
Nematode Community-Based Soil Food Web Analysis of Ferralsol, Lithosol and Nitosol Soil Groups in Ghana, Kenya and Malawi Reveals Distinct Soil Health Degradations

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Group and Sampling Site Selection, and Design
2.2. Sampling and Sample Processing
2.3. Soil Analysis
2.4. Nematode Extraction and Enumeration
2.5. Data and Statistical Analyses
3. Results
3.1. Nematode Abundance and Diversity
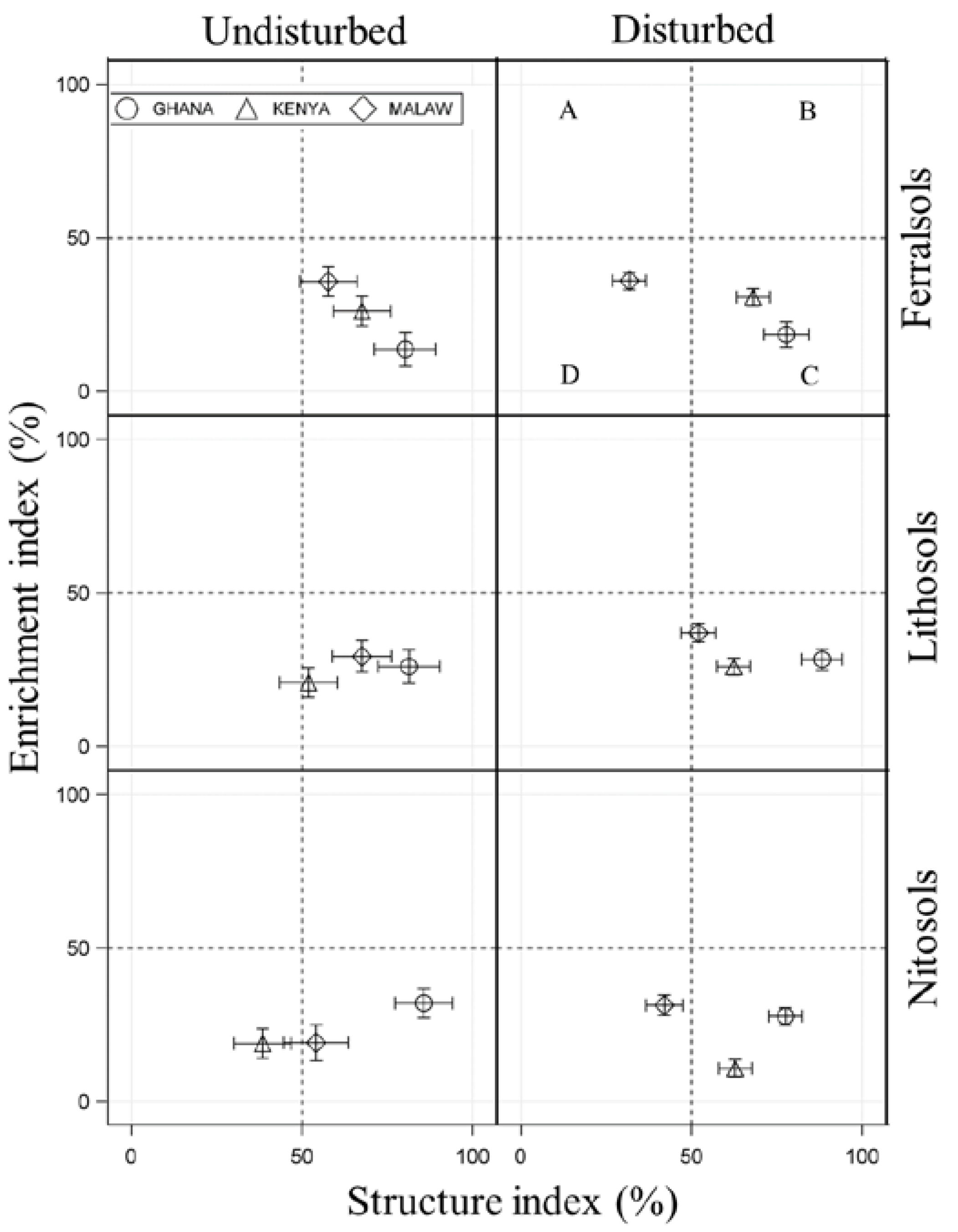
3.2. SFW Structure
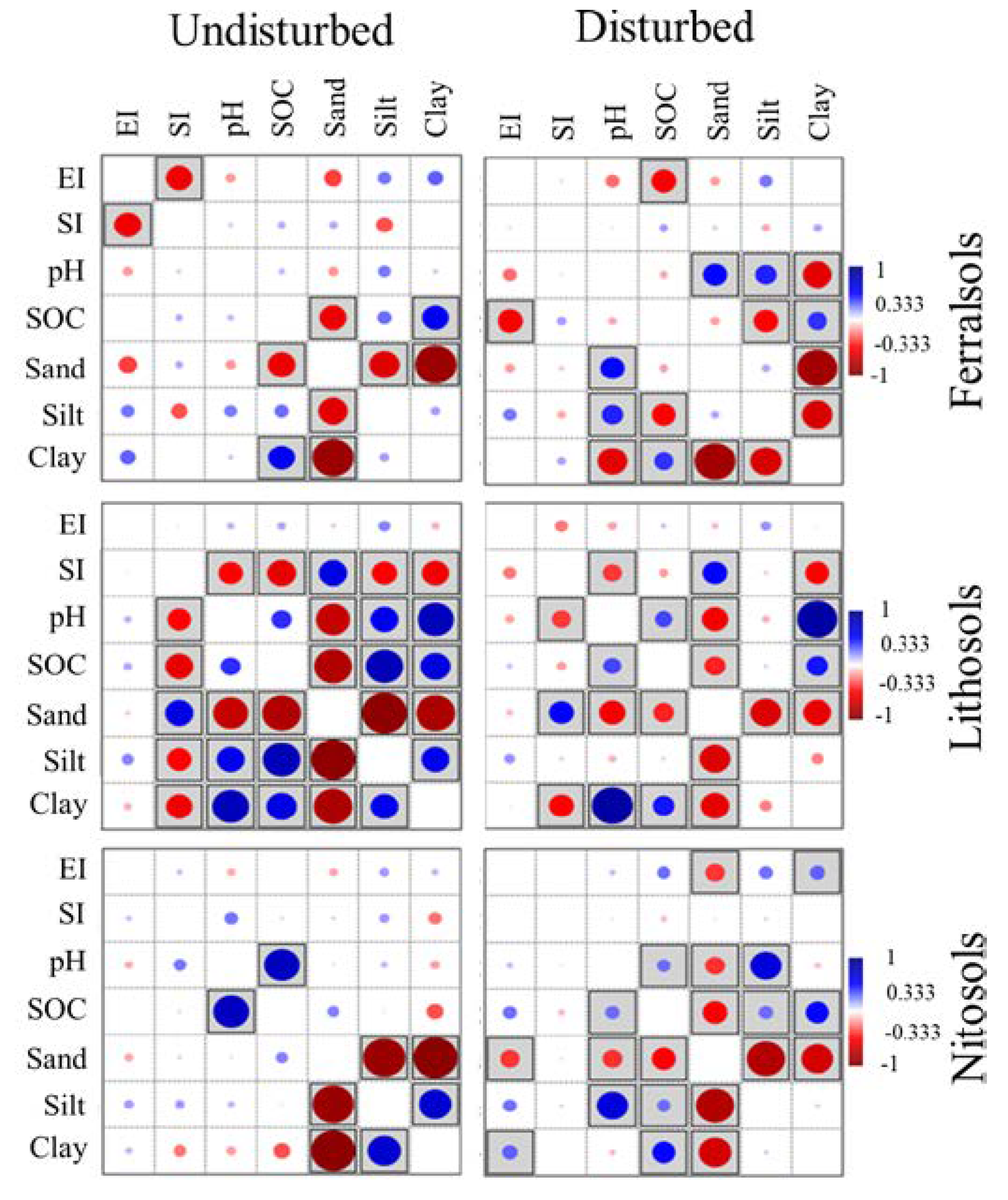
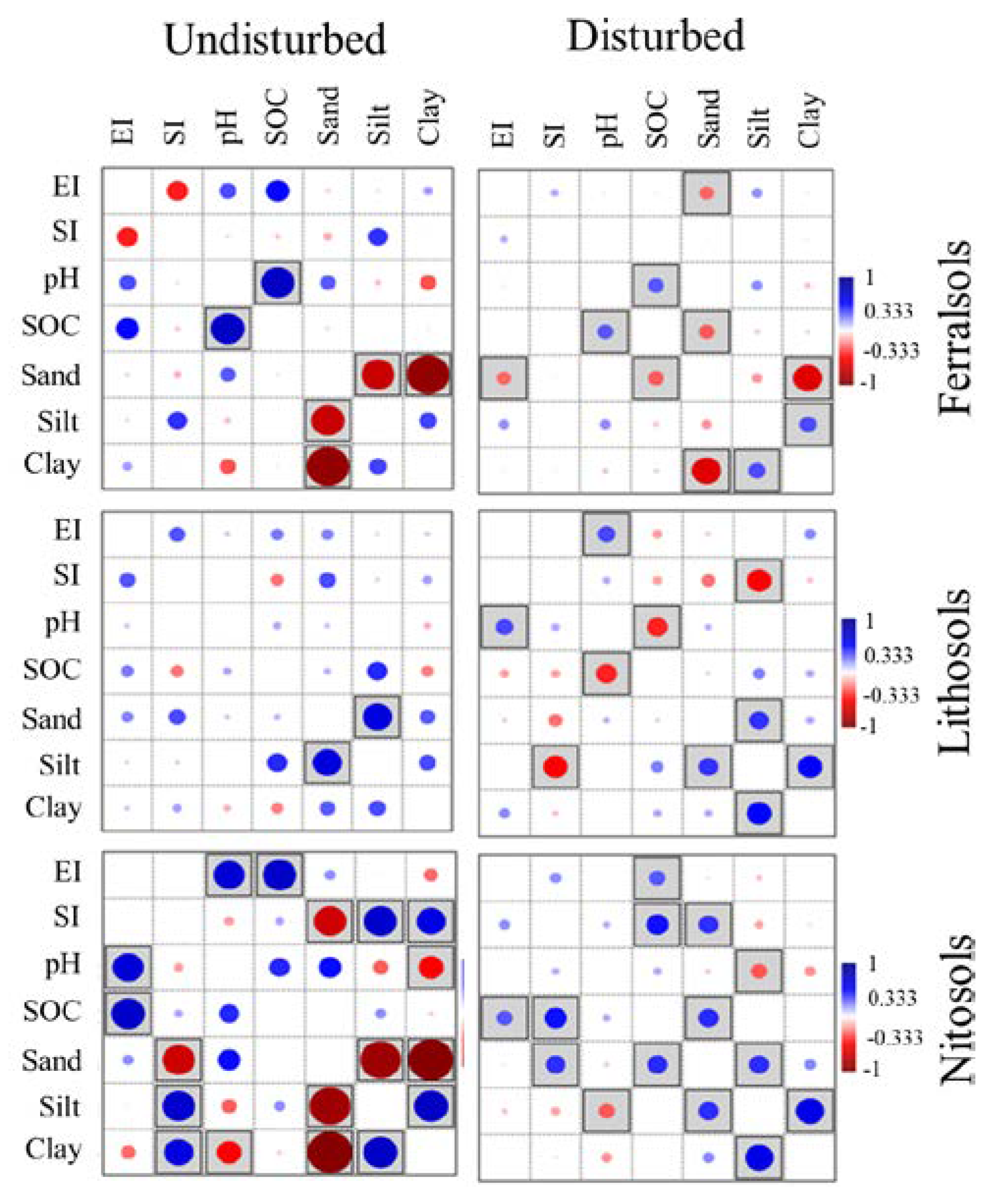
3.3. Bivariate Pearson Correlations of Biophysicochemical Parameters
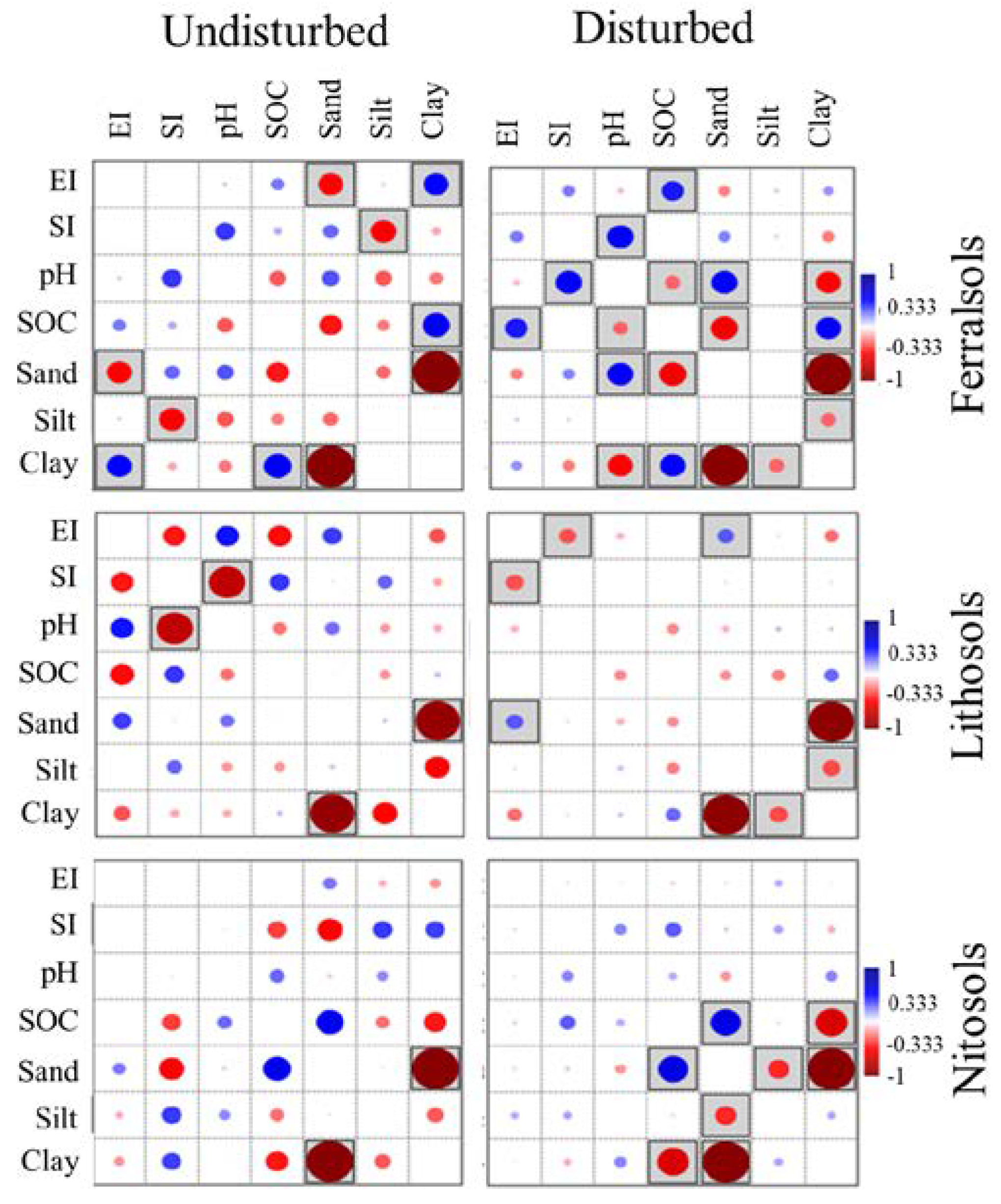
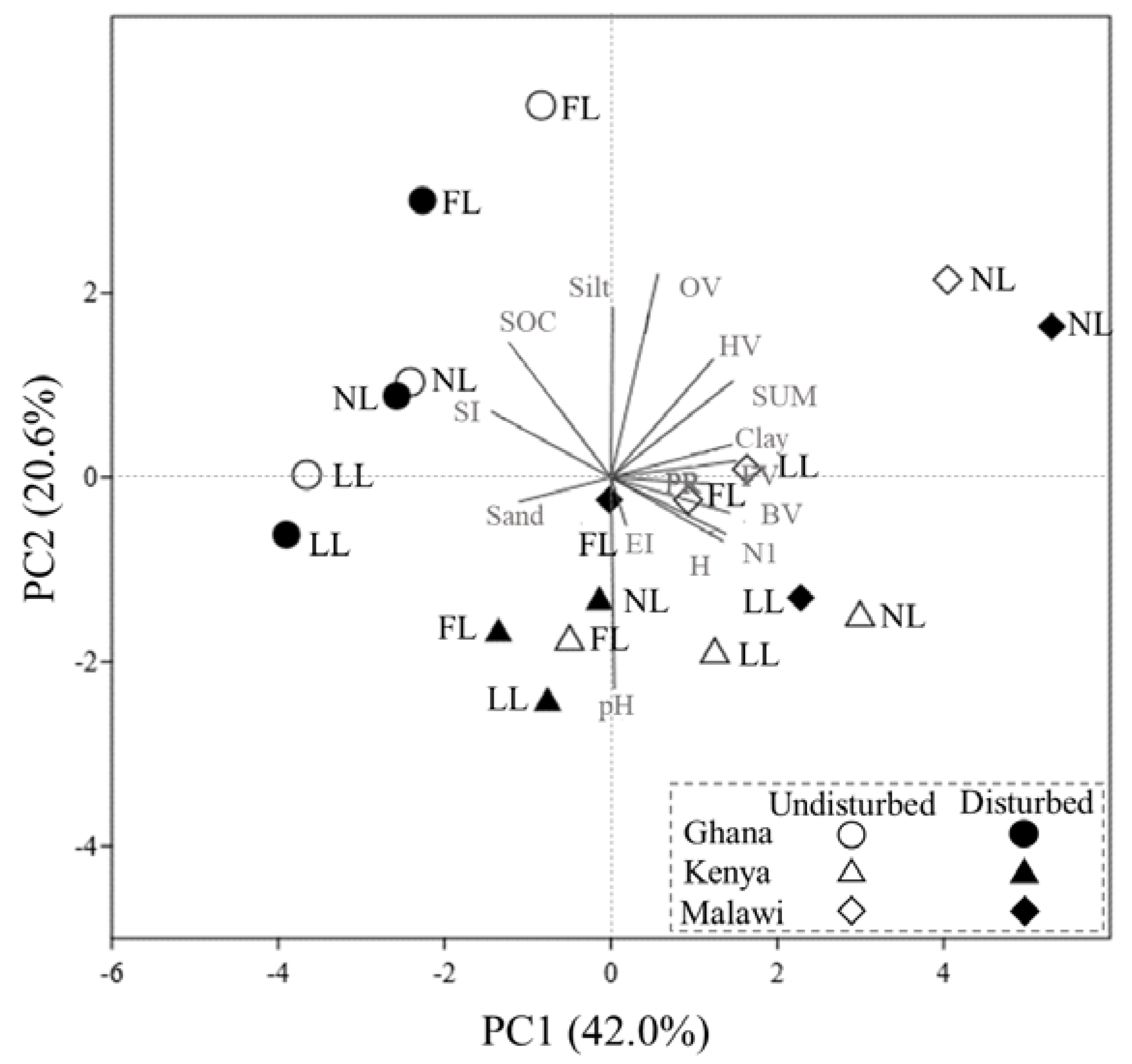
3.4. Multivariate Principal Component Analysis
4. Discussion
4.1. Nematode Trophic Group Abundance and Diversity
4.2. SFW Structure
4.3. Bivariate Correlations
4.4. Multivariate Correlations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beye, G. Impact of Foreign Assistance on Institutional Development of National Agricultural Research Systems in Sub-Saharan Africa; FAO Research and Technology Paper 10; FAO: Rome, Italy, 2002; p. 109. [Google Scholar]
- Diagana, B. Land Degradation in Sub-Saharan Africa: What Explains the Widespread Adoption of Unsustainable Farming practices? Department of Agricultural Economics and Economics, Montana State University: Bozeman, MT, USA, 2003; p. 17. [Google Scholar]
- Lal, R. Soil health and climate change: An overview. In Soil Health and Climate Change; Singh, B.P., Cowie, A.L., Chan, K.Y., Eds.; Springer: Berlin/Heidelberg, Germany; Dordrech, The Netherlands; London, UK; New York, NY, USA, 2011; pp. 3–24. [Google Scholar]
