
Effects of Pure and Mixed Pine and Oak Forest Stands on Carabid Beetles


Abstract
1. Introduction
- Differences in activity density, body size and sex ratios can be expected at the level of forest stand types for the five Carabid species analysed (Carabus arvensis (Hbst. 1784), C. coriaceus (L. 1758), C. hortensis (L. 1758), C. violaceus (L. 1758), Calosoma inquisitor (L. 1758)).
- Tree species-specific characteristics, such as proportions of crown cover, and the nature of the tree species spatial distribution (random, regular or aggregated) are suitable parameters to highlight the affinities of Carabid beetles, especially for the mixed oak and pine forest stand types.
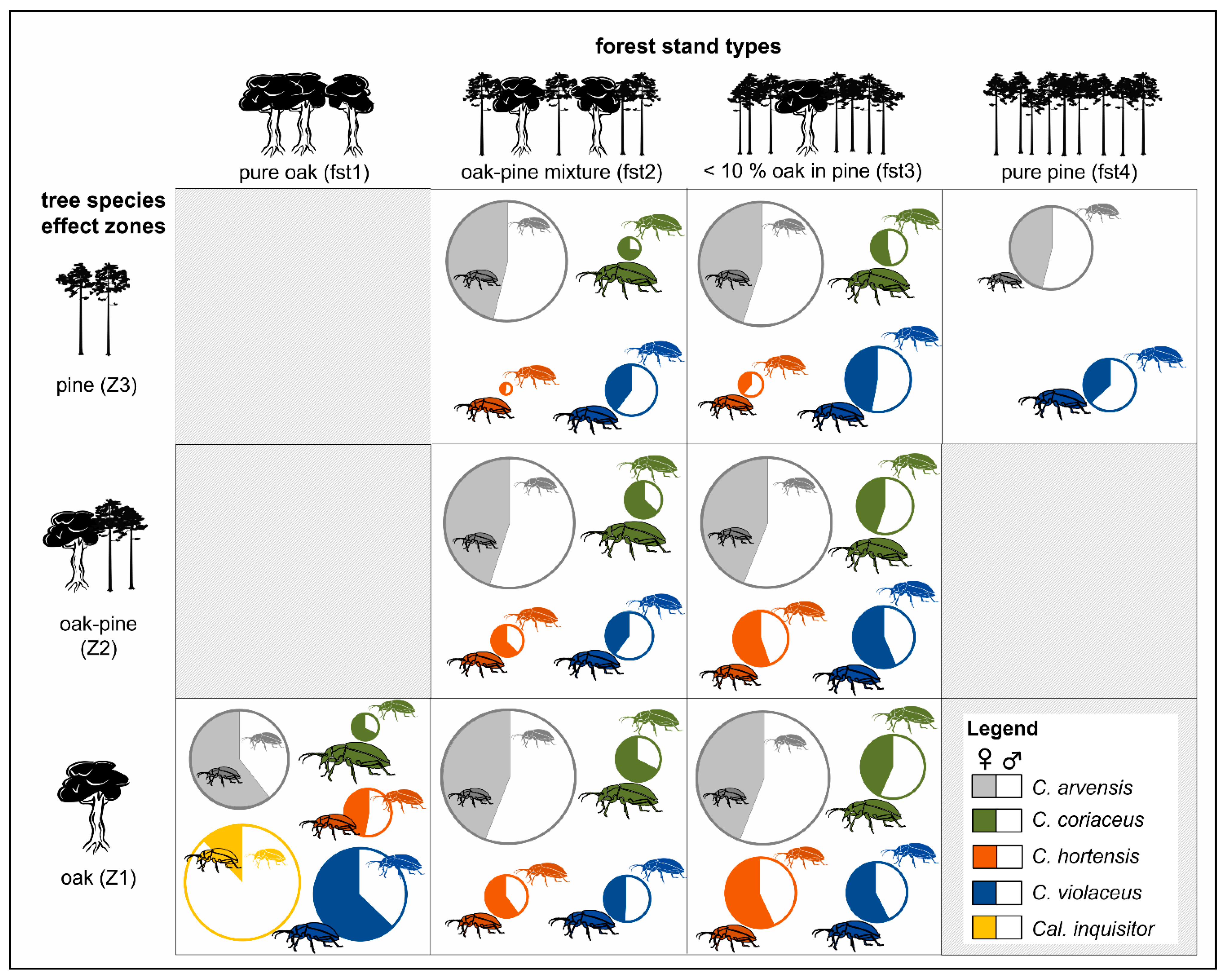
- Superordinate analyses are possible, where tree species effect zones (Z1—pure oak effect zone, Z2—mixed oak–pine effect zone, Z3—pure pine effect zone) are used to define small-scale habitat preferences and environmental niches of Carabid species.
2. Materials and Methods
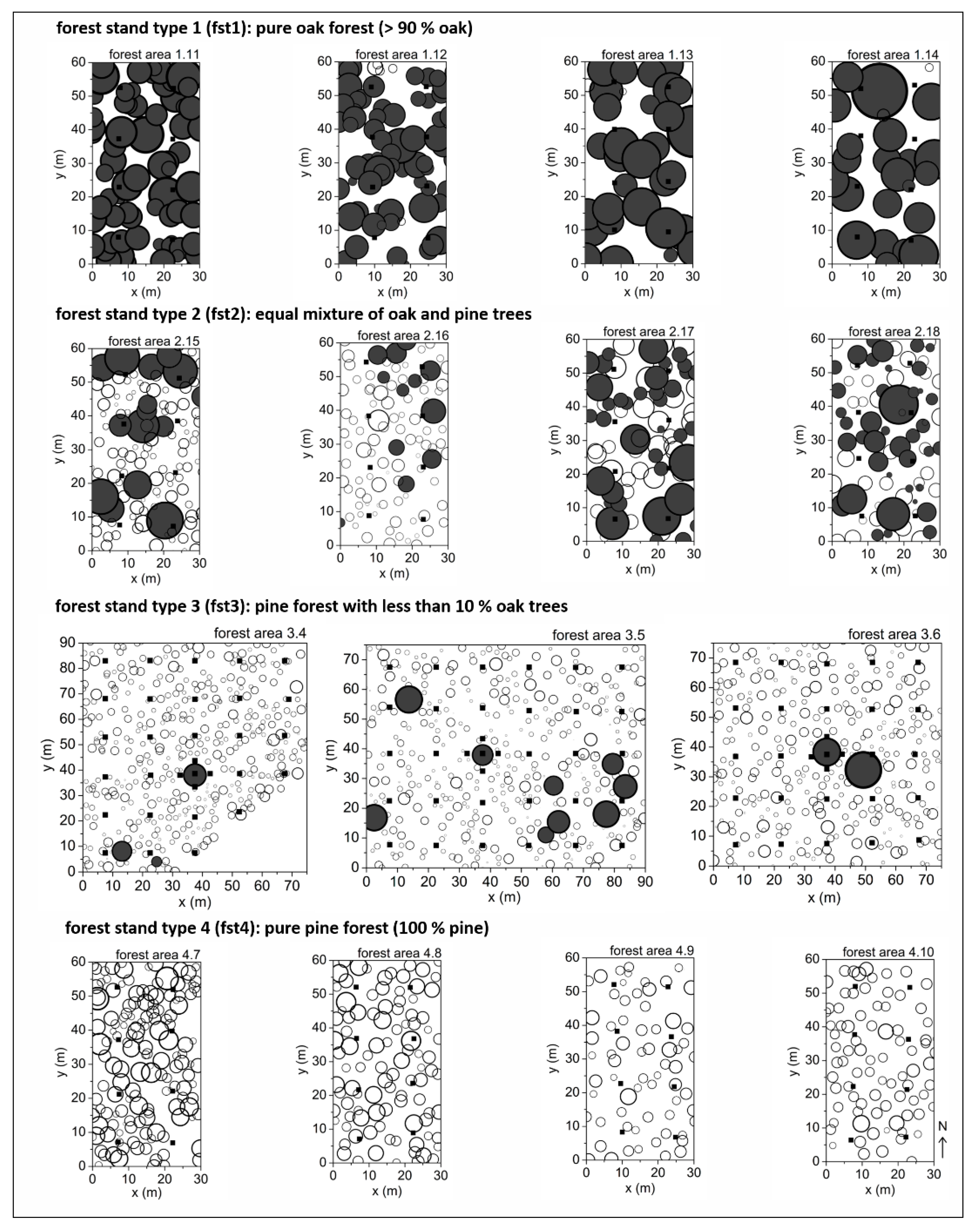
2.1. Study Areas and Tree Species Parameters
2.2. Trap Design and Beetle Parameters
2.3. Aggregation Index and Statistical Analyses
3. Results
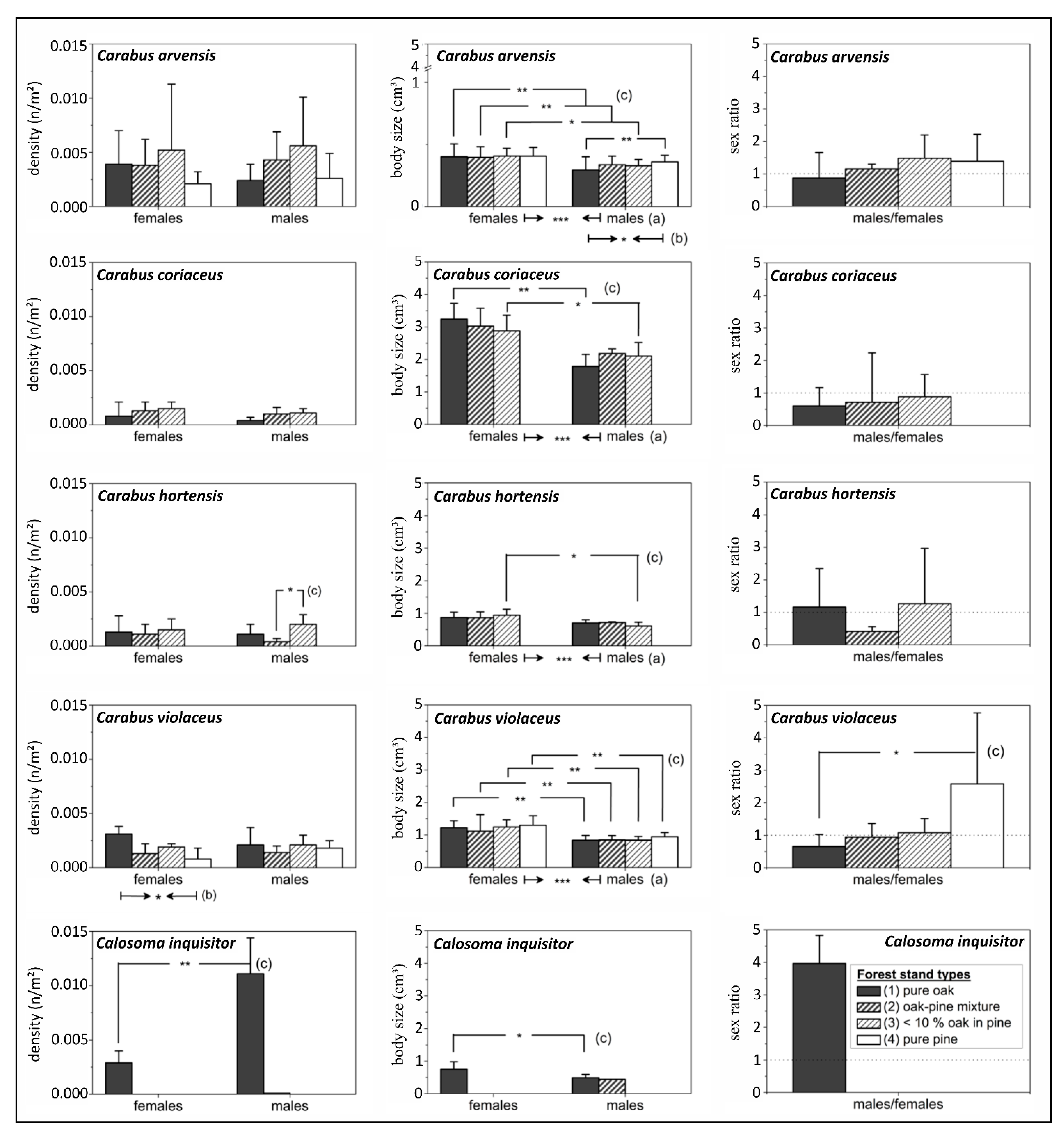
3.1. Effects of Forest Stand Types, Tree Species Proportions and Effect Zones
3.2. Model-Based Effects of Forest Stand Types and Different Structural Attributes
3.2.1. Modelling the Effects of Forest Stand Types and Oak Crown Cover
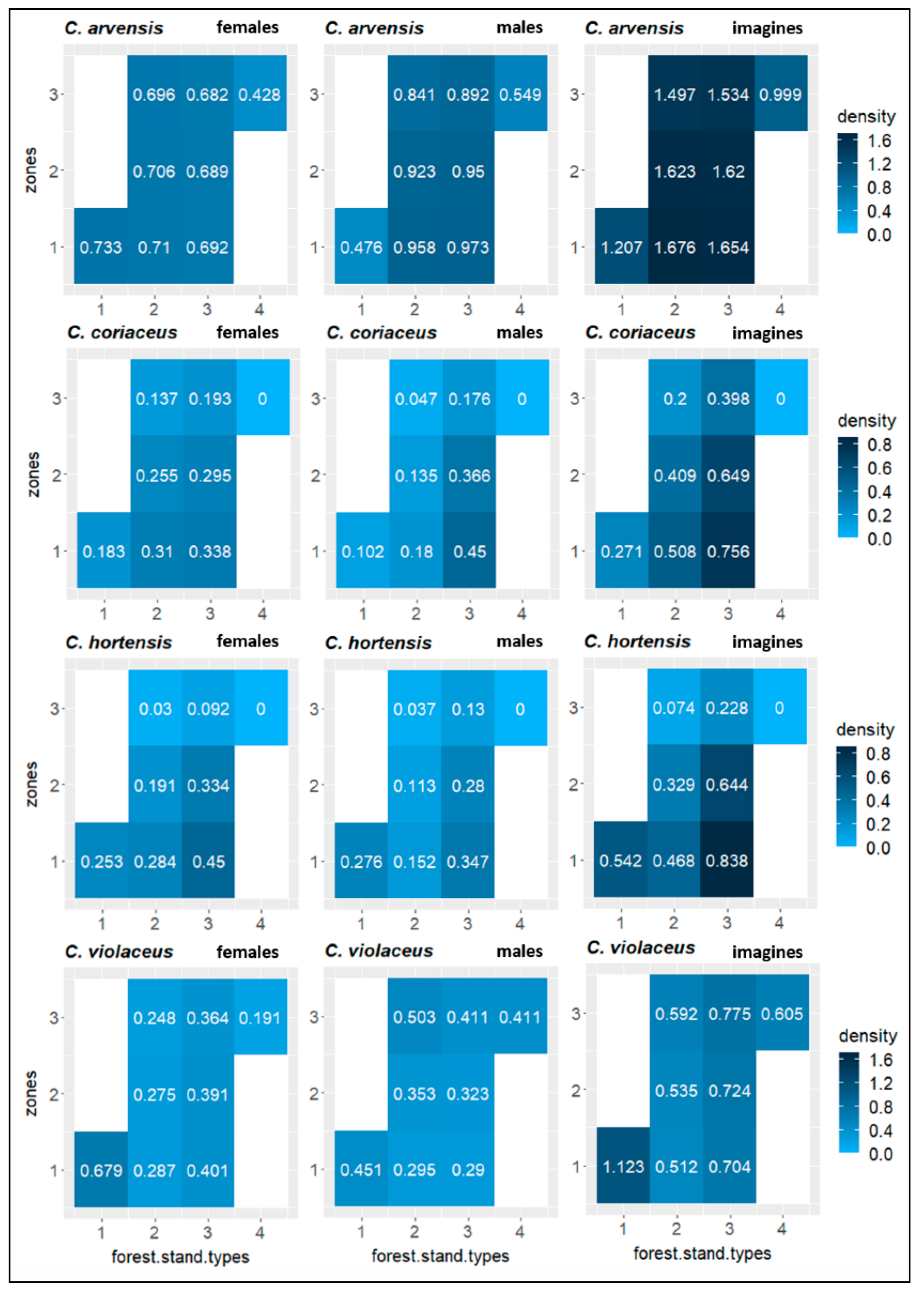
3.2.2. Modelling the Effects of Forest Stand Types and Effect Zones

4. Discussion
4.1. Spatial Scale of Pure and Mixed Forest Stand Types
4.2. Spatial Scale of Single Trees and their Ecological Effect Zones
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Oxbrough, A.; French, V.; Irwin, S.; Kelly, T.C.; Smiddy, P.; O’Halloran, J. Can mixed species stands enhance arthropod diversity in plantation forests? For. Ecol. Manag. 2012, 270, 11–18. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Pretzsch, H.; del Río, M. Dynamics, Silviculture and Management of Mixed Forests; Managing Forest Ecosystems Vol 31; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Anders, S.; Beck, W.; Bolte, A.; Hofmann, G.; Jenssen, M.; Krakau, U.; Müller, J. Ökologie und Vegetation der Wälder Nordostdeutschlands.—Einfluß von Niederschlagsarmut und erhöhtem Stickstoffeintrag auf Kiefern-, Eichen- und Buchen-Wald- und Forstökosysteme des Nordostdeutschen Tieflandes, 1st ed.; Verlag Dr. Kessel: Oberwinter, Germany, 2002. [Google Scholar]
- Goris, R.; Kint, V.; Haneca, K.; Geudens, G.; Beeckman, H.; Verheyen, K. Long-term dynamics in a planted conifer forest with spontaneous ingrowth of broad-leaved trees. Appl. Veg. Sci. 2007, 10, 219–228. [Google Scholar] [CrossRef]
- Pretzsch, H.; Steckel, M.; Heym, M.; Biber, P.; Ammer, C.; Ehbrecht, M.; Bielak, K.; Bravo, F.; Ordóñez, C.; Collet, C.; et al. Stand growth and structure of mixed-species and monospecific stands of Scots pine (Pinus sylvestris L.) and oak (Q. robur L., Quercus petraea (Matt.) Liebl.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2020, 139, 349–367. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Pretzsch, H.; Ammer, C.; Andenmatten, E.; Barbati, A.; Barreiro, S.; Brang, P.; Bravo, F.; Coll, L.; Corona, P.; et al. European Mixed Forests: Definition and research perspectives. For. Syst. 2014, 23, 518–533. [Google Scholar] [CrossRef]
- Jactel, H.; Brockerhoff, E.G. Tree diversity reduces herbivory by forest insects. Ecol. Lett. 2007, 10, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Knoke, T.; Ammer, C.; Stimm, B.; Mosandl, R. Admixing broadleaved to coniferous tree species: A review on yield, ecological stability and economics. Eur. J. For. Res. 2008, 127, 89–101. [Google Scholar] [CrossRef]
- Szmyt, J.; Tarasiuk, S. Species-specific spatial structure, species coexistence and mortality pattern in natural, uneven-aged Scots pine (Pinus sylvestris L.)-dominated forest. Eur. J. For. Res. 2018, 137, 1–16. [Google Scholar] [CrossRef]
