1. Introduction
Pelagic, floating mats of primarily
Sargassum fluitans and
Sargassum natans algae are found in both tropical and temperate waters of the Atlantic Ocean [
1,
2]. These species are characterized with highly branched thalli and small air bladders (pneumatocysts) that keep the algae afloat [
1].
S. natans differs from
S. fluitans with thinner blades and a spine located on the pneumatocysts [
3] (see
Figure A1 in
Appendix A).
These floating mats range greatly in size from small patches less than 0.5 m in horizontal diameter up to huge mats 50 m in diameter [
1,
4]. Several factors contribute to this broad range in sizes. For example, sustained calm conditions can allow
Sargassum mats to form large aggregations, but harsher weather conditions such as high winds can break up large mats of
Sargassum or create narrower windrows [
4]. As the algae approach the shoreline, wave action can break up mats into significantly smaller sizes. Boats and other watercrafts running through the mats could also cause further disruption of
Sargassum mat structure and size. Wind can also push
Sargassum on to shores in the Atlantic and Caribbean, sometimes in huge mass events [
5]. However,
Sargassum can also be found in the Sargasso Sea in high quantities [
1,
6].
Some associated species, such as
Sargassum shrimp, form a symbiosis with these algae and are rarely found elsewhere [
1]. With so many species reliant either part of or their entire lives on the
Sargassum habitat, it has been designated as an Essential Fish Habitat [
7]. While many studies have focused on species richness and diversity within the
Sargassum habitat, very little is known about details of interactions among the symbionts and host algae [
3].
Sargassum shrimp are the most abundant macro invertebrates in
Sargassum mats.
There are two species of
Sargassum shrimp,
Latreutes fucorum and
Leander tenuicornis, both of which exhibit camouflage and coloration closely resembling the brown and yellow colors of
Sargassum algae, making it challenging for the observer to locate them within the habitat [
1]. These shrimp occupy positions in the algal mats based on the frond characteristics and depth of the fronds in the water column. For example,
L. tenuicornis shows a preference for deeper floating
Sargassum patches (10–12 cm below the surface) than shallow patches. Additionally,
L. tenuicornis positions itself in a parallel alignment with the fronds, thus allowing the shrimp to blend with the fronds and exhibit algal morphology mimicry [
8]. Such behaviors by
L. tenuicornis and its innate camouflage maximize protection from predators such as jacks, sargassum fish (
Histrio histrio) and the gray triggerfish (
Balistes capriscus), the last of which is one of the most abundant fish in
Sargassum and feeds almost exclusively on
Sargassum shrimp [
1,
7]. Clearly,
Sargassum shrimp are vital components of the
Sargassum community and food chain. Therefore, this study focused on these two species of shrimp.
A mutualistic symbiosis is formed when two organisms live together and provide reciprocal benefits. In the symbiosis between
Sargassum and these shrimp, the alga is a host that provides shelter and protection to the shrimp [
9]. In return, shrimp and other animals provide nutrients to the algae [
10]. Mutual benefits to symbionts can help explain why an association occurs, but another critical aspect is understanding how the symbiosis is initiated and maintained. Typically, the smaller, more mobile symbiont is the one to seek out and form the symbiosis with the larger symbiont or host [
3].
This leads to the question about how shrimp symbionts locate the
Sargassum patches, which are temporally and spatially highly variable in abundance [
11,
12,
13]. Presumably, the shrimp must initially locate such patches of
Sargassum algae. DNA analysis of
L. fucorum shows the shrimp are highly dispersive during the planktonic larval stage [
2]. Additionally, juveniles and those individuals separated from mats by biotic perturbations (e.g., disruptive feeding actions by large, pelagic species such as dolphinfish,
Coryphaena hippurus [
1,
13,
14,
15]), and abiotic perturbations [(e.g., wind and waves that break up patches, or when mats are driven onto beaches, cf. [
5]); and anthropogenic events (e.g., algal mat disruption by boats)] must locate and re-establish association with new, displaced or larger algal mats.
Visual cues are mostly available during the daytime and would be potentially available at limited distances only, based on the visual abilities of the shrimp. However, chemical cues would likely be available most of the time, and distance would be potentially less of a problem than using visual cues exclusively.
There is an abundance of chemical cues in the marine habitat, as every organism releases some type of chemical signal into the environment via metabolic activity [
16]. Once a chemical is released into the water, the molecules disperse by undergoing diffusion or bulk flow, which is the movement of molecules from high to low pressure [
16]. There are many marine organisms that can detect chemicals via chemoreceptors [
17]. Receptors must be both highly specific and diverse for organisms to identify an array of specific chemicals within the environment [
16]. These chemicals are used to locate food, avoid predators, find a mate, find a suitable habitat, homing, recognize conspecifics, and mediate social behavior (such as forming a hierarchy) [
17]. Specifically, chemoreceptive organs on decapods contain sensilla, which are either hair like or rod shaped [
17]. The sensilla are usually located on the antennules but can also be found on parts of the mouth and legs [
17].
The American lobster (
Homarus americanus) can detect specific chemical cues to recognize individual lobsters and form dominance hierarchies [
16]. Another example of a chemoreceptive crustacean is the porcellanid crab (
Porcellana sayana), which uses chemical cues to locate sea anemones associated with hermit crabs [
18]. Yet another crustacean, the rock shrimp (
Rhynchocinetes typus) uses chemoreception to find and select mates [
19].
Dimethylsulfoniopropionate (DMSP) is a chemical cue excreted by some marine algae, e.g., dinoflagellates [
20,
21] and benthic
Sargassum species [
22], that is detected by a variety of marine animals [
23,
24,
25]. It can potentially attract zooplankton predators to minimize grazing on the phytoplankton [
26]. Potentially high DMSP concentrations are also present in areas where primary productivity and foraging activity are high, which include windrows where
Sargassum and other potential DMSP-producing organisms aggregate to form weedlines [
27]. DeBose et al. [
27] showed that some fishes (e.g., jacks,
Caranx hippos and
C. melampygus) associated with
Sargassum mats responded significantly to cues of DMSP. DMSP concentrations of 10
−9 M in the Sargasso Sea were found, which indicate that this compound could be used as a cue for symbionts to locate
Sargassum patches [
28].
