
Fanworms: Yesterday, Today and Tomorrow

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
3. Results
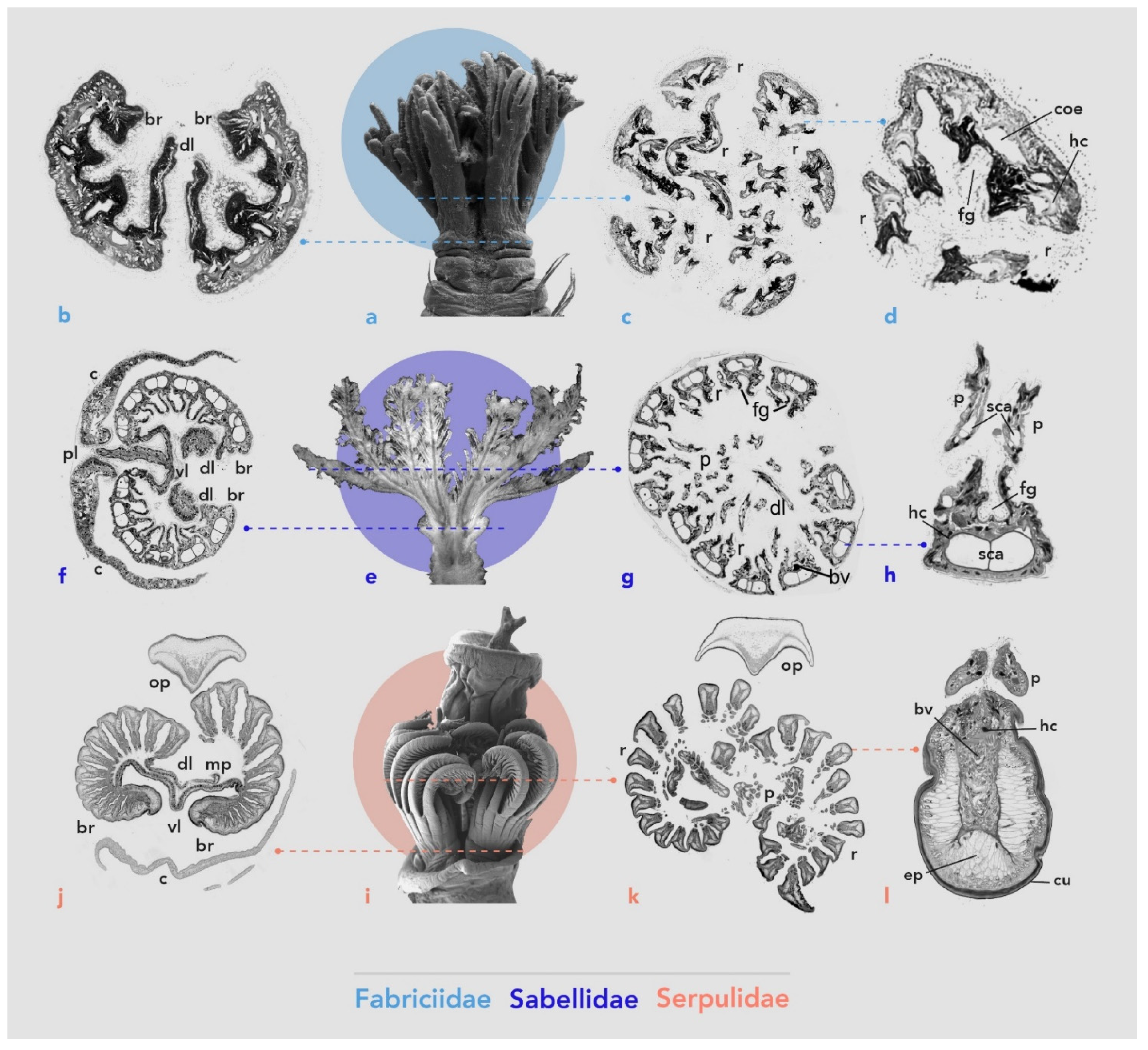
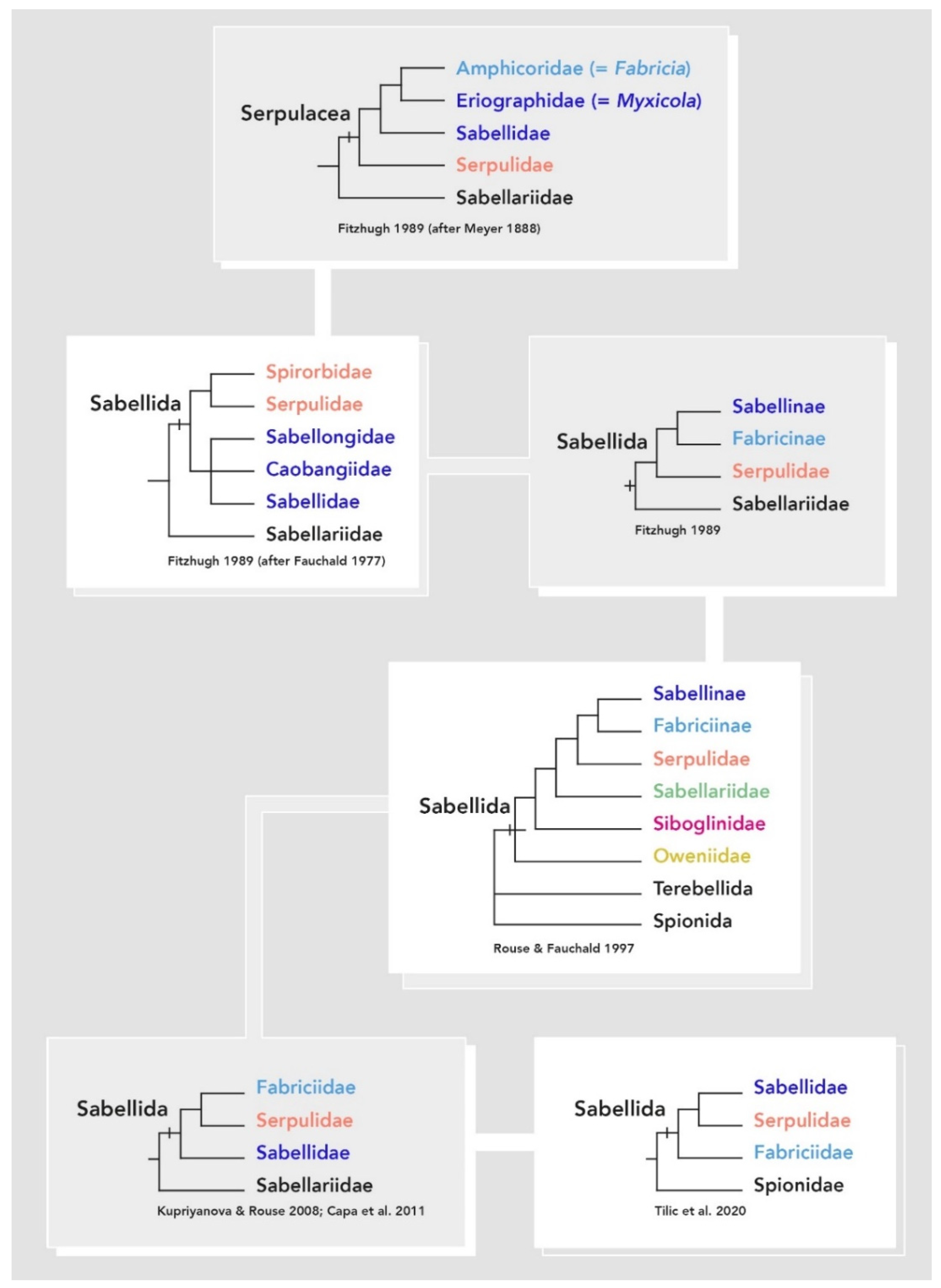
3.1. Systematics
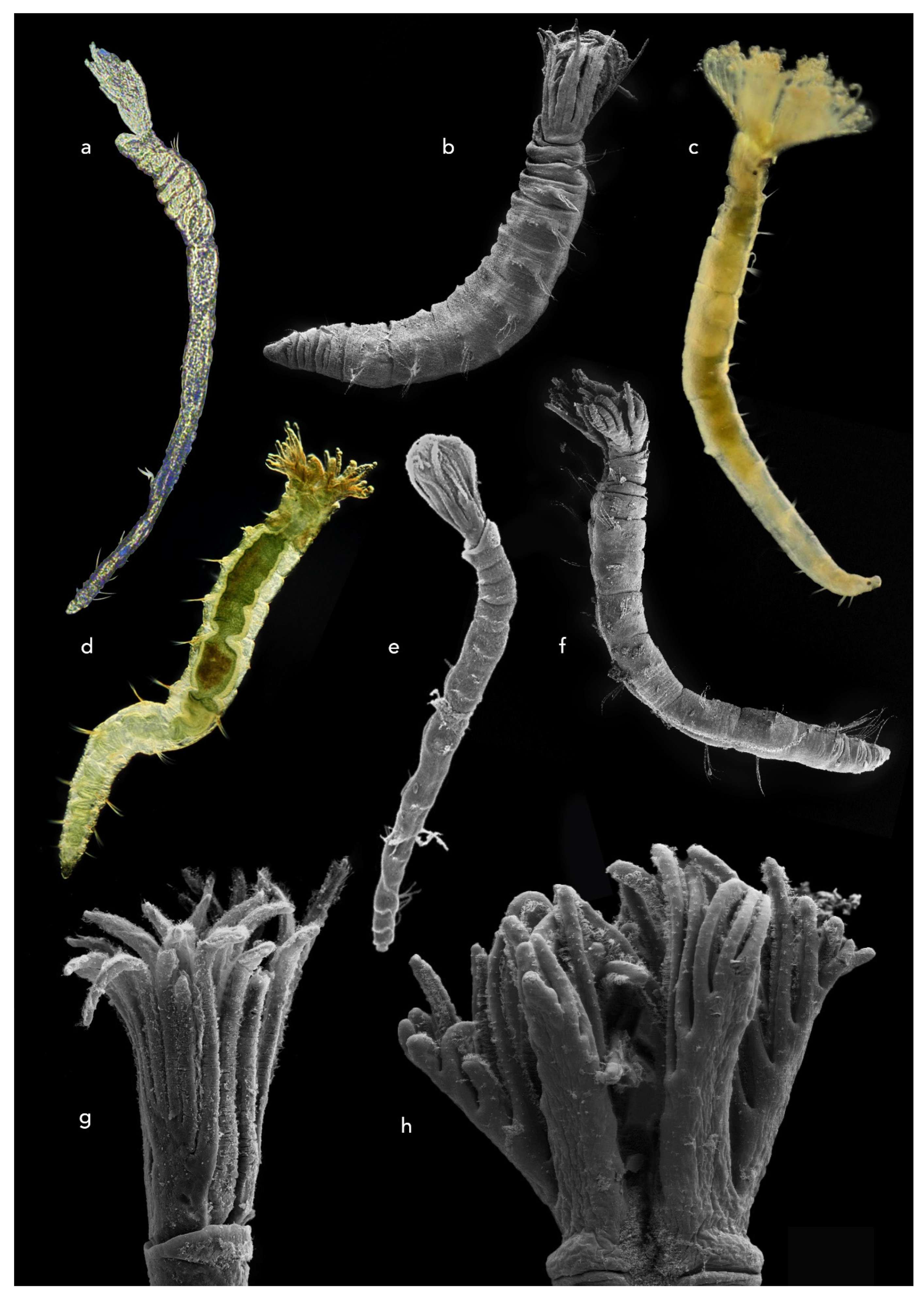
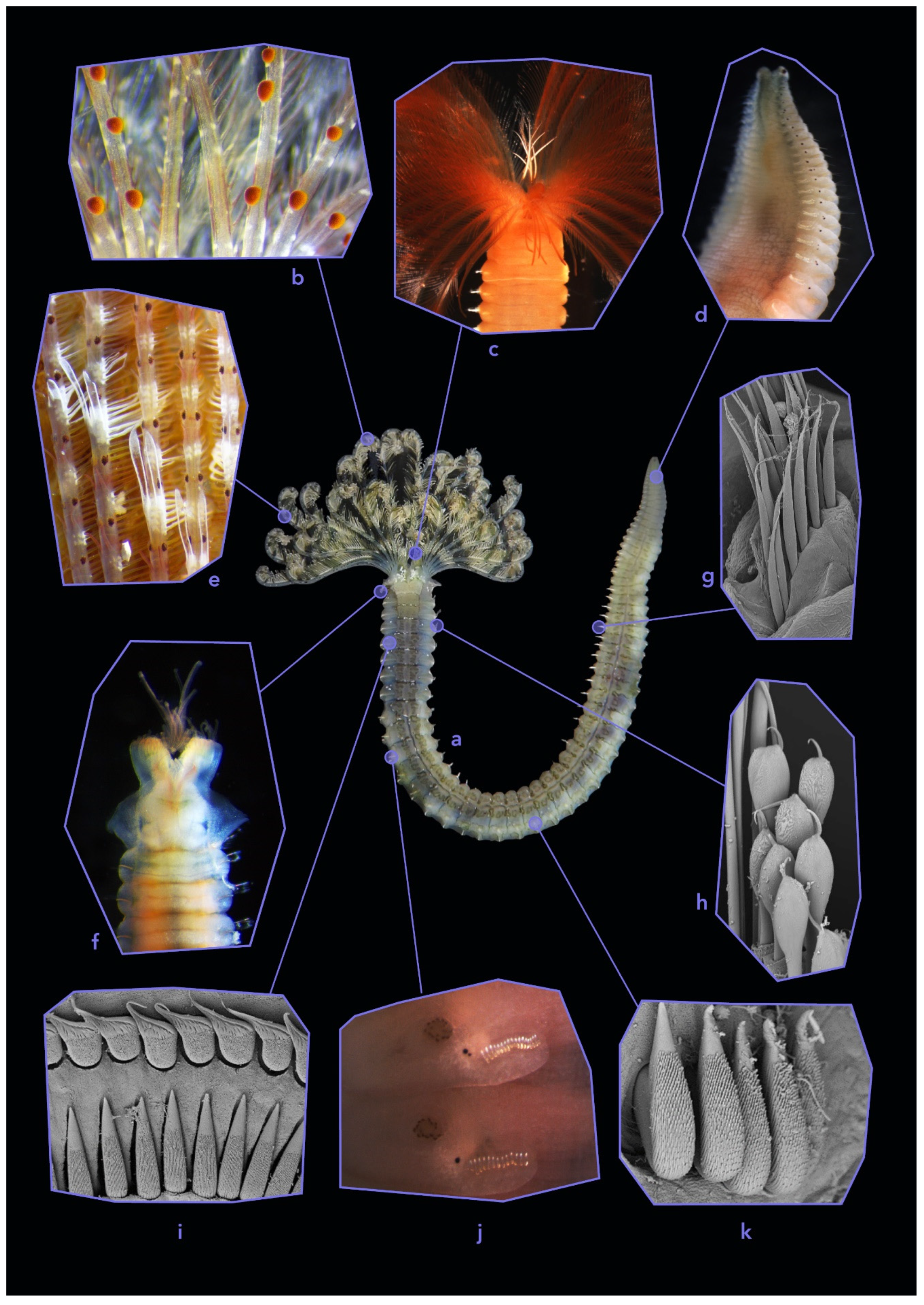
3.1.1. Fabriciidae
3.1.2. Sabellidae
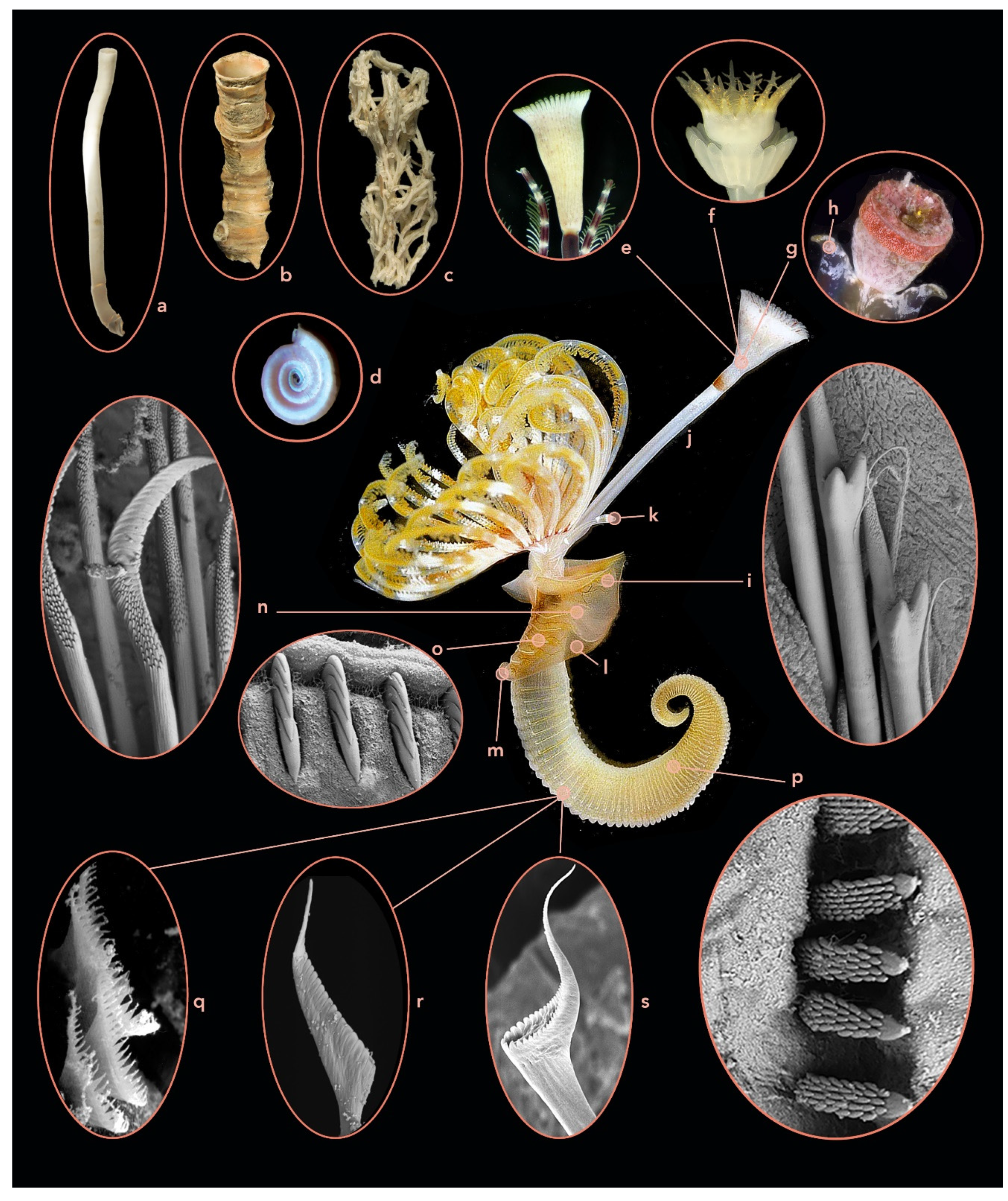
3.1.3. Serpulidae
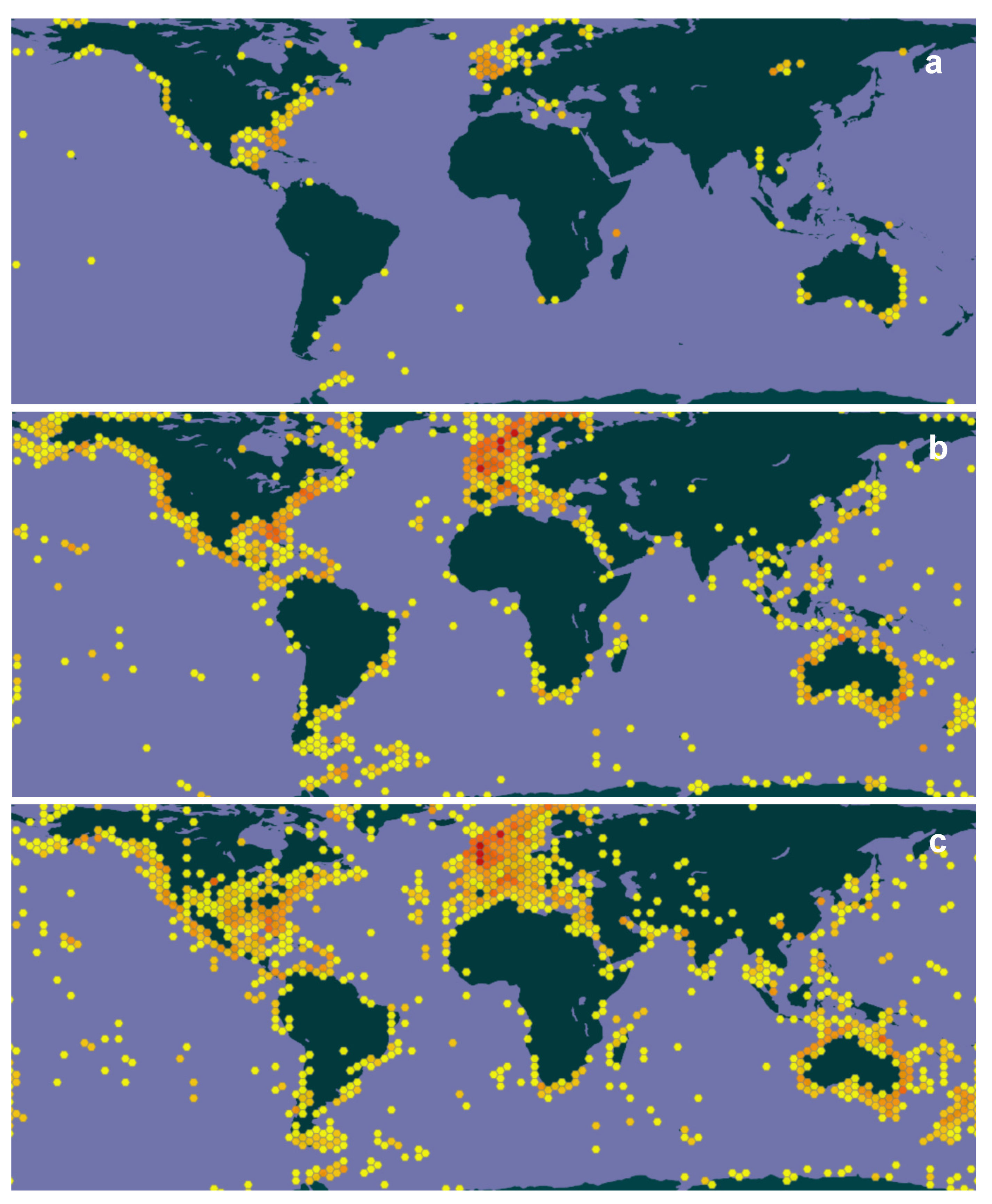
3.2. Diversity and Species Discovery
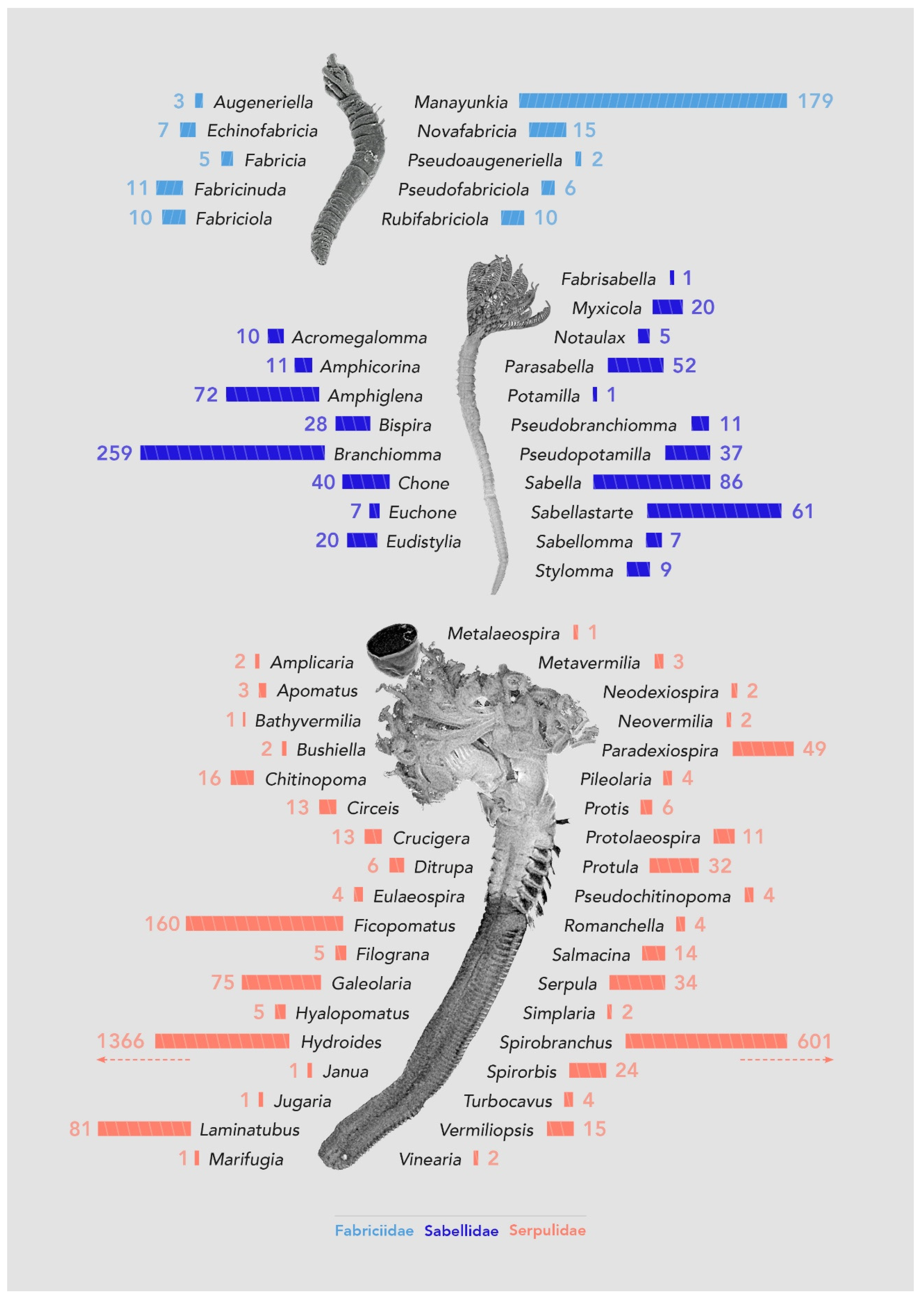
3.2.1. Number of Genera and Species
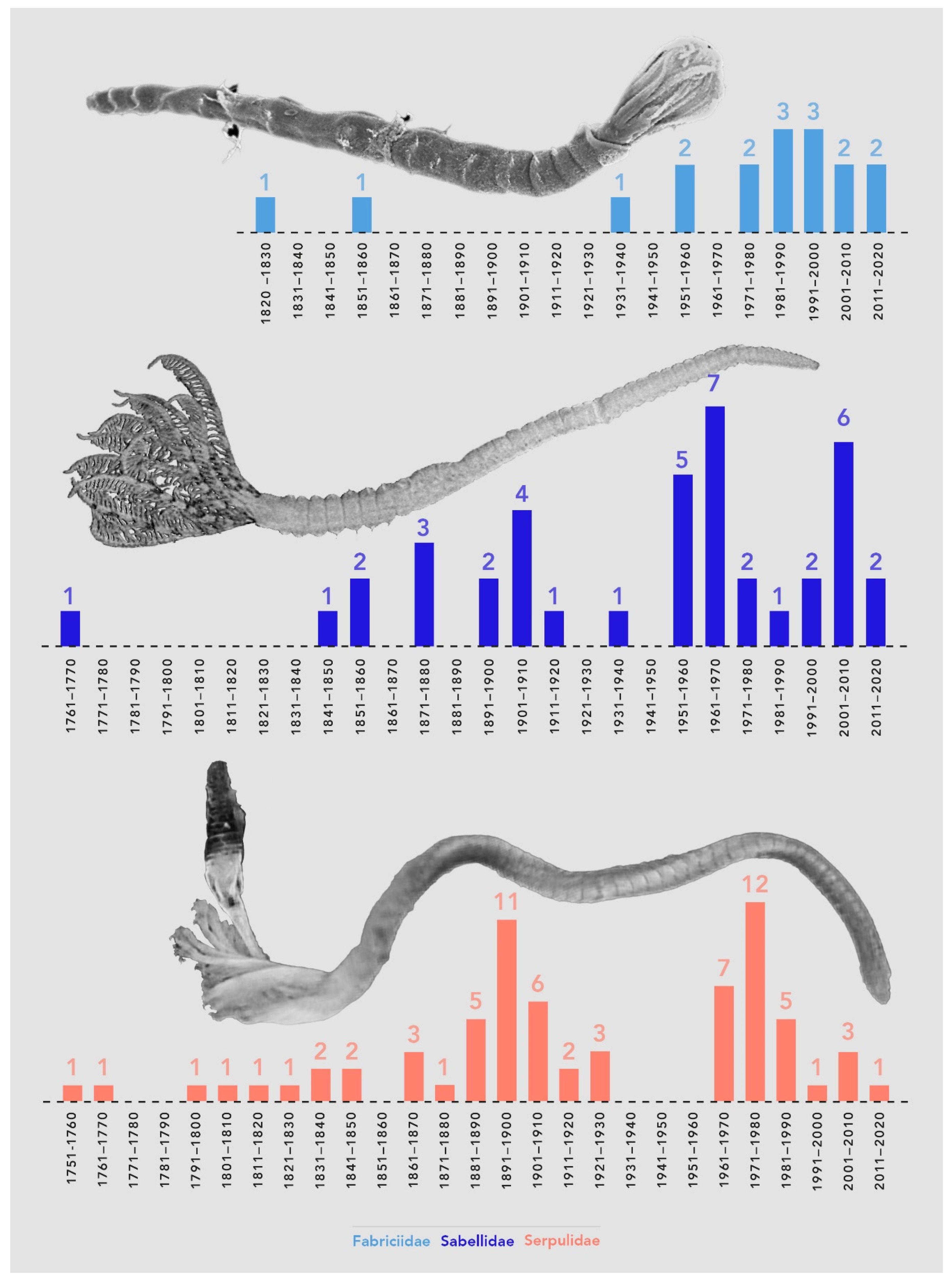
3.2.2. Taxonomists and Species Discovery
3.2.3. Identification Keys, Diversity Assessments, and Recent Regional Taxonomic Studies
Arctic
Temperate Northern Atlantic
Temperate Northern Pacific
Tropical Atlantic
Western Indo-Pacific
Central Indo-Pacific
Eastern Indo-Pacific
Tropical Eastern Pacific
Temperate South America
Temperate South Africa
Temperate Australasia
Southern Ocean
3.3. Diagnostic Characters and Techniques Used for Species Discrimination
3.3.1. Shortcuts to Identification of Fabriciidae
3.3.2. Shortcuts to Identification of Sabellidae
3.3.3. Serpulidae Diagnostic Characters
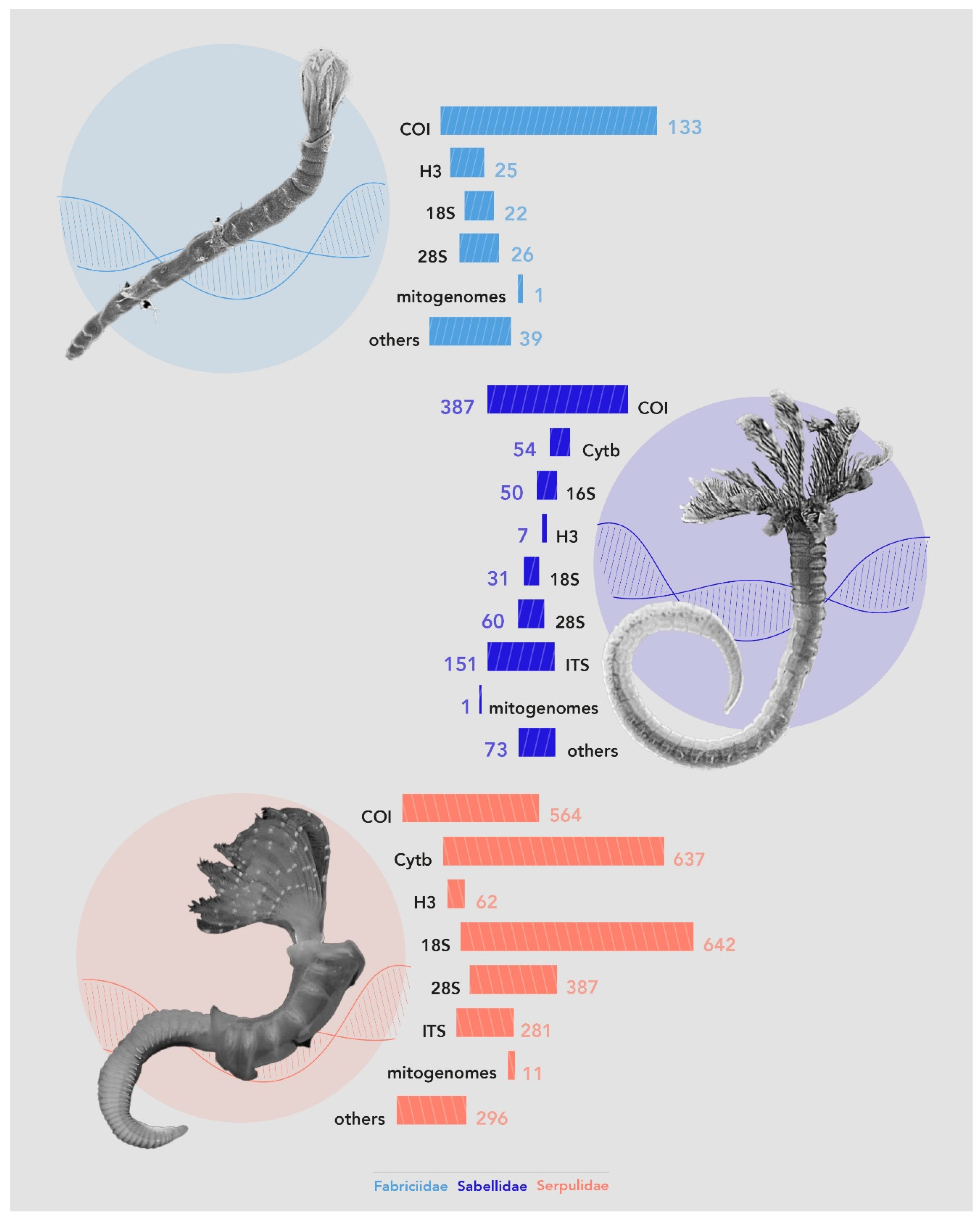
3.3.4. Data and Techniques Used for Species Identification and Systematics
Initial Collection, Observation and Fixation in the Field
Morphological Studies of Preserved Specimens
Genetic Data
Species Delimitation and Identification
3.4. Ecology, Distribution and Biogeography
3.4.1. Ecology
Fabriciidae
Sabellidae
Serpulidae
3.4.2. Biogeography, Distribution and Bathymetry
Fabriciidae
Sabellidae
Serpulidae
3.5. Non-Indigenous and Invasive Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Type Locality | Presumed Origin | Category | New Occurrences (Outside of Expected Natural Range) | Molecular Markers | Comments |
|---|---|---|---|---|---|---|
| Acromegalomma claparedei (Gravier, 1906) | Djibouti, Gulf of Aden | Red Sea | ? Non-indigenous | Western Mediterranean | none | Listed in the Mediterranean [382] but needs confirmation [130]. |
| Amphicorina pectinata (Banse, 1957) | Masked Island, New Zealand | Southern Island New Zealand | ? Non-indigenous | Western and Central Mediterranean | none | Redescribed from Italy and Spain without examination of types from New Zealand [383]. Included in the Mediterranean checklists [365,381,384] but requires confirmation. |
| Bispira polyomma Giangrande and Faasse in Faasse and Giangrande, 2012 | Yerseke Marina, the Netherlands | Unknown | ? Non-indigenous | NA | none | Found among cultured oysters, presumably as an introduction [385]. |
| Branchiomma bairdi (McIntosh, 1885) | Bermuda | Tropical Northwestern Atlantic | Non-indigenous | Gulf of California, Australia, Mediterranean, Suez Canal, northeastern Atlantic Ocean, California, Hawaii, Galapagos, Madeira | COI, Cytb, 16S, 28S, ITS | Widest distribution reported for a sabellid [72,126,129,147,185,189,376,377,379,386,387,388]. Some records need confirmation [185]. |
| Branchiomma boholense (Grube, 1878) | Bohol, Philippines | Western Coral Triangle | Non-indigenous | Mediterranean | COI | Reported from the Mediterranean [375,389,390]. Çinar [126] corrected his records as B. bairdi. Many records of B. bairdi in the Mediterranean are B. boholense [379]. |
| Branchiomma coheni Tovar-Hernández and Knight-Jones, 2006 | Panama, Pacific | Tropical Eastern Pacific | Non-indigenous | Florida, Gulf of California | none | Records from Tampa Bay and Gulf of California [391] first since description. In Florida it was an early detected non-indigenous species [391]. |
| Branchiomma conspersum (Ehlers, 1887) | Key West, Florida, USA | Floridian | Non-indigenous | Hawaii | ITS | Reported from Hawaii [185] |
| Branchiomma curtum (Ehlers, 1901) | Juan Fernández Island, Chile | Juan Fernández | Not translocated | ? Mexican Caribbean | none | Caribbean records may be erroneous [391]. Both syntypes of B. curtum and Caribbean specimens were juveniles produced by fission [392]. See comments in [391]. |
| Branchiomma luctuosum (Grube, 1870) | Red Sea | Red Sea | Non-indigenous | Mediterranean, Brazil | none | Common in the Mediterranean as Lessepsian migrant [129,389,390,393,394,395,396,397,398,399,400,401], and Brazil [234,402] |
| Desdemona ornata Banse, 1957 | South Africa | Agulhas | ? Non-indigenous | Iberian Peninsula, UK, Marmara Sea, Portugal | none | Reported as introduced in Australia [403], Spain [404], UK [405], Marmara Sea [406], Portugal [407] and the Netherlands [408], but types of D. ornata were not examined. |
| Euchone limnicola Reish, 1959 | Los Angeles, California, USA | Warm Temperate Northeast Pacific | ? Non-indigenous | Dunkerke, Australia | none | Reported from Australia by McArthur [409] and included in Hewitt et al. [380] based on McArthur dissertation. Reported by Guyonnet and Borg [410] from the French coast of North Sea. |
| Laonome calida Capa, 2007 | Queensland, Australia | Unknown | ? Non-indigenous | Australia (Queensland, Northern Territory, Western Australia), ? Europe (the Netherlands, Sea of Azov, Baltic Sea, Mosel River) | none | Reported in Australian in fully marine but also estuarine conditions, in both pristine and port environments [68]. Bick et al. [102] suggest that European records belong to Laonome xeprovala, not L. calida. |
| Laonome elegans Gravier, 1906 | Red Sea | Red Sea | ? Non-indigenous | East Mediterranean | none | Listed in Zenetos et al. [383]. Presence in the Mediterranean area as Lessepsian migrant needs to be confirmed. |
| Laonome triangularis Hutchings and Murray, 1984 | New South Wales, Australia | East Central Australian Shelf | ? Non-indigenous | Turkey | none | Reported as introduced in Turkey [126]. |
| Laonome xeprovala Bick and Bastrop in Bick et al. 2018 | Estonia, Baltic Sea | Unknown | Non-indigenous | the Netherlands, Sea of Azov, Baltic Sea, Mosel River | COI, 16S, 18S | Specimens from the Baltic Sea, the Netherlands and the Sea of Azov possessed identical genotypes, but unknown origin [102]. Confirmation that it is not the same as L. calida is needed. |
| Myxicola infundibulum (Renier in Meneghini, 1847) | Devon, UK | Northern European Seas | Non-indigenous | Australia, ?North America | COI, 16S | Analysed sequences of specimens from European and Australian waters belong to same species; sequences from Maine showed some differences [411]. |
| Parasabella fullo Grube, 1878) | Northern Japan | Temperate Northwest Pacific | Non-indigenous | Santa Barbara and San Diego, California, USA | none | Collected on ship hulls in California, and a resident population appears to exist in the region [412]. |
| Parasabella pallida Moore, 1923 | California, USA | Warm Temperate Northeast Pacific | Not translocated | NA | none | Included in the list of translocated species by mistake [294], as it was described from California [143] not the Caribbean, as later fixed in [73]. |
| Parasabella rugosa (Moore, 1904) | San Diego, California, USA | Warm Temperate Northeast Pacific | Non-indigenous | Australia | none | It was reported in Australia near an international port as sp. cf. P. rugosa Capa and Murray [73]. |
| Pseudobranchiomma emersoni Jones, 1962 | Port Jackson, Jamaica | Tropical Northwestern Atlantic | Non-indigenous | Australia | none | As cf. P. emersoni in Capa and Murray [74]. |
| Peudobranchiomma orientalis (McIntosh, 1885) | Hong Kong | Unknown | Non-indigenous | Australia | none | As cf. P. orientalis in Capa and Murray [74]. Ethanol fixed specimens are need for molecular analysis. |
| Pseudobranchiomma schizogenica Tovar-Hernández and Dean, 2014 | La Paz, Mexico | Tropical Northwestern Atlantic | Non-indigenous | Australia, Galapagos | ITS, Cytb | As cf. P. schizogenica in Capa and Murray [74] from Australia, also reported in Galapagos [183]. |
