
A Giant Ostrich from the Lower Pleistocene Nihewan Formation of North China, with a Review of the Fossil Ostriches of China



Abstract
1. Introduction
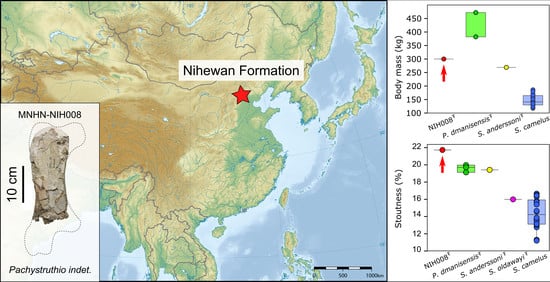
2. Discovery and Geological Setting of the Specimen
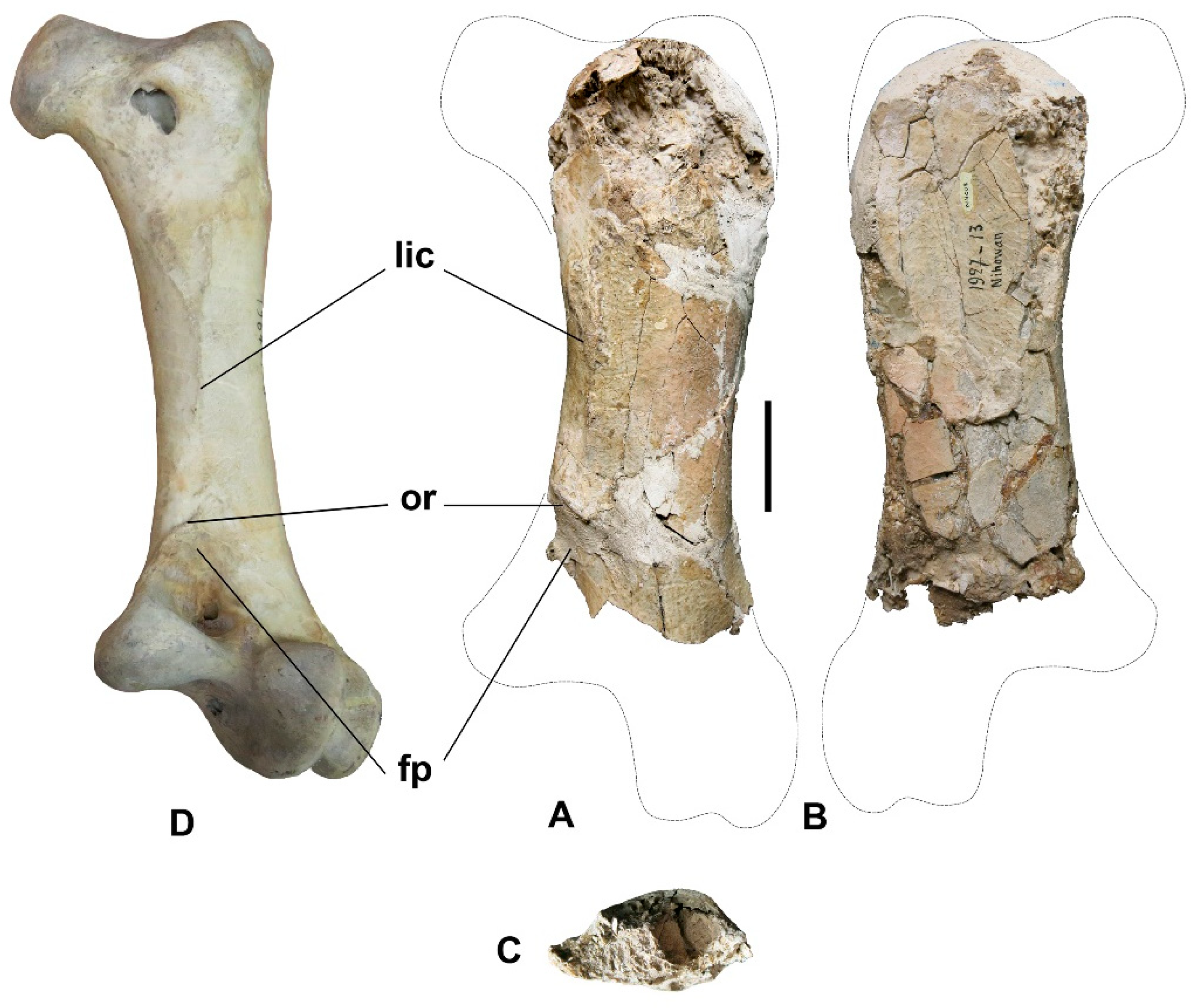
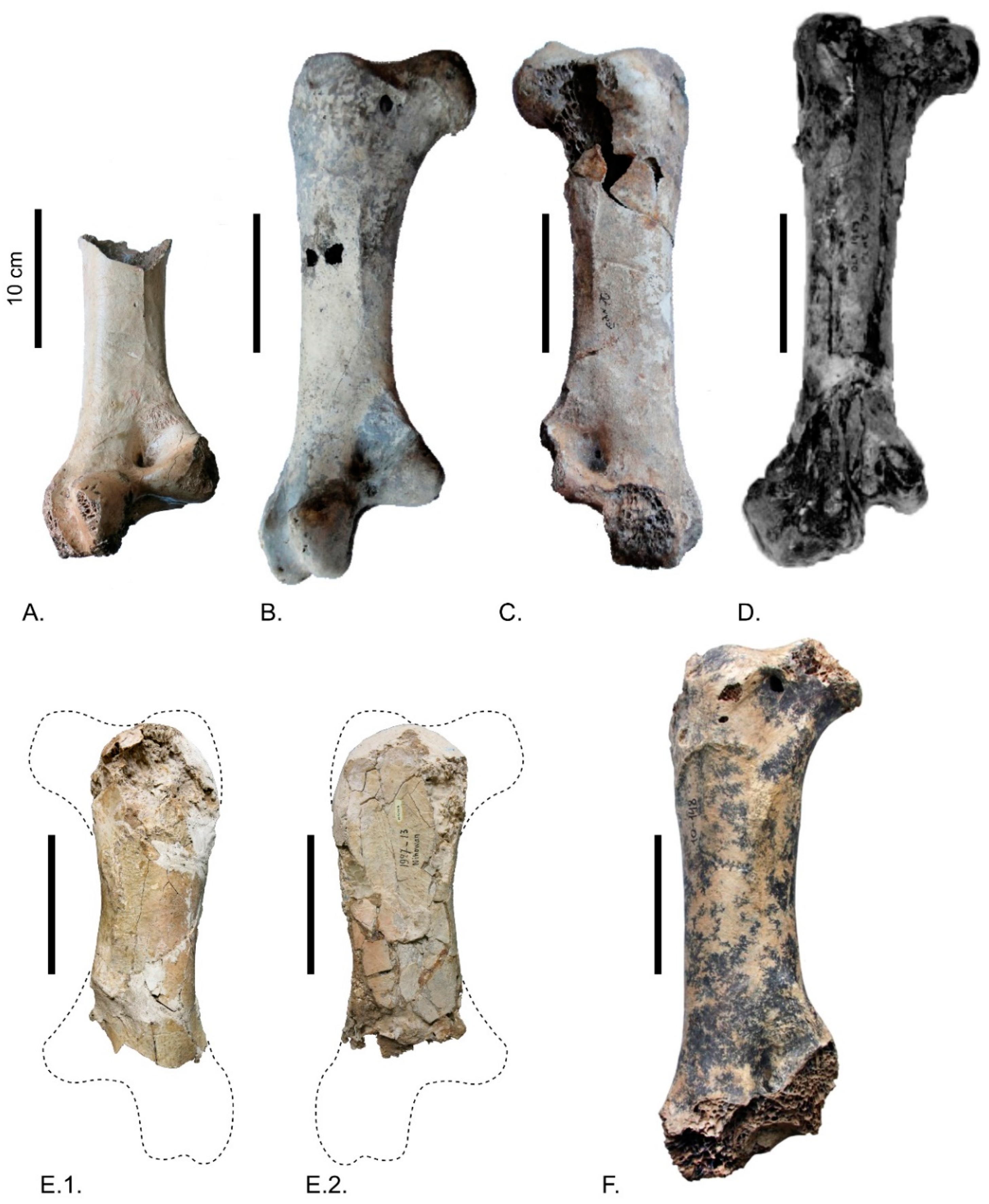
3. Description
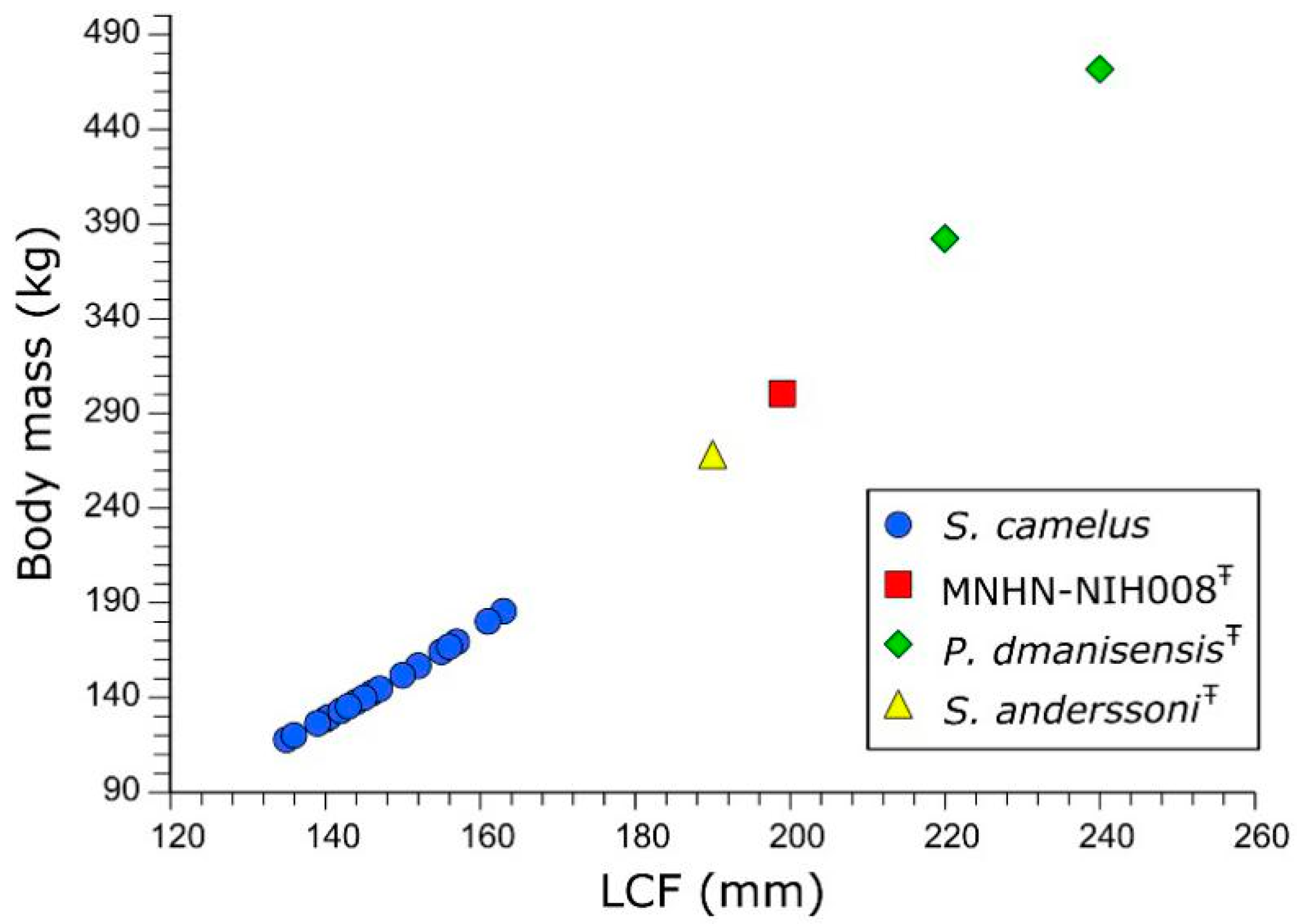
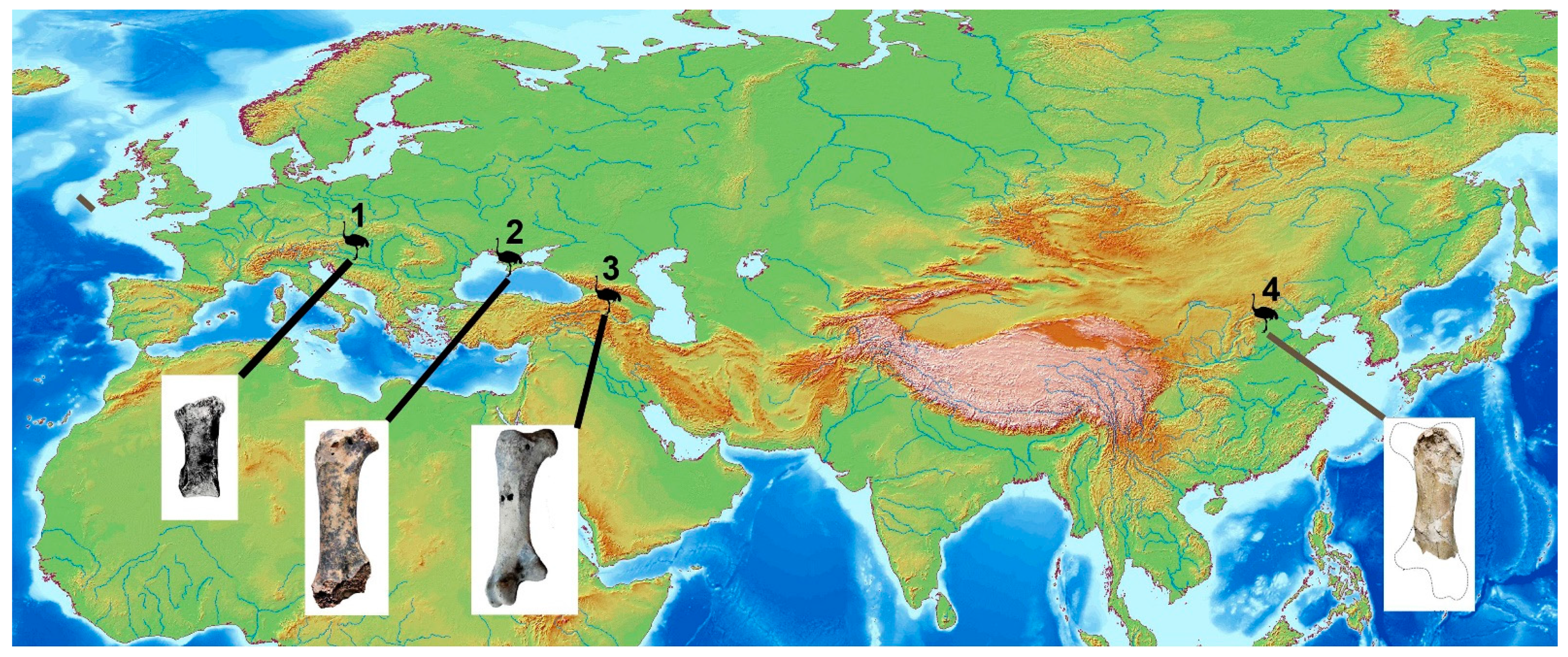