- Butterfield, J.; Benitez Malvido, J. Effect of mixed-species tree planting on the distribution of soil invertebrates. In The Ecology of Mixed-Species Stands of Trees; Cannell, M.G.R., Malcolm, D.C., Robertson, P.A., Eds.; Special Publication Number 11 of the British Ecological Society; Oxford-Blackwell Scientific Publications: London, UK, 1992; pp. 255–265. [Google Scholar]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef]
- Sobek, S.; Steffan-Dewenter, I.; Scherber, C.; Tscharntke, T. Spatiotemporal changes of beetle communities across a tree diversity gradient. Divers. Distrib. 2009, 15, 660–670. [Google Scholar] [CrossRef]
- du Bus de Warnaffe, G.; Dufrêne, M. To what extent can management variables explain species assemblages? A study of carabid beetles in forests. Ecography 2004, 27, 701–714. [Google Scholar] [CrossRef]
- Lassau, S.A.; Hochuli, D.F.; Cassis, G.; Reid, C.A.M. Effects of Habitat Complexity on Forest Beetle Diversity: Do Functional Groups Respond Consistently? Divers. Distrib. 2005, 11, 73–82. [Google Scholar] [CrossRef]
- Rainio, J.; Niemelä, J. Ground beetles (Coleoptera: Carabidae) as bioindicators. Biodivers. Conserv. 2003, 12, 487–506. [Google Scholar] [CrossRef]
- Work, T.T.; Koivula, M.; Klimaszewski, J.; Langor, D.; Sweeney, J.; Hébert, C. Evaluation of carabid beetles as indicators of forest change in Canada. Can. Entomol. 2008, 140, 393–414. [Google Scholar] [CrossRef]
- Koivula, M. Useful model organisms, indicators, or both? Ground beetles (Coleoptera, Carabidae) reflecting environmental conditions. ZooKeys 2011, 100, 287–317. [Google Scholar] [CrossRef]
- Walsh, P.J.; Day, K.R.; Leather, S.R.; Smith, A. The Influence of Soil Type and Pine Species on the Carabid Community of a Plantation Forest with a History of Pine Beauty Moth Infestation. Forestry 1993, 66, 135–146. [Google Scholar] [CrossRef]
- Pearce, J.L.; Venier, L.A. The use of ground beetles (Coleoptera: Carabidae) and spiders (Araneae) as bioindicators of sustainable forest management: A review. Ecol. Indic. 2006, 6, 780–793. [Google Scholar] [CrossRef]
- Lindroth, C.H. Ground Beetles (Carabidae) of Fennoscandia. A Zoogeographic Study. Part 1: Specific Knowledge Regarding the Species. Translation of: Die fennoskandischen Carabidae: Eine Tiergeographische Studie I. Spezieller Teil; Amerind Publishing Co. Pvt. Ltd.: New Delhi, India, 1992. [Google Scholar]
- Skłodowski, J.J.W. Interspecific body size differentiation in Carabus assemblages in the Białowieża Primeval Forest, Poland. In Proceedings of the 11th European Carabidologists’ Meeting, Århus, Denmark, 21–24 July 2003; Lövei, G.L., Toft, S., Eds.; DIAS Report, No. 114. Ministry of Food, Agriculture and Fisheries: Tjele, Denmark, 2005; pp. 291–303. [Google Scholar]
- Dahl, F. Die Tierwelt Deutschlands und der Angrenzenden Meeresteile nach ihren Merkmalen und Lebensweise 7.Teil: Coleoptera oder Käfer I: Carabidae (Laufkäfer), 1st ed.; Verlag von Gustav Fischer: Jena, Germany, 1928. [Google Scholar]
- Turin, H.; Penev, L.; Casale, A. The Genus Carabus in Europe. A Synthesis; Fauna Europaea Evertebrata No. 2; Pensoft Publishers & European Invertebrates Survey: Sofia, Bulgaria, 2003. [Google Scholar]
- Thiele, H.-U. Carabid Beetles in Their Environments: A Study on Habitat Selection by Adaptations in Physiology and Behaviour; Zoophysiology and Ecology Volume 10; Springer: Berlin/Heidelberg, Germany, 1977. [Google Scholar]
- Stork, N.E. The Role of Ground Beetles in Ecological and Environmental Studies; Intercept Ltd.: Andover, MA, USA, 1990. [Google Scholar]
- Sroka, K.; Finch, O.-D. Ground beetle diversity in ancient woodland remnants in north-western Germany (Coleoptera, Carabidae). J. Insect Conserv. 2006, 10, 335–350. [Google Scholar] [CrossRef]
- Lange, M.; Türke, M.; Pašalić, E.; Boch, S.; Hessenmöller, D.; Müller, J.; Prati, D.; Socher, S.A.; Fischer, M.; Weisser, W.W.; et al. Effects of forest management on ground-dwelling beetles (Coleoptera; Carabidae, Staphylinidae) in Central Europe are mainly mediated by changes in forest structure. For. Ecol. Manag. 2014, 329, 166–176. [Google Scholar] [CrossRef]
- Taboada, A.; Kotze, D.J.; Tárrega, R.; Salgado, J.M. Traditional forest management: Do carabid beetles respond to human-created vegetation structures in an oak mosaic landscape? For. Ecol. Manag. 2006, 237, 436–449. [Google Scholar] [CrossRef]