| Sabella spallanzanii (Gmelin, 1791) | Malta | Mediterranean | Non-indigenous | Australia, New Zealand | COI, H3, 18S, 28S, 16S, ITS | Considered an invasive pest in Australia and New Zealand [202,364,370,413]. |
| Sabellastarte spectabilis (Grube, 1878) | Bohol, Masalac, Philippines and Singapore | Western Coral Triangle | ? Non-indigenous | Sri Lanka, Solomon Islands, Mauritius, Japan, Taiwan, Hawaii, Malaysia, Saipan, Pakistan | COI, 16S | Reports of accidental introductions to Hawaii [414,415,416] rely on invalid morphological features [70]. Evidence for wide distribution exists [70]. |
| Terebrasabella heterouncinata Fitzhugh and Rouse, 1999 | South Africa | Unknown | Non-indigenous | California, Chile | none | Parasite of red abalone. Reported from abalone farms from California and Chile [66,300,301,302,303,304,305,306,307,308,309,310,311,312,313,314,315,316,317,318,319,320,321,322,323,324,325,326,327,328,329,330,331,332,333,334,335,336,337,338,339,340,341,342,343,344,345,346,347,348,349,350,351,352,353,354,355,356,357,358,359,360,361,362,363,364,365,366,367,368,369,370,371,372,373,374,375,376,377,378,379,380,381,382,383,384,385,386,387,388,389,390,391,392,393,394,395,396,397,398,399,400,401,402,403,404,405,406,407,408,409,410,411,412,413,414,415,416,417,418,419]. |
| Ficopomatus enigmaticus (Fauvel, 1923) | France, native range is unknown, likely southerm Australia | Unknown | Non-indigenous | Europe, New Zealand, Japan, USA (both coasts), Argentina, Tunisia, Egypt, Côte d’Ivoire, South Africa | Cytb, COI | Reviewed by Dittmann [420]. Styan et al. [204] revealed three species (not formally described) with overlapping ranges in Australia, one of which is morphologically distinct from the other two. Grosse et al. [421] found two other species within the complex Spain. |
| Ficopomatus uschakovi (Pillai, 1960) | Sri Lanka | Unknown | ? Non-indigenous | Indo-Pacific, Western Africa (Nigeria, Ivory Coast), Brazil, Venezuela, Colombian Caribbean and southern Mexican Pacific ?Australia | none | Several records given outside the native range [422,423,424,425]. Likely a species complex [204]. |
| Hydroides brachyacantha Rioja, 1941 | Pacific coast of Mexico | Warm Temperate Northeast Pacific | Native | Likely restricted to Pacific coast of Mexico, no evidence of translocations | 18S, cytb, ITS | Belongs to a complex of species, records from warm-temperate and tropical localities world-wide likely belong to other species of H. brachacantha complex [203]. |
| Hydroides dianthus (Verrill, 1873) | New England, USA | Either East Coast of the USA or the Mediterranean | Non-indigenous | Brazil, China, Japan, West Africa, the Mediterranean (or US East Coast), the Black Sea, Texas | COI | Observed higher haplotypes diversity in the Mediterranean contradicts the accepted native range of H. dianthus in the USA. The cryptic lineage found in Texas was evidently introduced to the Black Sea recently [250]. |
| Hydroides dirampha Mörch, 1863 | St. Thomas Island, US Virgin Islands | Tropical Northwestern Atlantic | Non-indigenous | Australia, Brazil, Japan, Hawaii, New Zealand, Panama, the Pacific from Mexico and California, and the Mediterranean | 18S, cytb, COI, 28S, ITS | Several records given outside the native range [426,427,428,429]. Molecular data from Australia, Brazil, Panama [78,79]. |
| Hydroides elegans (Haswell, 1883) | Sydney, Australia | Unknown | Non-indigenous | Sub-tropical world-wide: Mediterranean-Atlantic, Indo-West Pacific, tropical Pacific America, West Atlantic, East Atlantic, South Africa | Microsatellites, 18S, cytb, COI, 28S | See [106]. Biofouling has been shown as a major mode of dispersal for this species [430]. Molecular data from Panama, California, Australia, Brazil, Italy and Spain [79,421]. |
| Hydroides ezonesis Okuda, 1934 | Northern Japan | Warm Temperate Northwest Pacific | Non-indigenous | France, UK, Australia | 18S, 28S | Imported from Japan with oysters to the Atlantic coast of France [431,432] then the UK and Australia [359,432,433]. Molecular data from China, Japan [79]. |
| Hydroides operculata (Treadwell, 1929) | Gulf of Aden | Western Indo-Pacific | Native, some species in the complex maybe non-indigenous | A complex of species, no evidence of translocations for any species of the complex | 18S, cytb, COI, 28S, ITS | Reports from South and East Africa, India, Pakistan, Sri Lanka, Hong Kong, tropical Australia, and the eastern Mediterranean likely belong to other species of the H. operculata complex [251]. |
| Hydroides sanctaecrusis Krøyer [in] Mörch, 1863 | Saint Croix, Virgin Islands | Eastern Caribbean | Non-indigenous | Singapore, tropical Australia, Hong Kong, Taiwan, Florida, India. Not found in the Mediterranean [240] | 18S, COI, 28S, ITS | Records outside of the native range include [78,180,362,434]. Molecular data from Florida [77], Panama, Australia, India [79], Pacific Mexico [80]. |
| Spirobranchus kraussii (Baird, 1865) | Cape of Good Hope, South Africa | Agulhas | Native, some species in the complex non-indigenous, but not S. kraussi | Restricted to South Africa, no evidence of translocations | 18S, cytb | Reports from warm-temperate and tropical localities in the Indo-Pacific and Mediterranean Sea belong to other species of S. kraussii complex [84,193]. |
| Spirobranchus tetraceros (Schmarda, 1861) | NSW, Australia | East Central Australian Shelf | Native, some species in the complex likely non-indigenous, but not S. tetraceros | Likely restricted to south-eastern Australia, no evidence of translocations. | cytb | Reports from warm-temperate and tropical localities world-wide belong to other species of the S. tetraceros complex. At least one of these species was introduced and established in the Mediterranean [252]. |
3.6. Fanworms Are Important: Some Applications
3.6.1. Nuisance Fouling Species
3.6.2. Non-Indigenous and Invasive Species
3.6.3. Indicators of Pollution
3.6.4. Bioremeditators
3.6.5. Models Organisms in Research
3.6.6. Objects of Ornamental Trade
3.7. Future Perspectives in Fanworm Research
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Manayunkia siaukhu Annenkova, 1938 inquirenda. Based on the description, M. siaukhu has pygidial eyes [517] and thus does not fulfill diagnostic features for the genus.
- 2.
- Clymeneis Rathke, 1843 inquirenda
- 3.
- Sabella aculeata Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 4.
- Sabella ammonita Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 5.
- Sabella arenaria Montagu, 1803 indeterminable
- 6.
- Sabella arundinacea Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 7.
- Sabella clavata Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 8.
- Sabella compressa Montagu, 1803 indeterminable
- 9.
- Sabella conica Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 10.
- Sabella corticalis Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 11.
- Sabella dimidiata Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 12.
- Sabella flabellata Savigny, 1820 inquirenda
- 13.
- Sabella fixa Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 14.
- Sabella grossa Baird, 1865 inquirenda
- 15.
- Sabella helicina Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 16.
- Sabella nigra Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 17.
- Sabella sabulosa Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 18.
- Sabella setiformis Montagu, 1803 indeterminable
- 19.
- Sabella stagnalis Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 20.
- Sabella subcylindrica Montagu, 1803 indeterminable
- 21.
- Sabella teredula Chiereghini in Siebold, 1850 indeterminable
- 22.
- Sabella trigona Chiereghini in Siebold, 1850 indeterminable
- 23.
- Sabella uncinata Gmelin in Linnaeus, 1788. Insecta: Trichoptera.
- 24.
- Sabella vegetabilis Gmelin in Linnaeus, 1888. Insecta: Trichoptera.
- 25.
