
Quantifying Southern Pacific Rattlesnake (Crotalus oreganus helleri) Hunting Behavior through Community Science


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
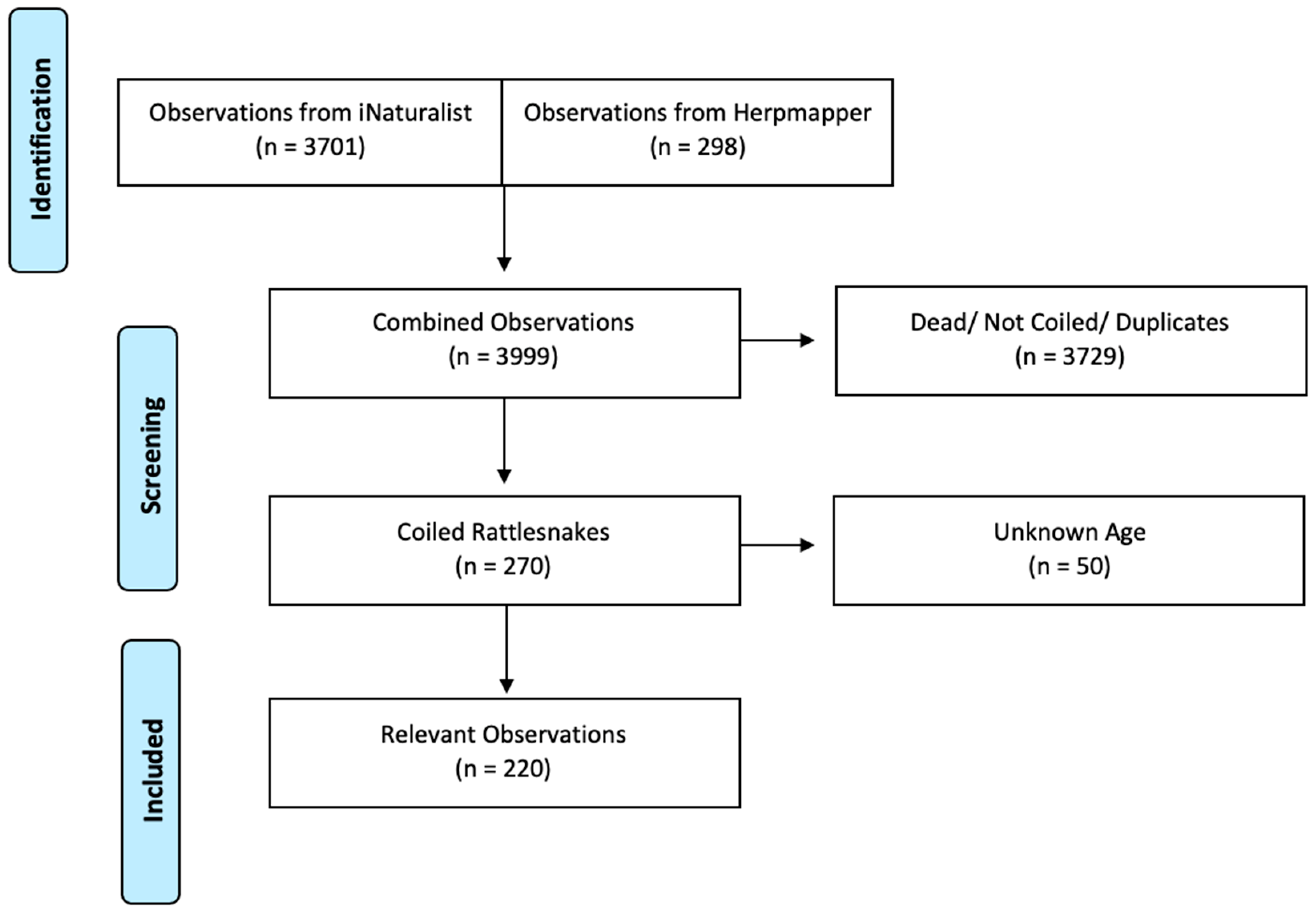
2.1. Identification of Community Science Data
2.2. Data Extraction
2.3. Statistical Analyses
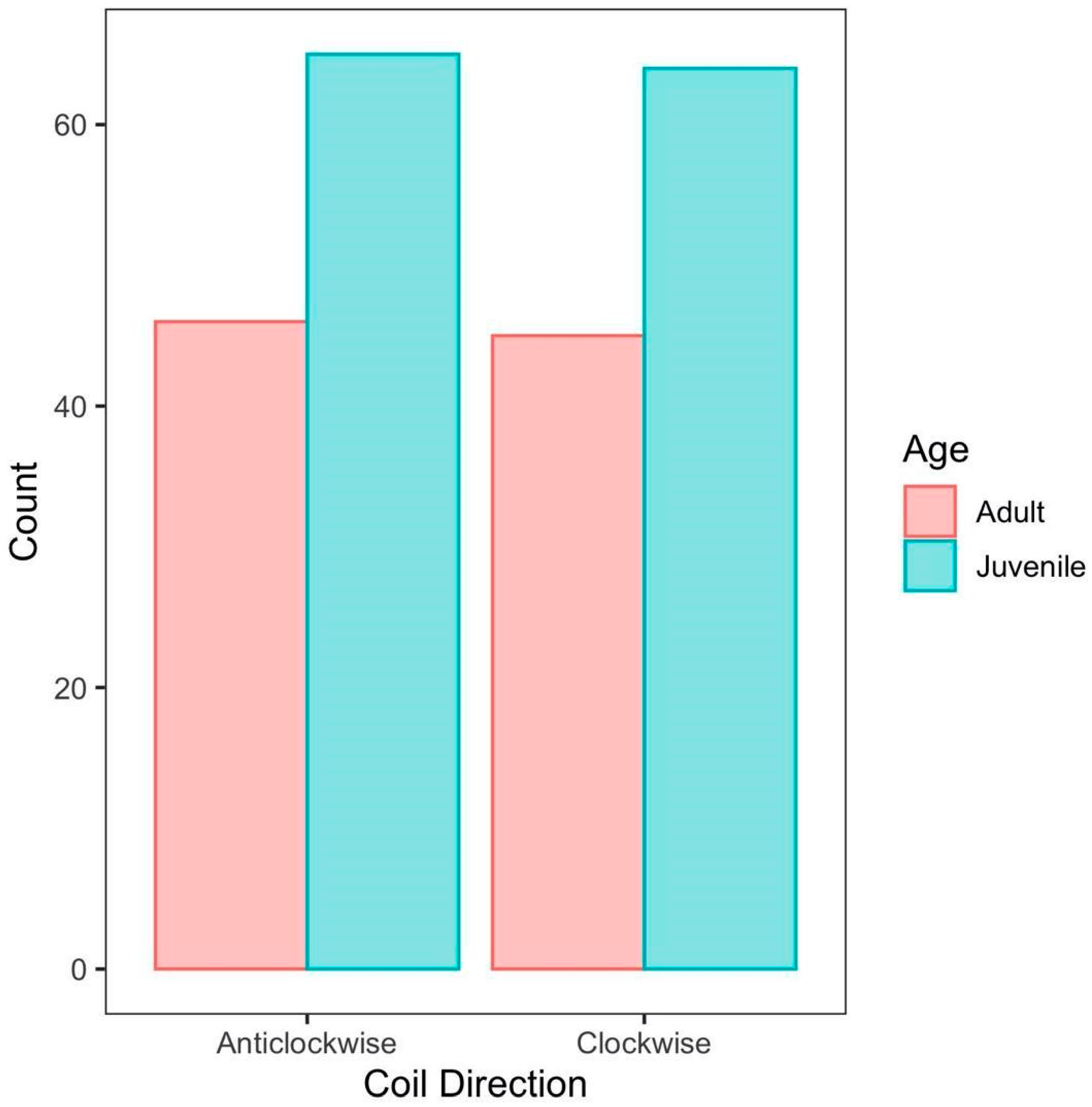
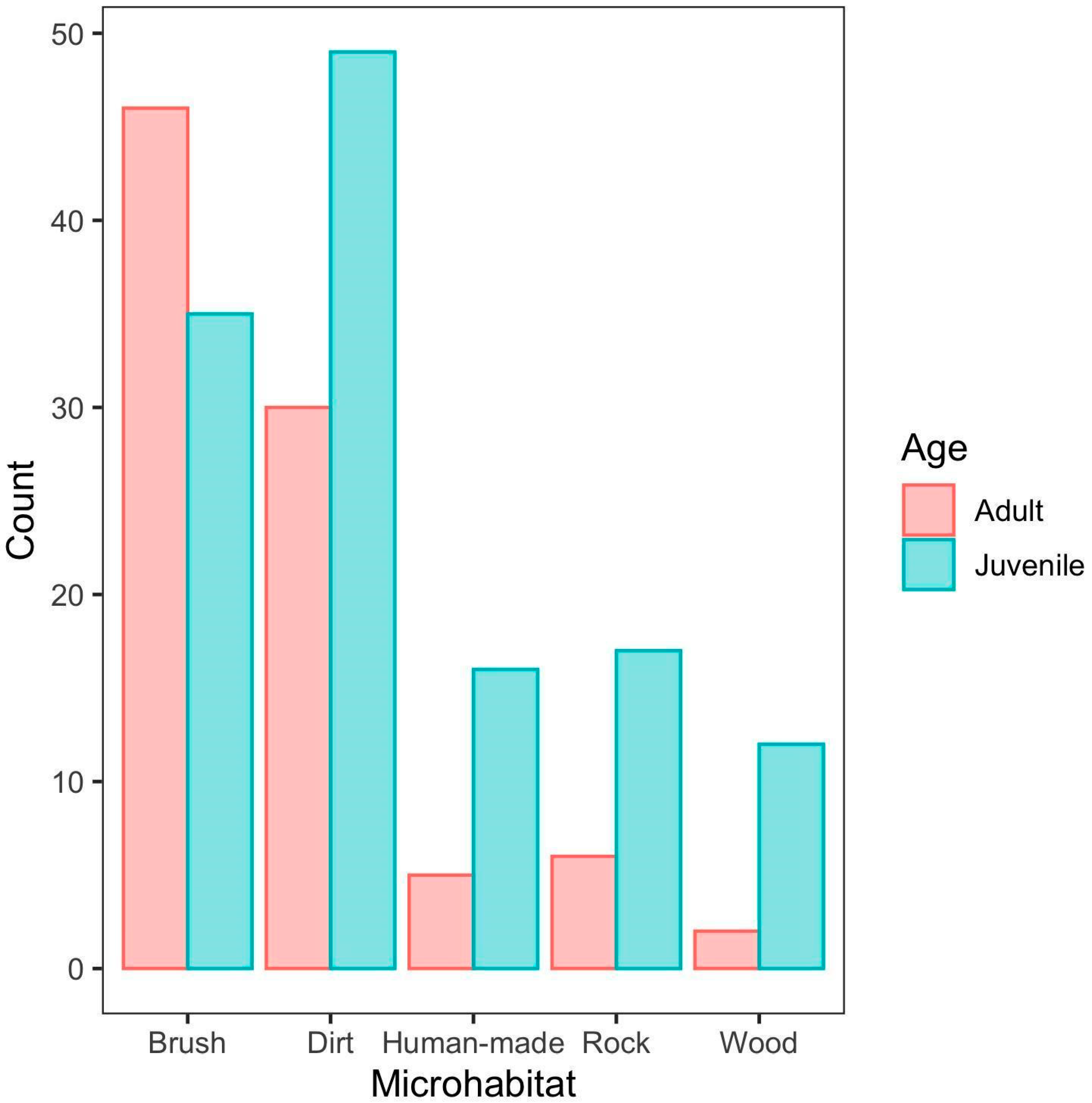
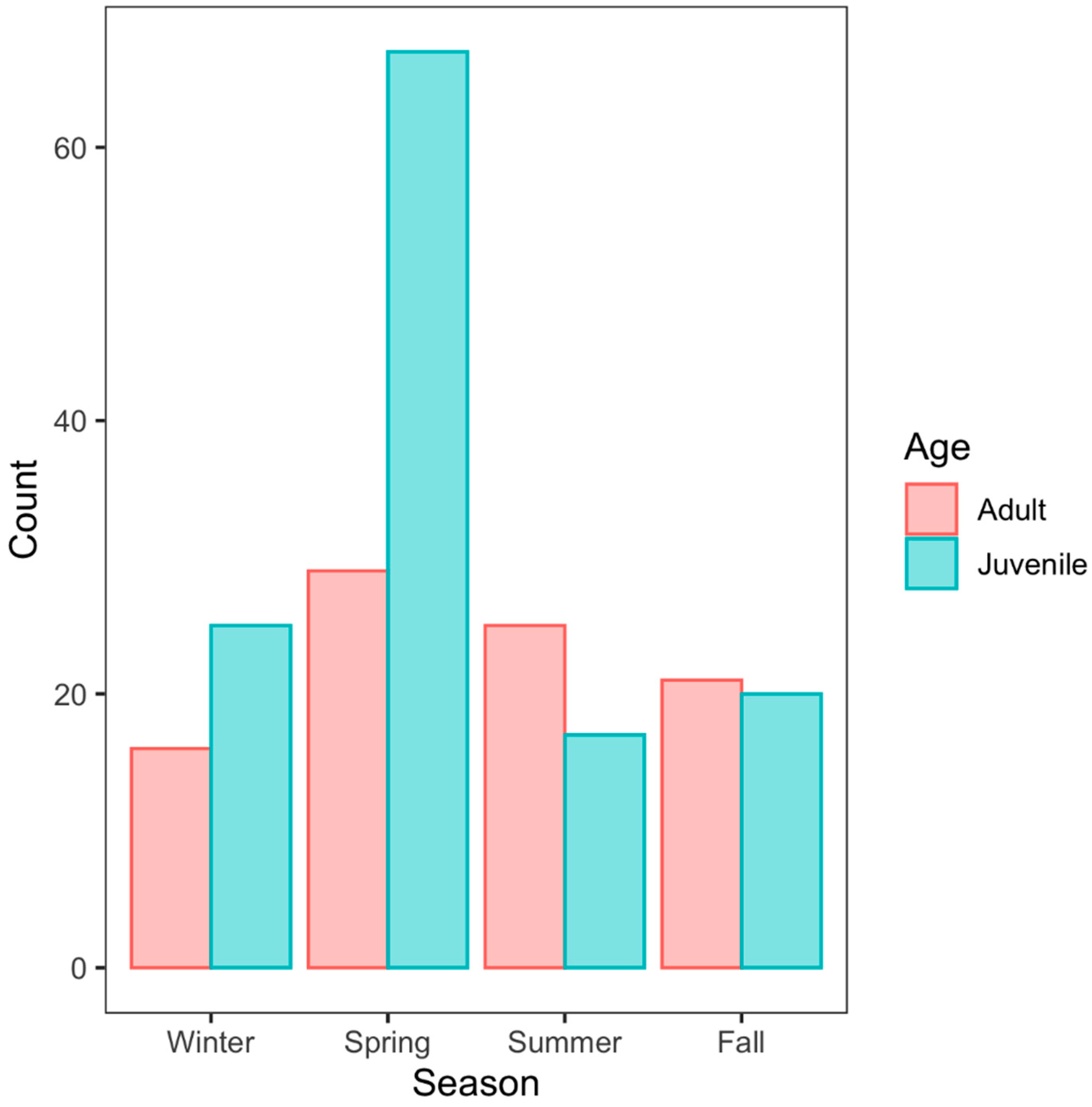
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burkholder, D.A.; Heithaus, M.R.; Fourqurean, J.W.; Wirsing, A.; Dill, L.M. Patterns of top-down control in a seagrass ecosystem: Could a roving apex predator induce a behaviour-mediated trophic cascade? J. Anim. Ecol. 2013, 82, 1192–1202. [Google Scholar] [CrossRef]
- Holt, A.R.; Davies, Z.G.; Tyler, C.; Staddon, S. Meta-Analysis of the effects of predation on animal prey abundance: Evidence from uk vertebrates. PLoS ONE 2008, 3, e2400. [Google Scholar] [CrossRef]
- Brown, J.S.; Kotler, B.P.; Smith, R.J.; Wirtz II, W.O. The effects of owl predation on the foraging behavior of heteromyid rodents. Oecologia 1988, 76, 408–415. [Google Scholar] [CrossRef]
- Woo, K.J.; Elliott, K.H.; Davidson, M.; Gaston, A.J.; Davoren, G.K. Individual specialization in diet by a generalist marine predator reflects specialization in foraging behaviour. J. Anim. Ecol. 2008, 77, 1082–1091. [Google Scholar] [CrossRef]
- Dézerald, O.; Céréghino, R.; Corbara, B.; Dejean, A.; Leroy, C. Functional trait responses of aquatic macroinvertebrates to simulated drought in a Neotropical bromeliad ecosystem. Freshw. Biol. 2015, 60, 1917–1929. [Google Scholar] [CrossRef]
- Brossard, D.; Lewenstein, B.; Bonney, R. Scientific knowledge and attitude change: The impact of a citizen science project. Int. J. Sci. Educ. 2005, 27, 1099–1121. [Google Scholar] [CrossRef]
- Cooper, C.B.; Dickinson, J.; Phillips, T.; Bonney, R. Citizen science as a tool for conservation in residential ecosystems. Ecol. Soc. 2007, 12, 11. [Google Scholar] [CrossRef]
