of Soil Science; Chinese Academy of Science. Soil Physical and Chemical Properties Analysis; Science and Technology Press in Shanghai: Shanghai, China, 1978. [Google Scholar]
- Wu, J.S.; Lin, Q.M.; Huang, Q.Y. Soil Microbial Biomass of Methods and Application; Meteorological Press: Beijing, China, 2006; pp. 55–88. [Google Scholar]
- Guang, S.Y. Soil Enzyme and Its Methodology; Agriculture Press: Beijing, China, 1986. [Google Scholar]
- Zhang, D.H.; Ye, Z.F. The effects of rotating plantation on the soil fertility of forest and the growth of stand. Scinen. Silv. Sin. 2001, 37, 10–15. [Google Scholar]
- Jin, Z.; Lei, J.; Xu, X.; Li, S.; Zhang, Z.; Peng, H.; Zhong, X. Effect of Drip Irrigation Water with Different Salinity on Soil Microbe of the Tarim Desert Highway Shelter-Forest. In Proceedings of the 2nd International Conference on Remote Sensing, Environment and Transportation Engineering, Nanjing, China, 1–3 June 2012; pp. 1–6. [Google Scholar] [CrossRef]
- Pang, H.C.; Li, Y.Y.; Yu, T.Y.; Liu, G.J.; Dong, L.H. Effects of microorganism agent on soil salination and alfalfa growth under different salt stress. Plant Nutr. Fertil. Sci. 2011, 17, 1403–1408. [Google Scholar]
- Kirk, J.L.; Beaudette, L.A.; Hart, M.; Moutoglis, P.; Klironomos, J.N.; Lee, H.; Trevors, J.T. Methods of studying soil microbial diversity. J. Microbiol. Methods 2004, 58, 169–188. [Google Scholar] [CrossRef]
- Yu, H.Y.; Li, T.X. Evolution of salt accumulation greenhouse soil in Liaoning province. J. Soil Water Conserv. 2005, 19, 80–83. [Google Scholar]
- Zhang, J.G.; Xu, X.W.; Li, S.Y.; Zhao, Y.; Zhang, A.F.; Zhang, T.B.; Jiang, R. Is the Taklimakan Desert Highway shelterbelt sus-tainable to long-term drip irrigation with high saline groundwater? PLoS ONE 2016, 11, 1–16. [Google Scholar]
- Wall, D.H.; Virginia, R.A. Controls on soil biodiversity: Insights from extreme environments. Appl. Soil Ecol. 1999, 13, 137–150. [Google Scholar] [CrossRef]
- Lei, J.; Li, S.; Jin, Z.; Fan, J.; Wang, H.; Fan, D.; Zhou, H.; Gu, F.; Qiu, Y.; Xu, B. Comprehensive eco-environmental effects of the shelter-forest ecological engineering along the Tarim Desert Highway. Sci. Bull. 2008, 53, 190–202. [Google Scholar] [CrossRef] [Green Version]
- Gu, F.X.; Pan, X.L.; Pan, B.R.; Wen, Q.K. Changing of eolian soil fertility in central Taklimakan Desert under impact of artificial vegetation. Ecol. Sin. 2002, 22, 1179–1188. [Google Scholar]
- Zhang, H.B.; Duan, C.Q.; Qu, L.H. Culture independent methods for studies on microbial ecology of soils. Chin. J. Ecol. 2003, 22, 131–136. [Google Scholar]
- Wang, Y.; Han, B.; Shi, Z.Q.; Shao, G.Q.; Jiang, X.D.; Ning, T.Y.; Jiao, N.Y.; Li, Z.J. Effects of conservation tillage on soil microbial characters and soil enzyme activities. J. Soil Water Conserv. 2006, 20, 120–122. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot No. | Salinity Value (g/L−1) | Location (Miles along the Desert Highway from Minfeng County) |
|---|---|---|
| S1 | 23.80 ± 1.12 | 140 km + 470 m |
| S2 | 18.10 ± 0.84 | 168 km + 800 m |
| S3 | 13.99 ± 0.66 | 176 km + 000 m |
| S4 | 8.90 ± 0.45 | 184 km + 500 m |
| S5 | 5.75 ± 0.29 | 277 km + 500 m |
| S6 | 2.58 ± 0.13 | 318 km + 100 m |
| Fertility Factor | Community | Fertility Factor | Community |
|---|---|---|---|
| Organic matter | 0.036 | DNA segments | 0.033 |
| Total nitrogen | 0.043 | Carbon source metabolism | 0.039 |