- Sabella zonalis Stimpson, 1854 inquirenda
References
- Kupriyanova, E.K.; Rouse, G.W. Yet another example of paraphyly in Annelida: Molecular evidence that Sabellidae contains Serpulidae. Mol. Phylogenet. Evol. 2008, 46, 1174–1181. [Google Scholar] [CrossRef]
- Capa, M.; Hutchings, P.; Aguado, M.T.; Bott, N.J. Phylogeny of Sabellidae (Annelida) and relationships with other taxa inferred from morphology and multiple genes. Cladistics 2011, 27, 449–469. [Google Scholar] [CrossRef]
- Tilic, E.; Sayyari, E.; Stiller, J.; Mirarab, S.; Rouse, G.W. More is needed—thousands of loci are required to elucidate the relationships of the ‘Flowers of the Sea’ (Sabellida, Annelida). Mol. Phylogenet. Evol. 2020, 151, 106892. [Google Scholar] [CrossRef]
- Bick, A. Fabriciidae Rioja, 1923. In Handbook of Zoology Annelida, Volume 3: Pleistoannelida, Sedentaria III and Errantia I; Purschke, G., Westheide, W., Böggemann, M., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2021; pp. 1–33. [Google Scholar]
- Savigny, J.-C. Systèmes des Annélides, Principalement de Celles des Côtes de l’Égypte et de la Syrie, Offrant les Caractères tant Distinctifs que Naturels des Ordres, Familles et Genres, Avec la Description des Espèces Système des Annelides (I); Commission des sciences et arts d’Egypte: Paris, France, 1822; Volume 1, pp. 1–128. [Google Scholar]
- Burmeister, H. Handbuch Der Naturgeschichte. Zum Gebrauch bei Voresungen; Zweite Abt. Zoologie.: Berlin, Germany, 1837; pp. 1–858. [Google Scholar]
- Grube, A.E. Die Familien der Anneliden. Archiv Für Naturgeschichte 1850, 16, 249–364. [Google Scholar]
- Grube, A.E. Beschreibungen Einiger von Georg Ritter von Frauenfeld Gesammelter Anneliden und Gephyreen des Rothen Meeres. Verhandlungen der Kaiserlich Königlichen Zoologisch-Botanischen Gesellschaft in Wien 1868, 18, 629–650. [Google Scholar]
- Malmgren, A.J. Nordiska Hafs-Annulater. Öfversigt af Königlich Vetenskapsakademiens Förhandlingar 1866, 22, 355–410. [Google Scholar]
- Hatschek, B. System Der Anneliden, ein vorläufiger Bericht. Sitzungsberichte des Deutschen Naturwissenschaftlich-Medicinischen Vereines für Böhmen”Lotos“ in Prag. Neue Folge 1893, 13, 123–126. [Google Scholar]
- Dales, R.P. The nature of the pigments in the crown of sabellid and serpulid polychaetes. J. Mar. Biol. Assoc. UK 1962, 42, 259–274. [Google Scholar] [CrossRef]
- Fauchald, K. The polychaete worms. Definitions and keys to the orders, families and genera. Nat. Hist. Mus. LA County Sci. Ser. 1977, 28, 1–188. [Google Scholar]
- Fauchald, K.; Rouse, G.W. Polychaete systematics: Past and present. Zool. Scr. 1997, 26, 71–138. [Google Scholar] [CrossRef]
- Andrade, S.C.S.; Novo, M.; Kawauchi, G.Y.; Worsaae, K.; Pleijel, F.; Giribet, G.; Rouse, G.W. Articulating “Archiannelids”: Phylogenomics and annelid relationships, with emphasis on meiofaunal taxa. Mol. Biol. Evol. 2015, 32, 2860–2875. [Google Scholar] [CrossRef] [PubMed]
- Weigert, A.; Bleidorn, C. Current status of annelid phylogeny. Org. Divers. Evol. 2016, 16, 345–362. [Google Scholar] [CrossRef]
- Helm, C.; Bok, M.J.; Hutchings, P.; Kupriyanova, E.; Capa, M. Developmental studies provide new insights into the evolution of sense organs in Sabellariidae (Annelida). BMC Evol. Biol. 2018, 18, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.P. The development of the sabellid Branchiomma vesiculosum. J. Cell Sci. 1936, 78, 534–603. [Google Scholar]
- Berrill, N. Functional morphology and development of segmental inversion in sabellid polychaetes. Biol. Bull. 1977, 153, 453–467. [Google Scholar] [CrossRef]
- Smith, R.S. Relationships within the Order Sabellida (Polychaeta). Ophelia Suppl. 1991, 5, 249–260. [Google Scholar]
- Bartolomaeus, T. Structure and formation of thoracic and abdominal uncini in Fabricia stellaris (Müller, 1774)–implication for the evolution of Sabellida (Annelida). Zool. Anz. J. Comp. Zool. 2002, 241, 1–17. [Google Scholar] [CrossRef]
- Kieselbach, D.; Hausen, H. Chaetal arrangement provides no support for a close relationship of Sabellidae and Sabellariidae (Annelida). J. Morphol. 2008, 269, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Capa, M.; Hutchings, P.; Peart, R. Systematic revision of Sabellariidae (Polychaeta) and their relationships with other polychaetes using morphological and DNA sequence data: Systematics of Sabellariidae. Zool. J. Linn. Soc. 2012, 164, 245–284. [Google Scholar] [CrossRef]
- Capa, M.; Parapar, J.; Hutchings, P. Phylogeny of Oweniidae (Polychaeta) based on morphological data and taxonomic revision of Australian fauna: Oweniid phylogeny and Australian fauna. Zool. J. Linn. Soc. 2012, 166, 236–278. [Google Scholar] [CrossRef]
- Capa, M.; Giangrande, A.; Nogueira, J.M.M.; Tovar-Hernández, M.A. Sabellidae Latreille, 1825. In Handbook of Zoology, Annelida, Volume 2: Pleistoannelida, Sedentaria II; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany, 2019; pp. 164–212. [Google Scholar]
- Capa, M.; Hutchings, P. Sabellariidae Johnston, 1865. In Handbook of Zoology, Annelida, Volume 2: Pleistoannelida, Sedentaria II; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany, 2019; pp. 144–163. [Google Scholar]
- Capa, M.; Parapar, J.; Hutchings, P.; Mortimer, K. 4. Palaeoannelida. Annelida; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany, 2019; pp. 91–132. ISBN 978-3-11-029158-2. [Google Scholar]
- Huang, D.; Fitzhugh, K.; Rouse, G.W. Inference of phylogenetic relationships within Fabriciidae (Sabellida, Annelida) using molecular and morphological data. Cladistics 2011, 27, 356–379. [Google Scholar] [CrossRef]
- Weigert, A.; Helm, C.; Meyer, M.; Nickel, B.; Arendt, D.; Hausdorf, B.; Santos, S.R.; Halanych, K.M.; Purschke, G.; Bleidorn, C.; et al. Illuminating the base of the annelid tree using transcriptomics. Mol. Biol. Evol. 2014, 31, 1391–1401. [Google Scholar] [CrossRef]
- Struck, T.H.; Golombek, A.; Weigert, A.; Franke, F.A.; Westheide, W.; Purschke, G.; Bleidorn, C.; Halanych, K.M. The evolution of annelids reveals two adaptive routes to the interstitial realm. Curr. Biol. 2015, 25, 1993–1999. [Google Scholar] [CrossRef]
- WoRMS 2020, Sabellida. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=882 (accessed on 2 March 2021).
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine ecoregions of the World: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef]
- Udvardy, M.D. A Classification of the Biogeographical Provinces of the World; IUCN Occasional Paper 18: Morges, Switzerland, 1975; pp. 1–49. [Google Scholar]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C. Terrestrial ecoregions of the world: A new map of life on Earth a new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- GBIF. 2020. Available online: https://www.gbif.org/es/ (accessed on 3 March 2021).
- NCBI. 2020. Available online: https://www.ncbi.nlm.nih.gov (accessed on 3 March 2021).
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Rafinesque, C.S. Analyse de la Nature ou Tableau de l’Universe et Des Corps Organize; De l’imprimerie de Jean Barravecchia: Palermo, Italy, 1815. [Google Scholar]
- Chamberlin, R.V. The Annelida Polychaeta. Mem. Mus. Comp. Zool. 1919, 48, 1–518. [Google Scholar]
- Rioja, E. Estudio sistemático de las especies Ibéricas del Suborden Sabelliformia. Trab. Mus. Nac. Cien. Nat. (Ser. Zool.) 1923, 48, 1–144. [Google Scholar]
- Pillai, G. Studies on a collection of spirorbids from Ceylon, together with a critical review and revision of spirorbid systematics and an account of their phylogeny and zoogeography. Ceylon J. Sci. (Biol. Sci.) 1970, 8, 100–172. [Google Scholar]
- Ten Hove, H.A. Towards a phylogeny in serpulids (Annelida; Polychaeta). In Proceedings of the First International Polychaete Conference; Hutchings, P.A., Ed.; Linnean Society of New South West: Sydney, Australia, 1984; pp. 181–196. [Google Scholar]
- Fitzhugh, K. A systematic revision of the Sabellidae-Caobangiidae-Sabellongidae complex (Annelida: Polychaeta). Bull. Am. Mus. Nat. Hist. 1989, 192, 1–104. [Google Scholar]
- Kupriyanova, E.K. Life history evolution in serpulimorph polychaetes: A phylogenetic analysis. Hydrobiologia 2003, 496, 105–114. [Google Scholar] [CrossRef]
- Macdonald, T.A. Phylogenetic relations among spirorbid subgenera and the evolution of opercular brooding. Hydrobiologia 2003, 496, 125–143. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Macdonald, T.A.; Rouse, G.W. Phylogenetic relationships within Serpulidae (Sabellida, Annelida) inferred from molecular and morphological data. Zool. Scr. 2006, 35, 421–439. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; ten Hove, H.A.; Sket, B.; Zakšek, V.; Trontelj, P.; Rouse, G.W. Evolution of the unique freshwater cave-dwelling tube worm Marifugia cavatica (Annelida: Serpulidae). Syst. Biodivers 2009, 7, 389–401. [Google Scholar] [CrossRef]
- Lehrke, J.; Harry, A.; Macdonald, T.A.; Bartolomaeus, T.; Bleidorn, C. Phylogenetic relationships of Serpulidae (Annelida: Polychaeta) based on 18S rDNA sequence data, and implications for opercular evolution. Org. Divers Evol. 2007, 7, 195–206. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Nishi, E. Serpulidae (Annelida, Polychaeta) from Patton-Murray Seamounts, Gulf of Alaska, North Pacific Ocean. Zootaxa 2010, 2665, 51–68. [Google Scholar] [CrossRef]
- Prentiss, N.K.; Vasileiadou, K.; Faulwetter, S.; Arvanitidis, C.; ten Hove, H.A. A new genus and species of Serpulidae (Annelida, Polychaeta, Sabellida) from the Caribbean Sea. Zootaxa 2014, 3900, 204–222. [Google Scholar] [CrossRef]
- Rzhavsky, A.V.; Kupriyanova, E.K.; Sikorski, A.V. Two new species of serpulid polychaetes (Annelida) from the Barents Sea. Fauna Norv. 2013, 32, 27–38. [Google Scholar] [CrossRef][Green Version]
- Johansson, K.E. Beiträge Zur Kenntnis der Polychaeten-Familien Hermellidae Sabellidae und Serpulidae. Zool. Bidr. Uppsala 1927, 11, 1–184. [Google Scholar]
- Fitzhugh, K. Further revisions of the Sabellidae subfamilies and cladistic relationships among the Fabriciinae (Annelida: Polychaeta). Zool. J. Linn. Soc. 1991, 102, 305–332. [Google Scholar] [CrossRef]
- Fitzhugh, K. Systematics of several fabriciin fan worms (Polychaeta: Sabellidae: Fabriciinae) previously referred to Fabricia or Fabriciola. J. Nat. Hist. 1991, 25, 1101–1120. [Google Scholar] [CrossRef]
- Fitzhugh, K. On the systematic position of Monroika africana (Monro) (Polychaeta: Sabellidae: Fabriciinae) and a description of a new fabriciin genus and species from Australia. Proc. Biol. Soc. Wash. 1992, 105, 116–131. [Google Scholar]
- Fitzhugh, K. Additions to the description of the fanworm genus Pseudofabricia Cantone, 1972 (Polychaeta: Sabellidae: Fabriciinae). Contrib. Sci. 1995, 456, 1–6. [Google Scholar]
- Fitzhugh, K. New fanworm species (Polychaeta: Sabellidae: Fabriciinae) in the genus Pseudofabriciola Fitzhugh. J. Nat. Hist. 1996, 30, 1267–1286. [Google Scholar] [CrossRef]
- Fitzhugh, K. New fan worm genera and species (Polychaeta, Sabellidae, Fabriciinae) from the Western Pacific, and cladistic relationships among genera. Zool. Scr. 1998, 27, 209–245. [Google Scholar] [CrossRef]
- Fitzhugh, K. New species of Fabricinuda Fitzhugh and Pseudofabriciola Fitzhugh (Polychaeta: Sabellidae: Fabriciinae), with an emendation of Pseudofabriciola australiensis (Hartmann-Schröder). J. Nat. Hist. 2002, 36, 893–925. [Google Scholar] [CrossRef]
- Nogueira, J.M.D.M.; Fitzhugh, K.; Rossi, M.C.S. A new genus and new species of fan worms (Polychaeta: Sabellidae) from Atlantic and Pacific Oceans—the formal treatment of taxon names as explanatory hypotheses. Zootaxa 2010, 2603, 1–52. [Google Scholar] [CrossRef]
- Fitzhugh, K.; Giangrande, A.; Simboura, N. New species of Pseudofabriciola Fitzhugh, 1990 (Polychaeta: Sabellidae: Fabriciinae), from the Mediterranean Sea. Zool. J. Linn. Soc. 1994, 110, 219–241. [Google Scholar] [CrossRef]
- Fitzhugh, K. Fabricinuda, a new genus of Fabriciinae (Polychaeta: Sabellidae). Proc. Biol. Soc. Wash. 1990, 103, 161–178. [Google Scholar]
- Fitzhugh, K. Two new genera of the Subfamily Fabriciinae (Polychaeta: Sabellidae). Am. Mus. Novit. 1990, 2967, 1–19. [Google Scholar]
- Fitzhugh, K. A new deep-water genus and species of Fabriciinae fanworm (Polychaeta: Sabellidae) from Antarctica. Contrib. Sci. 2001, 491, 1–8. [Google Scholar]
- Fitzhugh, K. A new species of Megalomma Johansson, 1927 (Polychaeta: Sabellidae: Sabellinae) from Taiwan, with comments on sabellid dorsal lip classification. Zool. Stud. 2003, 42, 106–134. [Google Scholar]
- Rouse, G.; Fitzhugh, K. Broadcasting fables: Is external fertilization really primitive? sex, size, and larvae in sabellid polychaetes. Zool Scr. 1994, 23, 271–312. [Google Scholar] [CrossRef]
- Fitzhugh, K.; Rouse, G.W. A remarkable new genus and species of fan worm (Polychaeta: Sabellidae: Sabellinae) associated with marine gastropods. Invertebr. Biol. 1999, 118, 357–390. [Google Scholar] [CrossRef]
- Cochrane, S. Snowflakes and feather-dusters–some challenges for soft-bottom fanworm systematics. Hydrobiologia 2003, 496, 49–62. [Google Scholar] [CrossRef]
- Capa, M. Taxonomic revision and phylogenetic relationships of apomorphic sabellids (Polychaeta) from Australia. Invertebr. Syst. 2007, 21, 537. [Google Scholar] [CrossRef]
- Capa, M. The genera Bispira Krøyer, 1856 and Stylomma Knight-Jones, 1997 (Polychaeta, Sabellidae): Systematic revision, relationships with close related taxa and new species from Australia. Hydrobiologia 2008, 596, 301–327. [Google Scholar] [CrossRef]
- Capa, M.; Bybee, D.R.; Bybee, S.M. Establishing species and species boundaries in Sabellastarte Krøyer, 1856 (Annelida: Sabellidae): An integrative approach. Org. Divers. Evol. 2010, 10, 351–371. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A. Phylogeny of Chone Krøyer, 1856 (Polychaeta: Sabellidae) and related genera. J. Nat. Hist. 2008, 42, 2193–2226. [Google Scholar] [CrossRef]
- Capa, M.; Pons, J.; Hutchings, P. Cryptic diversity, intraspecific phenetic plasticity and recent geographical translocations in Branchiomma (Sabellidae, Annelida). Zool. Scr. 2013, 42, 637–655. [Google Scholar] [CrossRef]
- Capa, M.; Murray, A. Integrative taxonomy of Parasabella and Sabellomma (Sabellidae: Annelida) from Australia: Description of new species, indication of cryptic diversity, and translocation of some species out of their natural distribution range: Australian Parasabella and Sabellomma. Zool. J. Linn. Soc. 2015, 175, 764–811. [Google Scholar] [CrossRef]
- Capa, M.; Murray, A. Combined morphological and molecular data unveils relationships of Pseudobranchiomma (Sabellidae, Annelida) and reveals higher diversity of this intriguing group of fan worms in Australia, including potentially introduced species. ZooKeys 2016, 622, 1–36. [Google Scholar] [CrossRef]
- Giangrande, A.; Licciano, M. The genus Euchone (Polychaeta, Sabellidae) in the Mediterranean Sea, addition of two new species and discussion on some closely related taxa. J. Nat. Hist. 2006, 40, 1301–1330. [Google Scholar] [CrossRef]
- Licciano, M.; Giangrande, A.; Gambi, M.C. A new genus of Sabellidae (Annelida, Polychaeta) from Antarctica, with discussion of relationships among plesiomorphic genera within Sabellinae. Zootaxa 2009, 2226, 28–42. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Bastida-Zavala, R.; Halt, M.N.; Lee, M.S.Y.; Rouse, G.W. Phylogeny of the Serpula-Crucigera-Hydroides clade (Serpulidae: Annelida) using molecular and morphological data: Implications for operculum evolution. Invertebr. Syst. 2008, 22, 425–437. [Google Scholar] [CrossRef]
- Sun, Y.; Kupriyanova, E.K.; Qiu, J.W. COI Barcoding of Hydroides: A road from impossible to difficult. Invertebr. Syst. 2012, 26, 539–547. [Google Scholar] [CrossRef]
- Sun, Y.; Wong, E.; Ahyong, S.T.; Williamson, J.E.; Hutchings, P.A.; Kupriyanova, E.K. Barcoding and multi-locus phylogeography of the globally distributed calcareous tubeworm genus Hydroides Gunnerus, 1768 (Annelida, Polychaeta, Serpulidae). Mol. Phylogenet. Evol 2018, 127, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Tovar-Hernández, M.A.; Villalobos-Guerrero, T.F.; Kupriyanova, E.K.; Sun, Y. A new fouling Hydroides (Annelida, Sabellida, Serpulidae) from Southern Gulf of California. J. Mar. Biol. Assoc. UK 2016, 96, 693. [Google Scholar] [CrossRef]
- Smith, A.M.; Henderson, Z.E.; Kennedy, M.; King, T.M.; Spencer, H.G. Reef formation versus solitariness in two New Zealand serpulids does not involve cryptic species. Aquat. Biol 2012, 16, 97–103. [Google Scholar] [CrossRef][Green Version]
- Willette, D.A.; Iniguez, A.R.; Kupriyanova, E.K.; Starger, C.J.; Varman, T.; Toha, A.H.; Maralit, B.A.; Barber, P.H. Christmas tree worms of Indo-Pacific coral reefs: Untangling the Spirobranchus corniculatus (Grube, 1862) complex. Coral Reefs 2015, 34, 899–904. [Google Scholar] [CrossRef]
- Perry, O.; Bronstein, O.; Simon-Blecher, N.; Atkins, A.; Kupriyanova, E.; ten Hove, H.; Levy, O.; Fine, M. On the genus Spirobranchus (Annelida, Serpulidae) from the Northern Red Sea, and a description of a new species. Inverterb. Syst. 2018, 32, 605. [Google Scholar] [CrossRef]
- Pazoki, S.; Rahimian, H.; Struck, T.H.; Katouzian, A.R.; Kupriyanova, E.K. A new species of the Spirobranchus kraussii-complex (Annelida, Serpulidae) from the Persian Gulf and Gulf of Oman. Zootaxa 2020, 4748, 401–430. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.H. Methods of brood protection as a basis for reclassification of the Spirorbinae (Serpulidae). Zool. J. Linn. Soc. 1969, 48, 387–407. [Google Scholar] [CrossRef]
- Thorp, C.H. The structure of the operculum in Pileolaria granulata (L.) (Polychaeta: Serpulidae) and related species. J. Exp. Mar. Biol. Ecol. 1975, 20, 215–235. [Google Scholar] [CrossRef]
- Knight-Jones, P.; Thorp, C.H. The opercular brood chambers of Spirorbidae. J. Linn. Soc. (Zool.) 1984, 80, 121–133. [Google Scholar] [CrossRef]
- Nishi, E. On the origin of brooding characteristics in spirorbids with the phylogeny of sabellids and serpulids (Annelida, Polychaeta, Sedentaria). Proc. Jap. Soc. Syst. Zool. 1993, 49, 6–12. [Google Scholar]
- Thorp, C.; Segrove, F. The opercular moult in Spirorbis spirorbis (L.) and S. pusilloides Bush (Polychaeta: Serpulidae). J. Exp. Mar. Biol. Ecol. 1975, 19, 117–143. [Google Scholar] [CrossRef]
- Rzhavsky, A.V.; Kupriyanova, E.K. Evolution of spirorbin brooding: A phylogenetic analysis and a test of an oxygen limitation hypothesis. Invertebr. Zool. 2019, 16, 409–430. [Google Scholar] [CrossRef]
- Pamungkas, J.; Glasby, C.J.; Read, G.B.; Wilson, S.P.; Costello, M.J. Progress and perspectives in the discovery of polychaete worms (Annelida) of the world. Helgol. Mar. Res. 2019, 73, 4. [Google Scholar] [CrossRef]
- Fitzhugh, K. A Revision of the genus Fabricia Blainville, 1828 (Polychaeta: Sabellidae: Fabriciinae). Sarsia 1990, 75, 1–16. [Google Scholar] [CrossRef]
- Cepeda, D.; Lattig, P. New reports and description of a new species of Sabellidae (Annelida) for the Iberian Peninsula and Balearic Archipelago. Mar. Biol. Res. 2017, 13, 832–853. [Google Scholar] [CrossRef]