- Thomson, R.C.; Wright, A.N.; Shaffer, H.B. California Amphibian and Reptile Species of Special Concern; University of California Press: Oakland, CA, USA, 2016. [Google Scholar]
- Martin, W.H.; Brown, W.S.; Possardt, E.; Sealy, J.B. The Biology of Rattlesnakes; Hayes, W.K., Beaman, K.R., Cardwell, M.D., Bush, S.P., Eds.; Loma Linda University Press: Loma Linda, CA, USA, 2008. [Google Scholar]
- Waldron, J.L.; Lanham, J.D.; Bennett, S.H. Using behaviorally-based seasons to investigate canebrake rattlesnake (Crotalus horridus) movement patterns and habitat selection. Herpetologica 2006, 62, 389–398. [Google Scholar] [CrossRef]
- Lowe, C.H. The Venomous Reptiles of Arizona; Arizona Game and Fish Department: Phoenix, AZ, USA, 1986. [Google Scholar]
- Ernst, C.H. Venomous Reptiles of North America; Smithsonian Books: Washington, DC, USA, 1999. [Google Scholar]
- Szymanski, J.; Pollack, C.; Ragan, L.; REdmer, M.; Clemency, L.; Voorhies, K.; Jaka, J. Species Status Assessment for the Eastern Massasauga Rattlesnake (Sistrurus catenatus); U.S. Fish and Wildlife: Bloomington, IN, USA, 2016.
- Martin, W.H.; Means, B.D. Distribution and habitat relationships of the eastern diamondback rattlesnake (Crotalus adamanteus). Herpetol. Nat. Hist. 2000, 7, 9–34. [Google Scholar]
- Stohlgren, K.M.; Spear, S.F.; Stevenson, D.J. A Status and Distribution Model for the Eastern Diamondback Rattlesnake (Crotalus adamanteus) in Georgia; U.S. Fish and Wildlife: Tiger, GA, USA, 2015.
- Hect, K.A.; Nickerson, M.A.; Colclough, P.B. Hellbenders (Cryptobranchus alleganiensis) may exhibit an ontogenetic dietary shift. Southeast. Nat. 2017, 16, 157–162. [Google Scholar] [CrossRef]
- Subalusky, A.; Fitzgerald, L.A.; Smith, L.L. Ontogenetic niche shifts in the american alligator establish functional connectivity between aquatic systems. Biol. Conserv. 2009, 142, 1507–1514. [Google Scholar] [CrossRef]
- Cundall, D.; Greene, H. Feeding: Form, Function, and Evolution in Tetrapod Vertebrates; Schwenk, K., Ed.; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Clark, R.W.; Tangco, S.; Barbour, M.A. Field video recordings reveal factors influencing predatory strike success of free-ranging rattlesnakes (Crotalus spp.). Anim. Behav. 2012, 84, 183–190. [Google Scholar] [CrossRef]