| Total phosphorus | 0.041 | Species of fatty acid | 0.038 |
| Total potassium | 0.037 | Catalase | 0.039 |
| Available nitrogen | 0.044 | Phosphatase | 0.040 |
| Available phosphorus | 0.041 | Urease | 0.043 |
| Available potassium | 0.041 | Cellulose | 0.033 |
| pH | 0.049 | Urease | 0.039 |
| Electrical conductivity | 0.034 | Protease | 0.036 |
| Total salt | 0.043 | Bacterial biomass carbon | 0.044 |
| Bulk density | 0.041 | Bacterial biomass nitrogen | 0.039 |
| Median diameter of soil particles | 0.040 | Bacterial biomass phosphorus | 0.041 |
| Moisture content | 0.047 |
| Plot | Carbon Mechanism | Genes | Fatty Acids | ||||||
|---|---|---|---|---|---|---|---|---|---|
| H | S | EH | H | S | EH | H | S | EH | |
| S1(23.80) | 2.85 ± 0.23 a | 21.75 ± 2.84 a | 0.93 ± 0.12 a | 0.82 ± 0.07 A | 12.34 ± 1.61 A | 0.25 ± 0.02 a | 1.30 ± 0.35 a | 20.08 ± 1.83 a | 0.33 ± 0.05 a |
| S2(18.10) | 3.14 ± 0.47 b | 25.75 ± 3.16 b | 0.97 ± 0.14 a | 0.87 ± 0.06 A | 12.96 ± 1.80 A | 0.29 ± 0.04 a | 1.47 ± 0.38 a | 21.69 ± 2.48 a | 0.41 ± 0.08 ab |
| S3(13.99) | 3.20 ± 0.35 b | 27.13 ± 3.60 c | 0.97 ± 0.17 a | 1.00 ± 0.06 A | 14.25 ± 2.57 A | 0.39 ± 0.04 b | 1.51 ± 0.33 ab | 23.5 ± 5.07 ab | 0.39 ± 0.09 a |
| S4(8.90) | 3.24 ± 0.39 b | 27.88 ± 2.82 c | 0.97 ± 0.18 a | 1.34 ± 0.32 B | 20.25 ± 3.18 B | 0.50 ± 0.11 c | 1.65 ± 0.60 b | 25.75 ± 2.56 b | 0.55 ± 0.15 bc |
| S5(5.75) | 3.26 ± 0.44 b | 28.63 ± 4.58 c | 0.97 ± 0.19 a | 1.78 ± 0.54 C | 35.00 ± 5.34 C | 0.48 ± 0.10 c | 1.73 ± 0.75 bc | 30.01 ± 4.91 c | 0.58 ± 0.11 c |
| S6(2.58) | 3.59 ± 0.45 c | 29.13 ± 4.22 c | 1.06 ± 0.21 a | 2.43 ± 0.47 D | 41.75 ± 5.9 D | 0.63 ± 0.13 d | 1.86 ± 0.92 c | 32 ± 8.42 c | 0.57 ± 0.14 c |
| p value | * | * | ** | ** | |||||
| Bands | Closest Species in the GenBank Database | Sequence Accession Number | Similarity (%) |
|---|---|---|---|
| C1 | Janthinobacterium sp. | AY88012 | 100 |
| C2 | Acidovorax sp. | AY88013 | 99 |
| C3 | Anaeromyxobacter | AY88014 | 96 |
| C4 | Thiobacillus denitrificans | AY88015 | 100 |
| C5 | Shewanella frigidimarina | AY88016 | 97 |
| Plot | CO32− | HCO3− | Cl− | SO42− | Ca2+ | Mg2+ | Na+ | K+ |
|---|---|---|---|---|---|---|---|---|
| S1 | 0.09 | 0.31 | 6.36 | 6.04 | 2.10 | 0.58 | 3.47 | 0.32 |
| S2 | 0.07 | 0.28 | 6.04 | 5.75 | 1.86 | 0.47 | 3.29 | 0.27 |
| S3 | 0.01 | 0.12 | 4.46 | 4.88 | 1.52 | 0.26 | 2.97 | 0.12 |
| S4 | 0.01 | 0.12 | 1.69 | 3.33 | 0.87 | 0.25 | 1.18 | 0.14 |
| S5 | 0.03 | 0.17 | 1.31 | 0.71 | 0.14 | 0.07 | 0.92 | 0.11 |
| S6 | 0.02 | 0.13 | 1.09 | 0.86 | 0.19 | 0.06 | 0.80 | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Jin, Z.; Wang, H. Effect of Saline Water for Drip Irrigation on Microbial Diversity and on Fertility of Aeolian Sandy Soils. Diversity 2021, 13, 379. https://doi.org/10.3390/d13080379
Yu X, Jin Z, Wang H. Effect of Saline Water for Drip Irrigation on Microbial Diversity and on Fertility of Aeolian Sandy Soils. Diversity. 2021; 13(8):379. https://doi.org/10.3390/d13080379
Chicago/Turabian StyleYu, Xiangxiang, Zhengzhong Jin, and Haifeng Wang. 2021. "Effect of Saline Water for Drip Irrigation on Microbial Diversity and on Fertility of Aeolian Sandy Soils" Diversity 13, no. 8: 379. https://doi.org/10.3390/d13080379
APA StyleYu, X., Jin, Z., & Wang, H. (2021). Effect of Saline Water for Drip Irrigation on Microbial Diversity and on Fertility of Aeolian Sandy Soils. Diversity, 13(8), 379. https://doi.org/10.3390/d13080379