- Clarke, R.T.; Thomas, J.A.; Elmes, G.W.; Hochberg, M.E. The Effects of Spatial Patterns in Habitat Quality on Community Dynamics within a Site. Proc. R. Soc. Lond. B 1997, 264, 347–354. [Google Scholar] [CrossRef]
- Tomppo, E.; Gschwantner, T.; Lawrence, M.; McRoberts, R.E. National Forest Inventories. Pathways for Common Reporting; Springer Science + Business Media B.V.: New York, NY, USA, 2010. [Google Scholar]
- Niemelä, J. Invertebrates and Boreal Forest Management. Conserv. Biol. 1997, 11, 601–610. [Google Scholar] [CrossRef]
- Barsoum, N.; Fuller, L.; Ashwood, F.; Reed, K.; Bonnet-Lebrun, A.-S.; Leung, F. Ground-dwelling spider (Araneae) and carabid beetle (Coleoptera: Carabidae) community assemblages in mixed and monoculture stands of oak (Quercus robur L./Quercus petraea (Matt.) Liebl.) and Scots pine (Pinus sylvestris L.). For. Ecol. Manag. 2014, 321, 29–41. [Google Scholar] [CrossRef]
- Wu, H.; Sharpe, P.J.H.; Walker, J.; Penridge, L.K. Ecological Field Theory: A Spatial Analysis of Resource Interference among Plants. Ecol. Model. 1985, 29, 215–243. [Google Scholar] [CrossRef]
- Pretzsch, H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For. Ecol. Manag. 2014, 327, 251–264. [Google Scholar] [CrossRef]
- Laca, E.A. Multi-Scape Interventions to Match Spatial Scales of Demand and Supply of Ecosystem Services. Front. Sustain. Food Syst. 2021, 4, 607276. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Pukkala, T. Effect of Scots pine seed trees on the density of ground vegetation and tree seedlings. Silva Fenn. 1989, 23, 159–167. [Google Scholar] [CrossRef][Green Version]
- Schua, K.; Fischer, H.; Lehmann, B.; Wagner, S. Single tree effects of sessile oak (Quercus petraea (Matt.) Liebl.) within pure stands (Pinus sylvestris L.) on topsoil properties. Allg. For. J. Ztg. 2007, 178, 172–179, (In German with English abstract). [Google Scholar]
- Gruba, P.; Mulder, J.; Pacanowskia, P. Combined effects of soil disturbances and tree positions on spatial variability of soil pHCaCl2 under oak and pine stands. Geoderma 2020, 376, 114537. [Google Scholar] [CrossRef]
- Wagner, S.; Herrmann, I.; Huth, F. Tools familiar, impact unexpected: Silviculture and ecosystem services on a small forest scale. Allg. For. J. Ztg. 2020, 190, 89–100. [Google Scholar] [CrossRef]
- Jonard, M.; Andre, F.; Ponette, Q. Tree species mediated effects on leaf litter dynamics in pure and mixed stands of oak and beech. Can. J. For. Res. 2008, 38, 528–538. [Google Scholar] [CrossRef]
- Wehnert, A.; Wagner, S. Niche partitioning in carabids: Single-tree admixtures matter. Insect Conserv. Divers. 2019, 12, 131–146. [Google Scholar] [CrossRef]
- Beniamino, F.; Ponge, J.F.; Arpin, P. Soil acidification under the crown of oak trees. I. Spatial distribution. For. Ecol. Manag. 1991, 40, 221–232. [Google Scholar] [CrossRef]
- Abella., S.R.; Springer, J.D. Canopy-tree influences along a soil parent material gradient in Pinus ponderosa-Quercus gambelii forests, northern Arizona. J. Torrey Bot. Soc. 2008, 135, 26–36. [Google Scholar] [CrossRef]
- Hassan, S.M.T.; Ghimire, C.P.; Lubczynski, M.W. Remote sensing upscaling of interception loss from isolated oaks: Sardon catchment case study, Spain. J. Hydrol. 2017, 555, 489–505. [Google Scholar] [CrossRef]
- Kádár, K.; Fazekas, J.P.; Sárospataki, M.; Lövei, G.L. Seasonal dynamics, age structure and reproduction of four Carabus species (Coleoptera: Carabidae) living in forested landscapes in Hungary. Acta Zool. Hung. 2015, 61, 57–72. [Google Scholar] [CrossRef]
- Paoletti, M.G.; Martin Cantarino, C. Sex ratio alterations in terrestrial woodlice populations (Isopoda: Oniscidea) from agroecosystems subjected to different agricultural practices in Italy. Appl. Soil Ecol. 2002, 19, 113–120. [Google Scholar] [CrossRef]
- Knapp, M.; Knappová, J. Measurement of body condition in a common carabid beetle, Poecilus cupreus: A comparison of fresh weight, dry weight, and fat content. J. Insect Sci. 2013, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Jervis, M.; Kidd, N. Insect Natural Enemies. Practical Approaches to Their Study and Evaluation; Chapman & Hall: London, UK, 1996. [Google Scholar]