- Giangrande, A.; Wasson, B.; Lezzi, M.; Licciano, M. Description of Euchone anceps sp. nov. (Annelida: Sabellidae) from the Mediterranean Sea and Northeast Atlantic, with remarks on the difficulty of generic definition. Eur. Zool. J. 2017, 84, 193–207. [Google Scholar] [CrossRef]
- Çinar, M.E.; Giangrande, A. A new species of Pseudobranchiomma (Sabellidae, Polychaeta) from the Sea of Marmara (Turkey). Mar. Biodivers 2018, 48, 1563–1569. [Google Scholar] [CrossRef]
- López, E.; Olivier, F.; Grant, C.; Archambault, P.A. new species and four new records of sedentary polychaetes from the Canadian High Arctic. J. Mar. Biol. Assoc. UK 2017, 97, 1685–1694. [Google Scholar] [CrossRef]
- Nishi, E.; Gil, J.; Tanaka, K.; Kupriyanova, E.K. Notaulax yamasui sp. n. (Annelida, Sabellidae) from Okinawa and Ogasawara, Japan, with notes on its ecology. ZooKeys 2017, 660, 1–16. [Google Scholar] [CrossRef][Green Version]
- Nishi, E.; Tanaka, K.; Tovar-Hernández, M.A. A new species of Claviramus (Annelida, Sabellida, Sabellidae) from the Ariake Inland Sea, Kyushu, Japan. ZooKeys 2019, 880, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Tovar-Hernández, M.A.; de León-González, J.A.; Bybee, D.R. Sabellid worms from the Patagonian Shelf and Humboldt Current System (Annelida, Sabellidae): Phyllis Knight-Jones’ and José María Orensanz’s collections. Zootaxa 2017, 4283, 1–64. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Harry, A.; Vinn, O.; Zatoń, M.; de León-González, J.A.; García-Garza, M.E. Fan worms (Annelida: Sabellidae) from Indonesia collected by the Snellius II Expedition (1984) with descriptions of three new species and tube microstructure. PeerJ 2020, 8, e9692. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; García-Garza, M.E.; de León-González, J.A. Sclerozoan and fouling sabellid worms (Annelida: Sabellidae) from Mexico with the establishment of two new species. Biodivers. Data J. 2020, 8, e57471. [Google Scholar] [CrossRef]
- Bick, A.; Bastrop, R.; Kotta, J.; Meißner, K.; Meyer, M.; Syomin, V. Description of a new species of Sabellidae (Polychaeta, Annelida) from fresh and brackish waters in Europe, with some remarks on the branchial crown of Laonome. Zootaxa 2018, 4483, 349–364. [Google Scholar] [CrossRef]
- Tilic, E.; Feerst, K.G.; Rouse, G.W. Two new species of Amphiglena (Sabellidae, Annelida), with an assessment of hidden diversity in the Mediterranean. Zootaxa 2019, 4648, 337–353. [Google Scholar] [CrossRef]
- Renier, S.A. Osservazioni postume di Zoologia Adriatica, del Professore Stefano Andrea Renier, Membro Effettivo dell’Instituto Italiano, Pubblicate per Cura dell’IR Istituto Veneto Di Scienze, Lettere er Arti a Studio Del Membro Effettivo Prof. G. Meneghini. Giovanni Cecchini: Venice, Italia, 1847; pp. 1–122. [Google Scholar]
- Linnaeus, C. Systema Naturae, 10th ed.; L. Salvius: Holmiae, Sweden, 1758. [Google Scholar]
- Pillai, T.G. A revision of the genera Galeolaria and Pyrgopolon (Polychaeta: Serpulidae), with discussions on opercular insertion as a character in their taxonomy and relationships, and their zoogeography. Zootaxa 2009, 2060, 47–58. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Ippolitov, A.P. Deep-sea serpulids (Annelida: Polychaeta) in tetragonal tubes: On a tube convergence path from the Mesozoic to Recent. Zootaxa 2015, 4044, 151–200. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A. Taxonomic update of the sabellids (Polychaeta: Sabellidae) from Chile and taxa established by prof. Ernst Ehlers, with a key to genera of Sabellinae. An. Inst. Patagon. 2010, 38, 7–29. [Google Scholar] [CrossRef][Green Version]
- Tovar-Hernández, M.A.; Fitzhugh, K. Sabellidae Latreille, 1825. In Poliquetos (Annelida: Polychaeta) de México y América Tropical, 2nd ed.; De León-González, J.A., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Salazar-Vallejo, S.I., Solís-Weiss, V., Tovar-Hernández, M.A., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2021; Volume 3. [Google Scholar]
- Ten Hove, H.A.; Kupriyanova, E.K. Taxonomy of Serpulidae (Annelida, Polychaeta): The state of affairs. Zootaxa 2009, 2036, 1–126. [Google Scholar] [CrossRef]
- Knight-Jones, P.; Darbyshire, T.; Petersen, M.E.; Tovar-Hernández, M. What is Pseudopotamilla reniformis (Sabellidae)? Comparisons of populations from Britain, Iceland and Canada with comments on Eudistylia and Schizobranchia. Zootaxa 2017, 4254, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Jirkov, I. Polikhety Severnogo Ledovitogo Okeana [Polychaeta of the Arctic Ocean.]; Janus-K Press: Moscow, Russia, 2001; pp. 1–632. (In Russian) [Google Scholar]
- Rzhavsky, A.V.; Kupriyanova, E.K.; Sikorski, A.V.; Dahle, S. Calcareous Tubeworms (Polychaeta, Serpulidae) of the Arctic Ocean; KMK Scientific Press: Moscow, Russia, 2014; pp. 1–187. [Google Scholar]
- Rzhavsky, A.; Kupriyanova, E.; Sikorski, A. Field guide to Calcareous Tubeworms (Polychaeta, Serpulidae) of the Arctic Ocean; KMK Scientific Press: Moscow, Russia, 2018; pp. 1–184. [Google Scholar]
- Fauvel, P. Polychètes sédentaires. Addenda aux errantes, Arachiannélides, Myzostomaires. In Faune de France; Paul Lechevalier: Paris, France, 1927; Volume 16, pp. 1–494. [Google Scholar]
- Zibrowius, H. Étude morphologique, systématique et écologique des Serpulidae (Annelida Polychaeta) de La région de Marseille. Recl. Trav. Stn. Mar. Endoume Bull. 1968, 43, 81–252. [Google Scholar]
- Bianchi, C.N. Les espèces de Serpuloidea (Annélides, Polychaètes) des lagunes côtières Italiennes. Rapp. Commis. Int. Mer. Médit. Monaco 1981, 27, 195–196. [Google Scholar]
- Knight-Jones, P. Sabellidae and Serpulidae. In The Marine Fauna of the British Isles and North West Europe; Hayward, P.J., Ryland, J.S., Eds.; Clarendon Press: Oxford, UK, 1990; Volume 1, pp. 271–287. [Google Scholar]
- López, E.; Cepeda, D. Familia Fabriciidae Rioja, 1923. In Fauna Ibérica; Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2020. [Google Scholar]
- Cepeda, D.; López, E. Familia Sabellidae Latreille, 1825. In Fauna Ibérica; Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2020. [Google Scholar]
- Bick, A. Redescription of Fabriciola tonerella Banse, 1959, and a new record of Novafabricia infratorquata (Fitzhugh, 1983) from the Mediterranean Sea, with a key for the Fabriciinae (Annelida: Polychaeta) of the Mediterranean Sea and the North-East Atlantic. Zool. Anz. J. Comp. Zool. 2005, 244, 137–152. [Google Scholar] [CrossRef]
- Bick, A.; Randel, N. Ontogenetic variations in characters of Euchone analis (Krøyer, 1856) (Polychaeta, Sabellidae, Sabellinae) from Spitsbergen, and new assignments of Oriopsis ingelorae Plate, 1995 and O. liefdefjordensis Plate, 1995. Acta Zool. 2005, 86, 145–157. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Licciano, M.; Giangrande, A. Revision of Chone Krøyer, 1856 (Polychaeta: Sabellidae) from the Eastern Central Atlantic and Mediterranean Sea with descriptions of two new species. Sci. Mar. 2007, 71, 315–338. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A. Revision of Chone Krøyer, 1856 (Polychaeta: Sabellidae) from North America and descriptions of four new species. J. Nat. Hist. 2007, 41, 511–566. [Google Scholar] [CrossRef]
- Çinar, M. Serpulid species (Polychaeta: Serpulidae) from the Levantine coast of Turkey (Eastern Mediterranean), with special emphasis on alien species. Aquat. Invasions 2006, 1, 223–240. [Google Scholar] [CrossRef]
- Çinar, M.E. Alien polychaete species (Annelida: Polychaeta) on the Southern coast of Turkey (Levantine Sea, Eastern Mediterranean), with 13 new records for the Mediterranean Sea. J. Nat. Hist. 2009, 43, 2283–2328. [Google Scholar] [CrossRef]
- Çinar, M.E.; Dağli, E.; Şahin, G.K. Checklist of Annelida from the coasts of Turkey. Turk. Zool. Derg. 2014, 38, 734–764. [Google Scholar] [CrossRef]
- Selim, S.A.; Rzhavsky, A.V.; Britayev, T.A. Dialychone and Paradialychone (Polychaeta: Sabellidae) from the Mediterranean Coast of Egypt with description of Dialychone egyptica sp. n. Invertebr. Zool. 2012, 9, 105–114. [Google Scholar] [CrossRef]
- Giangrande, A.; Cosentino, A.; Presti, C.L.; Licciano, M. Sabellidae (Annelida) from the Faro coastal lake (Messina, Ionian Sea), with the first record of the invasive species Branchiomma bairdi along the Italian Coast. Mediterr. Mar. Sci. 2012, 13, 283–293. [Google Scholar] [CrossRef][Green Version]
- Giangrande, A.; Caruso, L.; Mikac, B.; Licciano, M. The genus Megalomma (Annelida: Sabellidae) in the Mediterranean Sea, with description of two new species from Italian and Croatian Coasts. Ital. J. Zool. 2015, 82, 521–534. [Google Scholar] [CrossRef][Green Version]
- Mikac, B.A. Sea of worms: Polychaete checklist of the Adriatic Sea. Zootaxa 2015, 3943, 1–172. [Google Scholar] [CrossRef][Green Version]
- Mikac, B.; Licciano, M.; Jaklin, A.; Iveša, L.; Giangrande, A.; Musco, L. Diversity and distribution patterns of hard bottom polychaete assemblages in the North Adriatic Sea (Mediterranean). Diversity 2020, 12, 408. [Google Scholar] [CrossRef]
- Mikac, B.; Giangrande, A.; Licciano, M. Sabellidae and Fabriciidae (Polychaeta) of the Adriatic Sea with particular retrospect to the Northern Adriatic and the description of two new species. J. Mar. Biol. Assoc. UK 2013, 93, 1511–1524. [Google Scholar] [CrossRef]
- López, E.; Cepeda, D. Catálogo de las especies de Sabellidae Latreille, 1825 y Fabriciidae Rioja, 1923 (Annelida; Sabellida) citadas en la Península Ibérica e Islas Baleares y Chafarinas. Graellsia 2017, 73, e064. [Google Scholar] [CrossRef]
- Kurt Şahin, G.; Çınar, M.E. A check-list of polychaete species (Annelida: Polychaeta) from the Black Sea. J. Black Sea/Medit. Environ. 2012, 18, 10–48. [Google Scholar]
- Hartman, O. Atlas of the Sedentariate Polychaetous Annelids from California; Allan Hancock Foundation, University of Southern California: Los Angeles, CA, USA, 1969; pp. 1–812. [Google Scholar]
- Knight-Jones, P.; Knight-Jones, E.W.; Dales, R.P. Spirorbidae (Polychaeta Sedentaria) from Alaska to Panama. J. Zool. 1979, 189, 419–458. [Google Scholar] [CrossRef]
- Buzhinskaja, G. Polychaetes of the Far East. Seas of Russia and Adjacent Waters of the Pacific Ocean.: Annotated Checklist and Bibliography; KMK Scientific Press: Moscow, Russia, 2013; pp. 1–131. [Google Scholar]
- Nishi, E.; Tanaka, K.; Tovar-Hernández, M.A.; Giangrande, A. Dialychone, Jasmineira and Paradialychone (Annelida: Polychaeta: Sabellidae) from Japan and adjacent waters, including four new species descriptions. Zootaxa 2009, 2167, 1–24. [Google Scholar] [CrossRef]
- Yoshihara, T.; Hiruta, S.; Katoh, T.; Kajihara, H. Three species of Amphicorina (Annelida, Sabellida, Sabellidae) from Japan, with descriptions of two new species. Zookeys 2012, 187, 45–62. [Google Scholar] [CrossRef]
- Imajima, M. Kankeidoubutsu Tamourui (Annelida Polychaeta); Biological Research Co.: Tokyo, Japan, 1996; pp. 1–530. (In Japanese) [Google Scholar]
- Sun, R.; Yang, D. Polychaeta III. Sabellida. In Fauna Sinica: Invertebrata; Science Press: Beijing, China, 2014. [Google Scholar]
- Glasby, C.J.; Lee, Y.-L.; Hsueh, P.-W. Marine Annelida (excluding clitellates and siboglinids) from the South China Sea. Raffles Bull. Zool. 2016, 34, 178–234. [Google Scholar]
- Bastida-Zavala, J.R.; ten Hove, H.A. Revision of Hydroides Gunnerus, 1768 (Polychaeta: Serpulidae) from the Eastern Pacific Region and Hawaii. Beaufortia 2003, 53, 67–110. [Google Scholar]
- Bastida-Zavala, J.R. Serpulids (Annelida: Polychaeta) from the Eastern Pacific, including a brief mention of Hawaiian serpulids. Zootaxa 2008, 1722, 1–61. [Google Scholar]
- Bastida-Zavala, J.R.; McCann, L.D.; Keppel, E.; Ruiz, G.M. The fouling serpulids (Polychaeta: Serpulidae) from United States coastal waters: An overview. Eur. J. Taxon. 2017, 344, 1–76. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Méndez, N.; Villalobos-Guerrero, T.F. Fouling polychaete worms from the Southern Gulf of California: Sabellidae and Serpulidae. Syst. Biodivers 2009, 7, 319–336. [Google Scholar] [CrossRef]
- Bastida-Zavala, J.R.; Buelna, A.S.R.; De León-González, J.A.; Camacho-Cruz, K.A.; Carmona, I. New records of sabellids and serpulids (Polychaeta: Sabellidae, Serpulidae) from the Tropical Eastern Pacific. Zootaxa 2016, 4184, 401–457. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Carrera-Parra, L.F. Megalomma Johansson, 1925 (Polychaeta: Sabellidae) from America and other world-wide localities, and phylogenetic relationships within the genus. Zootaxa 2011, 2861, 1–71. [Google Scholar] [CrossRef]
- Zibrowius, H. Contribution a l’etude des Serpulidae (Polychaeta Sedentaria) du Bresil. Bol. Inst. Oceanogr. 1970, 19, 1–32. [Google Scholar] [CrossRef]
- Ten Hove, H.A. Serpulinae (Polychaeta) from the Caribbean: I-the genus Spirobranchus. Stud. Fauna Curaçao 1970, 32, 1–57. [Google Scholar]
- Ten Hove, H.A. Serpulinae (Polychaeta) from the Caribbean: II-the genus Sclerostyla. Stud. Fauna Curaçao 1973, 43, 1–21. [Google Scholar]
- ten Hove, H.A. Serpulinae (Polychaeta) from the Caribbean: III-the genus Pseudovermilia. Stud. Fauna Curaçao 1975, 47, 46–101. [Google Scholar]
- Ten Hove, H.A. Serpulinae (Polychaeta) from the Caribbean: IV Pseudovermilia madracicola sp. n., a symbiont of corals. In Natuurwetenschappelijke Studiekring voor Suriname en de Nederlandse Antillen; Van Der Steen, L.J., Ed.; 1989; Volume 123, pp. 135–144. [Google Scholar]
- Giangrande, A.; Licciano, M.; Gambi, M.C. A collection of Sabellidae (Polychaeta) from Carrie Bow Cay (Belize, Western Caribbean Sea) with the description of two new species. Zootaxa 2007, 1650, 41–53. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A. Redescription of Chone americana Day, 1973 (Polychaeta: Sabellidae) and description of five new species from the Grand Caribbean Region. Zootaxa 2005, 1070, 1–30. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Knight-Jones, P. Species of Branchiomma (Polychaeta: Sabellidae) from the Caribbean Sea and Pacific Coast of Panama. Zootaxa 2006, 1189, 1–37. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Salazar-Vallejo, S.I. Sabellids (Polychaeta: Sabellidae) from the Grand Caribbean. Zool. Stud. 2006, 45, 24–66. [Google Scholar]
- Bastida-Zavala, J.R.; ten Hove, H.A. Revision of Hydroides Gunnerus, 1768 (Polychaeta: Serpulidae) from the Western Atlantic Region. Beaufortia 2002, 52, 103–173. [Google Scholar]
- Bastida-Zavala, R. Serpula and Spiraserpula (Polychaeta, Serpulidae) from the Tropical Western Atlantic and Gulf of Guinea. ZooKeys 2012, 198, 1–23. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nogueira, J.; Knight-Jones, P.A. New species of Pseudobranchiomma Jones (Polychaeta: Sabellidae) found amongst Brazilian coral, with a redescription of P. punctata (Treadwell, 1906) from Hawaii. J. Nat. Hist. 2002, 36, 1661–1670. [Google Scholar] [CrossRef]
- Nogueira, J.M.D.M.; Abbud, A. Three new serpulids (Polychaeta: Serpulidae) from the Brazilian Exclusive Economic Zone. Zoosymposia 2009, 2, 201–227. [Google Scholar] [CrossRef]
- Amaral, A.C.Z.; Nallin, S.A.H.; Steiner, T.M.; Forroni, T.D.O.; Gomes-Filho, D. Catálogo das Espécies de Annelida Polychaeta do Brasil; Unicamp: Campinas, Brazil, 2013; pp. 1–159. [Google Scholar]
- Costa, D. de A.; Fernandes, H.F.; da Silva, F. de A.; Christoffersen, M.L. Checklist de espécies de Polychaeta (Annelida) da Praia do Seixas, João Pessoa, Estado Da Paraíba, Nordeste do Brasil. Rev. Bras. Gestão Amb. Sust. 2017, 4, 313–320. [Google Scholar] [CrossRef][Green Version]
- Bastida-Zavala, J.R.; Sánchez Ovando, J.P. Serpulidae Rafinesque 1815. In Poliquetos (Annelida: Polychaeta) de México y América Tropical, 2nd ed.; De León-González, J.A., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Salazar-Vallejo, S.I., Solís-Weiss, V., Tovar-Hernández, M.A., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2021; Volume 3. [Google Scholar]
- Fauvel, P. Annelida Polychaeta. In Fauna of India, Pakistan, Ceylon, Burma and Malaya; The Indian Press Ltd: Allahabad, India, 1953; pp. 1–507. [Google Scholar]
- Sivadas, S.K.; Carvalho, R. Marine Annelida of India: Taxonomy and status evaluation and an updated checklist. J. Threat. Taxa 2020, 12, 16647–16714. [Google Scholar]
- Pillai, G. Some marine and brackish-water serpulid polychaetes from Ceylon, including new genera and species. Ceylon J. Sci. (Biol. Sci.) 1960, 3, 1–40. [Google Scholar]
- Pillai, T.G. Annelida Polychaeta of Tambalagam Lake, Ceylon. Ceylon J. Sci. (Biol. Sci.) 1961, 4, 1–40. [Google Scholar]
- Wehe, T.; Fiege, D. Annotated checklist of the polychaete species of the seas surrounding the Arabian Peninsula: Red Sea, Gulf of Aden, Arabian Sea, Gulf of Oman, Arabian Gulf. Fauna Arabia 2002, 19, 7–238. [Google Scholar]
- Al-Kandari, M.; Sattari, Z.; Hussain, S.; Radashevsky, V.I.; Zhadan, A. Checklist of intertidal Polychaetes (Annelida) of Kuwait, Northern Part of the Arabian Gulf. Reg. Stud. Mar. Sci. 2019, 32, 100872. [Google Scholar] [CrossRef]
- Ben-Eliahu, M.N.; ten Hove, H.A. Serpulidae (Annelida: Polychaeta) from the Suez Canal—from a Lessepsian migration perspective (a monograph). Zootaxa 2011, 2848, 1–147. [Google Scholar] [CrossRef]
- Knight-Jones, E.W.; Knight-Jones, P.; Oliver, P.G.; Mackie, A.S.Y. A.S.Y. A new species of Hyalopomatus (Serpulidae: Polychaeta) which lacks an operculum: Is this an adaptation to low oxygen. In Interactions and Adaptation Strategies of Marine Organisms; Naumov, A.D., Hummel, H., Sukhotin, A.A., Ryland, J.S., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 145–151. [Google Scholar]
- Veron, J.E.; Devantier, L.M.; Turak, E.; Green, A.L.; Kininmonth, S.; Stafford-Smith, M.; Peterson, N. Delineating the Coral Triangle. Galaxea 2009, 11, 91–100. [Google Scholar] [CrossRef]
- Fitzhugh, K. Fan worm polychaetes (Sabellidae: Sabellinae) collected during the Thai-Danish BIOSHELF Project. Phuket Mar. Biol. Center Spec. Publ. 2002, 24, 353–424. [Google Scholar]
- Al-Hakim, I.; Glasby, C.J. Polychaeta (Annelida) of the Natuna Islands, South China Sea. Raffles Bull. Zool. 2004, 21, 25–45. [Google Scholar]
- Sun, Y.; ten Hove, H.A.; Qiu, J.W. Serpulidae (Annelida: Polychaeta) from Hong Kong. Zootaxa 2012, 3424, 1–42. [Google Scholar] [CrossRef]
- Murray, A.; Rouse, G.W. Two new species of Terebrasabella (Annelida: Sabellidae: Sabellinae) from Australia. Zootaxa 2007, 1434, 51–68. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Sun, Y.; ten Hove, H.A.; Wong, E.; Rouse, G.W. Serpulidae (Annelida) of Lizard Island, Great Barrier Reef, Australia. Zootaxa 2015, 4019, 275–353. [Google Scholar] [CrossRef]
- Sun, Y.; Wong, E.; ten Hove, H.A.; Hutchings, P.A.; Williamson, J.E.; Kupriyanova, E.K. Revision of the genus Hydroides (Annelida: Serpulidae) from Australia. Zootaxa 2015, 4009, 1–99. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.S.; Hutchings, P.A.; Glasby, C.J. Polychaetes: An. Interactive Identification Guide; CSIRO Publishing: Collingwood, VIC, Australia, 2003; pp. 1–24. [Google Scholar]
- Kupriyanova, E.K.; Wong, E.; Hutchings, P.A. Invasive Polychaete Identifier–an Australian Perspective. Version 1.1 2013. Available online: http://polychaetes.australianmuseum.net.au/ (accessed on 3 March 2021).