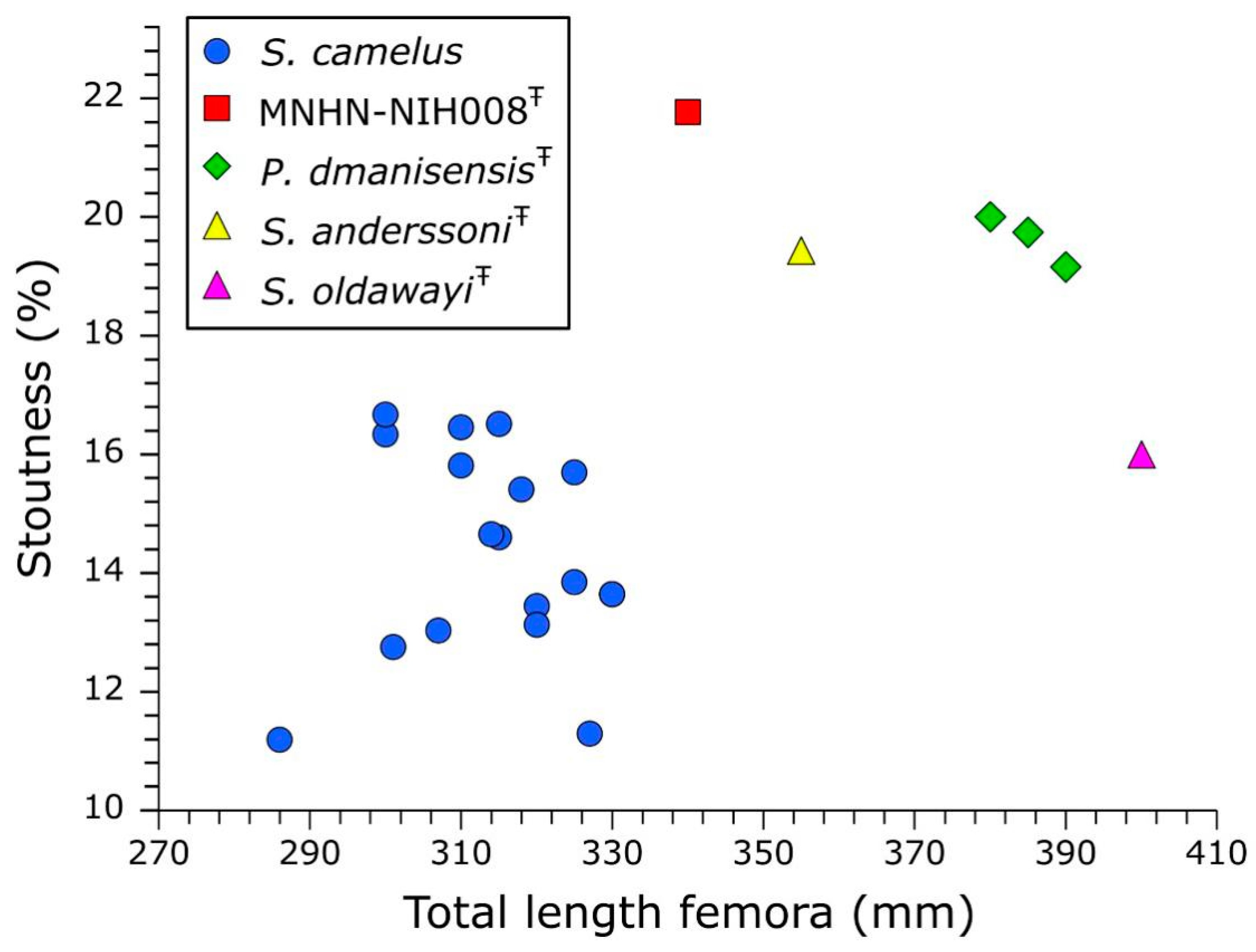
4. Body Mass Estimate, Comparison with Other Giant Ostriches and Identification
5. A Brief Review of the Fossil Record of Ostriches in China
6. Conclusions: The Giant Ostrich from Nihewan in Its Environment
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ao, H.; Deng, C.; Dekkers, M.J.; Sun, Y.; Liu, Q.; Zhu, R. New evidence for early presence of hominids in North China. Sci. Rep. 2013, 3, 2403. [Google Scholar] [CrossRef] [PubMed]