- Leather, S. Insect Sampling in Forest Ecosystems; Blackwell Science Ltd: Malden, MA, USA, 2005. [Google Scholar]
- Huk, T.; Kühne, B. Substrate selection by Carabus clatratus (Coleoptera, Carabidae) and its consequences for offspring development. Oecologia 1999, 121, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Pokluda, P.; Hauck, D.; Cizek, L. Importance of marginal habitats for grassland diversity: Fallows and overgrown tall-grass steppe as key habitats of endangered ground-beetle Carabus hungaricus. Insect Conserv. Divers. 2012, 5, 27–36. [Google Scholar] [CrossRef]
- LFB and LFE. 2017. Available online: http://www.forstliche-umweltkontrolle-bb.de/r3_meteo.php (accessed on 6 November 2017).
- Müller, J.K. Die Bedeutung der Fallenfang-Methode für die Lösung ökologischer Fragestellungen. Zool. Jb. Syst. 1984, 111, 281–305. [Google Scholar]
- Brown, G.R.; Matthews, I.M. A review of extensive variation in the design of pitfall traps and a proposal for a standard pitfall trap design for monitoring ground-active arthropod biodiversity. Ecol. Evol. 2016, 6, 3953–3964. [Google Scholar] [CrossRef] [PubMed]
- Ziesche, M. Effect of Forest Structure and Small-Scale Environmental Conditions on the Community of Epigeic Arthropods (Carabidae, Araneae). Ph.D. Thesis, Technische Universität Dresden, Fakultät Umweltwissenschaften, Tharandt, Germany, 2016. [Google Scholar]
- Heydemann, B. Agrarökologische Problematik (dargetan an Untersuchungen über die Tierwelt der Bodenoberfläche der Kulturfelder). Ph.D. Thesis, Hohen Philosophischen Fakultät an der Christian-Albrechts-Universität, Kiel, Germany, 1953. [Google Scholar]
- Scheller, H.V. Pitfall trapping as the basis for studying ground beetle (Carabidae) predation in spring barley. Tidsskr. Planteavl. 1984, 88, 317–324. [Google Scholar]
- Mühlenberg, M. Freilandökologie, 3rd ed.; Quelle & Meyer Verlag: Heidelberg/Wiesbaden, Germany, 1993. [Google Scholar]
- Langraf, V.; David, S.; Babosová, R.; Petrovičová, K.; Schlarmannová, J. Change of Ellipsoid Biovolume (EV) of Ground Beetles (Coleoptera, Carabidae) along an Urban–Suburban–Rural Gradient of Central Slovakia. Diversity 2020, 12, 475. [Google Scholar] [CrossRef]
- Szyszko, J.; Gryuntal, S.; Schwerk, A. Differences in Locomotory Activity Between Male and Female Carabus hortensis (Coleoptera: Carabidae) in a Pine Forest and a Beech Forest in Relation to Feeding State. Environ. Entomol. 2004, 33, 1442–1446. [Google Scholar] [CrossRef]
- Clark, P.J.; Evans, F.C. Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology 1954, 35, 445–453. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology, 2nd ed.; Addison Wesley Longman Inc., University of British Columbia: Vancouver, BC, Canada, 1999. [Google Scholar]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns. Methodology and Applications with R; Chapman & Hall/CRC Interdisciplinary Statistics Series; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Schober, P.; Boer, C.; Schwarte, L.A. Correlation Coefficients: Appropriate Use and Interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef]
- Zinke, P.J. The Pattern of Influence of Individual Forest Trees on Soil Properties. Ecology 1962, 43, 130–133. [Google Scholar] [CrossRef]
- Shamsutdinov, Z.; Ubaydullaev, S.; Shamsutdinov, N.; Mirzaev, B.; Mamatov, F.; Chorshabiyev, N. The concept of the phytogenic field: Theory, research experience and practical significance. IOP Conf. Ser. Earth Env. For. Sci. 2020, 614, 012164. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Vinatier, F.; Tixier, P.; Duyck, P.-F.; Lescourret, F. Factors and mechanisms explaining spatial heterogeneity: A review of methods for insect populations. Methods Ecol. Evol. 2011, 2, 11–22. [Google Scholar] [CrossRef]
- Magnusson, A.; Skaug, H.; Nielsen, A.; Berg, C.; Kristensen, K.; Maechler, M.; van Bentham, K.; Bolker, B.; Sadat, N.; Lüdecke, D.; et al. glm TMB: Generalized Linear Mixed Models Using Template Model Builder. Date/Publication 2020-07-02 11:30:17 UTC. 2020. Available online: https://cran.r-project.org/web/packages/glmmTMB/glmmTMB.pdf (accessed on 19 January 2021).