- Kupriyanova, E.; Hutchings, P.; Wong, E. A fully illustrated web-based guide to distinguish native and introduced polychaetes of Australia. Manag. Biol. Invasions 2016, 7, 305–312. [Google Scholar] [CrossRef]
- Bailey-Brock, J.H.; Magalhães, W.F. A new species and record of Serpulidae (Annelida: Polychaeta) from Cross Seamount in the Hawaiian Chain. Zootaxa 2012, 3192, 49–58. [Google Scholar] [CrossRef]
- Keppel, E.; Tovar-Hernández, M.A.; Ruiz, G. New records of the non-indigenous species Branchiomma bairdi and B. conspersum (Polychaeta: Sabellidae) on the Pacific Coast of North America. Bioinvasions Rec. 2018, 7, 229–236. [Google Scholar] [CrossRef]
- Bailey-Brock, J.H.; Magalhães, W.F.; Brock, R.E. Coral reef inhabiting tubeworms (Polychaeta: Serpulidae) from Enewetak, Kwajalein, Rongelap and Utirik Atolls, Marshall Islands. J. Mar. Biol. Assoc. UK 2012, 92, 967–988. [Google Scholar] [CrossRef]
- Capa, M.; López, E. Sabellidae (Annelida: Polychaeta) living in blocks of dead coral in the Coiba National Park, Panama. J. Mar. Biol. Assoc. UK 2004, 84, 63–72. [Google Scholar] [CrossRef]
- Yáñez-Rivera, B.; Tovar-Hernández, M.A.; Galván-Villa, C.M.; Ríos-Jara, E. Tubicolous polychaete worms (Annelida) from Bahía de Chamela Islands Sanctuary, Mexico, with the description of a new bamboo worm. Biodivers. Data J. 2020, 8, e57572. [Google Scholar] [CrossRef]
- Keppel, E.; Keith, I.; Ruiz, G.M.; Carlton, J.T. New records of native and non-indigenous Polychaetes (Annelida: Polychaeta) in the Galapagos Islands. Aquat. Invasions 2019, 14, 59–84. [Google Scholar] [CrossRef]
- Elías, R.; Jaubet, M.L.; Ferrando, A.; Saracho, M. Historia y perspectivas de los estudios sobre poliquetos en Argentina. In Poliquetos de Sudamérica; Díaz-Díaz, O., Bone, D., Rodríguez, C.T., Delgado-Blas, V.H., Eds.; Poliquetos de Sudamérica. Volumen especial del Boletín del Instituto Oceanográfico de Venezuela: Cumaná, Venezuela, 2017; pp. 3–23. [Google Scholar]
- Rozbaczylo, O.; Moreno, R.A.; Díaz-Díaz, Ó. Poliquetos Bentónicos en Chile. In Poliquetos de Sudamérica; Díaz-Díaz, Ó., Bone, D., Rodríguez, C.T., Delgado-Blas, V.H., Eds.; Volumen especial del Boletín del Instituto Oceanográfico de Venezuela: Cumaná, Venezuela, 2017; pp. 51–70. [Google Scholar]
- Kupriyanova, E.K.; ten Hove, H.A.; Nishi, E.A. Taxonomic revision of Pseudochitinopoma Zibrowius, 1969 (Annelida, Serpulidae) with description of two new species. Zootaxa 2012, 3507, 57–78. [Google Scholar] [CrossRef]
- Simon, C.A.; van Niekerk, H.H.; Burghardt, I.; ten Hove, H.A.; Kupriyanova, E.K. Not out of Africa: Spirobranchus kraussii (Baird, 1865) is not a global fouling and invasive serpulid of Indo-Pacific Origin. Aquat. Invasions 2019, 14, 221–249. [Google Scholar] [CrossRef]
- Day, J.H. A Monograph on the Polychaeta of Southern Africa. Part II. Brit. Mus. Nat. Hist. Publ. 1967, 656, 459–878. [Google Scholar]
- Knight-Jones, P. New species and a new subgenus of Spirorbinae (Serpulidae: Polychaeta) from Kenya. J. Zool. 1972, 166, 1–18. [Google Scholar] [CrossRef]
- Knight-Jones, P.; Knight-Jones, E.W. Spirorbinae (Polychaeta: Serpulidae) from South Africa, including three new species. Mar. Biol. 1974, 25, 253–261. [Google Scholar] [CrossRef]
- Knight-Jones, E.W.; Knight-Jones, P.; Llewellyn, L.C. Spirorbinae (Polychaeta: Serpulidae) from Southeastern Australia. Notes on their taxonomy, ecology and distribution. Rec. Aust. Mus. 1974, 29, 107–151. [Google Scholar] [CrossRef][Green Version]
- Vine, P.J. The marine fauna of New Zealand. Spirorbinae (Polychaeta: Serpulidae). NZOI Mem. 1977, 68, 1–66. [Google Scholar]
- Glasby, C.J.; Read, G.B. A chronological review of polychaete taxonomy in New Zealand. J. R. Soc. N. Z. 1998, 28, 347–374. [Google Scholar] [CrossRef]
- Capa, M.; Rouse, G.W. Phylogenetic relationships within Amphiglena Claparède, 1864 (Polychaeta: Sabellidae), description of five new species from Australia, a new species from Japan, and comments on previously described species. J. Nat. Hist. 2007, 41, 327–356. [Google Scholar] [CrossRef]
- Capa, M.; Murray, A. Review of the genus Megalomma (Polychaeta: Sabellidae) in Australia with description of three new species, new records and notes on certain features with phylogenetic implications. Rec. Aust. Mus. 2009, 61, 201–224. [Google Scholar] [CrossRef][Green Version]
- Murray, A.; Keable, S.J. First report of Sabella spallanzanii (Gmelin, 1791) (Annelida: Polychaeta) from Botany Bay, New South Wales, a northern range extension for the invasive species within Australia. Zootaxa 2013, 3670, 391–395. [Google Scholar] [CrossRef][Green Version]
- Sun, Y.; Wong, E.; Tovar-Hernández, M.A.; Williamson, J.E.; Kupriyanova, E.K. Is Hydroides brachyacantha (Serpulidae: Annelida) a widespread species? Invertebr. Syst. 2016, 30, 41–59. [Google Scholar] [CrossRef]
- Styan, C.; McCluskey, C.; Sun, Y.; Kupriyanova, E. Cryptic sympatric species across the Australian range of the global estuarine invader Ficopomatus enigmaticus (Fauvel, 1923) (Serpulidae, Annelida). Aquat. Invasions 2017, 12, 53–65. [Google Scholar] [CrossRef]
- Baird, W. Description of a new species of annelids belonging to the Family Amphinomidae. Trans. Linn. Soc. London 1864, 24, 449–450. [Google Scholar] [CrossRef]
- Baird, W. Description of several new species and varieties of tubicolous annelids (= Tribe Limivora of Grube), in the collection of the British Museum. J. Linn. Soc. Zool. 1865, 8, 10–22. [Google Scholar] [CrossRef]
- Ehlers, E. Polychaeten der Hamburger Magalhaenischen Sammelreise. Ergeb. Hambg. Magalhaenischen Sammelreise Liefersung 1897, 2, 1–147. [Google Scholar]
- Gravier, C. Sur les Annélides Polychètes Recueillies par l’Expédition Antarctique Française (Terebelliens, Serpuliens). Bull. Mus. d’Hist. Nat. Paris 1907, 13, 46–52. [Google Scholar]
- Pixell, H.L.M. Polychaeta of the Families Serpulidae and Sabellidae, collected by the Scottish National Antarctic Expedition. Trans. R. Soc. Edinburgh 1913, 49, 347–358. [Google Scholar] [CrossRef]
- Benham, W.B. Polychaeta. British Antarctic “Terra Nova” Expedition, 1910. Nat. Hist. Rep. Zool. 1927, 7, 47–182. [Google Scholar]
- Rzhavsky, A.V. Two new species of Pileolaria (Polychaeta: Spirorbidae) from the Southern hemisphere with a brief review of related species. Inverterb. Zool. 2010, 7, 81–91. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Yáñez-Rivera, B.; Giangrande, A.; Gambi, M.C. Notes on the species of Perkinsiana (Polychaeta: Sabellidae) from Antarctica with the description of P. brigittae sp. nov. Zootaxa 2012, 3485, 56–68. [Google Scholar] [CrossRef]
- Neal, L.; Paterson, G.L.; Blockley, D.; Scott, B.; Sherlock, E.; Huque, C.; Glover, A.G. Biodiversity data and new species descriptions of polychaetes from offshore waters of the Falkland Islands, an area undergoing hydrocarbon exploration. ZooKeys 2020, 938, 1–86. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Rzhavsky, A.; ten Hove, H.A. Serpulidae Rafinesque, 1815. In Handbook of Zoology, Annelida, Volume 2: Pleistoannelida, Sedentaria II; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany, 2019; pp. 213–275. [Google Scholar]
- Jones, M.L. Brandtika asiatica new genus, new species, from Southeastern Asia and a redescription of Monroika africana (Monro) (Polychaeta: Sabellidae). Proc. Biol. Soc. Wash. 1974, 87, 217–230. [Google Scholar]
- Atkinson, S.D.; Bartholomew, J.L.; Rouse, G.W. The invertebrate host of salmonid fish parasites Ceratonova shasta and Parvicapsula minibicornis (Cnidaria: Myxozoa), is a novel fabriciid annelid, Manayunkia occidentalis sp. nov. (Sabellida: Fabriciidae). Zootaxa 2020, 4751, 310–320. [Google Scholar] [CrossRef]
- Sitnikova, T.Y.; Shcherbakov, D.Y.; Kharchenko, V.V. On taxonomic status of polychaetes of the genus Manayunkia (Sabellidae, Fabriciinae) from the open Lake Baikal. Zool. Zhurnal 1997, 76, 16–27. [Google Scholar]
- Monro, C.C.A. On a collection of Polychaeta from near the Mouth of the River Congo. Rev. Zool. Bot. Afr. 1939, 32, 213–225. [Google Scholar]
- Rouse, G. New Fabriciola and Manayunkia species (Fabriciinae: Sabellidae: Polychaeta) from Papua New Guinea. J. Nat. Hist. 1996, 30, 1761–1778. [Google Scholar] [CrossRef]
- Baumhaker, H. Taxonomische Untersuchungen an Sabellidae und Fabriciidae (Annelida: Polychaeta) der Tiefsee des Südwest-Atlantiks. Master’s Thesis, University of Rostock, Rostock, Germany, 2012. [Google Scholar]
- Bok, M.J.; Capa, M.; Nilsson, D.-E. Here, there and everywhere: The radiolar eyes of fan worms (Annelida, Sabellidae). Integr. Comp. Biol. 2016, 56, 784–795. [Google Scholar] [CrossRef]
- Brown, S.D.; Emlet, R.B. Natural radiole damage and regeneration in the feather duster worm Schizobranchia insignis. Invertebr. Biol. 2020, e12307. [Google Scholar] [CrossRef]
- Orrhage, L. On the structure and homologues of the anterior end of the polychaete families Sabellidae and Serpulidae. Zoomorphology 1980, 96, 113–167. [Google Scholar] [CrossRef]
- Giangrande, A. The genus Chone (Polychaeta, Sabellidae) in the Mediterranean Sea with description of C. longiseta n. sp. Boll. Zool. 1992, 59, 517–529. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Sosa-Rodríguez, T. Redescription of Chone infundibuliformis Krøyer, 1856 (Polychaeta: Sabellidae) and histology of the branchial crown appendages, collar and glandular ridge. Zootaxa 2006, 1115, 31–59. [Google Scholar] [CrossRef]
- Capa, M.; Nogueira, J.M.D.M.; Rossi, M.C.S. Comparative internal structure of dorsal lips and radiolar appendages in Sabellidae (Polychaeta) and phylogenetic implications. J. Morphol. 2011, 272, 302–319. [Google Scholar] [CrossRef] [PubMed]
- Ippolitov, A.P.; Vinn, O.; Kupriyanova, E.K.; Jäger, M. Written in stone: History of serpulid polychaetes through time. Mem. Mus. Vic. 2014, 71, 123–160. [Google Scholar] [CrossRef]
- Wong, E.; Kupriyanova, E.K.; Hutchings, P.; Capa, M.; Radashevsky, V.I.; ten Hove, H.A. A graphically illustrated glossary of polychaete terminology: Invasive species of Sabellidae, Serpulidae and Spionidae. Mem. Mus. Vic. 2014, 71, 327–342. [Google Scholar] [CrossRef][Green Version]
- Handley, S.J. Spionid polychaetes in Pacific oysters, Crassostrea gigas (Thunberg) from Admiralty Bay, Marlborough Sounds, New Zealand. N. Z. J. Mar. Freshw. Res. 1995, 29, 305–309. [Google Scholar] [CrossRef][Green Version]
- Bick, A. Morphology and ecology of Dipolydora armata (Polychaeta, Spionidae) from the Western Mediterranean Sea. Acta Zool. 2001, 11, 177–187. [Google Scholar] [CrossRef]
- Bick, A. Polychaete communities associated with gastropod shells inhabited by the hermit crabs Clibanarius erythropus and Calcinus tubularis from Ibiza, Mediterranean Sea. J. Mar. Biol. Assoc. UK 2006, 86, 83–92. [Google Scholar] [CrossRef]
- Rouse, G.W.; Pleijel, F. Polychaetes; Oxford University Press: London, UK, 2001; pp. 1–354. [Google Scholar]
- Bick, A. Bedeutung Postlarvaler Ontogenetischer Merkmalsvariationen für die Taxonomie der Spionidae und Sabellidae; Habilitationsschrift an der Mathematisch-Naturwissenschaftliche Fakultät Fakultät der Universität Rostock: Rostock, Germany, 2007. [Google Scholar]
- Costa-Paiva, E.M.; Paiva, P.C. Anaesthetization and fixation effects on the morphology of sabellid polychaetes (Annelida: Polychaeta: Sabellidae). J. Mar. Biol. Assoc. UK 2007, 87, 1127–1132. [Google Scholar] [CrossRef]
- Coleman, C.O. “Digital inking”: How to make perfect line drawings on computers. Org. Div. Evol. Electr. Suppl. 2003, 14, 1–14. [Google Scholar] [CrossRef]
- Paterson, G.L.; Sykes, D.; Faulwetter, S.; Merk, R.; Ahmed, F.; Hawkins, L.E.; Dinley, J.; Ball, A.D.; Arvanitidis, C. The pros and cons of using micro-computed tomography in gross and micro-anatomical assessments of polychaetous annelids. Mem. Mus. Vic. 2014, 71, 237–246. [Google Scholar] [CrossRef]
- Parapar, J.; Candás, M.; Cunha-Veira, X.; Moreira, J. Exploring annelid anatomy using micro-computed tomography: A taxonomic approach. Zool. Anz. 2017, 270, 19–42. [Google Scholar] [CrossRef]
- Patti, F.P.; Gambi, M.C.; Giangrande, A. Preliminary study on the systematic relationships of Sabellinae (Polychaeta, Sabellidae), based on the C1 domain of the 28S rDNA, with discussion of reproductive features. Ital. J. Zool. 2003, 70, 269–278. [Google Scholar] [CrossRef]
- Halanych, K.M. A review of molecular markers used for annelid phylogenetics. Integr. Comp. Biol. 2006, 46, 533–543. [Google Scholar] [CrossRef]
- Carr, C.M.; Hardy, S.M.; Brown, T.M.; Macdonald, T.A.; Hebert, P.D.N. A Tri-Oceanic perspective: DNA barcoding reveals geographic structure and cryptic diversity in Canadian polychaetes. PLoS ONE 2011, 6, e22232. [Google Scholar] [CrossRef] [PubMed]
- Seixas, V.C.; de Moraes Russo, C.A.; Paiva, P.C. Mitochondrial genome of the Christmas Tree Worm Spirobranchus giganteus (Annelida: Serpulidae) reveals a high substitution rate among annelids. Gene 2017, 605, 43–53. [Google Scholar] [CrossRef]
- Sun, Y.; Daffe, G.; Zhang, Y.; Qiu, J.-W.; Kupriyanova, E.K. Another blow to the conserved gene order in Annelida: Mitogenomics reveals extensive gene rearrangement in the tubeworm genus Hydroides (Serpulidae). Mol. Phylogenet. Evol. 2021, 107124. [Google Scholar] [CrossRef] [PubMed]
- Tilic, E.; Atkinson, S.D.; Rouse, G.W. Mitochondrial genome of the freshwater annelid Manayunkia occidentalis (Sabellida: Fabriciidae). Mitochondrial DNA B 2020, 5, 3313–3315. [Google Scholar] [CrossRef] [PubMed]
- Daffe, G.; Sun, Y.; Ahyong, S.T.; Kupriyanova, E.K. Mitochondrial genome of Sabella spallanzanii (Sabellida: Sabellidae). Mitochondrial DNA B 2020, 6, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; McDougall, C.; Troscianko, J.; Chen, W.-C.; Jayaraman-Nagarajan, A.; Shimeld, S.M.; Ferrier, D.E. An EST screen from the annelid Pomatoceros lamarckii reveals patterns of gene loss and gain in animals. BMC Evol. Biol. 2009, 9, 1–17. [Google Scholar] [CrossRef]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef]
- Barraclough, T.G. Evolving entities: Towards a unified framework for understanding diversity at the species and higher levels. Phil. Trans. R. Soc. B Biol. Sci. 2010, 365, 1801–1813. [Google Scholar] [CrossRef] [PubMed]
- Nygren, A. Cryptic polychaete diversity: A review. Zool. Scr. 2014, 43, 172–183. [Google Scholar] [CrossRef]
- Halt, M.N.; Kupriyanova, E.K.; Cooper, S.J.B.; Rouse, G.W. Naming species with no morphological indicators: Species status of Galeolaria caespitosa (Annelida: Serpulidae) inferred from nuclear and mitochondrial gene sequences and morphology. Invertebr. Syst. 2009, 23, 205–222. [Google Scholar] [CrossRef]
- Sun, Y.; Wong, E.; Keppel, E.; Williamson, J.E.; Kupriyanova, E.K. A global invader or a complex of regionally distributed species? clarifying the status of an invasive calcareous tubeworm Hydroides dianthus (Verrill, 1873) (Polychaeta: Serpulidae) Using DNA Barcoding. Mar. Biol. 2017, 164, 28-1–28-12. [Google Scholar] [CrossRef]
- Sun, Y.; Al-Kandari, M.; Kubal, P.; Walmiki, N.; Kupriyanova, E.K. Cutting a Gordian knot of tubeworms with DNA data: The story of the Hydroides operculata-complex (Annelida, Serpulidae). Zootaxa 2017, 4323, 39–48. [Google Scholar] [CrossRef]
- Palero, F.; Torrado, H.; Perry, O.; Kupriyanova, E.K.; Ulman, A.; Genis-Armero, R.; ten Hove, H.A.; Capaccioni-Azzati, R. Following the Phoenician example: Western Mediterranean colonization by Spirobranchus cf. tetraceros (Annelida: Serpulidae). Sci. Mar. 2020, 84, 83–92. [Google Scholar] [CrossRef]
- Hebert, P.D.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. London Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef]
- Rech, S.; Borrell Pichs, Y.J.; García-Vazquez, E. Anthropogenic marine litter composition in coastal areas may be a predictor of potentially invasive rafting fauna. PLoS ONE 2018, 13, e0191859. [Google Scholar] [CrossRef] [PubMed]
- Langeneck, J.; Lezzi, M.; Del Pasqua, M.; Musco, L.; Gambi, M.C.; Castelli, A.; Giangrande, A. Non-indigenous polychaetes along the coasts of Italy: A critical review. Mediterr. Mar. Sci. 2020, 21, 238–275. [Google Scholar] [CrossRef]
- Rech, S.; Salmina, S.; Pichs, Y.J.B.; García-Vazquez, E. Dispersal of alien invasive species on anthropogenic litter from European mariculture areas. Mar. Poll. Bull. 2018, 131, 10–16. [Google Scholar] [CrossRef]
- Hutchings, P.; De Decker, P.; Geddes, M.C. A new species of Manayunkia (Polychaeta) from ephemeral lakes near the Coorong, South Australia. Trans. R. Soc. S. Aust. 1981, 105, 25–28. [Google Scholar]
- Schütz, L. Über Verbreitung, Ökologie und Biologie des Brackwasserpolychaeten Manayunkia aestuarina (Bourne) Insbesondere an den Küsten Schleswig-Holsteins. Faun. Mitt. Norddeut. 1965, 9, 226–234. [Google Scholar]
- Lewis, D.B. Some aspects of the ecology of Fabricia sabella (Ehr.) (Annelida, Polychaeta). J. Linn. Soc. (Zool.) 1968, 47, 515–526. [Google Scholar] [CrossRef]
- Light, W.J. Extension of range for Manayunkia aestuarina (Polychaeta: Sabellidae) to British Columbia. J. Fish. Board Canada 1969, 26, 3088–3091. [Google Scholar] [CrossRef]
- Bagheri, E.; McLusky, D. Population dynamics of oligochaetes and small polychaetes in the polluted forth estuary ecosystem. Neth. J. Sea Res. 1982, 16, 55–66. [Google Scholar] [CrossRef]
- Bick, A. Reproduction and larval development of Manayunkia aestuarina (Bourne, 1883) (Polychaeta, Sabellidae) in a coastal region of the Southern Baltic. Helgoländer Meeresunters 1996, 50, 287–298. [Google Scholar] [CrossRef]
- Giangrande, A.; Gambi, M.; Micheli, F.; Kroeker, K. Fabriciidae (Annelida, Sabellida) from a naturally acidified coastal system (Italy) with description of two new species. J. Mar. Biol. Assoc. UK 2014, 94, 1417. [Google Scholar] [CrossRef]
- Muus, B.J. The Fauna of Danish estuaries and lagoons: Distribution and ecology of dominating species in the shallow reaches of the mesohaline zone. Medd. Danmarks Fisk. Havunders (n. ser.) 1967, 5, 1–316. [Google Scholar]
- Lewis, D.B. Feeding and tube-building in the Fabriciinae (Annelida, Polychaeta). Proc. Linn. Soc. Lond. 1968, 179, 37–49. [Google Scholar] [CrossRef]
- Hartman, O. Fabricinae (Featherduster Polychaetous Annelids) in the Pacific. Pac. Sci. 1951, 5, 379–391. [Google Scholar]
- Bartholomew, J.L.; Atkinson, S.D.; Hallett, S.L. Involvement of Manayunkia speciosa (Annelida: Polychaeta: Sabellidae) in the life cycle of Parvicapsula minibicornis, a myxozoan parasite of Pacific salmon. J. Parasitol. 2006, 92, 742–748. [Google Scholar] [CrossRef]
- Alexander, J.D.; Hallett, S.L.; Stocking, R.W.; Xue, L.; Bartholomew, J.L. Host and parasite populations after a ten-year flood: Manayunkia speciosa and Ceratonova (Syn Ceratomyxa) shasta in the Klamath River. Northwest. Sci. 2014, 88, 219–233. [Google Scholar] [CrossRef]
- Giangrande, A.; Gambi, M.C. The genus Perkinsiana (Polychaeta, Sabellidae) from Antarctica, with descriptions of the new species P. milae and P. borsibrunoi. Zool. Scr. 1997, 26, 267–278. [Google Scholar] [CrossRef]
- Murray, J.; Watson, G.; Giangrande, A.; Licciano, M.; Bentley, M. Regeneration as a novel method to culture marine ornamental sabellids. Aquaculture 2013, 410, 129–137. [Google Scholar] [CrossRef]
- Chughtai, I.; Knight-Jones, W.E. Burrowing into limestone by sabellid polychaetes. Zool. Scr. 1988, 17, 231–238. [Google Scholar] [CrossRef]
- Fonseca, E.A.; Dean, H.K.; Cortés, J. Non-colonial coral macro-borers as indicators of coral reef status in the South Pacific of Costa Rica. Rev. Biol. Trop. 2006, 54, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Licciano, M.; Stabili, L.; Giangrande, A. Clearance rates of Sabella spallanzanii and Branchiomma luctuosum (Annelida: Polychaeta) on a pure culture of Vibrio alginolyticus. Water Res. 2005, 39, 4375–4384. [Google Scholar] [CrossRef]
- Licciano, M.; Terlizzi, A.; Giangrande, A.; Cavallo, R.A.; Stabili, L. Filter-feeder macroinvertebrates as key players in culturable bacteria biodiversity control: A case of study with Sabella spallanzanii (Polychaeta: Sabellidae). Mar. Environ. Res. 2007, 64, 504–513. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Licciano, M.; Giangrande, A. The mucus of Sabella spallanzanii (Annelida, Polychaeta): Its involvement in chemical defense and fertilization success. J. Exp. Mar. Biol. Ecol. 2009, 374, 144–149. [Google Scholar] [CrossRef]
- Giangrande, A.; Pierri, C.; Arduini, D.; Borghese, J.; Licciano, M.; Trani, R.; Corriero, G.; Basile, G.; Cecere, E.; Petrocelli, A. An innovative IMTA System: Polychaetes, sponges and macroalgae co-cultured in a southern Italian in-shore mariculture plant (Ionian Sea). J. Mar. Sci. Eng. 2020, 8, 733. [Google Scholar] [CrossRef]
- Selim, S.A. New records of sabellid species (Polychaeta: Sabellinae) from the coastal Egyptian waters. Egypt J. Aquat. Res. 2008, 34, 108–128. [Google Scholar]
- Giangrande, A.; Caruso, L.; Musco, L.; Licciano, M. Variability among Mediterranean populations of Sabella pavonina (Annelida: Sabellidae). Ital. J. Zool. 2014, 81, 100–111. [Google Scholar] [CrossRef]
- Magalhães, W.F.; Bailey-Brock, J.H.; Tovar-Hernández, M.A. An abundant new genus and species of fan worms (Polychaeta: Sabellidae) from Hawaii. Zootaxa 2020, 4763, 85–98. [Google Scholar] [CrossRef]
- Capa, M.; Nishi, E.; Tanaka, K.; Fujikura, K. First record of a Bispira species (Sabellidae: Polychaeta) from a hydrothermal vent. Mar. Biodivers. Rec. 2013, 6, e68. [Google Scholar] [CrossRef]
- Amon, D.J.; Glover, A.G.; Wiklund, H.; Marsh, L.; Linse, K.; Rogers, A.D.; Copley, J.T. The discovery of a natural whale fall in the Antarctic deep sea. Deep Sea Res. Part. II Top. Stud. Ocean. 2013, 92, 87–96. [Google Scholar] [CrossRef]
- Cordes, E.E.; Carney, S.L.; Hourdez, S.; Carney, R.S.; Brooks, J.M.; Fisher, C.R. Cold seeps of the deep Gulf of Mexico: Community structure and biogeographic comparisons to Atlantic Equatorial belt seep communities. Deep Sea Res. Part. I Ocean. Res. Pap. 2007, 54, 637–653. [Google Scholar] [CrossRef]
- Sellanes, J.; Quiroga, E.; Neira, C. Megafauna community structure and trophic relationships at the recently discovered Concepción methane seep area, Chile,∼ 36° S. ICES. Mar. Sci. 2008, 65, 1102–1111. [Google Scholar] [CrossRef]
- Goffredi, S.K.; Tilic, E.; Mullin, S.W.; Dawson, K.S.; Keller, A.; Lee, R.W.; Wu, F.; Levin, L.A.; Rouse, G.W.; Cordes, E.E. Methanotrophic bacterial symbionts fuel dense populations of deep-sea feather duster worms (Sabellida, Annelida) and extend the spatial influence of methane seepage. Sci. Adv. 2020, 6, eaay8562. [Google Scholar] [CrossRef]
- Hutchings, P.; Murray, A. Taxonomy of the polychaetes from the Hawkesbury River and the Southern estuaries of New South Wales, Australia. Rec. Aust. Mus. 1984, 36, 1–119. [Google Scholar] [CrossRef]
- Lardicci, C.; Castelli, A. Desdemona ornata Banse, 1957 (Polychaeta, Sabellidae, Fabricinae) New record in the Mediterranean Sea. Oebalia 1986, 13, 195–201. [Google Scholar]
- Castelli, A.; Galletti, C.; Lardicci, C. Structure of benthic communities of brackish-water microhabitats: Spatial and temporal variations. MAP Tech. Rep. Ser. (UNEP) 1988, 22, 19–46. [Google Scholar]
- Panagopoulos, D.; Nicolaidou, A. A population of Desdemona ornata Banse, 1957 (Polychaeta, Sabellidae) settled in a fully marine habitat of the Mediterranean. Oebalia 1989, 16, 35–39. [Google Scholar]
- Glasby, C.J.; Timm, T.; Muir, A.I.; Gil, J. Catalogue of non-marine Polychaeta (Annelida) of the World. Zootaxa 2009, 2070, 1–52. [Google Scholar] [CrossRef]
- Hossain, M.B.; Marshall, D.J. Benthic infaunal community structuring in an acidified tropical estuarine system. Aquat. Biosyst. 2014, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alcántara, P.; Cortés-Solano, J.D.; Medina-Cantú, N.M.; Avilés-Díaz, A.L.; Solís-Weiss, V. Polychaete diversity in the estuarine habitats of Términos lagoon, Southern Gulf of Mexico. Mem. Mus. Vic. 2014, 71, 97–107. [Google Scholar] [CrossRef]
- Hsieh, H.L. Laonome albicingillum, a new fan worm species (Polychaeta, Sabellidae, Sabellinae) from Taiwan. Proc. Biol. Soc. Wash. 1995, 108, 130–135. [Google Scholar]
- Hsieh, H.L. Self-fertilization-a potential fertilization mode in an estuarine sabellid polychaete. Mar. Ecol. Prog. Ser. 1997, 147, 143–148. [Google Scholar] [CrossRef][Green Version]
- Capa, M.; van Moorsel, G.; Tempelman, D. The Australian feather-duster worm Laonome calida Capa, 2007 (Annelida: Sabellidae) introduced into European inland waters? Bioinvasions Rec. 2014, 3, 1–11. [Google Scholar] [CrossRef]