- Harmel, M.V.; Crowell, H.L.; Whelan, J.M.; Taylor, E.N. Rattlesnake colouration affects detection by predators. J. Zool. 2020, 311, 260–268. [Google Scholar] [CrossRef]
- Taylor, E.N. Diet of the Baja California Rattlesnake, Crotalus enyo (Viperidae). Copeia 2001, 2001, 553–555. [Google Scholar] [CrossRef]
- Mociño-Deloya, E.; Setser, K.; Heacker, M.; Peurach, S. Diet of New Mexico ridge-nosed rattlesnake (Crotalus willardi obscurus) in the Sierra San Luis and Sierra Pan Duro, México. J. Herpetol. 2015, 48, 104–107. [Google Scholar] [CrossRef]
- Glaudas, X.; Jezkova, T.; Rodrıguez-Robles, J.A. Feeding ecology of the Great Basin Rattlesnake (Crotalus lutosus, Viperidae). Can. J. Zool. 2008, 86, 723–734. [Google Scholar] [CrossRef]
- Holycross, A.T.; Painter, C.W.; Prival, D.B.; Swann, D.E.; Schroff, M.J.; Edwards, T.; Schwalbe, C.R. Diet of Crotalus lepidus klauberi (Banded Rock Rattlesnake). J. Herpetol. 2002, 36, 589–597. [Google Scholar] [CrossRef]
- Clark, R.W.; Dorr, S.W.; Whitford, M.D.; Freymiller, G.A.; Putman, B.J. Activity cycles and foraging behaviors of free-ranging sidewinder rattlesnakes (Crotalus cerastes): The ontogeny of hunting in a precocial vertebrate. Zoology 2016, 119, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Avila-Villegas, H.; Martins, M.; Arnaud, G. Feeding ecology of the endemic rattleless rattlesnake, Crotalus catalinensis, of Santa Catalina Island, Gulf of California, Mexico. Copeia 2007, 1, 80–84. [Google Scholar] [CrossRef]
- Lomas, E.; Maida, J.R.; Bishop, C.A.; Larsen, K.W. Movement ecology of northern pacific rattlesnakes (Crotalus o. oreganus) in response to disturbance. Herpetologica 2019, 75, 153–161. [Google Scholar] [CrossRef]
- Harvey, D.S.; Weatherhead, P.J. Habitat selection as the mechanism for thermoregulation in a northern population of massasauga rattlesnakes (Sistrurus catenatus). Ecoscience 2010, 17, 411–419. [Google Scholar] [CrossRef]