- Assefa, F.; Elias, E.; Soromessa, T.; Ayele, G. Effect of changes in land-use management practices on soil physiochemical properties in Kabe Watershed, Ethiopia. Air Soil Water Res. 2020, 13. [Google Scholar] [CrossRef]
- Doran, J.W.; Zeiss, M.R. Soil health and sustainability: Managing the biotic component of soil quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- Awale, R.; Machado, S.; Ghimire, R.; Bista, P. Soil health. In Advances in Dryland Farming in the Inland Pacific Northwest; Yorgey, G., Kruger, C., Eds.; Washington State University Extension Publication: Pullman, WA, USA, 2017; pp. 47–98. [Google Scholar]
- Parrott, L. Complexity and limits of ecological engineering. Trans. Am. Soc. Agric. Eng. 2002, 45, 1697–1702. [Google Scholar] [CrossRef]
- Proulx, R. Ecological complexity for unifying ecological theory across scales: A field ecologist’s perspective. Ecol. Complex. 2007, 4, 85–92. [Google Scholar] [CrossRef]
- Richter, D.B.; Hofmockel, M.; Callahan, M.A.; Powelson, D.S.; Smith, P. Long-term soil experiments: Keys to managing earth’s rapidly changing ecosystems. Soil Sci. Soc. Am. J. 2007, 71, 266–279. [Google Scholar] [CrossRef]
- Jian, J.; Du, X.; Stewart, R.D. A database for global soil health assessment. Nat. Sci. Data 2020, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Kihara, J.; Bolo, P.; Kinyua, M.; Nyawira, S.S.; Sommer, R. Soil health and ecosystem services: Lessons from sub-Saharan Africa. Geoderma 2019, 370, 141342. [Google Scholar] [CrossRef]
- Stewart, Z.P.; Pierzynski, G.M.; Middendorf, B.J.; Prasad, P.V.V. Approaches to improve soil fertility in sub-Saharan Africa. J. Exp. Bot. 2020, 71, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Fine, A.K.; van Es, H.M.; Schindelbeck, R.R. Statistics, Scoring Functions, and Regional Analysis of a Comprehensive Soil Health Database. Soil Sci. Soc. Am. J. 2017, 81, 589. [Google Scholar] [CrossRef]