This current research paper is based on a prior study by Jobe and Brooks [
3] on the importance of chemical and visual cues for
Sargassum shrimp in locating and selecting habitat. The questions addressed by this previous study were whether chemical and visual cue responses to
Sargassum algae occurred by
L. tenuicornis or
L. fucorum, and whether these shrimp have preferences for a different
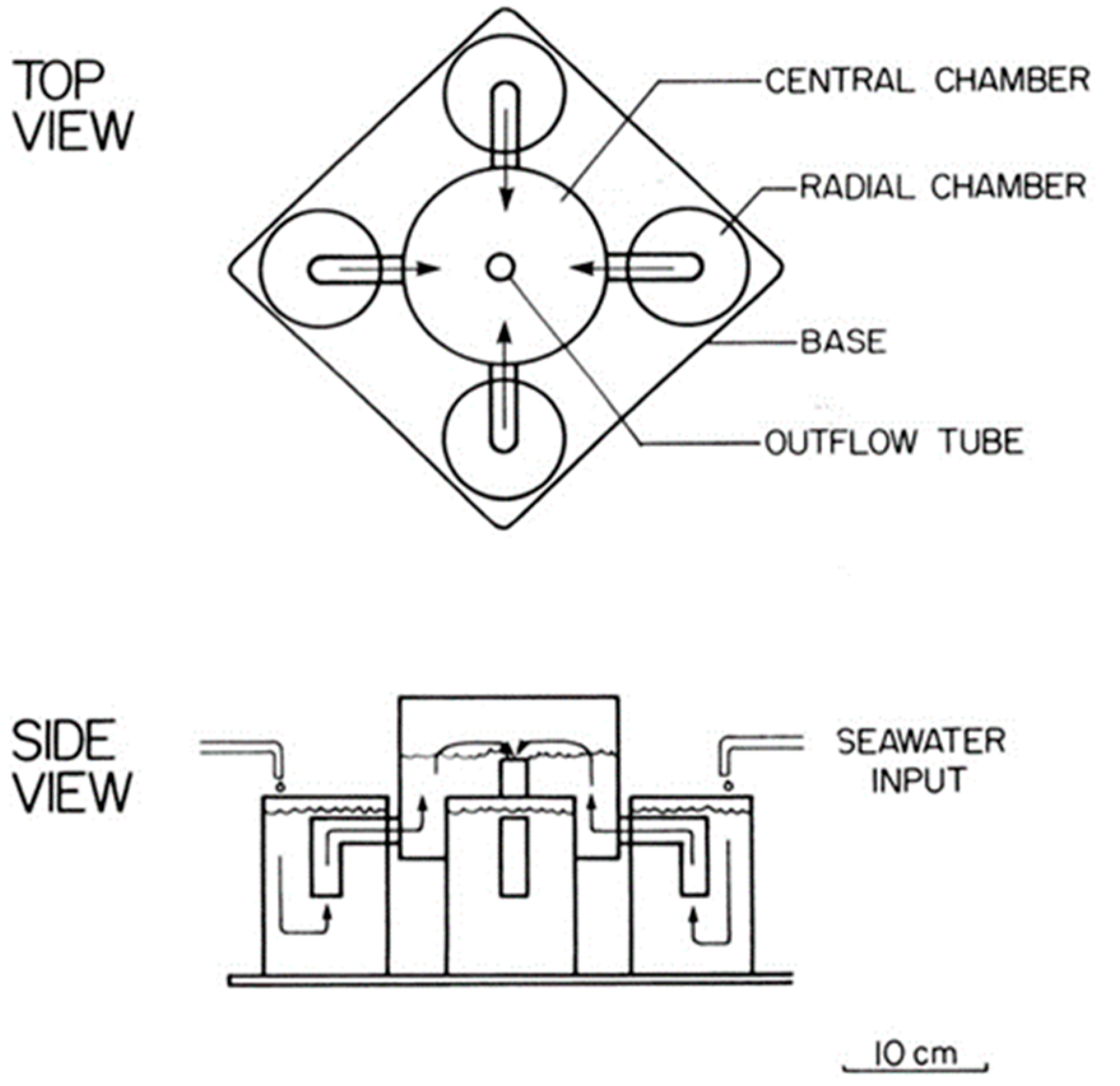
Sargassum species. In that study, two types of apparatus were used: (1) 4-chambered olfactometer (
Figure 1) to test for chemical cues only, and (2) aquarium with, first, chemical cues blocked and visual cues available (by placing the algae in beakers thereby allowing only visual cues), then chemical cues and visual cues by placing algae directly in the water with the shrimp.
Although there were no significant differences detected in the “chemical cues only” trials, there was significance in the “visual only” and the “visual plus chemical” cue trials. Specifically, “visual cues only” trials showed several specific significant results: (1) L. fucorum chose live Sargassum over artificial Sargassum and S. natans over S. fluitans; (2) small-sized L. tenuicornis chose S. natans over artificial Sargassum, and large-sized L. tenuicornis chose S. fluitans over S. natans and artificial Sargassum. However, when both visual and chemical cues were present, there were additional significant similar and differing results: (1) neither shrimp species had a significant preference for a specific Sargassum species, and (2) small L. tenuicornis selected S. fluitans over artificial algae. Although chemical cues from Sargassum were unavailable in the “visual cues only” trials, they were still present in the latter set of trials (i.e., both visual and chemical cues were available to the shrimp). While these results do not demonstrate that Sargassum shrimp responded to chemical cues in the 4-chambered apparatus trials, they do not preclude the possibility that chemoreception was employed by the shrimp.
The objective of this current research was to elaborate on the previous study to fully understand Sargassum shrimp chemoreception. Increases in sample sizes and both a 4-chambered and a Y-maze apparatus were used to test whether the shrimp could detect Sargassum cues, dimethylsulfoniopropionate (DMSP) (a chemical excreted by some marine algae), and conspecific cues.
Overall, these results showed the shrimp can detect chemical cues—in the absence of visual cues—that could affect initiating and maintaining this shrimp/algal symbiosis.