- Bonar, D.B. Feeding and tube construction in Chone mollis Bush (Polychaeta, Sabellidae). J. Exp. Mar. Biol. Ecol. 1972, 9, 1–18. [Google Scholar] [CrossRef]
- Fitzsimons, G. Feeding and tube building in Sabellastarte magnifica (Shaw) (Sabellidae: Polychaeta). Bull. Mar. Sci. 1965, 15, 642–671. [Google Scholar]
- Licciano, M.; Murray, J.M.; Watson, G.J.; Giangrande, A. Morphological comparison of the regeneration process in Sabella spallanzanii and Branchiomma luctuosum (Annelida, Sabellida). Invertebr. Biol. 2012, 131, 40–51. [Google Scholar] [CrossRef]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of worms emended: An update of polychaete feeding guilds. Ann. Rev. Mar. Sci 2015, 7, 497–520. [Google Scholar] [CrossRef]
- Jones, M.L. On the Caobangiidae, a new family of the Polychaeta, with a redescription of Caobangia billeti Giard. Smithson. Contr. Zool. 1974, 1–55. [Google Scholar] [CrossRef]
- Kuris, A.M.; Culver, C.S. An introduced sabellid polychaete pest infesting cultured abalones and its potential spread to other California gastropods. Invertebr. Biol. 1999, 118, 391–403. [Google Scholar] [CrossRef]
- Ruck, K.R.; Cook, P.A. Sabellid infestations in the shells of South African molluscs: Implications for abalone mariculture. J. Shellfish Res. 1998, 17, 693–699. [Google Scholar]
- Kupriyanova, E.K.; Badyaev, A.V. Ecological correlates of Arctic Serpulidae (Annelida, Polychaeta) distributions. Ophelia 1998, 49, 181–193. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Nishi, E.; ten Hove, H.A.; Rzhavsky, A.V. A review of life history patterns in serpulimorph polychaetes: Ecological and evolutionary perspectives. Ocean. Mar. Biol. Ann. Rev. 2001, 39, 1–101. [Google Scholar]
- Charles, F.; Jordana, E.; Amouroux, J.-M.; Gremare, A.; Desmalades, M.; Zudaire, L. Reproduction, recruitment and larval metamorphosis in the serpulid polychaete Ditrupa arietina (O.F. Muller). Estuar. Coast. Shelf Sci. 2003, 57, 435–443. [Google Scholar] [CrossRef]
- Goffredi, S.K.; Johnson, S.; Tunnicliffe, V.; Caress, D.; Clague, D.; Escobar, E.; Paduan, J.B.; Rouse, G.; Salcedo, D.L.; Soto, L.A.; et al. Hydrothermal vent fields discovered in the Southern Gulf of California clarify role of habitat in augmenting regional diversity. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170817. [Google Scholar] [CrossRef] [PubMed]
- Kupriyanova, E.K.; Nishi, E.; Kawato, M.; Fujiwara, Y. New records of Serpulidae (Annelida, Polychaeta) from hydrothermal vents of North Fiji, Pacific Ocean. Zootaxa 2010, 2389, 57–68. [Google Scholar] [CrossRef]
- Vinn, O.; Kupriyanova, E.K.; Kiel, S. Systematics of serpulid tubeworms (Annelida, Polychaeta) from Cretaceous and Cenozoic hydrocarbon-seep deposits in North America and Europe. Neues Jahrb. Geol. Paläontol. 2012, 265, 315–325. [Google Scholar] [CrossRef]
- Juniper, S.K.; Sibuet, M. Cold seep benthic communities in Japan subduction zones: Spatial organisation, trophic strategies and evidence for temporal evolution. Mar. Ecol. Prog. Ser. 1987, 40, 115–126. [Google Scholar] [CrossRef]
- Olu, K.; Duperret, A.; Sibuet, M.; Foucher, J.-P.; Fiala-Médioni, A. Structure and distribution of cold seep communities along the Peruvian Active Margin: Relationship to geological and fluid patterns. Mar. Ecol. Progr. Ser. 1996, 132, 109–125. [Google Scholar] [CrossRef]
- Segonzac, M.; Hekinian, R.; Auzende, J.; Francheteau, J. Recently discovered animal communities on the South East Pacific Rise (17-19 S and the Eastern Microplaque Region). Cah. Biol. Mar. 1997, 38, 140–141. [Google Scholar]
- López-González, P.; Rodríguez, E.; Gili, J.-M.; Segonzac, M. New records on sea anemones (Anthozoa: Actiniaria) from hydrothermal vents and cold seeps. Zool. Verh. 2003, 345, 215–243. [Google Scholar]
- Sibuet, M.; Olu, K. Biogeography, biodiversity and fluid dependence of deep-sea cold-seep communities at active and passive margins. Deep Sea Res. Part. II Top. Stud. Ocean. 1998, 45, 517–567. [Google Scholar] [CrossRef]
- Mak, P.; Huang, Z. The salinity tolerance of the serpulid polychaete, Hydroides elegans (Haswell, 1883), and its possible applications in bio-antifouling. The marine flora and fauna of Hong Kong and Southern China. Ecol. Morph. Behav. Phys. 1982, 1, 817–823. [Google Scholar]
- Knight-Jones, P. Behaviour, setal inversion and phylogeny of Sabellida (Polychaeta). Zool. Scr. 1981, 10, 183–202. [Google Scholar] [CrossRef]
- Dill, L.M.; Fraser, A.H.G. The worm re-turns: Hiding behavior of a tube-dwelling marine polychaete, Serpula vermicularis. Behav. Ecol. 1997, 8, 186–193. [Google Scholar] [CrossRef]
- Pan, J.; Marcoval, M.A. Top-down effects of an exotic serpulid polychaete on natural plankton assemblage of estuarine and brackish systems in the SW Atlantic. J. Coast. Res. 2014, 30, 1226–1235. [Google Scholar] [CrossRef]
- Jordana, E.; Charles, F.; Gremare, A.; Amouroux, J.-M.; Chretiennot-Dinet, M.-J. Food sources, ingestion and absorption in the suspension-feeding polychaete, Ditrupa arietina (O. F. Muller). J. Exp. Mar. Biol. Ecol. 2001, 266, 219–236. [Google Scholar] [CrossRef]
- Bailey-Brock, J.H. Polychaetes from Fijian coral reefs. Pac. Sci. 1985, 39, 195–220. [Google Scholar]
- Nishi, E. Serpulid polychaetes associated with living and dead corals at Okinawa Island, Southwest Japan. Publ. Seto Mar. Biol. Lab. 1996, 36, 305–318. [Google Scholar] [CrossRef][Green Version]
- Marsden, J.R. Coral preference behaviour by planktotrophic larvae of Spirobranchus giganteus corniculatus (Serpulidae: Polychaeta). Coral Reefs 1987, 6, 71–74. [Google Scholar] [CrossRef]
- Hunte, W.; Conlin, B.E.; Marsden, J.R. Habitat selection in the tropical polychaete Spirobranchus giganteus. I. Distribution on corals. Mar. Biol. 1990, 104, 87–92. [Google Scholar] [CrossRef]
- Marsden, J.R. Light responses of the planktotrophic larva of the serpulid polychaete Spirobranchus polycerus. Mar. Ecol. Prog. Ser. 1990, 58, 225–233. [Google Scholar]
- Williams, G.B. The effect of extracts of Fucus serratus in promoting the settlement of larvae of Spirorbis borealis (Polychaeta). J. Mar. Biol. Assoc. UK 1964, 44, 397–414. [Google Scholar] [CrossRef]
- Gee, J.M. Chemical stimulation of settlement in larvae of Spirorbis rupestris (Serpulidae). Anim. Behav. 1965, 13, 181–186. [Google Scholar] [CrossRef]
- Hartman, O. Catalogue of the polychaetous annelids of the world. Part II. Occ. Pap. Allan Hancock Found. 1959, 23, 355–628. [Google Scholar]
- Darling, J.A.; Carlton, J.T. A framework for understanding marine cosmopolitanism in the Anthropocene. Front. Mar. Sci. 2018, 5, 1–293. [Google Scholar] [CrossRef]
- Hutchings, P.; Kupriyanova, E. Cosmopolitan polychaetes–fact or fiction? Personal and historical perspectives. Invertebr. Syst. 2018, 32, 1–9. [Google Scholar] [CrossRef]
- Briggs, J.C. Marine Zoogeography; McGraw-Hill: New York, NY, USA, 1974; pp. 1–475. [Google Scholar]
- McClain, C.R.; Schlacher, T.A. On some hypotheses of diversity of animal life at great depths on the sea floor. Mar. Ecol. 2015, 36, 849–872. [Google Scholar] [CrossRef]
- Higgs, N.D.; Attrill, M. Biases in biodiversity: Wide-ranging species are discovered first in the deep sea. Front. Mar. Sci. 2015, 2, 61. [Google Scholar] [CrossRef]
- Giangrande, A.; Licciano, M. Factors influencing latitudinal pattern of biodiversity: An example using Sabellidae (Annelida, Polychaeta). Biodiv. Conserv. 2004, 13, 1633–1646. [Google Scholar] [CrossRef]
- Banse, K. Beiträge zur kenntnis der gattungen Fabricia, Manayunkia und Fabriciola (Sabellidae, Polychaeta). Zool. Jahrb. (Abt. Syst. Okol. Geogr. Tiere) 1956, 84, 415–438. [Google Scholar]
- Bourne, A.G. On Haplobranchus, a new genus of Capitobranchiate annelids. J. Cell Sci. 1883, 23, 168–176. [Google Scholar]
- Annenkova, N.P. Über Die Pontokaspischen Polychaeten. II. Die Gattungen Hypaniola, Parhypania, Fabricia und Manajunkia. Ann. Mussée Zool. L’Academie des Sci. L’URSS, Leningrad 1928, 30, 13–20. [Google Scholar]
- Day, J.H. The polychaete fauna of South Africa Part 8: New species and records from grab samplings and dreddings. Bull. Brit. Mus. (Nat. Hist.) Zool. 1963, 10, 381–445. [Google Scholar] [CrossRef]
- Perkins, T.H. Revision of Demonax Kinberg, Hypsicomus Grube, and Notaulax Tauber, with a review of Megalomma Johansson from Florida (Polychaeta: Sabellidae). Proc. Biol. Soc. Wash. 1984, 97, 285–368. [Google Scholar]
- Knight-Jones, P.; Perkins, T.H. A revision of Sabella and Bispira and Stylomma. (Sabellidae: Polychaeta). Zool. J. Linn. Soc. 1998, 123, 385–467. [Google Scholar] [CrossRef]
- Knight-Jones, P.; Mackie, A.S. A revision of Sabellastarte (Polychaeta: Sabellidae). J. Nat. Hist. 2003, 37, 2269–2301. [Google Scholar] [CrossRef]
- Hartman, O. Abyssal polychaetous annelids from the Mozambique Basin off Southeast Africa, with a compendium of abyssal polychaetous annelids from world-wide Areas. J. Fish. Res. Board Can. 1971, 28, 1407–1428. [Google Scholar] [CrossRef]
- Uschakov, P.V. Bathypelagic and deepwater forms of marine polychaetes from Kamchatka Area of the Pacific. Issled. Fauny Morej USSR 1952, 3, 103–112. [Google Scholar]
- Levenstein, R.Y. Polychaeta from deep water in the Bering Sea. Proc. Inst. Oceanol. USSR Acad. Sci 1961, 46, 147–178. [Google Scholar]
- Levenstein, R.Y. Polychaeta. In Pacific Ocean. Biology. Deep-Sea Fauna; Zenkevitch, L.A., Ed.; Nauka: Moscow, Russia, 1969; pp. 52–62. (In Russian) [Google Scholar]
- Belyaev, G.M. Hadal Bottom Fauna of the World Ocean; Institute of Oceanology, USSR Academy of Sciences: Moscow, Russia, 1972; pp. 1–199. [Google Scholar]
- Belyaev, G.M. Glubokovodnye Okeanicheskie Zheloba i Ikh Fauna [Deep-Sea Oceanic Trenches and Their Fauna]; Nauka: Moscow, Russia, 1989; pp. 1–256. (In Russian) [Google Scholar]
- Moore, J.P. Additional new species of Polychaeta from the North Pacific. Proc. Acad. Nat. Sci. Phil. 1906, 58, 217–260. [Google Scholar]
- Moore, J.P. The Polychætous annelids Dredged by the U. S. S. “Albatross” off the Coast of Southern California in 1904. IV. Spionidae to Sabellariidae. Proc. Acad. Nat. Sci. Phil. 1923, 7, 179–259. [Google Scholar]
- Hartman, O. Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic Areas. Occ. Pap. Allan Hancock Found. 1965, 28, 1–378. [Google Scholar]
- Méndez, N. Trophic categories of soft-bottom epibenthic deep-sea polychaetes from the Southeastern Gulf of California (Mexico) in relation with environmental variables. Pan Am. J. Aquat. Sci. 2013, 8, 299–311. [Google Scholar]
- Zibrowius, H. Review of Serpulidae (Polychaeta) from depths exceeding 2000 meters. In Essays on Polychaetous Annelids in Memory of Dr. Olga Hartman; Reish, D.J., Fauchald, K., Eds.; Allan Hancock Press: Los Angeles, CA, USA, 1977; pp. 289–306. [Google Scholar]
- Kirkergaard, J.B. Benthic Polychaeta from depths exceeding 6000 m. Galathea Rep. 1956, 2, 63–78. [Google Scholar]
- Paterson, G.L.; Glover, A.G.; Froján, C.R.B.; Whitaker, A.; Budaeva, N.; Chimonides, J.; Doner, S. A Census of abyssal polychaetes. Deep Sea Res. Part. II Top. Stud. Ocean. 2009, 56, 1739–1746. [Google Scholar] [CrossRef]
- Kupriyanova, E.K. Deep-water Serpulidae (Annelida, Polychaeta) from the Kurile-Kamchatka Trench. 2. Genera Bathyditrupa, Bathyvermilia and Protis. Zool. Zhurnal 1993, 72, 21–28. [Google Scholar]
- Kupriyanova, E.K. Deep-water Serpulidae (Annelida, Polychaeta) from the Kurile-Kamchatka Trench. 1. Genus Hyalopomatus. Zool. Zhurnal 1993, 72, 145–152. [Google Scholar]
- Sanfilippo, R. New species of Hyalopomatus Marenzeller, 1878 (Annelida, Polychaeta, Serpulidae) from Recent Mediterranean deep-water coral mounds and comments on some congeners. Zoosystema 2009, 31, 147–161. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Bailey-Brock, J.; Nishi, E. New records of Serpulidae (Annelida, Polychaeta) collected by R/V “Vityaz” from bathyal and abyssal depths of the Pacific Ocean. Zootaxa 2011, 2871, 43–60. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Vinn, O.; Taylor, P.D.; Schopf, J.W.; Kudryavtsev, A.B.; Bailey-Brock, J. Serpulids living deep: Calcareous tubeworms beyond the abyss. Deep Sea Res. Part. I Ocean. Res. 2014, 90, 91–104. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Nishi, E. New records of the deep-sea Nogrobs grimaldii (Serpulidae: Annelida). Mar. Biodivers. Rec. 2011, 4, e74. [Google Scholar] [CrossRef]
- Rouse, G.W.; Kupriyanova, E.K. Laminatubus (Serpulidae, Annelida) from Eastern Pacific hydrothermal vents and methane seeps, with description of two new species. Zootaxa 2021, 4915, 1–27. [Google Scholar] [CrossRef]
- Thorp, C.H.; Pyne, S.; West, S.A. Hydroides ezoensis Okuda, a fouling serpulid new to British coastal waters. J. Nat. Hist. 1987, 21, 863–877. [Google Scholar] [CrossRef]
- Zibrowius, H. A propos des pretendud “Recifs de Serpulidae” de l’lle Rousse, Corse (Mediterranee Nord-Occidentale). Mesogee 1991, 50, 71–74. [Google Scholar]
- Zibrowius, H. Assessing scale and impact of ship-transported alien fauna in the Mediterranean. In Alien Marine Organisms Introduced by Ships in the Mediterranean and Black Seas; Briand, F., Ed.; CIESM Workshop Monographs: Monaco, France, 2002; Volume 20, pp. 62–68. [Google Scholar]
- Lewis, J.A.; Watson, C.; Harry, A. Establishment of the Caribbean serpulid tubeworm Hydroides sanctaecrucis Krøyer [in] Mörch, 1863, in Northern Australia. Biol. Invasions 2006, 8, 665–671. [Google Scholar] [CrossRef]
- Giangrande, A.; Licciano, M.; Pagliara, P.; Gambi, M.C. Gametogenesis and larval development in Sabella spallanzanii (Polychaeta: Sabellidae) from the Mediterranean Sea. Mar. Biol. 2000, 136, 847–861. [Google Scholar] [CrossRef]
- Read, G.; Inglis, G.; Stratford, P.; Ahyong, S. Arrival of the alien fanworm Sabella spallanzanii (Gmelin, 1791) (Polychaeta: Sabellidae) in two New Zealand harbours. Aquat. Invasions 2011, 6, 273–279. [Google Scholar] [CrossRef]
- Çinar, M.E. Alien polychaete species worldwide: Current status and their impacts. J. Mar. Biol. Assoc. UK 2013, 93, 1257–1278. [Google Scholar] [CrossRef]
- Morais, P.; Reichard, M. Cryptic invasions: A review. Sci. Total Environ. 2018, 613, 1438–1448. [Google Scholar] [CrossRef]
- Chapman, J.W.; Carlton, J.T. A test of criteria for introduced species: The global invasion by the isopod Synidotea laevidorsalis (Miers, 1881). J. Crust. Biol. 1991, 11, 386–400. [Google Scholar] [CrossRef]
- Wolff, W.J. Non-Indigenous marine and estuarine species in the Netherlands. Zool. Med. Leiden 2005, 79, 1–116. [Google Scholar]
- Patti, F.; Gambi, M. Phylogeography of the invasive polychaete Sabella spallanzanii (Sabellidae) based on the nucleotide sequence of internal transcribed spacer 2 (ITS2) of nuclear rDNA. Mar. Ecol. Prog. Ser. 2001, 215, 169–177. [Google Scholar] [CrossRef]
- Ahyong, S.T.; Kupriyanova, E.; Burghardt, I.; Sun, Y.; Hutchings, P.A.; Capa, M.; Cox, S.L. Phylogeography of the invasive Mediterranean fan worm, Sabella spallanzanii (Gmelin, 1791), in Australia and New Zealand. J. Mar. Biol. Assoc. UK 2017, 97, 985–991. [Google Scholar] [CrossRef]
- Pabis, K.; Krodkiewska, M.; Cebulska, K. Alien freshwater polychaetes Hypania invalida (Grube 1860) and Laonome calida Capa, 2007 in the Upper Odra River (Baltic Sea catchment area). Knowl. Manag. Aquat. Ecosyst. 2017, 418, 46. [Google Scholar] [CrossRef]
- Boltachova, N.; Lisitskaya, E.; Frolenko, L.; Kovalev, E.; Barabashin, T. The finding of polychaete Laonome calida Capa, 2007 (Annelida: Sabellidae) in the Southeast Sea of Azov. Russ. J. Biol. Invasions 2017, 8, 303–306. [Google Scholar] [CrossRef]
- Tamulyonis, A.Y.; Gagaev, S.Y.; Stratanenko, E.A.; Zuyev, Y.A.; Potin, V.V. Invasion of the Polychaeta Laonome xeprovala Bick & Bastrop, 2018 (Sabellidae, Polychaeta) into the estuary of the Luga and Khabolovka Rivers (Luga Bay, Gulf of Finland). Russ. J. Biol. Invasions 2020, 11, 148–154. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Yáñez-Rivera, B.; Bortolini-Rosales, J.L. Reproduction of the invasive fan worm Branchiomma bairdi (Polychaeta: Sabellidae). Mar. Biol. Res. 2011, 7, 710–718. [Google Scholar] [CrossRef]
- Roman, S.; Perez-Ruzafa, A.; Lopez, E. First record in the Western Mediterranean Sea of Branchiomma boholense (Grube, 1878) (Polychaeta: Sabellidae), an alien species of Indo-Pacific origin. Cah. Biol. Mar. 2009, 50, 241–250. [Google Scholar]
- Arias, A.; Giangrande, A.; Gambi, M.C.; Anadón, N. Biology and new records of the invasive species Branchiomma bairdi (Annelida: Sabellidae) in the Mediterranean Sea. Mediterr. Mar. Sci. 2013, 10, 162–171. [Google Scholar] [CrossRef]
- Ramalhosa, P.; Camacho-Cruz, K.; Bastida-Zavala, R.; Canning-Clode, J. First record of Branchiomma bairdi McIntosh, 1885 (Annelida: Sabellidae) from Madeira Island, Portugal (Northeastern Atlantic Ocean). Bioinvasions Rec. 2014, 3, 235–239. [Google Scholar] [CrossRef]
- Cepeda, D.; Rodríguez-Flores, P. First record of the invasive worm Branchiomma bairdi (Annelida: Sabellidae) in the Balearic Sea (Western Mediterranean). J. Mar. Biol. Assoc. UK 2018, 98, 1955–1963. [Google Scholar] [CrossRef]
- Del Pasqua, M.; Schulze, A.; Tovar-Hernández, M.A.; Keppel, E.; Lezzi, M.; Gambi, M.C.; Giangrande, A. Clarifying the taxonomic status of the alien species Branchiomma bairdi and Branchiomma boholense (Annelida: Sabellidae) using molecular and morphological evidence. PLoS ONE 2018, 13, e0197104. [Google Scholar] [CrossRef]
- Hewitt, C.L.; Campbell, M.L.; Thresher, R.E.; Martin, R.B.; Boyd, S.; Cohen, B.F.; Currie, D.R.; Gomon, M.F.; Keough, M.J.; Lewis, J.A.; et al. Introduced and cryptogenic species in Port Phillip Bay, Victoria, Australia. Mar. Biol. 2004, 144, 183–202. [Google Scholar] [CrossRef]
- Zenetos, A.; Çinar, M.E.; Pancucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Gravili, C.; Belmonte, G.; Cecere, E.; Denitto, F.; Giangrande, A.; Guidetti, P.; Longo, C.; Mastrototaro, F.; Moscatello, S.; Petrocelli, A. Nonindigenous species along the Apulian Coast, Italy. Chem. Ecol. 2010, 26, 121–142. [Google Scholar] [CrossRef]
- Giangrande, A.; Montanaro, P.; Castelli, A. On some Amphicorina (Polychaeta, Sabellidae) species from the Mediterranean coast, with the description of A. grahamensis. Ital. J. Zool. 1999, 66, 195–203. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Çinar, M.E.; Garcia Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediter. Mar. Sci. 2010, 11, 381. [Google Scholar] [CrossRef]
- Faasse, M.A.; Giangrande, A. Description of Bispira polyomma n. sp. (Annelida: Sabellidae): A probable introduction to the Netherlands. Aquat. Invasions 2012, 7, 591–598. [Google Scholar] [CrossRef]
- Capa, M.; Murray, A. A taxonomic guide to the fanworms (Sabellidae, Annelida) of Lizard Island, Great Barrier Reef, Australia, including new species and new records. Zootaxa 2015, 4019, 98–167. [Google Scholar] [CrossRef] [PubMed]
- Khedhri, I.; Tovar-Hernández, M.A.; Bonifácio, P.; Ahmed, A.; Aleya, L. First report of the invasive species Branchiomma bairdi McIntosh, 1885 (Annelida: Sabellidae) along the Tunisian Coast (Mediterranean Sea). Bioinvasions Rec. 2017, 6, 139–145. [Google Scholar] [CrossRef]
- Abdelnaby, F. Alien polychaete species and the first record of Branchiomma bairdi (McIntosh, 1885) from the Suez Canal and the Mediterranean coast of Egypt. Egypt. J. Aquat. Biol. Fish. 2020, 24, 13–32. [Google Scholar] [CrossRef]
- Knight-Jones, P.; Knight-Jones, W.; Ergen, Z. Sabelliform Polychaetes, mostly from Turkey’s Aegean coast. J. Nat. Hist. 1991, 25, 837–858. [Google Scholar]
- Çinar, M.E. Polychaetes from the coast of Northern Cyprus (Eastern Mediterranean Sea), with Two new records for the Mediterranean Sea. Cah. Biol. Mar. 2005, 46, 143–160. [Google Scholar]
- Keppel, E.; Tovar Hernández, M.A.; Ruiz, G.M. First record and establishment of Branchiomma coheni (Polychaeta: Sabellidae) in the Atlantic Ocean and review of non indigenous species of the genus. Zootaxa 2015, 4058, 499–518. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Dean, H.A. New gregarious sabellid worm from the Gulf of California reproducing by dpontaneous fission (Polychaeta, Sabellidae). J. M. Biol. Assoc. UK 2014, 94, 935–946. [Google Scholar] [CrossRef]
- Bianchi, C. Serpuloidea (Annelida, Polychaeta) Delle Lagune Costiere Laziali e Campane. Annali del Museo Civico di Storia Naturale di Genova 1983, 84, 231–243. [Google Scholar]
- Arvanitides, C. Polychaete fauna of the Aegean Sea: Inventory and new information. Bull. Mar. Sci. 2000, 66, 73–96. [Google Scholar]
- Simboura, N.; Nicolaidou, A. The Polychaetes (Annelida, Polychaeta) of Greece: Checklist, distribution and ecological characteristics. Monographs on Marine Sciences, Ser. 4; National Center for Marine Research: Athens, Greece, 2001; pp. 1–115. [Google Scholar]
- Licciano, M.; Giangrande, A.; Gambi, M.C. Reproduction and simultaneous hermaphroditism in Branchiomma luctuosum (Polychaeta, Sabellidae) from the Mediterranean Sea. Invertebr. Biol. 2002, 121, 55–65. [Google Scholar] [CrossRef]
- Mastrototaro, F.; Petrocelli, A.; Cecere, E.; Matarrese, A. Non indigenous species settle down in the Taranto Seas. Biogeographia 2004, 25, 47–54. [Google Scholar] [CrossRef]
- Çinar, M.E.; Bilecenoglu, M.; Öztürk, B.; Can, A. New records of alien species on the Levantine coast of Turkey. Aquat. Invasions 2006, 1, 84–90. [Google Scholar] [CrossRef]
- El Haddad, M.; Capaccioni Azzati, R.; García-Carrascosa, A.M. Branchiomma luctuosum (Polychaeta: Sabellidae): A non-indigenous species at Valencia Port (Western Mediterranean Sea, Spain). Mar. Biodivers. Rec. 2008, 1, e61. [Google Scholar] [CrossRef]
- Giangrande, A.; Licciano, M. Revision of the species of Megalomma (Polychaeta, Sabellidae) from the Mediterranean Sea, with the description of M. messapicum n. sp. Ital. J. Zool. 2008, 75, 207–217. [Google Scholar] [CrossRef]
- Tanduo, V.; Golemaj, A.; Crocetta, F. Citizen-science detects the arrival and establishment of Branchiomma luctuosum (Grube, 1870) (Annelida: Polychaeta: Sabellidae) in Albania. Biodivers. Data J. 2020, 8, e54790. [Google Scholar] [CrossRef]
- De Matos Nogueira, J.M.; Rossi, M.C.S.; López, E. Intertidal species of Branchiomma Kolliker and Pseudobranchiomma Jones (Polychaeta: Sabellidae: Sabellinae) occurring on rocky shores along the state of Sao Paulo, Southeastern Brazil. Zool. Stud. 2006, 45, 586–610. [Google Scholar]
- Hartmann-Schröder, G. Teil 8. Die Polychaeten der Subtropisch-Antiborealen Westküste Australiens (Zwischen Cervantes im Norden und Cape Naturliste im Suden). Mitt. Hamb. Zool. Mus. Inst. 1982, 79, 51–118. [Google Scholar]
- Ceberio, A.; Martínez, J.; Aguirrezabalaga, F. Presencia de Desdemona ornata Banse, 1957 (Polychaeta, Sabellidae) en las costas de La Península Ibérica, Golfo de Vizcaya. Munibe 1998, 50, 37–41. [Google Scholar]
- Smith, P.; Perrett, J.; Garwood, P.; Moore, G. Two additions to the UK marine fauna: Desdemona ornata Banse, 1957 (Polychaeta, Sabellidae) and Grandidierella japonica Stephensen, 1938 (Amphipoda, Gammaridea). Newsl. Porcupine Mar. Nat. Hist. Soc. 1999, 2, 8–11. [Google Scholar]
- Çinar, M.E.; Dagli, E.; Açik, S. Annelids (Polychaeta and Oligochaeta) from the Sea of Marmara, with descriptions of five new species. J. Nat. Hist. 2011, 45, 2105–2143. [Google Scholar] [CrossRef]
- Carvalho, S.; Pereira, P.; Pereira, F.; de Pablo, H.; Vale, C.; Gaspar, M.B. Factors structuring temporal and spatial dynamics of macrobenthic communities in a eutrophic coastal lagoon (Óbidos Lagoon, Portugal). Mar. Environ. Res. 2011, 71, 97–110. [Google Scholar] [CrossRef]
- Faasse, M. Dispersal of the invasive tubeworms Desdemona ornata and Pseudopolydora paucibranchiata to the Netherlands (Polychaeta: Sedentaria). Nederl. Faun. Med. 2016, 46, 49–56. [Google Scholar]
- McArthur, M. Systematics and Introduced Status of Euchone (Polychaeta: Sabellidae) in Port Phillip Bay, and the Feeding Biology of Euchone limnicola. Bachelor’s Thesis, Victoria University of Technology, Melbourne, Australia, 1997. [Google Scholar]
- Guyonnet, B.; Borg, D. Premier signalement de l’espèce introduite Euchone limnicola Reish, 1959 (Polychaeta: Sabellidae) sur ses Côtes Françaises de la Mer du Nord (Grand Port Maritime de Dunkerque). Cah. Nat. Observat. Mar. 2015, 4, 15–23. [Google Scholar]
- 411. Dane, L Morphological and Genetic Variation in the Cryptic Species Complex Myxicola infundibulum (Polychaeta, Sabellidae), and its Introduction to Australia. Master’s Thesis, University of Melbourne, Melbourne, Australia, 2008.