- Lind, C.M.; Beaupre, S.J. Natural variation in steroid hormone profiles of male Timber Rattlesnakes, Crotalus horridus, in northwest Arkansas. Gen. Comp. Endocrinol. 2014, 206, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Capehart, G.D.; Escallón, C.; Vernasco, B.J.; Moore, I.T.; Taylor, E.N. No drought about it: Effects of supplemental hydration on the ecology, behavior, and physiology of free-ranging rattlesnakes. J. Arid Environ. 2016, 134, 79–86. [Google Scholar] [CrossRef]
- Jaramillo-Alba, J.L.; Díaz de la Vega-Pérez, A.H.; Bucio-Jiménez, L.E.; Méndez-De la Cruz, F.R.; Pérez-Mendoza, H.A. Comparative thermal ecology parameters of the mexican dusky rattlesnake (Crotalus triseriatus). J. Therm. Biol. 2020, 92, 102695. [Google Scholar] [CrossRef] [PubMed]
- Hale, V.L.; Macgowan, B.; Corriveau, L.; Huse, D.C.; Currylow, A.F.T.; Thompson, S. Radio transmitter implantation and movement in the wild timber rattlesnake (Crotalus horridus). J. Wildl. Dis. 2017, 53, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Mata-Silva, V.; Desantis, D.L.; Wagler, A.E.; Johnson, J.D. Spatial ecology of Rock Rattlesnakes (Crotalus lepidus) in far West Texas. Herpetologica 2018, 74, 245–254. [Google Scholar] [CrossRef]
- Amarello, M.; Nowak, E.M.; Taylor, E.N.; Schuett, G.W.; Repp, R.A.; Rosen, P.C.; Hardy, D.L. Potential environmental influences on variation in body size and sexual size dimorphism among Arizona populations of the western diamond-backed rattlesnake (Crotalus atrox). J. Arid Environ. 2010, 74, 1443–1449. [Google Scholar] [CrossRef]
- Petersen, C.E.; Goetz, S.M.; Dreslik, M.J.; Kleopfer, J.D.; Savitzky, A.H. Sex, mass, and monitoring effort: Keys to understanding spatial ecology of timber rattlesnakes (Crotalus horridus). Herpetologica 2019, 75, 162–174. [Google Scholar] [CrossRef]
- Wallace, R.L.; Diller, L. V Feeding Ecology of the Rattlesnake, Crotalus viridis oreganus, in Northern Idaho. J. Herpetol. 1990, 24, 246–253. [Google Scholar] [CrossRef]