- Barbaro, L.; Pontcharraud, L.; Vetillard, F.; Guyon, D.; Jactel, H. Comparative responses of bird, carabid, and spider assemblages to stand and landscape diversity in maritime pine plantation forest. Écoscience 2005, 12, 110–121. [Google Scholar] [CrossRef]
- Samways, M.J.; McGeoch, M.A.; New, T.R. Insect Conservation: A Handbook of Approaches and Methods; Techniques in Ecology & Conservation Series; Oxford University Press: London, UK, 2013. [Google Scholar]
- Davis, D.E.; Gagné, S.A. Boundaries in ground beetle (Coleoptera: Carabidae) and environmental variables at the edges of forest patches with residential developments. PeerJ 2018, 6, e4226. [Google Scholar] [CrossRef] [PubMed]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Tree 1995, 10, 58–62. [Google Scholar] [CrossRef]
- Didham, R.K. An overview of invertebrate responses to forest fragmentation. In Forests and Insects; Watt, A.D., Stork, N.E., Hunter, M.D., Eds.; Chapman & Hall: London, UK, 1997; pp. 305–320. [Google Scholar]
- Hamberg, L.; Lehvävirta, S.; Kotze, D.J. Forest edge structure as a shaping factor of understorey vegetation in urban forests in Finland. For. Ecol. Manag. 2009, 257, 712–722. [Google Scholar] [CrossRef]
- Ries, L.; Sisk, T.D. What is an edge species? The implications of sensitivity to habitat edges. Oikos 2010, 119, 1636–1642. [Google Scholar] [CrossRef]
- Fahrig, L. Rethinking patch size and isolation effects: The habitat amount hypothesis. J. Biogeogr. 2013, 40, 1649–1663. [Google Scholar] [CrossRef]
- Niemelä, J.; Haila, Y.; Halme, E.; Pajunen, T.; Punttila, P. Small-scale heterogeneity in the spatial distribution of carabid beetles in the southern Finnish taiga. J. Biogeogr. 1992, 19, 173–181. [Google Scholar] [CrossRef]
- Day, K.R.; Marshall, S.; Heaney, C. Association between Forest Type and Invertebrates: Ground Beetle Community Patterns in a Natural Oakwood and Juxtaposed Conifer Plantation. Forestry 1993, 66, 37–50. [Google Scholar] [CrossRef]
- Hunter, M.L. Maintaining Biodiversity in Forest Ecosystems; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Kraus, D.; Krumm, F. (Eds.) Integrative Approaches as an Opportunity for the Conservation of Forest Biodiversity; European Forest Institute: Freiburg, Germany, 2013. [Google Scholar]
- Niemelä, J.; Koivula, M.; Kotze, D.J. The effects of forestry on carabid beetles (Coleoptera: Carabidae) in boreal forest. J. Insect Conserv. 2007, 11, 5–18. [Google Scholar] [CrossRef]
- Szujecki, A. Ecology of Forest Insects; Series Entomologica; Dr. W. Junk Publishers: Dordrecht, The Netherlands, 1987; Volume 26. [Google Scholar]
- Jukes, M.R.; Peace, A.J.; Ferris, R. Carabid beetle communities associated with coniferous plantations in Britain: The influence of site, ground vegetation and stand structure. For. Ecol. Manag. 2001, 148, 271–286. [Google Scholar] [CrossRef]
- Pretzsch, H.; Forrester, D.I.; Bauhus, J. Mixed-Species Forests: Ecology and Management; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar] [CrossRef]
- Schowalter, T.D. Insect Ecology. An Ecosystem Approach, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Moya-Laraño, J.; Macías-Ordóñez, R.; Blanckenhorn, W.U.; Fernández-Montraveta, C. Analysing Body Condition: Mass, Volume or Density? J. Anim. Ecol. 2008, 77, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Langraf, V.; Petrovičová, K.; David, S.; Ábelová, M.; Schlarmannová, J. Body volume in ground beetles (Carabidae) reflects biotope disturbance. Folia Oecologica 2017, 44, 114–120. [Google Scholar] [CrossRef]
- Loreau, M. Annual activity and life cycles of carabid beetles in two forest communities. Holarct. Ecol. 1985, 8, 228–235. [Google Scholar] [CrossRef]
- Lövei, G.L.; Sunderland, K.D. Ecology and behavior of Ground Beetles (Coleoptera: Carabidae). Annu. Rev. Entomol. 1996, 41, 231–256. [Google Scholar] [CrossRef]
- Müller-Motzfeld, G. Laufkäfer in Wäldern Deutschlands. Angew. Carab. Suppl. 2001, II, 9–20. [Google Scholar]
- Wehnert, A.; Wagner, S.; Huth, F. Spatio-Temporal Distribution of Carabids Influenced by Small-Scale Admixture of Oak Trees in Pine Stands. Diversity 2020, 12, 398. [Google Scholar] [CrossRef]
- Shibuya, S.; Kubota, K.; Ohsawa, M.; Kikvidze, Z. Assembly rules for ground beetle communities: What determines community structure, environmental factors or competition? Eur. J. Entomol. 2011, 108, 453–459. [Google Scholar] [CrossRef]
- Magura, T.; Tóthmérész, B.; Elek, Z. Effects of leaf-litter addition on carabid beetles in a non-native Norway spruce plantation. Acta Zool. Acad. Sci. Hung. 2004, 50, 9–23. [Google Scholar]
- Tyler, G. Variability in colour, metallic lustre, and body size of Carabus arvensis Herbst, 1784 (Coleoptera: Carabidae) in relation to habitat properties. Entomol. Fenn. 2010, 21, 90–96. [Google Scholar] [CrossRef]
- Jelaska, L.Š.; Dumbović, V.; Kučinić, M. Carabid beetle diversity and mean individual biomass in beech forests of various age. ZooKeys 2011, 100, 393–405. [Google Scholar] [CrossRef]
- Ohsawa, M. The role of isolated old oak trees in maintaining beetle diversity within larch plantations in the central mountainous region of Japan. For. Ecol. Manag. 2007, 250, 215–226. [Google Scholar] [CrossRef]
- Wiezik, M.; Svitok, M.; Dovčiak, M. Conifer introductions decrease richness and alter composition of litter-dwelling beetles (Coleoptera) in Carpathian oak forests. For. Ecol. Manag. 2007, 247, 61–71. [Google Scholar] [CrossRef]
- Worthen, W.B.; Merriman, D.C.G. Relationships between Carabid Beetle Commmunities and Forest Stand Parameters: Taxon Congruencer or Habitat Association. Southeast. Nat. 2013, 12, 379–386. [Google Scholar] [CrossRef]
- Skłodowski, J.; Szczeszek, J. Dead wood modifies mobility of ground beetles. Baltic J. Coleopterol. 2015, 15, 91–98. [Google Scholar]
- Andrési, D.; Bali, L.; Tuba, K.; Szinetár, C. Comparative study of ground beetle and ground-dwelling spider assemblages of artificial gap openings. Community Ecol. 2018, 19, 133–140. [Google Scholar] [CrossRef]