4. Discussion
4.1. Four-Chambered Apparatus Trials
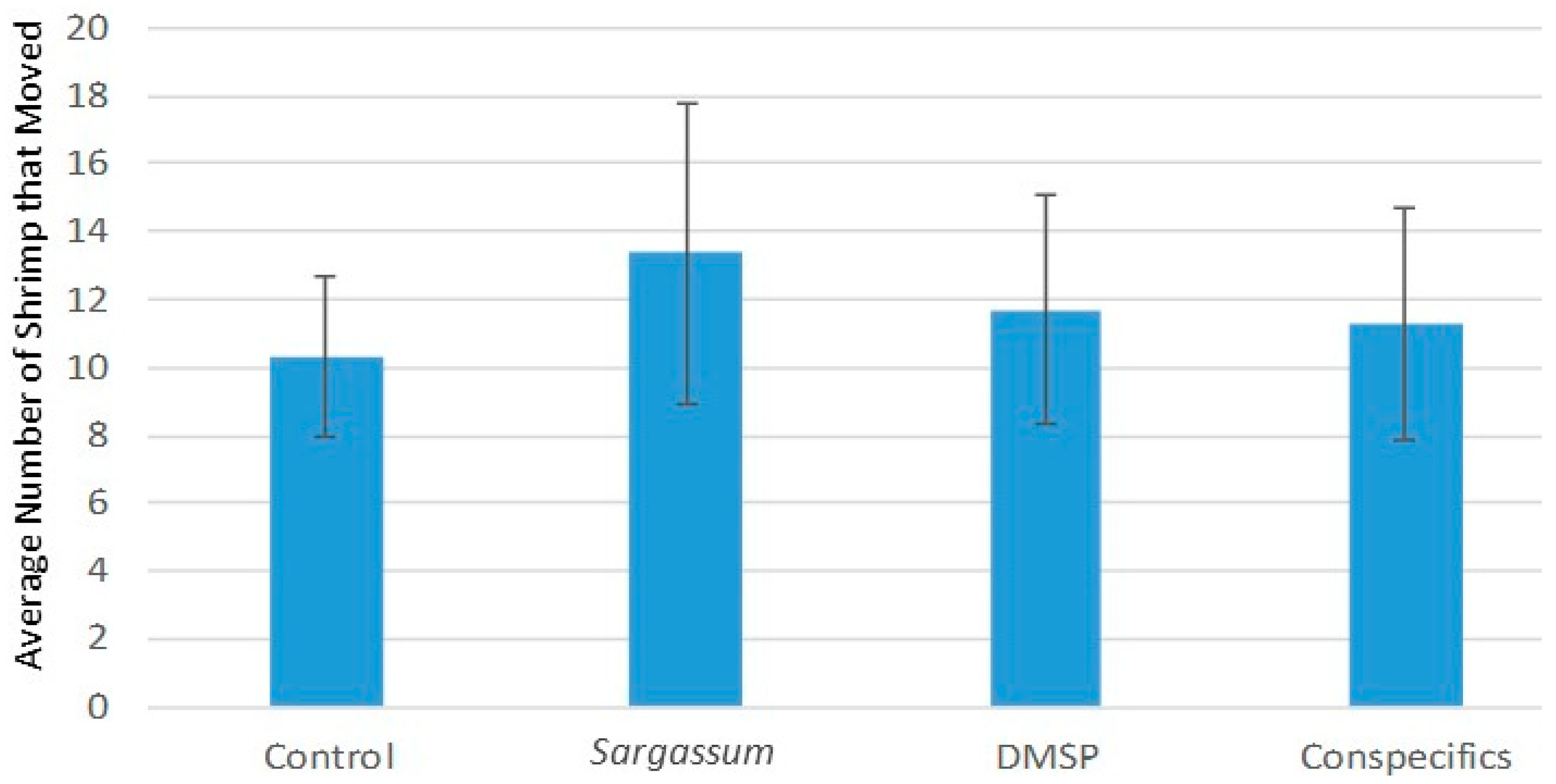
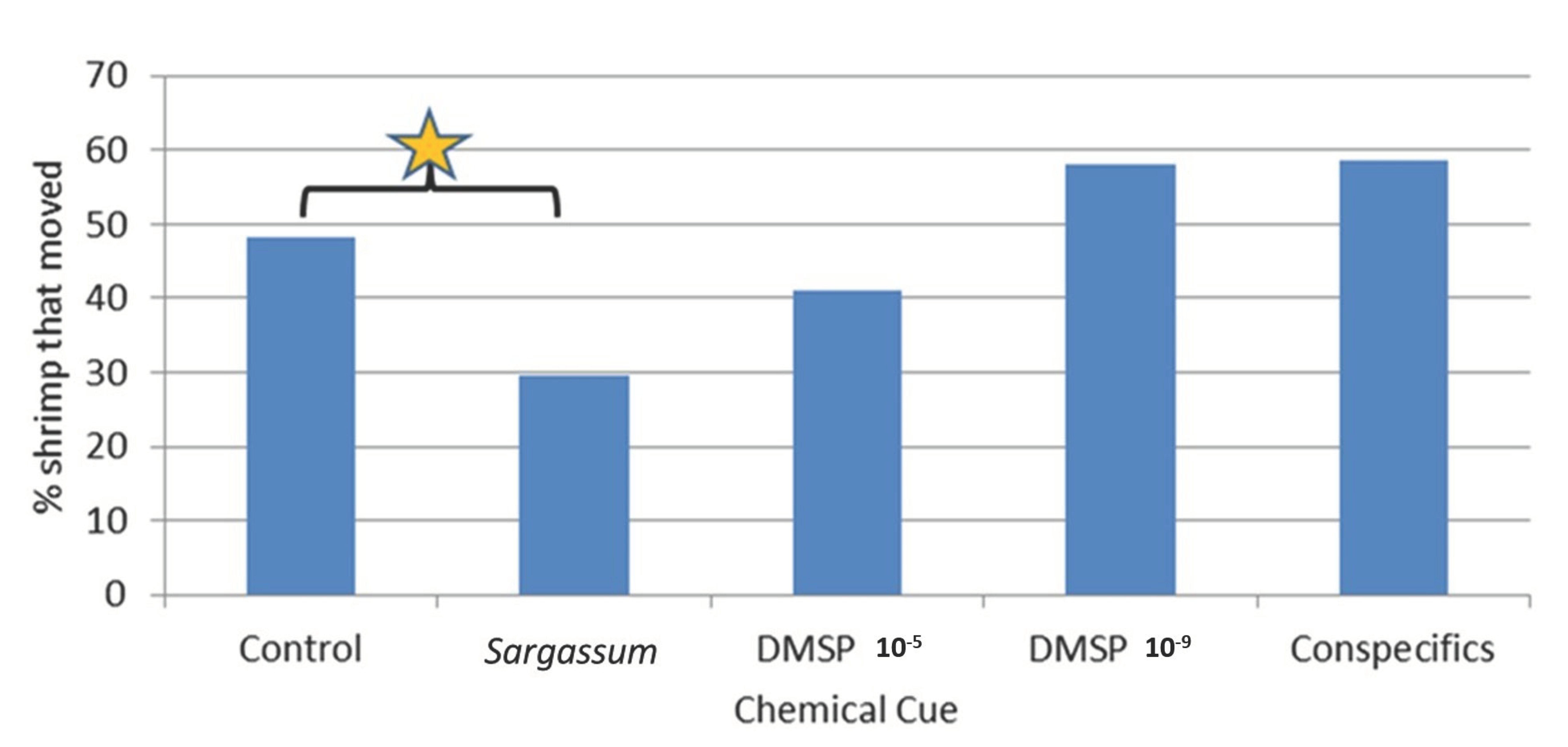
Results of these trials for
Latreutes fucorum, overall, were similar to those in the 2009 study by Jobe and Brooks [
3], which showed minimal response by the shrimp to chemical odors when visual cues were essentially unavailable. However, in our current study a post hoc
t-test comparing shrimp movements in the presence of
Sargassum odors showed a significant increase. This increase in movement did not show more shrimp specifically moving into the radial chamber with the
Sargassum chemical source but rather more shrimp moved into all of the four radial chambers in the presence of
Sargassum compared to the other treatments. This result suggests
L. fucorum increased its general searching behavior but was not precise in locating the directional aspects of the chemical cue source.