- Bonney, R.; Phillips, T.B.; Ballard, H.L.; Enck, J.W. Can citizen science enhance public understanding of science? Public Underst. Sci. 2016, 25, 2–16. [Google Scholar] [CrossRef]
- Newman, G.; Chandler, M.; Clyde, M.; McGreavy, B.; Haklay, M.; Ballard, H.; Gray, S.; Scarpino, R.; Hauptfeld, R.; Mellor, D.; et al. Leveraging the power of place in citizen science for effective conservation decision making. Biol. Conserv. 2016, 208, 55–64. [Google Scholar] [CrossRef]
- Kobori, H.; Dickinson, J.L.; Washitani, I.; Sakurai, R.; Amano, T.; Komatsu, N.; Kitamura, W.; Takagawa, S.; Koyama, K.; Ogawara, T.; et al. Citizen science: A new approach to advance ecology, education, and conservation. Ecol. Res. 2016, 31, 1–19. [Google Scholar] [CrossRef]
- Losey, J.E.; Perlman, J.E.; Hoebeke, E.R. Citizen scientist rediscovers rare nine-spotted lady beetle, Coccinella novemnotata, in eastern North America. J. Insect Conserv. 2007, 11, 415–417. [Google Scholar] [CrossRef]
- Ziter, C.D.; Herrick, B.M.; Johnston, M.R.; Turner, M.G. Ready, set, go: Community science field campaign reveals habitat preferences of nonnative asian earthworms in an urban landscape. Bioscience 2021, 71, 280–291. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Bino, G.; Major, R.E.; Martin, J.M.; Lyons, M.B.; Kingsford, R.T. Heterogeneous urban green areas are bird diversity hotspots: Insights using continental-scale citizen science data. Landsc. Ecol. 2019, 34, 1231–1246. [Google Scholar] [CrossRef]
- Lye, G.C.; Osborne, J.L.; Park, K.J.; Goulson, D. Using citizen science to monitor Bombus populations in the UK: Nesting ecology and relative abundance in the urban environment. J. Insect Conserv. 2012, 16, 697–707. [Google Scholar] [CrossRef]
- Putman, B.J.; Williams, R.; Li, E.; Pauly, G.B. The power of community science to quantify ecological interactions in cities. Sci. Rep. 2021, 11, 3069. [Google Scholar] [CrossRef]
- Klauber, L.M. Rattlesnakes, Their Habits, Life Histories, and Influence on Mankind; Abridged; University of California Press: Berkeley, CA, USA, 1982; ISBN 0520040392, 9780520040397. [Google Scholar]