- Matveinen-Huju, K.; Niemelä, J.; Rita, H.; O´Hara, R.B. Retention-tree groups in clear-cuts: Do they constitute "life-boats" for spiders and carabids? For. Ecol. Manag. 2006, 230, 119–135. [Google Scholar] [CrossRef]
- Koch Widerberg, M. Oak as Retention Tree in Commercial Spruce Forests. Effects on Species Diversity of Saproxylic Beetles and Wood Production. Ph.D. Thesis, Swedish University of Agricultural Sciences, Alnarp, Sweden, 2013. No. 2013:66. [Google Scholar]
- Koivula, M.; Niemelä, J. Gap felling as a forest harvesting method in boreal forests: Responses of carabid beetles (Coleoptera, Carabidae). Ecography 2003, 26, 179–187. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Hanula, J.L.; Horn, S.; Kilgo, J.C.; Moorman, C.E. The response of ground beetles (Coleoptera: Carabidae) to selection cutting in a South Carolina bottomland hardwood forest. Biodivers. Conserv. 2006, 15, 261–274. [Google Scholar] [CrossRef]
- Vítková, L.; Bače, R.; Kjučukov, P.; Svoboda, M. Deadwood management in Central European forests: Key considerations for practical implementation. For. Ecol. Manag. 2018, 429, 394–405. [Google Scholar] [CrossRef]
- Weber, P.; Bardgett, R.D. Influence of single trees on spatial and temporal patterns of belowground properties in native pine forest. Soil Biol. Biochem. 2011, 43, 1372e1378. [Google Scholar] [CrossRef]
- Vrška, T.; Janík, D.; Pálková, M.; Adam, D.; Trochta, J. Below- and above-ground biomass, structure and patterns in ancient lowland coppices. iForest 2016, 10, 23–31. [Google Scholar] [CrossRef]
- García, L.V.; Maltez-Mouro, S.; Pérez-Ramos, I.M.; Freitas, H.; Marañón, T. Counteracting gradients of light and soil nutrients in the understorey of Mediterranean oak forests. Web Ecol. 2006, 6, 67–74. [Google Scholar] [CrossRef]
- Skłodowski, J. Consequence of the transformation of a primeval forest into a managed forest for carabid beetles (Coleoptera: Carabidae)—A case study from Białowieża (Poland). Eur. J. Entomol. 2014, 111, 639–648. [Google Scholar] [CrossRef]
- Heliölä, J.; Koivula, M.; Niemelä, J. Distribution of Carabid Beetles (Coleoptera, Carabidae) across a Boreal Forest-Clearcut Ecotone. Conserv. Biol. 2001, 15, 370–377. [Google Scholar] [CrossRef]
- Wagner, S.; Fischer, H.; Huth, F. Canopy effects on vegetation caused by harvesting and regeneration treatments. Eur. J. For. Res. 2011, 130, 17–40. [Google Scholar] [CrossRef]
- Lintunen, A.; Kaitaniemi, P.; Perttunen, J.; Sievänen, R. Analysing species-specific light transmission and related crown characteristics of Pinus sylvestris and Betula pendula using a shoot-level 3D model. Can. J. For. Res. 2013, 43. [Google Scholar] [CrossRef]
- Perot, T.; Mårell, A.; Korboulewsky, N.; Seigner, V.; Balandier, P. Modeling and predicting solar radiation transmittance in mixed forests at a within-stand scale from tree species basal area. For. Ecol. Manag. 2017, 390, 127–136. [Google Scholar] [CrossRef]
- Grüm, L. Spatial Distribution of Males and Females of Carabus arcensis Hbst. in the Breeding Season. In The Role of Ground Beetles in Ecological and Environmental Studies; Stork, N.E., Ed.; Intercept Ltd.: Andover, MA, USA, 1990; pp. 277–287. [Google Scholar]
- Loreau, M. Vertical distribution of activity of carabid beetles in a beech forest floor. Pedobiologia 1987, 30, 173–178. [Google Scholar]
- Hawes, C.; Stewart, A.J.A.; Evans, H.F. The impact of wood ants (Formica rufa) on the distribution and abundance of ground beetles (Coleoptera: Carabidae) in a Scots pine plantation. Oecologia 2002, 131, 612–619. [Google Scholar] [CrossRef]
- Luff, M.L. Biology and ecology of immature stages of ground beetles (Carabidae). In Proceedings of the 11th European Carabidologists’ Meeting, Århus, Denmark, 21–24 July 2003; Lövei, G.L., Toft, S., Eds.; DIAS Report, No. 114. Ministry of Food, Agriculture and Fisheries: Tjele, Denmark, 2005; pp. 183–208. [Google Scholar]
- Koivula, M.; Niemelä, J. Boreal Carabid Beetles (Coleoptera, Carabidae) in Managed Spruce Forests—A Summary of Finnish Case Studies. Silva Fenn. 2002, 36, 423–436. [Google Scholar] [CrossRef][Green Version]
- Krissl, K.; Müller, F. Zweckmäßige Dauermischungsformen und Mischungsregulierung. Österr. Forstztg. 1990, 3, 29–32. [Google Scholar]
- Perot, T.; Picard, N. Mixture enhances productivity in a two-species forest: Evidence from a modeling approach. Ecol. Res. Ecol. Soc. JPN. 2012, 27, 83–94. [Google Scholar] [CrossRef]
- Šiška, B.; Eliašová, M.; Kollár, J. Carabus Population Response to Drought in Lowland Oak Hornbeam Forest. Water 2020, 12, 3284. [Google Scholar] [CrossRef]
- Arndt, E.; Arndt, M. Auswertung der Bodenfallenfänge von Carabidenlarven (Coleoptera) im Hakel (Nordharzvorland). Hercynia 1987, 24, 22–33. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Area | No. 1.11 | No. 1.12 | No. 1.13 | No. 1.14 | No. 2.15 | No. 2.16 | No. 2.17 | No. 2.18 | |||||||||||
| Parameters | Units | Pine | Oak | Pine | Oak | Pine | Oak | Pine | Oak | Pine | Oak | Pine | Oak | Pine | Oak | Pine | Oak | ||
| age | (years) | fst 1 | 122 | 57 | 111 | 37 | 144 | 49 | 197 | fst 2 | 62 | 94 | 99 | 59 | 139 | 104 | 81 | 92 | |
| no. of trees | (n/ha) | 372 | 28 | 371 | 11 | 167 | 11 | 122 | 600 | 106 | 483 | 67 | 206 | 217 | 194 | 256 | |||
| prop. of trees | (%) | 100 | 7 | 93 | 6 | 94 | 8 | 92 | 85 | 15 | 88 | 12 | 49 | 51 | 43 | 57 | |||
| crown cover | (%) | 62.4 | 1.0 | 53.9 | 0.2 | 61.7 | 0.2 | 57.4 | 20.0 | 30.5 | 22.1 | 10.9 | 18.6 | 41.0 | 14.3 | 36.8 | |||
| dbh (mean) | (cm) | 30.92 | 16.44 | 28.25 | 17.10 | 47.26 | 24.08 | 49.93 | 23.48 | 31.50 | 25.68 | 29.78 | 38.86 | 19.34 | 32.55 | 23.21 | |||
| height (mean) | (m) | 17.67 | 14.21 | 16.55 | 10.80 | 21.76 | 17.70 | 20.29 | 19.37 | 19.35 | 19.78 | 17.99 | 24.40 | 14.53 | 22.67 | 19.16 | |||
| crown diameter (mean) | (m) | 6.05 | 3.62 | 5.12 | 3.59 | 8.49 | 3.27 | 8.41 | 3.69 | 6.53 | 4.22 | 6.36 | 5.04 | 5.89 | 4.66 | 4.75 | |||