- Liu, T.; Hu, F.; Li, H. Spatial ecology of soil nematodes: Perspectives from global to micro scales. Soil Biol. Biochem. 2019, 137, 107565. [Google Scholar] [CrossRef]
- Rayamajhi, N.; Manandhar, B. Impact of climate change and adaptation measures on transhumance herding system in Gatlang, Rasuwa. Air Soil Water Res. 2020, 13. [Google Scholar] [CrossRef]
- Wander, M.M.; Cihacek, L.J.; Coyne, M.; Drijber, R.A.; Grossman, J.M.; Gutknecht, J.L.M.; Horwath, W.R.; Jagandamma, S.; Olk, D.C.; Ruark, M.; et al. Developments in agricultural soil quality and health: Reflections by the research committee on soil organic matter management. Front. Environ. Sci. 2019, 7, 1–9. [Google Scholar] [CrossRef]
- Bongers, T.; Ferris, H. Nematode community structure as a bioindicator in environmental monitoring. Trends Evol. Ecol. 1999, 14, 224–228. [Google Scholar] [CrossRef]
- Sánchez-Moreno, S. Biodiversity and soil health: The role of the soil food web in soil fertility and suppressiveness to soil-borne diseases. Acta Hortic. 2018, 1196, 95–104. [Google Scholar] [CrossRef]
- Kimenju, J.W.; Karanja, N.K.; Mutua, G.K.; Rimberia, B.M.; Wachira, P.M. Nematode community structure as influenced by land use and intensity of cultivation. Trop. Subtrop. Agroecosyst. 2009, 11, 353–360. [Google Scholar]
- Levi, T.; Sherman, C.; Pen-Mouratov, S.; Steinberger, Y. Changes in soil free-living nematode communities and their trophic composition along a climatic gradient. Open Ecol. J. 2012, 2, 79–89. [Google Scholar] [CrossRef][Green Version]