- Keppel, E.; Ruiz, G.; Tovar–Hernández, M. Re-description of Parasabella fullo (Grube, 1878) (Polychaeta: Sabellidae) and diagnostic characteristics for detection in California. Eur. Zool. J. 2020, 87, 105–115. [Google Scholar] [CrossRef]
- Currie, D.; McArthur, M.; Cohen, B. Exotic Marine Pests in the Port. of Geelong, Victoria. Report 8; Marine and Freshwater Resources Institute: Queenscliff, Australia, 1998; pp. 1–57. [Google Scholar]
- Coles, S.L. Historical and recent introductions of non-indigenous marine species into Pearl Harbor, Oahu, Hawaiian Islands. Mar. Biol. 1999, 135, 147–158. [Google Scholar] [CrossRef]
- Eldredge, L.G.; Smith, C.A. Guidebook of Introduced Marine Species in Hawaii; Bishop Museum Technical Report 21; Bishop Museum: Honolulu, Hawaii, USA, 2001. [Google Scholar]
- Coles, S.; Eldredge, L.G. Nonindigenous species introductions on coral reefs: A need for information. Pac. Sci. 2002, 56, 191–209. [Google Scholar] [CrossRef]
- Culver, C.S.; Kuris, A.M.; Beede, B. Identification and Management of the Exotic Sabellid Pest in California Cultured Abalone; Californian Sea Grant College System: La Jolla, CA, USA, 1997. [Google Scholar]
- Moore, J.D.; Juhasz, C.I.; Robbins, T.T.; Grosholz, E.D. The introduced sabellid polychaete Terebrasabella heterouncinata in California: Transmission, methods of control and survey for presence in native gastropod populations. J. Shellfish Res. 2007, 26, 869–876. [Google Scholar] [CrossRef]
- Moreno, R.A.; Neill, P.E.; Rozbaczylo, N. Native and non-indigenous boring polychaetes in Chile: A threat to native and commercial mollusc species. Rev. Chil. Hist. Nat. 2006, 79, 263–278. [Google Scholar] [CrossRef]
- Dittmann, S.; Rolston, A.; Benger, S.N.; Kupriyanova, E.K. Habitat Requirements, Distribution and Colonisation of the Tubeworm Ficopomatus Enigmaticus in the Lower Lakes and Coorong; Report for the South Australian Murray-Darling Basin Natural Resources Management Board; Department of Environment and water: Adelaide, Australia, 2009; pp. 1–99. [Google Scholar]
- Grosse, M.; Pérez, R.; Juan-Amengual, M.; Pons, J.; Capa, M. The elephant in the room-first record of invasive gregarious species of serpulids (Calcareous Tube Annelids) in Majorca (Western Mediterranean). Sci. Mar. 2021, in press. [Google Scholar]
- Assis, J.D.; Alonso, C.; Christoffersen, M.L. First record of Ficopomatus uschakovi (Pillai, 1960) Serpulidae (Polychaeta: Annelida) for the Western Atlantic. Rev. Nord. Biol. 2008, 19, 51–58. [Google Scholar]
- Bastida-Zavala, R.; García-Madrigal, S. First record in the Tropical Eastern Pacific of the exotic species Ficopomatus uschakovi (Polychaeta, Serpulidae). ZooKeys 2012, 238, 45–55. [Google Scholar] [CrossRef]
- Liñero-Arana, I.; Díaz-Díaz, Ó. Presencia del poliqueto exótico Ficopomatus uschakovi (Polychaeta: Serpulidae) en Venezuela: Descripción y comentarios sobre su distribución. Interciencia 2012, 37, 234–237. [Google Scholar]
- Arteaga-Flórez, C.; Fernández-Rodríguez, V.; Londoño-Mesa, M.H. First record of the polychaete Ficopomatus uschakovi (Pillai, 1960) (Annelida, Serpulidae) in the Colombian Caribbean, South America. Zookeys 2014, 371, 1–11. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zibrowius, H. Les espèces Méditerrannéennes du genre Hydroides (Polychaeta Serpulidae). Remarques sur le prétendu polymorphisme de Hydroides uncinata. Téthys 1971, 2, 691–746. [Google Scholar]
- Zibrowius, H. Introduced invertebrates: Examples of success and nuisance in the European Atlantic and in the Mediterranean. Introduced species in European coastal waters. Ecosyst. Res. Rep. 1994, 8, 44–49. [Google Scholar]
- Cohen, A.N.; Harris, L.H.; Bingham, B.L.; Carlton, J.T.; Chapman, J.W.; Lambert, C.C.; Lambert, G.; Ljubenkov, J.C.; Murray, S.N.; Rao, L.C. Project Report for the Southern California Exotics Expedition; A Rapid Assessment Survey of Exotic Species in Sheltered Coastal Waters California Department of Fish and Game, Sacramento CA State Water Resources Control Board, Sacramento CA National Fish and Wildlife Foundation: San Francisco, CA, USA, 2002; pp. 1–37. [Google Scholar]
- Carlton, J.T.; Eldredge, L.G. Marine bioinvasions of Hawaiʻi; Bernice, P., Ed.; Bishop Museum Press: Honolulu, HI, USA, 2009; pp. 1–202. [Google Scholar]
- Pettengill, J.; Wendt, D.; Schug, M.D.; Hadfield, M. Biofouling likely serves as a major mode of dispersal for the polychaete tubeworm Hydroides elegans as inferred from microsatellite loci. Biofouling 2007, 23, 161–169. [Google Scholar] [CrossRef]
- Gruet, Y.; Hèral, M.; Robert, J.M. Premières observations sur l’introduction de la faune associée au naissain d’huîtres Japonaises Crassostrea gigas (Thunberg), importé sur la Côte Atlantique Française. Cah. Biol. Mar. 1976, 17, 173–184. [Google Scholar]
- Zibrowius, H. Introduction du polychete Serpulidae Japonais Hydroides ezoensis sur la Cote, Atlantique Frnacoise et remarques sur la propagation d’autre espeies de Serpulidae. Tethys 1978, 8, 141–150. [Google Scholar]
- Hayes, K.; Sliwa, C.; Migus, S.; McEnnulty, F.; Dunstan, P. National Priority Pests: Part II Ranking of Australian Marine Pests. In An independent report undertaken for the Department of Environment and Heritage by CSIRO Marine Research; Australian Government Department of the Environment and Heritage: Canberra, Australia, 2005; pp. 1–106. [Google Scholar]
- Ferrario, J.; Minchin, D. Spread of the non-indigenous serpulid Hydroides sanctaecrucis Krøyer in Mörch, 1863 in the Pacific Ocean: A new record from Taiwan. Bioinvasions Rec. 2017, 6, 33–38. [Google Scholar] [CrossRef]
- Lee, A.L.; Dafforn, K.A.; Hutchings, P.A.; Johnston, E.L. Reproductive strategy and gamete development of an invasive fanworm, Sabella spallanzanii (Polychaeta: Sabellidae), a field study in Gulf St Vincent, South Australia. PLoS ONE 2018, 13, e0200027. [Google Scholar]
- Clapin, G.; Evans, D.R. The status of the introduced marine fanworm Sabella spallanzanii in Western Australia: A preliminary investigation. Cent. Res. Introd. Mar. Pests Tech. Rep. 1995, 2, 1–34. [Google Scholar]
- Yee, A.; Mackie, J.; Pernet, B. The distribution and unexpected genetic diversity of the non-indigenous annelid Ficopomatus enigmaticus in California. Aquat. Invasions 2019, 14, 250–266. [Google Scholar] [CrossRef]
- Oliva, M.; De Marchi, L.; Sanches, M.V.; Pires, A.; Cuccaro, A.; Baratti, M.; Chiellini, F.; Morelli, A.; Freitas, R.; Pretti, C. Atlantic and Mediterranean populations of the widespread serpulid Ficopomatus enigmaticus: Developmental responses to carbon nanotubes. Mar. Pollut. Bull. 2020, 156, 111265. [Google Scholar] [CrossRef] [PubMed]
- Zibrowius, H. Serpulidae (Annelida Polychaeta) de l’Ocean Indien arrives sur des Coques de Bateaux a Toulon (France, Mediterranee). Rapports et Proces Verbaux des Reunions Commission Internationale pour l’Exploration Scientifique de la Mer Mediterranee Monaco 1979, 25–26, 133–134. [Google Scholar]
- Ben-Eliahu, M.N. Red Sea serpulids (Polychaeta) in the Eastern Mediterranean. Ophelia Suppl. 1991, 5, 515–528. [Google Scholar]
- Zenetos, A.; Çinar, M.E.; Crocetta, F.; Golani, D.; Rosso, A.; Servello, G.; Shenkar, N.; Turon, X.; Verlaque, M. Uncertainties and validation of alien species catalogues: The Mediterranean as an example. Estuar. Coast. Shelf Sci. 2017, 191, 171–187. [Google Scholar] [CrossRef]
- Streftaris, N.; Zenetos, A. Alien marine species in the Mediterranean-the 100 ‘worst invasives’ and their impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef]
- Dürr, S.; Watson, D.I. Biofouling and antifouling in aquaculture. Biofouling 2010, 12, 267–287. [Google Scholar]
- Wang, J.; Huang, Z. Fouling polychaetes of Hong Kong and adjacent saters. Asian Mar. Biol. 1993, 10, 1–12. [Google Scholar]
- Mohan, P.C.; Aruna, C. The biology of serpulid worms in relation to biofouling. In Recent Developments in Biofouling Control; Thompson, M.F., Nagabhushanam, R., Sarojini, R., Fingerman, M., Eds.; Oxford & IBH Publishing Company: New Delhi, India, 1994; pp. 59–64. [Google Scholar]
- Fitridge, I.; Dempster, T.; Guenther, J.; De Nys, R. The impact and control of biofouling in marine aquaculture: A review. Biofouling 2012, 28, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Schloesser, D.W.; Malakauskas, D.M.; Malakauskas, S.J. Freshwater polychaetes (Manayunkia speciosa) near the Detroit River, Western Lake Erie: Abundance and life-history characteristics. J. Great Lakes Res. 2016, 42, 1070–1083. [Google Scholar] [CrossRef]
- Read, G.B.; Gordon, D.P. Adventive occurrence of the fouling serpulid Ficopomatus enigmaticus (Polychaeta) in New Zealand. N. Z. J. Mar. Freshw. Res. 1991, 25, 269–273. [Google Scholar] [CrossRef]
- Probert, P.K. First record of the introduced fouling tubeworm Ficopomatus enigmaticus (Polychaeta: Serpulidae) in Hawke Bay, New Zealand. N. Z. J. Zool. 1993, 20, 35–36. [Google Scholar] [CrossRef]
- Pernet, B.; Barton, M.; Fitzhugh, K.; Harris, L.; Lizárraga, D.; Ohl, R.; Whitcraft, C. Establishment of the reef-forming tubeworm Ficopomatus enigmaticus (Fauvel, 1923) (Annelida: Serpulidae) in Southern California. Bioinvasions Rec. 2016, 5, 13–19. [Google Scholar] [CrossRef]
- Heiman, K.W.; Micheli, F. Non-native ecosystem engineer alters Estuarine communities. Integr. Comp. Biol. 2010, 50, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Ulman, A.; Ferrario, J.; Forcada, A.; Seebens, H.; Arvanitidis, C.; Occhipinti-Ambrogi, A.; Marchini, A. Alien species spreading via biofouling on recreational vessels in the Mediterranean Sea. J. Appl. Ecol. 2019, 56, 2620–2629. [Google Scholar] [CrossRef]
- Secretaría de Medio Ambiente y Recursos Naturales, Estados Unidos Mexicanos. Acuerdo por el que se Determina la Lista de las Especies Exóticas Invasoras Para México; Secretaría de Medio Ambiente y Recursos Naturales: Ciudad de México, Mexico, 2016. [Google Scholar]
- Mackie, G.L.; Qadri, S.U. A polychaete, Manayunkia speciosa, from the Ottawa River, and Its North American distribution. Canad. J. Zool. 1971, 49, 780–782. [Google Scholar] [CrossRef]
- Stocking, R.W.; Bartholomew, J.L. Distribution and habitat characteristics of Manayunkia speciosa and infection prevalence with the parasite Ceratomyxa shasta in the Klamath River, Oregon–California. J. Parasitol. 2007, 93, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Piazzolla, D.; Cafaro, V.; Mancini, E.; Scanu, S.; Bonamano, S.; Marcelli, M. Preliminary investigation of microlitter pollution in low-energy hydrodynamic basins using Sabella spallanzanii (Polychaeta: Sabellidae) tubes. Bull. Environ. Contam. Toxicol. 2020, 104, 345–350. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Thilagam, H.; Raja, P.V. Comparison of heavy metal toxicity in life stages (spermiotoxicity, egg toxicity, embryotoxicity and larval toxicity) of Hydroides elegans. Chemosphere 2008, 71, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Thilagam, H.; Gopalakrishnan, S.; Vijayavel, K.; Raja, P.V. Effluent toxicity test using developmental stages of the marine polychaete Hydroides elegans. Arch. Environ. Contam. Toxicol. 2008, 54, 674–683. [Google Scholar] [CrossRef]
- Vijayaragavan, S.; Raja, P.V. The toxic effects of phorate on early embryonic stages of sedentary polychaete Hydroides elegans (Haswell, 1883). J. Basic Appl. Zool. 2018, 79, 45. [Google Scholar] [CrossRef]
- Bellante, A.; Piazzese, D.; Cataldo, S.; Parisi, M.G.; Cammarata, M. Evaluation and comparison of trace metal accumulation in different tissues of potential bioindicator organisms: Macrobenthic filter feeders Styela plicata, Sabella spallanzanii, and Mytilus galloprovincialis. Environ. Toxicol. Chem. 2016, 35, 3062–3070. [Google Scholar] [CrossRef]
- Giangrande, A.; Licciano, M.; Del Pasqua, M.; Fanizzi, F.P.; Migoni, D.; Stabili, L. Heavy metals in five Sabellidae species (Annelida, Polychaeta): Ecological implications. Environ. Sci. Pollut. Res. 2017, 24, 3759–3768. [Google Scholar] [CrossRef]
- Xie, Z.-C.; Wong, N.C.; Qian, P.-Y.; Qiu, J.-W. Responses of polychaete Hydroides elegans life stages to copper stress. Mar. Ecol. Prog. Ser. 2005, 285, 89–96. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Thilagam, H.; Raja, P. Toxicity of heavy metals on embryogenesis and larvae of the marine sedentary polychaete Hydroides elegans. Arch. Environ. Contamin. Toxicol. 2007, 52, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Ross, K.E.; Bidwell, J.R. A 48-h larval development toxicity test using the marine polychaete Galeolaria caespitosa Lamarck (Fam. Serpulidae). Arch. Environ. Contamin. Toxicol. 2001, 40, 489–496. [Google Scholar]
- Giangrande, A.; Cavallo, A.; Licciano, M.; Mola, E.; Pierri, C.; Trianni, L. Utilization of the filter feeder polychaete Sabella. Aquac. Int. 2005, 13, 129–136. [Google Scholar] [CrossRef]
- Licciano, M.; Stabili, L.; Giangrande, A.; Cavallo, R.A. Bacterial accumulation by Branchiomma luctuosum (Annelida: Polychaeta): A tool for biomonitoring marine systems and restoring polluted waters. Mar. Environ. Res. 2007, 63, 291–302. [Google Scholar] [CrossRef]
- Pierri, C.; Fanelli, G.; Giangrande, A. Experimental co-culture of low food-chain organisms, Sabella spallanzanii (Polychaeta, Sabellidae) and Cladophora prolifera (Chlorophyta, Cladophorales), in Porto Cesareo area (Mediterranean Sea). Aquac. Res. 2006, 37, 966–974. [Google Scholar] [CrossRef]
- Stabili, L.; Licciano, M.; Giangrande, A.; Longo, C.; Mercurio, M.; Marzano, C.N.; Corriero, G. Filtering Activity of Spongia officinalis var. adriatica (Schmidt) (Porifera, Demospongiae) on bacterioplankton: Implications for bioremediation of polluted seawater. Water Res. 2006, 40, 3083–3090. [Google Scholar] [CrossRef]
- Stabili, L.; Licciano, M.; Lezzi, M.; Giangrande, A. Microbiological accumulation by the Mediterranean invasive alien species Branchiomma bairdi (Annelida, Sabellidae): Potential tool for bioremediation. Mar. Pollut. Bull. 2014, 86, 325–331. [Google Scholar] [CrossRef]
- Stabili, L.; Licciano, M.; Giangrande, A.; Gerardi, C.; De Pascali, S.A.; Fanizzi, F.P. First insight on the mucus of the annelid Myxicola infundibulum (Polychaeta, Sabellidae) as a potential prospect for drug discovery. Mar. Drugs 2019, 17, 396. [Google Scholar] [CrossRef]
- Soto, D. Integrated Mariculture: A Global Review.; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2009. [Google Scholar]
- Stewart, B.; Davies, B. Effects of macrophyte harvesting on invertebrates associated with Potamogeton pectinatus L. in the Marina Da Gama, Zandvlei, Western Cape. Trans. R. Soc. S. Africa 1986, 46, 35–49. [Google Scholar] [CrossRef]
- Joyce, C.B.; Vina-Herbon, C.; Metcalfe, D.J. Biotic variation in coastal water bodies in Sussex, England: Implications for saline lagoons. Estuar. Coast. Shelf Sci. 2005, 65, 633–644. [Google Scholar] [CrossRef]
- Davies, B.; Stuart, V.; De Villiers, M. The filtration activity of a serpulid polychaete population (Ficopomatus enigmaticus (Fauvel) and its effects on water quality in a coastal marina. Estuar. Coast. Shelf Sci. 1989, 29, 613–620. [Google Scholar] [CrossRef]
- Licciano, M.; Watson, G.J.; Murray, J.M.; Giangrande, A. Evidence of regenerative ability in Myxicola infundibulum (Annelida, Sabellida): Evolutionary and systematic implications. Invertebr. Biol. 2015, 134, 48–60. [Google Scholar] [CrossRef]
- Bely, A.E. Distribution of segment regeneration ability in the Annelida. Integr. Comp. Biol. 2006, 46, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Prentiss, N.K.; Tyler, M.S.; Dean, D.A. Morphological and histological investigation of the regeneration in Myxicola infundibulum (Montagu, 1808) (Sabellida, Annelida). Mar. Biol. Assoc. UK 2017, 97, 1155–1165. [Google Scholar] [CrossRef]
- Nicol, J. The giant nerve-fibres in the central nervous system of Myxicola (Polychaeta, Sabellidae). Q. J. Micros. Sci. 1948, 3, 1–45. [Google Scholar]
- Roberts, M.B.V. The Rapid response of Myxicola infundibulum (Grube). J. Mar. Biol. Assoc. U.K. 1962, 42, 527–539. [Google Scholar] [CrossRef]
- Gilbert, D. Axoplasm architecture and physical properties as seen in the Myxicola giant axon. J. Physiol. 1975, 253, 257–301. [Google Scholar] [CrossRef]
- Barry, J.F.; Turner, M.J.; Schloss, J.M.; Glenn, D.R.; Song, Y.; Lukin, M.D.; Park, H.; Walsworth, R.L. Optical magnetic detection of single-neuron action potentials using quantum defects in diamond. Proc. Natl. Acad. Sci. USA 2016, 113, 14133–14138. [Google Scholar] [CrossRef]
- Purschke, G. Annelida: Basal groups and Pleistoannelida. In Structure and Evolution of Invertebrate Nervous Systems; Schmidt-Rhaesa, A., Harzsch, S., Purschke, G., Eds.; Oxford University Press: Oxford, UK, 2016; pp. 254–312. [Google Scholar]
- Schauf, C.; Davis, F.A.; Marder, J. Effects of carbamazepine on the ionic conductances of Myxicola giant axons. J. Pharmacol. Exp. Ther. 1974, 189, 538–543. [Google Scholar]
- Nedved, B.T.; Hadfield, M.G. Hydroides elegans (Annelida: Polychaeta): A model for biofouling research. In Marine and Industrial Biofouling; Flemming, H.-S., Sriyutha Murthy, P., Venkatesan, R., Cooksey, K., Eds.; Springer Series on Biofilms: Los Angeles, CA, USA, 2008; Volume 4, pp. 203–218. [Google Scholar]
- Hadfield, M.G. The D.P. Wilson Lecture. Research on settlement and metamorphosis of marine invertebrate larvae: Past, present and future. Biofouling 1998, 12, 9–29. [Google Scholar] [CrossRef]
- Hadfield, M.G. Biofilms and marine invertebrate larvae: What bacteria produce that larvae use to choose settlement sites. Ann. Rev. Mar. Sci 2011, 3, 453–470. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.-Y. Larval settlement of polychaetes. Hydrobiologia 1999, 402, 239–253. [Google Scholar] [CrossRef]
- Hadfield, M.G.; Nedved, B.T.; Wilbur, S.; Koehl, M. Biofilm cue for larval settlement in Hydroides elegans (Polychaeta): Is contact necessary? Mar. Biol. 2014, 161, 2577–2587. [Google Scholar] [CrossRef]
- Shikuma, N.J.; Antoshechkin, I.; Medeiros, J.M.; Pilhofer, M.; Newman, D.K. Stepwise metamorphosis of the tubeworm Hydroides elegans is mediated by a bacterial inducer and MAPK Signaling. Proc. Natl. Acad. Sci. USA 2016, 113, 10097–10102. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, N.; Lema, K.A.; Nedved, B.T.; Hadfield, M.G. Microbiomes of the polychaete Hydroides elegans (Polychaeta: Serpulidae) across its life-history stages. Mar. Biol. 2019, 166, 19. [Google Scholar] [CrossRef]
- Toonen, R.J.; Pawlik, J.R. Foundations of gregariousness. Nature 1994, 370, 511–512. [Google Scholar] [CrossRef]
- Toonen, R.J.; Pawlik, J.R. Settlement of the gregarious tube worm Hydroides dianthus (Polychaeta: Serpulidae): Cues for gregarious settlement. Mar. Biol. 1996, 126, 725–733. [Google Scholar] [CrossRef]
- Toonen, R.; Pawlik, J. Settlement of the gregarious tube worm Hydroides dianthus (Polychaeta: Serpulidae). I. gregarious and nongregarious settlement. Mar. Ecol. Prog. Ser. 2001, 224, 103–114. [Google Scholar] [CrossRef]
- Kenny, N.J.; Shimeld, S.M. Additive multiple k-mer transcriptome of the keelworm Pomatoceros lamarckii (Annelida; Serpulidae) reveals annelid trochophore transcription factor cassette. Dev. Genes Evol. 2012, 222, 325–339. [Google Scholar] [CrossRef]
- Barton-Owen, T.B.; Szabó, R.; Somorjai, I.M.; Ferrier, D.E. A revised spiralian homeobox gene classification incorporating new polychaete transcriptomes reveals a diverse TALE class and a divergent hox gene. Genome Biol. Evol. 2018, 10, 2151–2167. [Google Scholar] [CrossRef] [PubMed]