This slight difference between the past study and the current study, both of which used the same shrimp species, apparatus, and chemical cue type, could be related to improvements made on the preparation of the chemical cue in this study. The earlier study used a single algal frond placed in the source water at the beginning of each trial. Whereas in the current study we soaked the algal frond in seawater for 48 h prior to the start of the trials. This significantly longer algal soaking period prior to testing could potentially produce a stronger chemical cue source concentration for the shrimp to detect. Additionally, our current study increased the replications for the 4-chambered trials from 10 (in the 2009 study) to 14, which could also potentially increase the likelihood of detecting a significant response. Due to these initial, relatively low responses, serial dilutions involving potentially weaker concentrations of Sargassum chemical cue were not tested for either apparatus.
4.2. Y-Maze Apparatus Trials
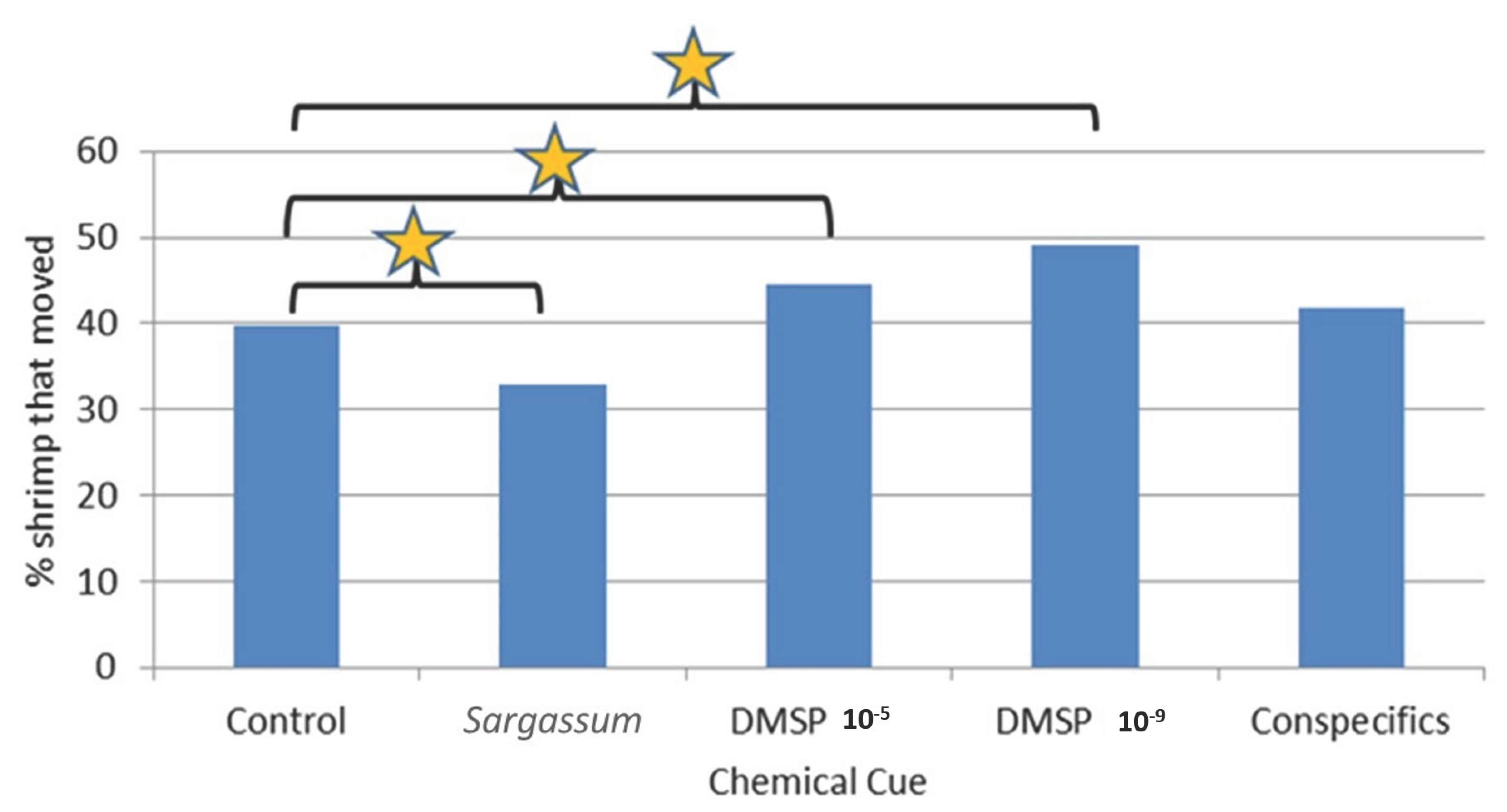
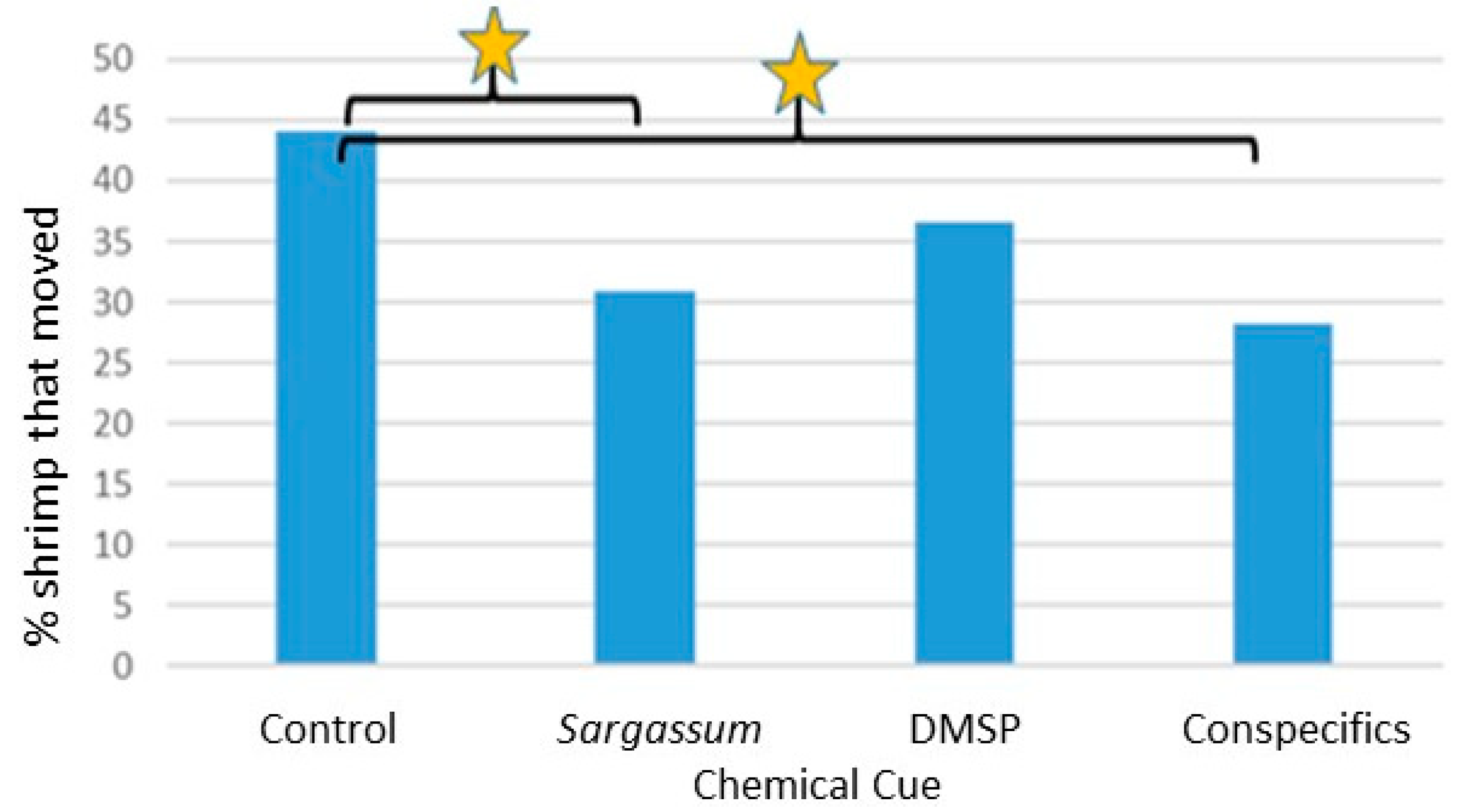
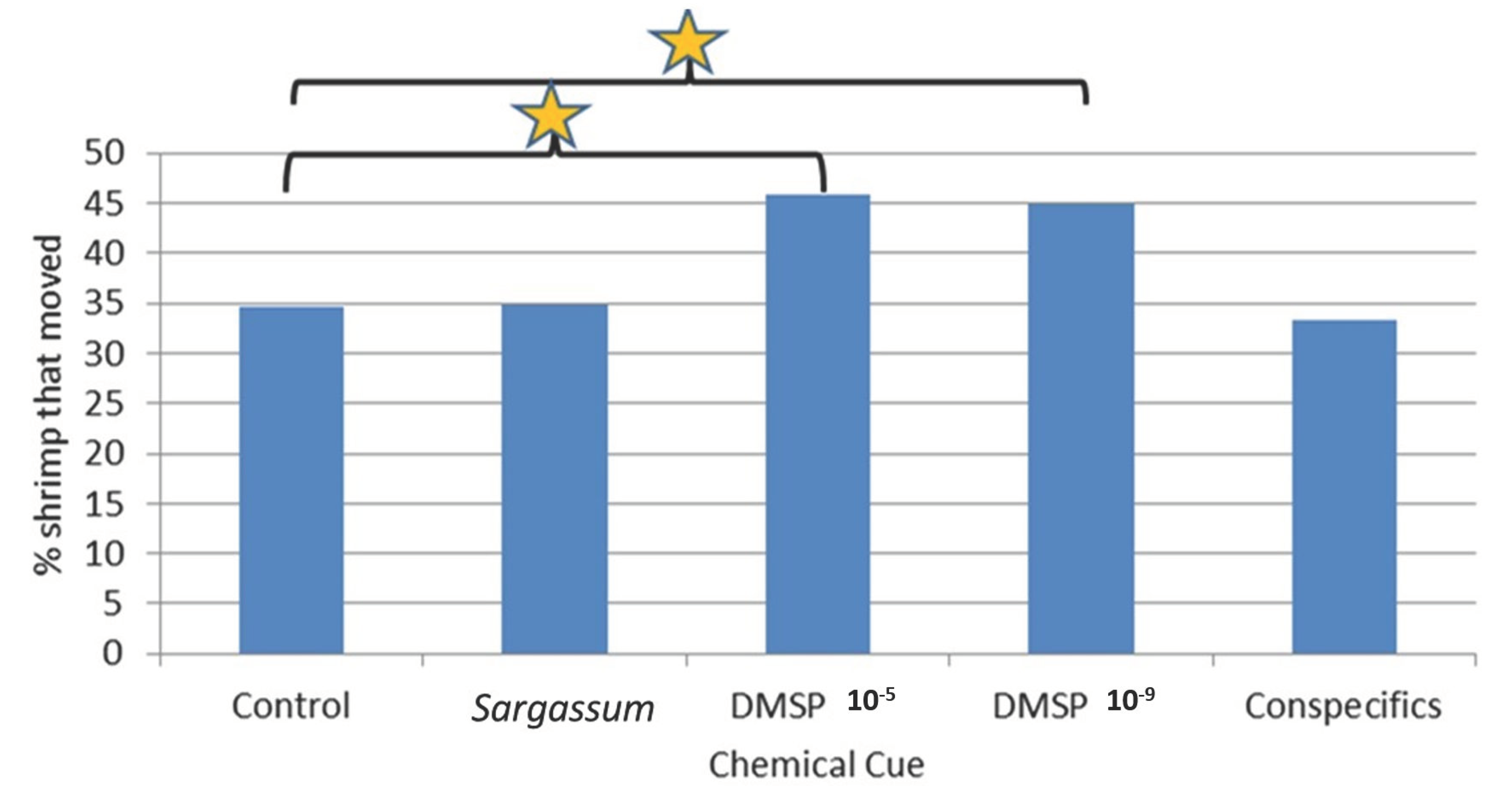
Results of these trials for Latreutes fucorum and Leander tenuicornis were compelling in that they showed a clear ability of both shrimp species to detect and respond to chemical cues associated with Sargassum (including DMSP) and conspecifics Such a strong response with the Y-maze compared to outcomes with the 4-chambered apparatus (for L. fucorum only, due to