- Gebremikael, M.T.; Steel, H.; Bert, W.; Maenhout, P.; Sleutel, S.; De Neve, S. Quantifying the contribution of entire free-living nematode communities to carbon mineralization under contrasting C and N availability. PLoS ONE 2015, 10, e0136244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ferris, H.; Mitchell, J.; Liang, W. Ecosystem services of the soil food web after long-term application if agricultural management practices. Soil Biol. Biochem. 2017, 111, 36–43. [Google Scholar] [CrossRef]
- Treonis, A.M.; Undangst, S.K.; Kepler, R.M.; Buyer, J.S.; Cavegelli, M.A.; Mirsky, S.B.; Maul, J.E. Characterization of soil nematode communities in three cropping systems through morphological and DNA metabarcoding approaches. Nat. Sci. Rep. 2018, 8, 2004. [Google Scholar] [CrossRef] [PubMed]
- Van der Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi,, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Renco, M.; Gomoryova, E.; Cerevkova, A. The effect of soil type and ecosystems on the soil nematode and microbial communities. Helminthologia 2020, 57, 129–144. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index, an ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Yeates, G.W. Modification and qualification of the nematode maturity index. Pedobiologia 1995, 38, 97–101. [Google Scholar]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Sánchez-Moreno, S.; Nicola, N.L.; Ferris, H.; Zalom, F.G. Effects of agricultural management on nematode–mite assemblages: Soil food web indices as predictors of mite community composition. Appl. Soil Ecol. 2009, 41, 107–117. [Google Scholar] [CrossRef]
- Ruess, L. Nematode soil faunal analysis of decomposition pathways in different ecosystems. Nematology 2003, 5, 179–181. [Google Scholar] [CrossRef]