- Hughes, A.L.; Cauthen, J.; Driscoll, C. Testing for behavioral lateralization in observational data: A monte carlo approach applied to neck-looping in american flamingos. Wilson J. Orinthol. 2014, 126, 345–352. [Google Scholar] [CrossRef]
- Frasnelli, E. Brain and behavioral lateralization in invertebrates. Front. Psychol. 2013, 4, 939. [Google Scholar] [CrossRef]
- Shine, R.; Olsson, M.M.; LeMaster, M.P.; Moore, I.T.; Mason, R.T. Are snakes right-handed? Asymmetry in hemipenis size and usage in gartersnakes (Thamnophis sirtalis). Behav. Ecol. 2000, 11, 411–415. [Google Scholar] [CrossRef]
- Roth, E.D. “Handedness” in snakes? Lateralization of coiling behaviour in a cottonmouth, Agkistrodon piscivorus leucostoma, population. Anim. Behav. 2003, 66, 337–341. [Google Scholar] [CrossRef]
- Hoso, M.; Asami, T.; Hori, M. Right-handed snakes: Convergent evolution of asymmetry for functional specialization. Biol. Lett. 2007, 3, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, R.C.; McGinnis, S.M. Amphibians and Reptiles of California: Revised Edition, 1st ed.; University of California Press: Berkely, CA, USA, 2012; ISBN 9780520270510. [Google Scholar]
- Putman, B.J.; Barbour, M.A.; Clark, R.W. The Foraging Behavior of Free-ranging Rattlesnakes (Crotalus oreganus) in California Ground Squirrel (Otospermophilus beecheyi) Colonies. Herpetologica 2016, 72, 55–63. [Google Scholar] [CrossRef]
- Wittenberg, R.D. Foraging ecology of the Timber Rattlesnake (Crotalus horridus) in a fragmented agricultural landscape. Herpetol. Conserv. Biol. 2012, 7, 449–461. [Google Scholar]
- Webber, M.M.; Glaudas, X.; Rodríguez-Robles, J.A. Do sidewinder rattlesnakes (Crotalus cerastes, Viperidae) cease feeding during the breeding season? Copeia 2012, 100–105. [Google Scholar] [CrossRef]