limited availability of L. tenuicornis) are likely related to notable differences between the experimental procedures used with these two different types of apparatus. For example, each replicate for the 4-cambered apparatus used 30 individuals simultaneously, which means that conspecific cues from each sex were presumably readily available to each shrimp throughout those trials. Additionally, shrimp tested in groups may respond differently to shrimp tested individually. That is, movement and choices may be directly influenced by the chemical presence and direct behavior of conspecifics. Because of the high numbers of shrimp typically found in situ within Sargassum fronds (personal observation), it is likely that testing in groups is closer to natural conditions. However, the Y-maze has some advantages in that the number of movement choices by the shrimp is limited to two arms (versus four radial chambers) and chemical cues can be more highly controlled (e.g., conspecific, including sex, cues can be delivered individually).
In the Y-maze trials, several specific and significant responses by both shrimp species to chemical cues were observed. Although rarely was the significant difference in selection of the arm with the cue versus non-cue arm detected, there were several significant differences observed between where the shrimp were eventually located, i.e., comparing shrimp that stayed in the holding area C versus moving to either arm of the Y-maze.
Interestingly, significant responses in several instances were to actually move less than that observed in controls without chemical cues, while others involved directional movements toward the chemical odor source (but not necessarily selecting the arm with the chemical cue, i.e., increase in searching behavior but not necessarily locating the cue source, e.g., significantly more L. fucorum moved in the presence of both DMSP chemical cue concentrations). This dichotomy of chemical cues triggering search behavior decreases in some situations and increases in others will be discussed below for the different cues.
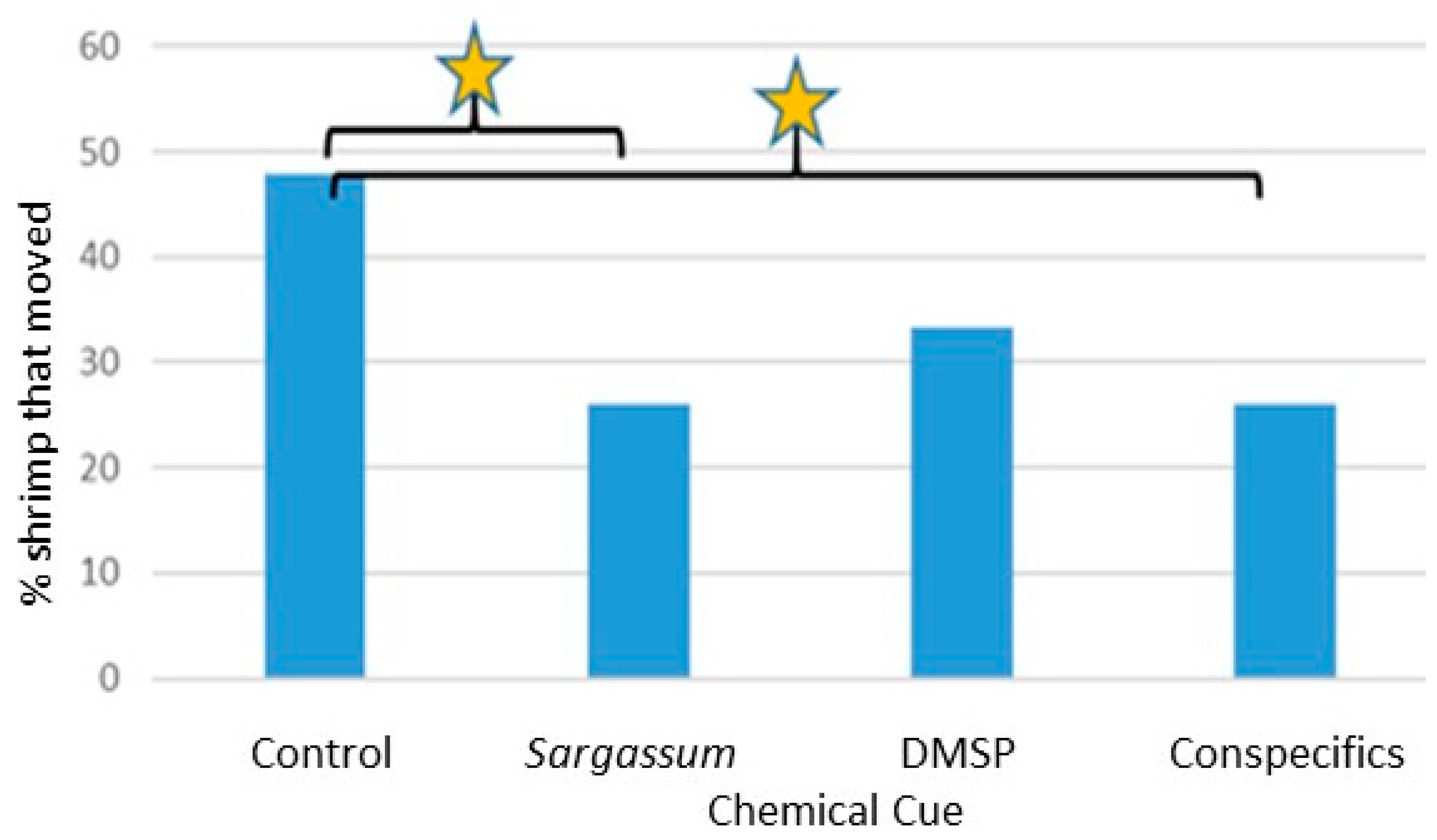
Significant responses to
Sargassum for both shrimp species involved fewer individuals moving out of the holding area C. Additionally, fewer
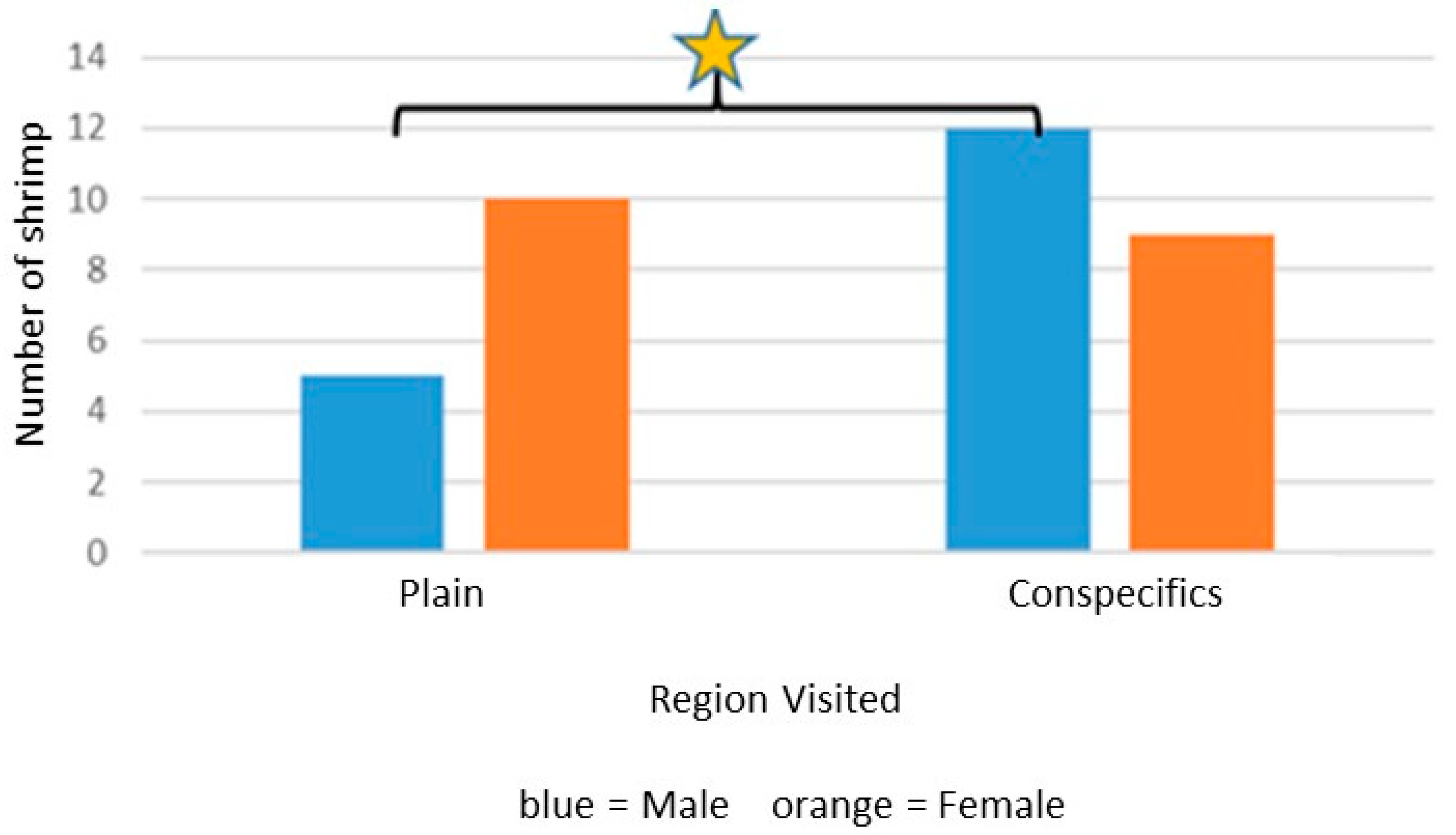
L. tenuicornis moved out of area C in the presence of conspecific chemical cues.
L. tenuicornis is the larger of the two species of shrimp; therefore, interactions observed in the lab between conspecifics have occasionally been aggressive, especially larger shrimp towards smaller ones. In general, the shrimp seemed to disperse when kept together in a small container. Conspecific chemical cues for
L. tenuicornis may have acted as a movement deterrent. For example, shrimp moving less in response to a conspecific chemical cue could be caused by conspecific shrimp releasing stress chemicals. A study on the crayfish
Procambarus clarkii showed that individuals moved away from conspecifics because of the release of chemicals (i.e., stress cues) into the water [
31]. In our study, handling of the shrimp when transferring them into the holding container (from which water dripped into an arm of the Y-maze) may have triggered the release of stress chemicals as well thereby affecting the response to conspecifics.
As pointed out previously, these shrimp are typically found in high concentrations in the fronds of
Sargassum. Isolating a shrimp in the Y-maze may trigger anti-predator behavior. For example, the freshwater shrimp
Atya lanipes will reduce its movement by up to 55% to avoid predation [
32]. It is possible the chemical cues released from
Sargassum effluents might have triggered a response to remain stationary. However, this would not explain why
L. fucorum moved more in the presence of the DMSP cues. This contradiction could indicate the chemical cues released into the water from
Sargassum are more complex than the single cue of DMSP (which presumably is released from
Sargassum, too, along with other compounds).
This phenomenon of chemical cues triggering less search activity also occurred with L. tenuicornis when exposed to conspecific chemical cues. Perhaps such cues indicated to this shrimp that moving towards the source would involve interaction with additional individuals. Potential negative interactions (e.g., competition or aggression or stress chemicals released, as discussed above) among these shrimp are not well understood, even though they are found in high abundance in situ.