- Asiedu, O.; Kwoseh, C.K.; Melakeberhan, H.; Adjeigyapong, T. Nematode distribution in cultivated and undisturbed soils of Guinea Savannah and Semi-deciduous Forest zones of Ghana. Geosci. Front. 2019, 10, 381–387. [Google Scholar] [CrossRef]
- FAO. Land and Water Development Division; FAO: Rome, Italy, 2003. [Google Scholar]
- Thuo, A.K.; Karuku, G.N.; Kimenju, J.W.; Kariuku, G.M.; Wendot, P.K.; Melakeberhan, H. Seasonal variation of nematode assemblage and diversity on selected soil groups in Kenya: Vertisols, Cambisols and Arenosols. Trop. Subtrop. Agroecosystems 2020, 23, 63. [Google Scholar]
- Thuo, A.K.; Karuku, G.N.; Kimenju, J.W.; Kariuku, G.M.; Wendot, P.K.; Melakeberhan, H. Factors influencing the relationship between nematode communities and edaphic factors on selected soil groups in Kenya: Vertisols, Cambisols and Arenosols. Trop. Subtrop. Agroecosystems 2020, 23, 49. [Google Scholar]
- Lima da Silva, J.V.C.; Hirschfeld, M.N.C.; Cares, J.E.; Esteves, A.M. Land use, soil properties and climate variables influence the nematode communities in the Caatinga dry forest. Appl. Soil Ecol. 2019, 103474. [Google Scholar] [CrossRef]
- Hánĕl, L. Soil nematodes in cambisol agroecosystem of the Czeck Republic. Biol. Brast. 2003, 22, 205–216. [Google Scholar]
- Lišková, M.; Čerevková, A.; Hanel, L. Nematode communities of forest ecosystems in association with various soil orders. Russ. J. Nematol. 2008, 16, 129–142. [Google Scholar]
- Sant’Anna, R. Major Soils for Food Production in Africa; FAO Soils Bulletin 69; FAO: Rome, Italy, 1993; Available online: http://www.fao.org/docrep/t1696e/t1696e07.htm (accessed on 4 February 2021).