- Lind, C.M.; Husak, J.F.; Eikenaar, C.; Moore, I.T.; Taylor, E.N. The relationship between plasma steroid hormone concentrations and the reproductive cycle in the Northern Pacific rattlesnake, Crotalus oreganus. Gen. Comp. Endocrinol. 2010, 166, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Putman, B.J.; Clark, R.W. Behavioral thermal tolerances of free-ranging rattlesnakes (Crotalus oreganus) during the summer foraging season. J. Therm. Biol. 2017, 65, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Putman, B.J.; Lind, C.; Taylor, E.N. Does size matter? Factors influencing the spatial ecology of northern pacific rattlesnakes (Crotalus oreganus oreganus) in Central California. Copeia 2013, 2013, 485–492. [Google Scholar] [CrossRef]
- Kardong, K.V. Morphology of the respiratory system and its musculature in different snake genera. II. Charina bottae. Gegenbaurs Morphol. Jahrb. 1972, 117, 364–376. [Google Scholar] [PubMed]
- van Soldt, B.J.; Metscher, B.D.; Poelmann, R.E.; Vervust, B.; Vonk, F.J.; Muller, G.B.; Richardson, M.K. Heterochrony and early left-right asymmetry in the development of the cardiorespiratory system of snakes. PLoS ONE 2015, 10, e116416. [Google Scholar] [CrossRef] [PubMed]
- Saviola, A.J.; Chiszar, D.; Mackessy, S.P. Ontogenetic shift in response to prey-derived chemical cues in prairie rattlesnakes Crotalus viridis viridis. Curr. Zool. 2012, 58, 549–555. [Google Scholar] [CrossRef]
- Goetz, S.M.; Petersen, C.E.; Rose, R.K.; Kleopfer, J.D.; Savitzky, A.H. Diet and foraging behaviors of Timber Rattlesnakes, Crotalus horridus, in Eastern Virginia. J. Herpetol. 2016, 50, 520–526. [Google Scholar] [CrossRef]
- Grolle, E.K.; Lopez, M.C.; Gerson, M.M. Flight initiation distance differs between populations of Western Fence Lizards (Sceloporus occidentalis) at a rural and an urban Site. Bull. South. Calif. Acad. Sci. 2014, 113, 42–46. [Google Scholar] [CrossRef][Green Version]
- Samia, D.S.M.; Nakagawa, S.; Nomura, F.; Rangel, T.F.; Blumstein, D.T. Increased tolerance to humans among disturbed wildlife. Nat. Commun. 2015, 6, 9877. [Google Scholar] [CrossRef]
- Webber, M.M.; Jezkova, T.; Rodríguez-Robles, J.A. Feeding Ecology of Sidewinder Rattlesnakes, Crotalus cerastes (Viperidae). Herpetologica 2016, 72, 324–330. [Google Scholar] [CrossRef]