Alternatively, significantly more L. fucorum moved in the presence of both DMSP chemical cue concentrations. This demonstrates the possibility this cue could be used by shrimp in locating Sargassum patches, too.
Again, while an explanation of the variation of response for some chemical cues causing less shrimp movement by some and more movement for others is problematic, what is clear is that a significant difference in behaviors of the shrimp was observed for the chemical-cue treatments of Sargassum, DMSP and conspecifics. We were also able to further parse the significant movement differences related to the sex of the shrimp. Below, we have summarized the significant differences to the chemical cues based on shrimp species and sex.
Sargassum Cue: L. fucorum males and L. tenuicornis females moved significantly less in the Y-maze in the presence of the Sargassum chemical cue.
DMSP Cue: L. fucorum females moved statistically more in the Y-maze in the presence of both DMSP dilutions.
Conspecific Cue:
- a.
L. tenuicornis females moved in the Y-maze statistically less in the presence of conspecific cues.
- b.
L. fucorum males that moved in the Y-maze significantly chose the conspecific cue side initially.
A limitation of our study is that the shrimp were not sexed until after use in a trial. Because female shrimp are larger than males [
33], when choosing shrimp from the holding aquarium, it is likely there was bias towards choosing females. This bias was especially noticeable for
L. fucorum trials, as this species is smaller and difficult to locate. Additionally, only shrimp with a total length of 10 mm or greater were used in the Y-maze to ensure visibility in the recorded videos. This bias caused fewer males to be tested than females (
L. fucorum, 1.88:1, female:male ratio;
L. tenuicornis, 1.6:1, female:male ratio). Having lower replications for male shrimp decreases the power of statistical analyses, and limits conclusions that can be made about these results. Future studies may choose to incorporate all sizes of shrimp and more replicates with males. Doing so, however, may be problematic as handling smaller shrimp while preventing injury, etc., is challenging.
The two size groups of shrimp did not have any statistically different responses to any of the chemical cues tested (chi-square, all
p > 0.05). While this may suggest no difference in response between life history stages of the shrimp, future studies should attempt to record larval shrimp responses to chemical cues, as it may be more important for the larvae to detect patches and settle into the habitat. Larvae of other species of invertebrate rely heavily on habitat chemical cues for settlement. For example, swimming larvae of the hydrozoan
Coryne uchidai settle in response to chemical cues from
Sargassum tortile [
34].
L. fucorum has a planktonic larval stage that lasts 18–30 days in the lab [
35]. Such a long period of time would indicate that the larvae must have a means to locate suitable habitat.
In some cases, an organism responds to certain chemical cues as a larva but may lose the capability to detect the same chemical cues as an adult. Fiddler crab (
Uca spp.) larvae can detect cues from food it typically consumes, but adults are insensitive to those same chemical cues [
36]. Those results suggest that the life history stage of an organism may influence which chemical cues they respond to based on requirements of the organism at different stages in their life history.
5. Conclusions
Sargassum habitat is part of a significant marine, pelagic holobiont (i.e., host and collective symbionts), which is a complex and diverse community, much like a floating version of a coral reef.
Sargassum is an Essential Fish Habitat [
7], and entire food chains and marine ecosystems rely on the success of this community of symbionts. Understanding how these biotic connections are established and maintained is critical. This study focused on two shrimp species that are abundant in
Sargassum; specifically, to expand on the previous study by Jobe and Brooks [
3] and clarify the role of available chemical cues for the shrimp to form and maintain their role in the
Sargassum holobiont.
Unequivocally, we show that both
Latreutes fucorum and
Leander tenuicornis can detect and respond to chemical cues in the absence of visual cues from
Sargassum patches. It is important to note that while chemoreception may be utilized, these shrimp are likely utilizing all senses available to maintain the association. As such, we envision that shrimp would use visual and chemical cues when both are available, as demonstrated by Jobe and Brooks [
3], such as during the day and nights associated with full (or near) moons. The current results demonstrate responses to chemical cues exclusively could be important when visual cues are mostly limited. Additionally, both species of
Sargassum shrimp are brooders and carry embryos until hatching [
37]. Because the shrimp are hatched within the
Sargassum patches, it is possible that planktonic larval and juvenile shrimp do not need to travel long distances to establish this symbiosis. However, natural disturbances such as storms and strong currents can disrupt and break apart
Sargassum patches thereby potentially separating shrimp and other symbionts from their resident patch. This would require relocating patches for all ages and sizes of shrimp. Having access to both chemical and visual cues would seem advantageous.
A study on chemical feeding cues for
L. tenuicornis found that only 5 out of the 28 single compounds used as cues were significantly stimulating the shrimp, which feed primarily on hydroids and bryozoans [
38]. This shows that chemoreception can be highly specific, and thus searching for an organism’s ability to use chemoreception may require testing several cues. Our study attempted to potentially isolate components of
Sargassum by using DMSP, which did separately trigger responses. Differences in shrimp response between
Sargassum and DMSP indicate that
Sargassum effluents are likely highly complex, involving potentially multiple specific compounds. Future studies should conduct a chemical assay on
Sargassum algae, potentially identifying more specific bioactive compounds that can be used to gain further insight into how this crucial mutualistic (alga supplies habitat and fish and shrimp provides nutrients via waste products); cf. [
10], symbiosis is formed and maintained.
It is abundantly clear how important the shrimp are to the continuity of the
Sargassum community. In addition to these shrimp species, several other
Sargassum organisms have been studied. The
Sargassum crab
Portunus sayi is abundant in this community and was also found to detect
Sargassum chemical cues [
39]. When looking at both this current study and the previously mentioned studies on this community, it is clear that chemoreception is a major mechanism being used by several organisms in the
Sargassum holobiont. Another study looked at habitat selection by both
L. tenuicornis and the common fish species,
Histrio histrio, and found that the organisms have preferred habitats based on structural complexity [
8]. It is likely that all of these organisms are interconnected and play an important role in maintaining this symbiotic community, and the use of chemoreception is one vital component to the success of this symbiosis.
Finally, the
Sargassum holobiont is in global crisis. Nutrient influx from anthropogenic sources has contributed to producing massive and sometimes harmful blooms of
Sargassum that are not only potentially affecting the components within this ecosystem but have already profoundly affected coastal ecosystems [
40] (Lapointe et al., 2021). It is more important than ever that we understand fully how
Sargassum communities function.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