- Melakeberhan, H.; Maung, Z.T.Z.; Lee, C.-L.; Poindexter, S.; Stewart, J. Soil type-driven variable effects on cover- and rotation-crops, nematodes and soil food web in sugar beet fields reveal a roadmap for developing healthy soils. Eur. J. Soil Biol. 2018, 85, 53–63. [Google Scholar] [CrossRef]
- Nelson, E.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis Part III; Sparks, D.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Glendon, W.G.; Doni, O.R. Particle-size analysis. In Methods of Soil Analysis Part. IV; Dane, J.H., Topp, G.C., Eds.; Soil Science Society of America: Madison, WI, USA, 2002; pp. 264–289. [Google Scholar]
- Bouyoucos, G.J. A recalibration of the hydrometer method for making mechanical analysis of soils. Agron. J. 1951, 43, 434–438. [Google Scholar] [CrossRef]
- Ayoub, S.M. Plant Nematology: An Agricultural Training Aid; California Department of Food and Agriculture, NemaAid Publications: Sacramento, CA, USA, 1977; p. 195. [Google Scholar]
- Hooper, D.J. Handling, fixing, staining and mounting nematodes. In Laboratory Methods for Work with Plant and Soil Nematodes; Southey, J.F., Ed.; MAFF: London, UK, 1986; pp. 59–80. [Google Scholar]
- Bongers, T. De Nematoden van Nederland, 2nd ed.; KNNV Uuitgaeverij: Schoorl, The Netherlands, 1994; p. 408. [Google Scholar]
- Okada, H.; Kadota, I. Host status of 10 fungal isolates for two nematode species, Filenchus misellus and Aphelenchus avenae. Soil Biol. Biochem. 2003, 35, 1601–1607. [Google Scholar] [CrossRef]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; de Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera an outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1998; p. 144. [Google Scholar]
- SAS Institute Inc. SAS OnlineDoc 9.4; SAS Institute Inc.: Cary, NC, USA, 2018. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Van Diepeningen, A.D.; de Vos, O.J.; Korthals, G.W.; van Bruggen, A.H.C. Effects of organic versus conventional management on chemical and biological parameters in agricultural soils. Appl. Soil Ecol. 2006, 31, 120–135. [Google Scholar] [CrossRef]
- Hánĕl, L. Recovery of soil nematode populations from cropping stress by natural secondary succession to meadow land. Appl. Soil Ecol. 2003, 22, 255–270. [Google Scholar] [CrossRef]
- Talwana, H.L.; Butseya, M.M.; Tusiime, G. Occurrence of plant parasitic nematodes and factors that enhance population build-up in cereal-based cropping systems in Uganda. Afr. Crop. Sci. J. 2008, 16, 119–131. [Google Scholar]
- Ferris, H.; Griffiths, B.S.; Porazinska, D.L.; Powers, T.O.; Wang, K.; Tenuta, M. Reflections on Plant and Soil Nematode Ecology: Past, Present and Future. J. Nematol. 2012, 44, 115–126. [Google Scholar] [PubMed]
- Ferris, H. Contribution of nematodes to the structure and function of the soil food web. J. Nematol. 2010, 42, 63–67. [Google Scholar] [PubMed]
- McSorley, R.; Wang, K.H.; Church, G. Suppression of root-knot nematodes in natural and agricultural soils. Appl. Soil Ecol. 2008, 39, 291–298. [Google Scholar] [CrossRef]
- Hartemink, A.E.; Krasilnikov, P.; Bockheim, J.G. Soil maps of the world. Geoderma 2013, 208, 256–267. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trophic Group | c-p | Trophic Group | c-p | Trophic Group | c-p |
|---|---|---|---|---|---|
| Herbivores | Bacterivores | Predators | |||
| Cephalenchus | 2 | Panagrolaimus | 1 | Tripyla | 3 |
| Ecphyadophora | 2 | Rhabditis | 1 | Clarkus | 4 |
| Gracilacus | 2 | Acrobeles | 2 | Mononchidae | 4 |
| Paratylenchus | 2 | Acrobeloides | 2 | Mononchus | 4 |