- Stebbins, R.C. A Field Guide to Western Reptiles and Amphibians: Field Marks of all Species in Western North America, 18th ed.; Houghton Mifflin: Boston, MA, USA, 1966. [Google Scholar]
- Clark, R.W. Fixed videography to study predation behavior of an ambush forgaing snake, Crotalus horridus. Copeia 2006, 2, 181–187. [Google Scholar] [CrossRef]
- Lang, S.D.J.; Mann, R.P.; Farine, D.R. Temporal activity patterns of predators and prey across broad geographic scales. Behav. Ecol. 2019, 30, 172–180. [Google Scholar] [CrossRef]
- Jungen, M.T. Eastern Diamondback Rattlesnake (Crotalus adamanteus) Telemetry Techniques and Translocation; Marshall University: Huntington, WV, USA, 2018. [Google Scholar]
- Bela, G.; Peltola, T.; Young, J.C.; Balázs, B.; Arpin, I.; Pataki, G.; Hauck, J.; Kelemen, E.; Kopperoinen, L.; Van Herzele, A.; et al. Learning and the transformative potential of citizen science. Conserv. Biol. 2016, 30, 990–999. [Google Scholar] [CrossRef]
- MacPhail, V.J.; Gibson, S.D.; Colla, S.R. Community science participants gain environmental awareness and contribute high quality data but improvements are needed: Insights from bumble bee watch. PeerJ 2020, 8, e9141. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urquidi, E.R.; Putman, B.J. Quantifying Southern Pacific Rattlesnake (Crotalus oreganus helleri) Hunting Behavior through Community Science. Diversity 2021, 13, 349. https://doi.org/10.3390/d13080349
Urquidi ER, Putman BJ. Quantifying Southern Pacific Rattlesnake (Crotalus oreganus helleri) Hunting Behavior through Community Science. Diversity. 2021; 13(8):349. https://doi.org/10.3390/d13080349
Chicago/Turabian StyleUrquidi, Emily R., and Breanna J. Putman. 2021. "Quantifying Southern Pacific Rattlesnake (Crotalus oreganus helleri) Hunting Behavior through Community Science" Diversity 13, no. 8: 349. https://doi.org/10.3390/d13080349
APA StyleUrquidi, E. R., & Putman, B. J. (2021). Quantifying Southern Pacific Rattlesnake (Crotalus oreganus helleri) Hunting Behavior through Community Science. Diversity, 13(8), 349. https://doi.org/10.3390/d13080349