- Szabó, R.; Ferrier, D.E.K. Cell proliferation dynamics in regeneration of the operculum head appendage in the annelid Pomatoceros lamarckii. J. Exp. Zool. Part. B Mol. Dev. Evol. 2014, 322, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Szabó, R.; Ferrier, D.E. Another biomineralising protostome with an msp130 gene and conservation of msp130 gene structure across Bilateria. Evol. Dev. 2015, 17, 195–197. [Google Scholar] [CrossRef]
- San Chan, V.B.; Li, C.; Lane, A.C.; Wang, Y.; Lu, X.; Shih, K.; Zhang, T.; Thiyagarajan, V. CO2-driven ocean acidification alters and weakens integrity of the calcareous tubes produced by the serpulid tubeworm, Hydroides elegans. PLoS ONE 2012, 7, e42718. [Google Scholar] [CrossRef]
- Lane, A.C.; Mukherjee, J.; Chan, V.B.; Thiyagarajan, V. Decreased pH does not alter metamorphosis but compromises juvenile calcification of the tube worm Hydroides elegans. Mar. Biol. 2013, 160, 1983–1993. [Google Scholar] [CrossRef][Green Version]
- Díaz-Castañeda, V.; Cox, T.E.; Gazeau, F.; Fitzer, S.; Delille, J.; Alliouane, S.; Gattuso, J.-P. Ocean acidification affects calcareous tube growth in adults and reared offspring of serpulid polychaetes. J. Exp. Biol. 2019, 222, jeb196543. [Google Scholar] [CrossRef]
- Wabnitz, C. From Ocean to Aquarium: The Global Trade in Marine Ornamental Species; UNEP/Earthprint: Cambridge, UK, 2003. [Google Scholar]
- Murray, J.M.; Watson, G.J.; Giangrande, A.; Licciano, M.; Bentley, M.G. Managing the marine aquarium trade: Revealing the data gaps using ornamental polychaetes. PLoS ONE 2012, 7, e29543. [Google Scholar] [CrossRef] [PubMed]
- Bybee, D.R.; Murray, J.M. Polychaetes. Mar. Ornam. Spec. Aquacult. 2017, 1, 565–579. [Google Scholar]
- Bybee, D.R.; Bailey-Brock, J.H.; Tamaru, C.S. Larval development of Sabellastarte spectabilis (Grube, 1878) (Polychaeta: Sabellidae) in Hawaiian waters. Sci. Mar. 2006, 70, 279–286. [Google Scholar] [CrossRef][Green Version]
- Bybee, D.R.; Bailey-Brock, J.H.; Tamaru, C.S. Evidence for sequential hermaphroditism in Sabellastarte spectabilis (Polychaeta: Sabellidae) in Hawai’i. Pac. Sci. 2006, 60, 541–547. [Google Scholar] [CrossRef]
- Bybee, D.R.; Bailey-Brock, J.H.; Tamaru, C.S. Gametogenesis and spawning periodicity in the fan worm Sabellastarte spectabilis (Polychaeta: Sabellidae). Mar. Biol. 2007, 151, 639–648. [Google Scholar] [CrossRef]
- Murray, J.M. Regeneration and Reproduction in Sabella pavonina (Savigny): Developing a Novel Method to Culture Marine Ornamental Sabellids. Ph.D. Thesis, University of Portsmouth, Portsmouth, UK, 2010. [Google Scholar]
- Dávila-Jiménez, Y.; Tovar-Hernández, M.A.; Simōes, N. The social feather duster worm Bispira brunnea (Polychaeta: Sabellidae): Aggregations, morphology and reproduction. Mar. Biol. Res. 2017, 13, 782–796. [Google Scholar] [CrossRef]
- Alalykina IL, Polychaeta. A review of the deep-sea benthic polychaetes along the NW Pacific. In Biogeographical Atlas of the Deep NW Pacific Fauna; Pensoft Publishers: Sofia, Bulgaria, 2020; pp. 175–212. [Google Scholar]
- Gunton, L.M.; Kupriyanova, E.K.; Alvestad, T.; Avery, L.; Blake, J.A.; Biriukova, O.; Böggemann, M.; Borisova, P.; Budaeva, N.; Burghardt, I.; et al. Annelids of the eastern Australian abyss collected by the 2017 RV ‘Investigator’voyage. ZooKeys 2021, 1020, 1–198. [Google Scholar] [CrossRef]
- Simon, C. The wonderful world of worms. Quest 2014, 10, 32–34. [Google Scholar]
- Ippolitov, A.; Rzhavsky, A. Tube morphology, ultrastructures and mineralogy in recent Spirorbinae (Annelida; Polychaeta; Serpulidae). I. General introduction. Tribe Paralaeospirini. Invertebr. Zool. 2014, 11, 293–314. [Google Scholar] [CrossRef]
- Sigvaldadóttir, E.; Mackie, A.S. Prionospio steenstrupi, P. fallax and P. dubia (Polychaeta, Spionidae): Re-evaluation of identity and status. Sarsia 1993, 78, 203–219. [Google Scholar] [CrossRef]
- Martin, D.; Britayev, T.A.; San Martín, G.; Gil, J. Inter-population variability and character description in the sponge-associated Haplosyllis spongicola complex (Polychaeta: Syllidae). In Advances in Polychaete Research; Springer: Dordrecht, The Netherlands, 2003; pp. 145–162. [Google Scholar]
- Ten Hove, H.A.; Jansen-Jacobs, M.J. A Revision of the genus Crucigera (Polychaeta; Serpulidae); A proposed methodical approach to serpulids, with special reference to variation in Serpula and Hydroides. In Proceedings of the First International Polychaete Conference; Hutchings, P.A., Ed.; Linnean Society of New South West: Sydney, Australia, 1984; pp. 143–180. [Google Scholar]
- Watanabe, M.E. The evolution of natural history collections: New research tools move specimens, data to center stage. Bioscience 2019, 69, 163–169. [Google Scholar] [CrossRef]
- Annenkova, N.P. Polychaeta of the North Japan Sea and their horizontal and vertical distribution. USSR Hydrobiological Expedition in 1934 to the Japan Sea. Trudy Daln. Filiala Akademii Nauk SSSR 1938, 1, 81–230. (In Russian) [Google Scholar]
- Oug, E.; Bakken, T.; Kongsrud, J.A. Original specimens and type localities of early described polychaete species (Annelida) from Norway, with particular attention to species described by O. F. Müller and M. Sars. Mem. Mus. Vic. 2014, 71, 217–236. [Google Scholar] [CrossRef][Green Version]
- Montagu, G. Testacea Brittanica or Natural History of British Shells, Marine, Land and Fresh-Water, Including the Most Minute: Systematically Arranged and Embellished with Figures. Part II; White: London, UK, 1803; pp. 293–606. [Google Scholar]
- Von Siebold, C.T. Bericht Über die Leistungen in der Naturgeschichte der Würmer, Zoophyten und Protozoen Während der Jahre 1845, 1846 und 1847. Arch. Naturgesch. 1850, 16, 351–468. [Google Scholar]













| Feature | Fabriciidae | Sabellidae | Serpulidae |
|---|---|---|---|
| Tube material | mucus and sediment/none | mucus and sediment * | calcium carbonate |
| Radiolar lobes | separated | fused | separated |
| Vacuolated cells supporting radioles | absent | present | absent |
| Operculum | absent | absent | absent or present |
| Thoracic membrane | absent | absent | present |
| Thoracic uncini | acicular | avicular ** | avicular |
| Number of abdominal chaetigers | usually three (exceptionally two or four) | more than three | more than three |
| Abdominal uncini | with elongate and wide handle (manubrium) | with short handle or lacking handles | lacking handles |
| Branchial hearts | present | absent | absent |
| Genera | Number of Species * | Taxonomic Revision | Phylogenetic Studies |
|---|---|---|---|
| Augeneriella Banse, 1957 | 5 | Banse 1957, Fitzhugh 1983, 1990a, 1993 | Fitzhugh 1991a-b, 1992, 1993, 2010 |
| Bansella Fitzhugh, 2010 | 1 | Fitzhugh 2010 | Fitzhugh 2010 |
| Brandtika Jones, 1974 | 1 | Fitzhugh 2001 | NO |
| Brifacia Fitzhugh, 1998 | 2 | Fitzhugh 1998; Giangrande et al. 2014 | Fitzhugh 2010 |
| Echinofabricia Huang, Fitzhugh and Rouse, 2011 | 4 | Huang et al. 2011 | Fitzhugh 1991a, 1992, 1993, 1998, 2010 (as genus A) |
| Fabricia de Blainville, 1828 | 1 | Fitzhugh 1991b, 2010 | Fitzhugh 1991a, 1992, 1993, 1998, 2010 |
| Fabricinuda Fitzhugh, 1990b | 7 | Fitzhugh 1983, 1990b, 2002a; López and Rodríguez 2008 | Fitzhugh 1991a, 1992, 1993, 1998, 2002a, 2010; López and Rodríguez 2008 |
| Fabriciola Friedrich, 1939 | 6 | Fitzhugh 1991b, 1992, 1999; Bick 2005 | Fitzhugh 1991a, 1992, 1993, 1998, 1999, 2010 |
| Manayunkia Leidy, 1859 | 10 | Sitnikova et al. 2014, Atkinson et al. 2020 | Fitzhugh 1991a, 1992, 1993, 1998, 2010; Sitnikova et al. 2014; Pudovkina et al. 2016 |
| Monroika Hartman, 1951 | 1 | Fitzhugh 1992 | Fitzhugh 1992, 1998, 2010 |
| Novafabricia Fitzhugh, 1990c | 11 | Fitzhugh 1983, 1990c, 1998; Bick 2005 | Fitzhugh 1991a, 1992, 1993, 1998, 2010 |
| Parafabricia Fitzhugh, 1992 | 2 | Fitzhugh 1992 | Fitzhugh 1992, 1993, 1998, 2010 |
| Pseudoaugeneriella Fitzhugh, 1998 | 5 | Fitzhugh 1998 | Fitzhugh 1998, 2010 |
| Pseudofabricia Cantone, 1972 | 1 | Fitzhugh 1995 | Fitzhugh 1992, 1998, 2010 |
| Pseudofabriciola Fitzhugh, 1990b | 13 | Fitzhugh 1990c, 1991b, 1993, 1996, 2002a; Fitzhugh et al. 1994, Fitzhugh and Simboura 1995 | Fitzhugh 1991a-b, 1993, 1994, 1996, 1998, 2002a, 2010; Fitzhugh et al. 1994, Fitzhugh and Simboura 1995 |
| Raficiba Fitzhugh, 2001 | 1 | Fitzhugh 2001 | Fitzhugh 2010 |
| Rubifabriciola Huang, Fitzhugh and Rouse, 2011 | 10 | Huang et al. 2011 | Huang et al. 2011 |
| Genera | Number of Species * | Revision | Phylogenetic Analyses |
|---|---|---|---|
| Acromegalomma Gil and Nishi, 2017 | 38 | Perkins 1984, Tovar-Hernández and Salazar-Vallejo 2008; Capa and Murray 2009; Tovar-Hernández and Carrera-Parra 2011, Gil and Nishi 2017 | Capa and Murray 2009, Tovar-Hernández and Carrera-Parra 2011 |
| Amphicorina Claparède, 1864 | 46 | Rouse 1990 (as Oriopsis), Cochrane 2003 | Cochrane 2003 |
| Amphiglena Claparède, 1864 | 14 | Capa and Rouse 2007; Tilic et al. 2019 | Capa and Rouse 2007, Tilic et al. 2019 |
| Anamobaea Krøyer, 1856 | 2 | Tovar-Hernández et al. 2020 | NO |
| Aracia Nogueira, Fitzhugh and Rossi, 2004 | 3 | Nogueira et al. 2004; Tovar-Hernández 2014 | Nogueira et al. 2010 |
| Bispira Krøyer, 1856 | 24 | Knight-Jones and Perkins 1998; Capa 2008 | Capa 2008 |
| Branchiomma Kölliker, 1859 | 30 | Tovar-Hernández and Knight-Jones 2006 | Capa et al. 2013; del Pasqua et al. 2018 |
| Caobangia Giard, 1893 | 7 | Jones 1974 | NO |
| Chone Krøyer, 1856 | 20 | Cochrane 2003, Tovar-Hernández 2005, 2006, 2007a, b, c, 2008 | Tovar-Hernández 2008 |
| Claviramus Fitzhugh, 2002 | 5 | Fitzhugh 2002; Nishi et al. 2019 | NO |
| Dialychone Claparède, 1869 | 19 | Tovar-Hernández 2008 | Tovar-Hernández 2008 |
| Euchone Malmgren, 1866 | 35 | Cochrane 2003, Giangrande and Licciano 2006, Giangrande et al. 2017 | Cochrane 2003 |
| Euchoneira Licciano, Giangrande and Gambi, 2009 | 1 | Licciano et al. 2009 | Licciano et al. 2009 |
| Eudistylia Bush, 1905 | 5 | Hartman 1938, Banse 1979 | NO |
| Hypsicomus Grube, 1870 | 1 | Perkins 1984 | NO |
| Jasmineira Langerhans, 1880 | 19 | Cochrane 2003, Capa and Murray 2015 | Cochrane 2003 |
| Laonome Malmgren, 1866 | 10 | Fitzhugh 2002, Capa 2007, Bick et al. 2018 | Capa 2007 |
| Notaulax Tauber, 1879 | 26 | Perkins 1984 | NO |
| Paradialychone Tovar-Hernández, 2008 | 16 | Tovar-Hernández 2008 | Tovar-Hernández 2008 |
| Parasabella Bush, 1905 | 29 | Perkins 1984, Giangrande 1994, Tovar-Hernández and Harris 2010, Capa and Murray 2015b, Keppel et al. 2020 | Capa and Murray 2015b |
| Perkinsiana Knight-Jones, 1983 | 16 | Knight-Jones 1983, Giangrande and Gambi 1997, Tovar-Hernández et al. 2012 | NO |
| Potamethus Chamberlin, 1919 | 11 | Knight-Jones 1983 | NO |
| Potamilla Malmgren, 1866 | 10 | Knight-Jones 1983 | NO |
| Potaspina Hartman, 1969 | 2 | Capa 2007 | Capa 2007 |
| Pseudobranchiomma Jones, 1962 | 19 | Knight-Jones 1994, Knight-Jones and Giangrande 2003 | Capa and Murray 2016 |
| Pseudopotamilla Bush, 1904 | 23 | Knight-Jones et al. 2017 | Capa 2007 |
| Sabella Linnaeus, 1767 | 39 | Knight-Jones and Perkins 1998 | NO |
| Sabellastarte Krøyer, 1856 | 8 | Knight-Jones and Mackie 2003 | Capa et al. 2010 |
| Sabellomma Nogueira, Fitzhugh and Rossi, 2010 | 4 | Nogueira et al. 2010, Capa and Murray 2015 | Nogueira et al. 2010 |
| Stylomma Knight-Jones, 1997 | 2 | Knight-Jones and Perkins 1998, Capa 2007 | Capa 2007 |
| Terebrasabella Fitzhugh and Rouse, 1999 | 3 | Murray and Rouse 2007 | Murray and Rouse 2007 |
| Genera | Number of Species * | Revision | Phylogenetic Analyses |
|---|---|---|---|
| Bathyditrupa Kupriyanova, 1993 | 1 | Kupriyanova and Ippolitov 2015 | NO |
| Bathyvermilia Zibrowius, 1973 | 7 | Zibrowius 1973 | NO |
| Crucigera Benedict, 1887 | 5 | ten Hove and Jansen-Jacobs 1984 | Kupriyanova et al. 2008 |
| Ditrupa Berkeley, 1835 | ten Hove and Smith 1990 | NO | |
| Ficopomatus Southern, 1921 | 6 | ten Hove and Weerdenburg 1978 | Kupriyanova et al. 2009; Styan et al. 2017 |
| Galeolaria Lamarck, 1818 | 3 | NO | Halt et al. 2009; Smith et al. 2012 |
| Hydroides Gunnerus, 1768 | 99 | Bastida-Zavala and ten Hove 2002, 2003; Sun et al. 2015; Sun et al. 2018 | Sun et al. 2018 |
| Laminatubus ten Hove and Zibrowius, 1986 | 3 | Rouse and Kupriyanova 2021 | Rouse and Kupriyanova 2021 |
| Marifugia Absolon and Hrabe, 1930 | 1 | Kupriyanova et al. 2009 | Kupriyanova et al. 2009 |
| Metavermilia Bush, 1905 | 15 | Zibrowius 1971; Nishi et al. 2007 | NO |
| Pseudochitinopoma Zibrowius, 1969 | 5 | Kupriyanova et al. 2012 | NO |
| Pyrgopolon de Montfort, 1808 | 3 | ten Hove 1973 | NO |
| Serpula Linnaeus, 1758 | 30 | NO | Kupriyanova et al. 2008 |
| Spirobranchus de Blainville, 1818 | 36 | ten Hove 1970 | Willette et al. 2015; Perry et al. 2019; Pazoki et al. 2020 |
| Spiraserpula Regenhardt, 1961 | 18 | Pillai and ten Hove 1994 | NO |
| Spirodiscus Fauvel, 1909 | 2 | Kupriyanova and Nishi 2011, Kupriyanova and Ippolitov 2015 | NO |
| Taxon | Depth (m) | References |
|---|---|---|
| FABRICIIDAE | ||
| Fabriciola sp. (Okhotsk Sea) | >2000 | Alalykina 2020 |
| Fabriciola sp. | 4600 | Baumhaker 2012 |
| Novafabricia sp. | 4600 | Baumhaker 2012 |
| SABELLIDAE | ||
| Jasmineira bermudensis Hartman, 1965 | 1000 | Original description |
| Potaspina australiensis Capa, 2007 | 1000 | Original description |
| Euchone magna (Fauchald, 1972) | 1071 | Original description |
| Perkinsiana assimilis (McIntosh, 1885) | 1100 | Original description |
| Chone gracilis Moore, 1906 | 1244 | Méndez 2006 |
| Bispira wireni (Johansson, 1922) | 1335 | Capa et al. 2013 |
| Potamethus filiformis Hartmann-Schröder, 1977 | 1430 | Original description |
| Pseudopotamilla intermedia Moore, 1905 | 1682 | Original description |
| Bispira sp. (Costa Rica) | 1887 | Goffredi et al. 2020 |
| Chone sp. (Okhotsk Sea and N. Pacific abyss) | >2000 | Alalykina 2020 |
| Euchone sp. (Okhotsk Sea and N. Pacific abyss) | >2000 | Alalykina 2020 |
| Jasmineira sp. (Okhotsk Sea and N. Pacific abyss) | >2000 | Alalykina 2020 |
| Potamethus sp. 1 (Okhotsk Sea and N. Pacific abyss) | >2000 | Alalykina 2020 |
| Potamethus sp. 2 (Okhotsk Sea) | >2000 | Alalykina 2020 |
| Potamethus singularis Hartman, 1965 | 2000 | Original description |
| Potamilla neglecta (Sars, 1851) | 2030 | Hansen 1882 |
| Potamethus malmgreni (Hansen, 1878) | 2222 | Original description |
| Euchone cf. incolor Hartman, 1965 | 2500 | Alalykina 2020 |
| Fabrisabella similis Fauchald, 1972 | 2520 | Original description |
| Potamethus scotiae (Pixell, 1913) | 2578 | Original description |
| Euchone papillosa (Sars, 1851) | 2900 | Uschakov 1955; Levenstein 1969 |
| Jasmineria pacifica Annenkova, 1937 | 2900 | Original description |
| Jasmineira schaudinni Augener, 1912 | 3500 | Augener 1912 (abyssal, no depth given); Jirkov 1982, 2001 |
| Chone infundibuliformis Krøyer, 1856 | 3521 | Wesenberg-Lund 1950 |
| cf Sabellidae species 1 (Clarion-Clipperton Zone) | 4029 | Amon et al. 2017 |
| Potamethus mucronatus (Moore, 1923) | 4131 | Original description, as Notaulax |
| Potamilla abyssicola Uschakov, 1952 | 4200 | Original description; Levenstein 1961 1969; Alalykina 2020 |
| Potamethus spathiferus (Ehlers, 1887) | 4360 | Fauvel 1914 |
| Euchone incolor Hartman, 1965 | 4862 | Original description; Hartman 1971 |
| ?Potamethus sp. Mozambique Basin | 5068 | Hartman 1971 |
| Potamehus dubius (Eliason, 1951) | 5860 | Original description |
| Jasmineira sp. Japan | 6207 | Levenstein 1961b |
| Potamethus singularis Hartman, 1965 | 6023 | Original description; Hartman 1971 |
| Sabellidae sp. Pacific Ocean | 8042 | Levenstein 1969; Lemche et al. 1976 |
| Potamilla sp. Kurile-Kamchatka Trench | 8100 | Uschakov 1952; Belyaev 1989 |
| Jasmineira sp. Kermadec Trench Trench | 8300 | Kirkegaard 1956; Hartman and Fauchald 1971; Belayev 1972 |
| Potamethus sp. Izu-Bonin Trench | 8735 | Belayev 1989 |
| Jasmineira filitovae Levenstein, 1961 (as Potamethus) | 9735 | Levenstein 1969, 1973; Belyaev 1989 |
| SERPULIDAE | ||
| Laminatubus joicebrooksae Rouse and Kupriyanova 2021 | 1011 | Original description |
| Hyalopomatus madreporae Sanfilippo, 2009 | 1146 | Original description |
| Neovermilia falcigera (Roule, 1898) | 1580 | Zibrowius and ten Hove 1987 |
| Metavermilia ogasawaraensis Nishi, Kupriyanova and Tachikawa, 2007 | 1603 | Original description |
| Zibrovermilia zibrowii Kupriyanova and Ippolitov, 2015 | 1710 | Original description |
| Hyalopomatus dieteri Kupriyanova and Ippolitov, 2015 | 1980 | Original description |
| Hyalopomatus biformis (Hartman, 1960) | 1982 | Kupriyanova and Nishi 2010 |
| Metavermilia zibrowii Bailey-Brock and Magalhães, 2012 | 2013 | Original description |
| Vermiliopsis notialis Monro, 1930 | 2016 | Averintsev 1974 |
| Bushiella (Jugaria) atlantica (Knight-Jones, 1978) | 2100 | Original description |
| Bathyvermilia islandica Sanfilippo, 2001 | 2399 | Original description |
| Filogranula stellata (Southward, 1963) | 2464 | Ben-Eliahu and Fiege 1996 |
| Hyalopomatus variorugosus Ben-Eliahu and Fiege, 1996 | 2474 | Original description |
| Laminatubus paulbrooksi Rouse and Kupriyanova, 2021 | 2478 | Original description |
| Protis browni (Pixell, 1913) | 2585 | Original description |
| Protis hydrothermica ten Hove and Zibrowius 1986 | 2620 | ten Hove and Zibrowius 1986 |
| Hyalopomatus marenzelleri Langerhans, 1884 | 2800 | Zibrowius 1968, 1969, 1977 |
| Laminatubus alvini ten Hove and Zibrowius, 1986 | 2842 | Original description |
| Neovermilia cf. sphaeropomata (Benham, 1927) | 3261 | Rouse and Kupriyanova 2021 |
| Apomatus globifer Théel, 1878 | 3384 | Uschakov 1957 |
| Hyalopomatus claparedii Marenzeller, 1878 | 3550 | Kupriyanova and Jirkov 1997 |
| Hyalopomatus jirkovi Kupriyanova, 1993 | 3949 | Kupriyanova et al. 2011 |
| Spirodiscus grimaldii Fauvel, 1909 | 4124 | ten Hove and Kupriyanova 2009 |
| Bathyvermilia kupriyanovae Bastida-Zavala, 2008 | 4190 | Original description |
| Apomatus similis Marion and Bobretzky, 1875 | 4400 | Fauvel (1909), Fauvel (1914) |
| Spirodiscus groenlandicus (McIntosh, 1877) | 4440 | Kupriyanova and Ippolitov 2015 |
| Bathyvermilia zibrowiusi Kupriyanova, 1993 | 4550 | Kupriyanova et al. 2011 |
| Hyalopomatus sikorskii Kupriyanova, 1993 | 4550 | Kupriyanova et al. 2011 |
| Protis simplex Ehlers, 1887 | 4810 | Knight-Jones et al. (1997) |
| Protis polyoperculata Kupriyanova, 1993 | 5110 | Original description |
| Hyalopomatus mironovi Kupriyanova, 1993 | 5216 | Rouse and Kupriyanova 2021 |
| Protis arctica (Hansen, 1879) | 5300 | Zibrowius 1969 |
| Bathyvermilia challengeri Zibrowius, 1973 | 5719 | Original description |
| Bathyvermilia langerhansi Fauvel, 1909 | 5987 | Eliason 1951 |
| Bathyvermilia gregrousei Kupriyanova and Ippolitov, 2015 | 6050 | Original description |
| Nidificaria levensteinae (Bailey-Brock and Knight-Jones, 1977) | 6096 | Original description |
| Bathyditrupa hovei Kupriyanova, 1993 | 6330 | Kupriyanova et al. 2011 |
| Protis sp. 2 | 8345 | Kupriyanova et al. 2014 |
| Protis sp. 1 | 9735 | Kupriyanova et al. 2014 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capa, M.; Kupriyanova, E.; Nogueira, J.M.d.M.; Bick, A.; Tovar-Hernández, M.A. Fanworms: Yesterday, Today and Tomorrow. Diversity 2021, 13, 130. https://doi.org/10.3390/d13030130
Capa M, Kupriyanova E, Nogueira JMdM, Bick A, Tovar-Hernández MA. Fanworms: Yesterday, Today and Tomorrow. Diversity. 2021; 13(3):130. https://doi.org/10.3390/d13030130
Chicago/Turabian StyleCapa, María, Elena Kupriyanova, João Miguel de Matos Nogueira, Andreas Bick, and María Ana Tovar-Hernández. 2021. "Fanworms: Yesterday, Today and Tomorrow" Diversity 13, no. 3: 130. https://doi.org/10.3390/d13030130
APA StyleCapa, M., Kupriyanova, E., Nogueira, J. M. d. M., Bick, A., & Tovar-Hernández, M. A. (2021). Fanworms: Yesterday, Today and Tomorrow. Diversity, 13(3), 130. https://doi.org/10.3390/d13030130