| aggregation index (R) | 1.095 | 0.877 | 1.010 | 0.450 | 1.146 | 0.856 | 1.295 * | 1.190 * | 0.925 | 1.281 * | 1.147 | 1.238 * | 1.079 | 1.359 * | 1.008 | ||||
| Forest Area | No. 3.4 | No. 3.5 | No. 3.6 | No. 4.7 | No. 4.8 | No. 4.9 | No. 4.10 | ||||||||||||
| Parameters | Units | Pine | Oak | Pine | Oak | Pine | Oak | Pine | Pine | Pine | Pine | ||||||||
| age | (years) | fst 3 | 53 | 150 | 57 | 150 | 60 | 150 | fst 4 | 57 | 60 | 87 | 104 | ||||||
| no. of trees | (n/ha) | 645 | 5 | 547 | 13 | 526 | 5 | 817 | 544 | 433 | 456 | ||||||||
| prop. of trees | (%) | 99 | 1 | 98 | 2 | 99 | 1 | 100 | 100 | 100 | 100 | ||||||||
| crown cover | (%) | 24.0 | 1.8 | 19.3 | 5.8 | 24.1 | 3.6 | 61.5 | 60.9 | 26.4 | 34.5 | ||||||||
| dbh (mean) | (cm) | 24.05 | 38.02 | 24.36 | 44.03 | 25.69 | 48.32 | 24.09 | 24.71 | 30.41 | 29.74 | ||||||||
| height (mean) | (m) | 20.45 | 16.07 | 20.12 | 18.66 | 21.18 | 17.87 | 21.58 | 19.63 | 20.93 | 22.71 | ||||||||
| crown diameter (mean) | (m) | 2.97 | 8.13 | 2.84 | 10.33 | 3.34 | 12.84 | 3.10 | 3.15 | 2.35 | 2.65 | ||||||||
| aggregation index (R) | 1.116 * | 0.635 | 1.196 * | 1.029 | 1.249 * | 0.220 | 1.158 * | 1.360 * | 1.225 * | 1.316 * | |||||||||
| No. of Traps per Zone | Proportion of Zones (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Forest Stand Type | Stand No. | Z1 (oak) | Z2 (oak-pine) | Z3 (pine) | Z1 (oak) | Z2 (oak-pine) | Z3 (pine) | |
| fst1 | pure oak | 1.11 | 8 | 0 | 0 | 100 | 0 | 0 |
| 1.12 | 8 | 0 | 0 | 100 | 0 | 0 | ||
| 1.13 | 8 | 0 | 0 | 100 | 0 | 0 | ||
| 1.14 | 8 | 0 | 0 | 100 | 0 | 0 | ||
| fst2 | oak–pine mix. | 2.15 | 3 | 5 | 0 | 34 | 66 | 0 |
| 2.16 | 3 | 5 | 0 | 26 | 68 | 6 | ||
| 2.17 | 3 | 5 | 0 | 30 | 69 | 2 | ||
| 2.18 | 2 | 6 | 0 | 26 | 74 | 0 | ||
| fst 3 | pine with <10% oak | 3.4 | 2 | 6 | 23 | 2 | 14 | 84 |
| 3.5 | 1 | 19 | 14 | 6 | 48 | 45 | ||
| 3.6 | 1 | 8 | 20 | 2 | 15 | 83 | ||
| fst 4 | pure pine | 4.7 | 0 | 0 | 8 | 0 | 0 | 100 |
| 4.8 | 0 | 0 | 8 | 0 | 0 | 100 | ||
| 4.9 | 0 | 0 | 8 | 0 | 0 | 100 | ||
| 4.10 | 0 | 0 | 8 | 0 | 0 | 100 | ||
| Forest Stand Parameters | Carabid Parameters | Carabid Species | Oaks | Pines | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Females | p | Males | p | Imagines | p | Females | p | Males | p | Imagines | p | |||
| crown cover (%) | beetle density | C. arvensis | 0.018 | n.s. | −0.277 | n.s. | −0.122 | n.s. | −0.179 | n.s. | 0.098 | n.s. | −0.046 | n.s. |
| C. coriaceus | 0.294 | n.s. | 0.407 | n.s. | 0.236 | n.s. | −0.412 | . | −0.466 | . | −0.420 | . | ||
| C. hortensis | 0.508 | * | 0.259 | n.s. | 0.462 | . | −0.516 | * | −0.394 | n.s. | −0.548 | * | ||
| C. violaceus | 0.676 | ** | −0.054 | n.s. | 0.485 | . | −0.526 | * | 0.191 | n.s. | −0.441 | . | ||
| Cal. inquisitor | 0.814 | *** | 0.806 | *** | 0.811 | *** | −0.668 | ** | −0.670 | ** | −0.670 | ** | ||
| body size | C. arvensis | 0.165 | n.s. | −0.649 | ** | −0.335 | n.s. | −0.340 | n.s. | 0.492 | . | 0.100 | n.s. | |
| C. coriaceus | 0.080 | n.s. | 0.330 | n.s. | 0.204 | n.s. | −0.360 | n.s. | −0.487 | . | −0.466 | . | ||
| C. hortensis | 0.446 | . | 0.560 | * | 0.566 | * | −0.552 | * | −0.565 | * | −0.636 | ** | ||
| C. violaceus | 0.286 | n.s. | −0.479 | . | 0.034 | n.s. | −0.132 | n.s. | 0.501 | * | 0.207 | n.s. | ||
| Cal. inquisitor | 0.872 | *** | 0.830 | *** | 0.880 | *** | NA | - | NA | - | NA | - | ||
| aggregation index trees (R) | beetle density | C. arvensis | 0.231 | n.s. | 0.132 | n.s. | 0.193 | n.s. | −0.291 | n.s. | 0.135 | n.s. | −0.060 | n.s. |
| C. coriaceus | 0.435 | . | 0.540 | * | 0.454 | . | −0.436 | . | −0.287 | n.s. | −0.268 | n.s. | ||
| C. hortensis | 0.610 | . | 0.468 | . | 0.671 | ** | −0.510 | . | −0.380 | n.s. | −0.506 | * | ||
| C. violaceus | 0.423 | n.s. | −0.145 | n.s. | 0.274 | n.s. | 0.507 | * | 0.492 | * | 0.497 | * | ||
| Cal. inquisitor | 0.436 | n.s. | 0.396 | n.s. | 0.406 | n.s. | −0.782 | ** | −0.737 | * | −0.752 | ** | ||
| body size | C. arvensis | 0.189 | n.s. | −0.159 | n.s. | 0.007 | n.s. | −0.217 | n.s. | 0.811 | *** | 0.452 | . | |
| C. coriaceus | 0.293 | n.s. | −0.105 | n.s. | 0.118 | n.s. | −0.415 | n.s. | 0.784 | * | 0.297 | n.s. | ||
| C. hortensis | −0.003 | n.s. | 0.424 | n.s. | −0.007 | n.s. | 0.264 | n.s. | 0.130 | n.s. | 0.130 | n.s. | ||
| C. violaceus | 0.258 | n.s. | −0.119 | n.s. | −0.039 | n.s. | −0.114 | n.s. | 0.387 | n.s. | 0.155 | n.s. | ||
| Cal. inquisitor | −0.961 | * | 0.749 | n.s. | −0.059 | n.s. | NA | - | NA | - | NA | - | ||
| Carabid Species | Oak Zone (Z1) | Oak–Pine Zone (Z2) | Pine Zone (Z3) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fem | p | Mal | p | Imag | p | Fem | p | Mal | p | Imag | p | Fem | p | Mal | p | Imag | p | ||
| beetle density | C. arv. | 0.051 | n.s. | −0.250 | n.s. | −0.099 | n.s. | 0.009 | n.s. | 0.215 | n.s. | 0.124 | n.s. | −0.052 | n.s. | 0.084 | n.s. | 0.007 | n.s. |
| C. cor. | 0.180 | n.s. | 0.214 | n.s. | 0.074 | n.s. | 0.341 | n.s. | 0.443 | . | 0.501 | . | −0.390 | n.s. | −0.490 | . | −0.401 | n.s. | |
| C. hort. | 0.375 | n.s. | 0.220 | n.s. | 0.358 | n.s. | 0.269 | n.s. | 0.105 | n.s. | 0.273 | n.s. | −0.520 | * | −0.269 | n.s. | −0.507 | * | |
| C. viol. | 0.654 | * | 0.004 | n.s. | 0.497 | * | −0.277 | n.s. | −0.305 | n.s. | −0.339 | n.s. | −0.408 | n.s. | 0.201 | n.s. | −0.225 | n.s. | |
| Cal. in. | 0.925 | *** | 0.915 | *** | 0.920 | *** | NA | - | NA | - | NA | - | NA | - | NA | - | NA | - | |
| body size | C. arv. | 0.254 | n.s. | −0.639 | ** | −0.265 | n.s. | −0.100 | n.s. | 0.007 | n.s. | −0.065 | n.s. | −0.164 | n.s. | 0.575 | * | 0.284 | n.s. |
| C. cor. | 0.541 | . | 0.366 | n.s. | 0.475 | n.s. | −0.077 | n.s. | 0.593 | . | 0.404 | n.s. | −0.240 | n.s. | 0.593 | . | 0.023 | n.s. | |
| C. hort. | 0.502 | . | 0.631 | * | 0.537 | * | −0.004 | n.s. | 0.041 | n.s. | −0.096 | n.s. | −0.002 | n.s. | −0.630 | . | −0.035 | n.s. | |