| Psilenchus | 2 | Cephalobus | 2 | Seinura | 4 |
| Tylenchus | 2 | Cervidellus | 2 | Discolaimoides | 5 |
| Amplimerlinius | 3 | Chiloplacus | 2 | Discolaimus | 5 |
| Antarctylus | 3 | Eucephalobus | 2 | Nygolaimus | 5 |
| Criconema | 3 | Heterocephalobus | 2 | ||
| Criconemella | 3 | Leptolaimidae | 2 | Omnivores | |
| Criconemoides | 3 | Monhysteridae | 2 | Campydora | 4 |
| Discocriconemella | 3 | Plectus | 2 | Dorylaimellus | 4 |
| Helicotylenchus | 3 | Wilsonema | 2 | Dorylaimidae | 4 |
| Hemicycliophora | 3 | Zeldia | 2 | Dorylaimoides | 4 |
| Heterodera | 3 | Achromadora | 3 | Dorylaimus | 4 |
| Hirschmanniella | 3 | Prismatolaimus | 3 | Ecumericus | 4 |
| Hoplolaimus | 3 | Alaimus | 4 | Enchodelus | 4 |
| Meloidogyne | 3 | Labronema | 4 | ||
| Merlinius | 3 | Fungivores | Mesodorylaimus | 4 | |
| Nagelus | 3 | Aphelenchoides | 2 | Paraxonchium | 4 |
| Paratrophurus | 3 | Aphelenchus | 2 | Pungentus | 4 |
| Pratylenchus | 3 | Deladenus | 2 | Actinolaimidae | 5 |
| Rotylenchulus | 3 | Ditylenchus | 2 | Aparcelaimus | 5 |
| Rotylenchus | 3 | Filenchus | 2 | Aporcelaimellus | 5 |
| Trichotylenchus | 3 | Diphterophora | 3 | Belondira | 5 |
| Trophurus | 3 | Leptonchus | 4 | Belondirella | 5 |
| Tylenchorhynchus | 3 | Tylencholaimus | 4 | Egtitus | 5 |
| Paratrichodorus | 4 | Tyleptus | 4 | Fuschelia | 5 |
| Trichodorus | 4 | Prodorylaimus | 5 | ||
| Longidorus | 5 | ||||
| Xiphinema | 5 |
| Factors | Nematodes Trophic Group | SUM | Diversity | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| HV | BV | FV | PR | OV | H | N1 | ||||
| Ghana | SG | FL | 28 ± 0.25 | 5 ± 0.14 a | 1 ± 0.19 | 1 ± 0.11 | 7 ± 0.14 a | 47 ± 0.17 | 1.6 ± 0.12 | 5.9 ± 0.75 |
| LL | 17 ± 0.23 | 4 ± 0.13 ab | 2 ± 0.18 | 0 ± 0.10 | 3 ± 0.13 b | 31 ± 0.16 | 1.5 ± 0.11 | 5.2 ± 0.71 | ||
| NL | 26 ± 0.20 | 3 ± 0.11 b | 2 ± 0.16 | 0 ± 0.09 | 4 ± 0.11 b | 42 ± 0.14 | 1.5 ± 0.10 | 4.9 ± 0.60 | ||
| RG | N | 25 ± 0.20 | 5 ± 0.12 a | 1 ± 0.16 | 0 ± 0.09 | 5 ± 0.12 | 41 ± 0.14 | 2.0 ± 0.10 a | 5.9 ± 0.60 | |
| S | 22 ± 0.16 | 3 ± 0.08 b | 2 ± 0.13 | 1 ± 0.07 | 4 ± 0.08 | 38 ± 0.12 | 1.0 ± 0.08 b | 4.8 ± 0.47 | ||
| LS | U | 32 ± 0.22 a | 5 ± 0.12 a | 1 ± 0.17 | 0 ± 0.10 | 5 ± 0.12 a | 50 ± 0.15 a | 1.58 ± 0.11 | 5.5 ± 0.65 | |
| D | 17 ± 0.14 b | 3 ± 0.08 b | 2 ± 0.11 | 0 ± 0.06 | 3 ± 0.08 b | 31 ± 0.10 b | 1.55 ± 0.07 | 5.2 ± 0.40 | ||
| F values | RG*LS | 0.00 | 0.47 | 3.55 | 0.01 | 1.07 | 0.01 | 0.14 | 0.13 | |
| SG*LS | 0.10 | 2.87 | 0.07 | 1.07 | 1.68 | 0.23 | 0.35 | 0.37 | ||
| RG*SG | 4.01 | 6.93 * | 1.21 | 0.59 | 9.42 * | 4.81 * | 0.02 | 0.49 | ||
| RG*SG*LS | 0.90 | 0.82 | 0.16 | 0.38 | 0.85 | 0.64 | 0.60 | 0.47 | ||
| Kenya | SG | FL | 13 ± 0.18 b | 7 ± 0.15b a | 3 ± 0.16 ba | 0 ± 0.11 b | 3 ± 0.13 | 29 ± 0.16 b | 1.8 ± 0.08 | 7 ± 0.46 b |
| LL | 16 ± 0.18 b | 11 ± 0.15 a | 2 ± 0.16 b | 1 ± 0.11 a | 2 ± 0.13 | 37 ± 0.16 ba | 1.7 ± 0.08 | 7 ± 0.46 b | ||
| NL | 27 ± 0.18 a | 6 ± 0.15 b | 5 ± 0.16 a | 1 ± 0.11 a | 3 ± 0.13 | 47 ± 0.16 a | 1.9 ± 0.08 | 8 ± 0.46 a | ||
| RG | N | 14 ± 0.15 | 5 ± 0.12 b | 3 ± 0.13 | 1 ± 0.09 | 2 ± 0.10 | 30 ± 0.13 b | 1.9 ± 0.06 | 7 ± 0.36 | |
| S | 21 ± 0.15 | 13 ± 0.12 a | 3 ± 0.13 | 1 ± 0.09 | 2 ± 0.10 | 45 ± 0.13 a | 1.9 ± 0.06 | 7 ± 0.36 | ||
| LS | U | 29 ± 0.18 a | 13 ± 0.15 a | 4 ± 0.16 a | 1 ± 0.11 | 3 ± 0.13 | 56 ± 0.16 a | 2a ± 0.08 | 8 ± 0.46 a | |
| D | 11 ± 0.10 b | 5 ± 0.08 b | 2 ± 0.09 b | 1 ± 0.06 | 2 ± 0.07 | 24 ± 0.09 b | 1b ± 0.04 | 7 ± 0.25 b | ||
| F values | RG*LS | 0.39 | 1.78 | 6.51 * | 1.60 | 3.67 | 0.73 | 0.51 | 1.86 | |
| SG*LS | 0.67 | 0.72 | 1.98 | 0.43 | 3.64 | 0.91 | 1.32 | 2.56 | ||