| C. viol. | 0.196 | n.s. | −0.352 | n.s. | −0.059 | n.s. | 0.269 | n.s. | −0.142 | n.s. | −0.012 | n.s. | −0.088 | n.s. | 0.414 | n.s. | 0.062 | n.s. | |
| Cal. in. | NA | - | 0.688 | n.s. | 0.819 | . | NA | - | NA | - | NA | - | NA | - | NA | - | NA | - | |
| No. of Beetles | Females | Males | Imagines | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | std. Error | p-Value | Estimate | std. Error | p-Value | Estimate | std. Error | p-Value | |||||
| C. arvensis | interc. | −0.3486 | 0.6861 | 0.611 | −0.9919 | 0.6883 | 0.150 | −0.0281 | 0.5520 | 0.959 | |||
| fst 2 | −0.0135 | 0.6499 | 0.983 | 0.8178 | 0.5915 | 0.167 | 0.4309 | 0.5255 | 0.421 | ||||
| fst 3 | −0.0336 | 0.7484 | 0.964 | 0.8762 | 0.6981 | 0.209 | 0.4549 | 0.6162 | 0.460 | ||||
| fst 4 | −0.5006 | 0.8035 | 0.533 | 0.3924 | 0.7666 | 0.609 | 0.0271 | 0.6484 | 0.967 | ||||
| oak cc | 0.0006 | 0.0079 | 0.944 | 0.0036 | 0.0079 | 0.651 | 0.0031 | 0.0060 | 0.608 | ||||
| C. coriaceus | interc. | −3.2320 | 0.9403 | 0.001 | *** | −4.7730 | 1.0460 | 0.000 | *** | −3.0466 | 0.7934 | 0.000 | *** |
| fst 2 | 1.2420 | 0.7370 | 0.092 | . | 1.7040 | 0.8671 | 0.049 | * | 1.4339 | 0.6162 | 0.020 | * | |
| fst 3 | 1.5810 | 0.8680 | 0.068 | . | 3.0240 | 0.9322 | 0.002 | ** | 2.1183 | 0.7173 | 0.003 | ** | |
| fst 4 | −18.7500 | 10,760.00 | 0.999 | −17.6800 | 1383.00 | 0.999 | −18.4916 | 9404.59 | 0.999 | ||||
| oak cc | 0.0214 | 0.0114 | 0.060 | . | 0.0344 | 0.0114 | 0.003 | ** | 0.0243 | 0.0092 | 0.008 | ** | |
| C. hortensis | interc. | −5.4180 | 1.3420 | 0.000 | *** | −3.8935 | 1.5625 | 0.013 | * | −3.9899 | 1.1684 | 0.001 | *** |
| fst 2 | 1.9110 | 0.9020 | 0.034 | * | 0.5937 | 1.0665 | 0.578 | 1.3733 | 0.7862 | 0.081 | . | ||
| fst 3 | 3.0090 | 1.1146 | 0.009 | ** | 1.8370 | 1.3474 | 0.173 | 2.4921 | 1.2026 | 0.015 | * | ||
| fst 4 | −16.6500 | 10,720.00 | 0.999 | −17.6840 | 9896.70 | 0.999 | −17.5303 | 9620.49 | 0.999 | ||||
| oak cc | 0.0548 | 0.0151 | 0.000 | *** | 0.0359 | 0.0185 | 0.053 | . | 0.0461 | 0.0134 | 0.001 | *** | |
| C. violaceus | interc. | −0.6633 | 0.6856 | 0.333 | 0.2344 | 0.7949 | 0.768 | 0.3961 | 0.5529 | 0.474 | |||
| fst 2 | −0.7316 | 0.5462 | 0.180 | −0.9195 | 0.6441 | 0.153 | −0.9192 | 0.4485 | 0.040 | * | |||
| fst 3 | −0.3481 | 0.6424 | 0.588 | −1.1208 | 0.7760 | 0.149 | −0.6468 | 0.5300 | 0.220 | ||||
| fst 4 | −0.9944 | 0.8048 | 0.217 | −1.1232 | 0.8556 | 0.189 | −0.8987 | 0.6131 | 0.143 | ||||
| oak cc | 0.0039 | 0.0092 | 0.668 | −0.0515 | 0.0109 | 0.169 | −0.0040 | 0.0075 | 0.592 | ||||
| No. of Beetles | Females | Males | Imagines | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | Std. Error | p-Value | Estimate | Std. Error | p-Value | Estimate | Std. Error | p-Value | |||||
| C. arvensis | interc. | −0.3108 | 0.3914 | 0.427 | −0.7406 | 0.3767 | 0.049 | * | 0.1922 | 0.3400 | 0.572 | ||
| fst 2 | −0.3304 | 0.6209 | 0.595 | 0.4308 | 0.5528 | 0.436 | 0.0119 | 0.5114 | 0.981 | ||||
| fst 3 | −0.4359 | 0.6658 | 0.513 | 0.5275 | 0.5902 | 0.371 | −0.0104 | 0.5490 | 0.985 | ||||
| fst 4 | −0.8993 | 0.7052 | 0.202 | 0.1446 | 0.6511 | 0.824 | −0.3969 | 0.5691 | 0.486 | ||||
| Z2 | 0.4232 | 0.3779 | 0.263 | 0.3400 | 0.3576 | 0.342 | 0.4111 | 0.2743 | 0.134 | ||||
| Z3 | 0.3594 | 0.4061 | 0.376 | −0.0076 | 0.3961 | 0.985 | 0.2010 | 0.2986 | 0.501 | ||||
| C. coriaceus | interc. | −1.7804 | 0.5022 | 0.000 | *** | −2.3671 | 0.5774 | 0.000 | *** | −1.3491 | 0.3950 | 0.001 | *** |
| fst 2 | 1.1268 | 0.6541 | 0.085 | . | 1.3251 | 0.7374 | 0.072 | . | 1.3132 | 0.5043 | 0.009 | ** | |
| fst 3 | 1.7444 | 0.7056 | 0.013 | * | 2.0437 | 0.6780 | 0.000 | *** | 2.3143 | 0.5064 | 0.000 | *** | |
| fst 4 | −18.3099 | 9565.85 | 0.998 | −15.0008 | 3718.91 | 0.997 | −18.0114 | 8434.89 | 0.998 | ||||
| Z2 | −1.3491 | 0.4878 | 0.006 | ** | −1.8893 | 0.4555 | 0.000 | *** | −1.6282 | 0.3704 | 0.000 | *** | |
| Z3 | −1.7433 | 0.5535 | 0.002 | ** | −2.5402 | 0.5014 | 0.000 | *** | −2.0328 | 0.4078 | 0.000 | *** | |
| C. hortensis | interc. | −1.2211 | 0.4309 | 0.005 | ** | −1.5070 | 0.0513 | 0.006 | ** | −0.6934 | 0.4057 | 0.087 | . |
| fst 2 | 0.3203 | 0.6390 | 0.616 | −0.0189 | 0.0858 | 0.982 | 0.2843 | 0.5953 | 0.633 | ||||
| fst 3 | 1.2173 | 0.6624 | 0.066 | . | 2.0460 | 0.7610 | 0.007 | ** | 1.6898 | 0.5911 | 0.004 | ** | |
| fst 4 | −16.5116 | 9282.76 | 0.999 | −15.7900 | 10,690.00 | 0.999 | −16.7419 | 9698.62 | 0.999 | ||||
| Z2 | −0.8988 | 0.5207 | 0.084 | . | −1.6910 | 0.5020 | 0.001 | *** | −1.2569 | 0.3776 | 0.001 | *** | |
| Z3 | −3.8908 | 1.1116 | 0.000 | *** | −4.6770 | 1.0710 | 0.000 | *** | −4.2323 | 0.7833 | 0.000 | *** | |
| C. violaceus | interc. | −0.3880 | 0.2243 | 0.083 | . | −0.7902 | 0.2895 | 0.006 | ** | 0.1213 | 0.1835 | 0.509 | |
| fst 2 | −0.8097 | 0.5280 | 0.125 | −0.8244 | 0.7313 | 0.260 | −1.0273 | 0.4775 | 0.032 | * | |||
| fst 3 | −0.4747 | 0.5967 | 0.426 | −1.2281 | 0.8164 | 0.133 | −0.9516 | 0.5377 | 0.077 | . | |||
| fst 4 | −1.1461 | 0.7571 | 0.130 | −1.3330 | 0.8914 | 0.135 | −1.2776 | 0.6122 | 0.037 | * | |||
| Z2 | −0.1240 | 0.5418 | 0.819 | 0.7072 | 0.7213 | 0.327 | 0.3956 | 0.4871 | 0.417 | ||||
| Z3 | −0.1228 | 0.5913 | 0.835 | 1.2444 | 0.7907 | 0.116 | 0.6598 | 0.5298 | 0.213 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wehnert, A.; Wagner, S.; Huth, F. Effects of Pure and Mixed Pine and Oak Forest Stands on Carabid Beetles. Diversity 2021, 13, 127. https://doi.org/10.3390/d13030127
Wehnert A, Wagner S, Huth F. Effects of Pure and Mixed Pine and Oak Forest Stands on Carabid Beetles. Diversity. 2021; 13(3):127. https://doi.org/10.3390/d13030127
Chicago/Turabian StyleWehnert, Alexandra, Sven Wagner, and Franka Huth. 2021. "Effects of Pure and Mixed Pine and Oak Forest Stands on Carabid Beetles" Diversity 13, no. 3: 127. https://doi.org/10.3390/d13030127
APA StyleWehnert, A., Wagner, S., & Huth, F. (2021). Effects of Pure and Mixed Pine and Oak Forest Stands on Carabid Beetles. Diversity, 13(3), 127. https://doi.org/10.3390/d13030127