| RG*SG | 1.55 | 0.94 | 3.22 | 1.50 | 4.09 * | 0.87 | 4.37 * | 4.08 * | ||
| RG*SG*LS | 0.23 | 3.18 | 1.07 | 0.61 | 3.24 | 0.29 | 3.21 | 2.83 | ||
| Malawi | SG | FL | 19 ± 0.37 b | 9 ± 0.15 b | 6 ± 0.19 b | 2 ± 0.11 | 5 ± 0.16 b | 51 ± 0.25 b | 1.6 ± 0.10 | 7.7 ± 0.62 |
| LL | 18 ± 0.38 b | 21 ± 0.16 a | 11 ± 0.20 ba | 2 ± 0.11 | 4 ± 0.16 b | 70 ± 0.26 b | 1.9 ± 0.11 | 7.1 ± 0.65 | ||
| NL | 92 ± 0.41 a | 16 ± 0.19 a | 16 ± 0.23 a | 2 ± 0.13 | 9 ± 0.18 a | 156 ± 0.28 a | 1.8 ± 0.12 | 6.4 ± 0.79 | ||
| RG | N | 25 ± 0.30 | 13 ± 0.13 | 11 ± 0.16 | 2 ± 0.09 | 4 ± 0.13 b | 62 ± 0.21 | 1.74 ± 0.09 | 7 ± 0.51 | |
| S | 40 ± 0.31 | 16 ± 0.14 | 10 ± 0.17 | 2 ± 0.10 | 7 ± 0.14 a | 95 ± 0.21 | 1.8 ± 0.09 | 7 ± 0.55 | ||
| LS | U | 38 ± 0.39 | 13 ± 0.17 | 8 ±0.21 b | 2 ± 0.12 | 7 ± 0.17 a | 75 ± 0.26 | 1.8 ± 0.11 | 7.5 ± 0.69 | |
| D | 26 ± 0.21 | 16 ± 0.09 | 12 ± 0.12 a | 2 ± 0.07 | 4 ± 0.09 b | 78 ± 0.15 | 1.7 ± 0.06 | 6.6 ± 0.35 | ||
| F values | RG*LS | 0.40 | 0.05 | 0.00 | 2.22 | 2.19 | 0.06 | 0.02 | 0.01 | |
| SG*LS | 2.70 | 5.39 * | 0.32 | 3.82 | 0.29 | 0.19 | 0.34 | 0.58 | ||
| RG*SG | 0.15 | 7.98 * | 0.10 | 0.14 | 4.61 * | 1.11 | 2.65 | 1.62 | ||
| RG*SG*LS | 1.10 | 1.48 | 0.18 | 0.69 | 2.26 | 1.05 | 1.32 | 2.30 | ||
| SG | Country | Undisturbed | Disturbed | ||
|---|---|---|---|---|---|
| SI | EI | SI | EI | ||
| FL | GA | 80.3 ± 9.1 ABC | 13.7 ± 5.5 | 77.8 ± 6.6 AB | 18.5 ± 4.1 ED |
| KY | 67.7 ± 8.4 BCD | 26.1 ± 4.8 | 68.2 ± 4.9 BC | 30.6 ± 2.8 ABC | |
| ML | 57.8 ± 8.4 a D | 35.7 ± 4.8 | 31.9 ± 4.9 b E | 36.0 ± 2.8 AB | |
| LL | GA | 81.4 ± 9.1 AB | 26.0 ± 5.5 | 88.3 ± 5.8 A | 28.3 ± 3.4 ABC |
| KY | 52.0 ± 8.4 b ED | 20.8 ± 4.8 | 62.4 ± 4.9 a CD | 26.0 ± 2.8 CD | |
| ML | 67.7 ± 8.7 a BCD | 29.3 ± 5.2 | 52.1 ± 5.0 b D | 37.0 ± 2.9 A | |
| NL | GA | 85.8 ± 8.4 A | 32.0 ± 4.8 | 77.7 ± 4.9 AB | 27.7 ± 2.8 BCD |
| KY | 38.5 ± 8.4 b E | 18.8 ± 4.8 | 62.8 ± 4.9 a CD | 10.8 ± 2.8 E | |
| ML | 54.1 ± 9.5 a CD | 19.2 ± 5.9 | 42.1 ± 5.4 b E | 31.3 ± 3.3 ABC | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melakeberhan, H.; Maung, Z.; Lartey, I.; Yildiz, S.; Gronseth, J.; Qi, J.; Karuku, G.N.; Kimenju, J.W.; Kwoseh, C.; Adjei-Gyapong, T. Nematode Community-Based Soil Food Web Analysis of Ferralsol, Lithosol and Nitosol Soil Groups in Ghana, Kenya and Malawi Reveals Distinct Soil Health Degradations. Diversity 2021, 13, 101. https://doi.org/10.3390/d13030101
Melakeberhan H, Maung Z, Lartey I, Yildiz S, Gronseth J, Qi J, Karuku GN, Kimenju JW, Kwoseh C, Adjei-Gyapong T. Nematode Community-Based Soil Food Web Analysis of Ferralsol, Lithosol and Nitosol Soil Groups in Ghana, Kenya and Malawi Reveals Distinct Soil Health Degradations. Diversity. 2021; 13(3):101. https://doi.org/10.3390/d13030101
Chicago/Turabian StyleMelakeberhan, Haddish, ZinThuZar Maung, Isaac Lartey, Senol Yildiz, Jenni Gronseth, Jiaguo Qi, George N. Karuku, John W. Kimenju, Charles Kwoseh, and Thomas Adjei-Gyapong. 2021. "Nematode Community-Based Soil Food Web Analysis of Ferralsol, Lithosol and Nitosol Soil Groups in Ghana, Kenya and Malawi Reveals Distinct Soil Health Degradations" Diversity 13, no. 3: 101. https://doi.org/10.3390/d13030101
APA StyleMelakeberhan, H., Maung, Z., Lartey, I., Yildiz, S., Gronseth, J., Qi, J., Karuku, G. N., Kimenju, J. W., Kwoseh, C., & Adjei-Gyapong, T. (2021). Nematode Community-Based Soil Food Web Analysis of Ferralsol, Lithosol and Nitosol Soil Groups in Ghana, Kenya and Malawi Reveals Distinct Soil Health Degradations. Diversity, 13(3), 101. https://doi.org/10.3390/d13030101