- Teilhard de Chardin, P.; Piveteau, J. Les mammifères fossiles de Nihowan (Chine). Ann. Paléont. 1930, 19, 1–134. [Google Scholar]
- Wang, M.; O’Connor, J.K.; Zhou, Z. The first fossil crow (Corvus sp. indet.) from the Early Pleistocene Nihewan Paleolithic sites in North China. J. Archaeol. Sc. 2013, 40, 1623–1628. [Google Scholar] [CrossRef]
- Licent, E. Voyage aux terrasses du Sang Kan Ho, à l’entrée de la plaine de Sining Hien. Pub. Mus. Hoang Ho Pai Ho 1924, 4, 1–14. [Google Scholar]
- Barbour, G.B. Preliminary observations in the Kalgan area. Bull. Geol. Soc. China 1924, 3, 153–168. [Google Scholar] [CrossRef]
- Barbour, G.B.; Licent, E.; Teilhard de Chardin, P. Geological study of the deposits of the Sangkanho basin. Bull. Geol. Soc. China 1926, 5, 263–278. [Google Scholar] [CrossRef]
- Barbour, G.B. The deposits of the Sang Kan Ho valley. Bull. Geol. Soc. China 1925, 4, 53–55. [Google Scholar] [CrossRef]
- Boule, M.; Breuil, H.; Licent, E.; Teilhard, P. Le Paléolithique de la Chine. Arch. Inst. Paléont. Hum. 1928, 4, 1–138. [Google Scholar]
- Lowe, P.R. Struthious remains from Northern China and Mongolia, with descriptions of Struthio wimani, Struthio anderssoni and Struthio mongolicus, spp. nov. Palaeont. Sin. C 1931, 6, 1–47. [Google Scholar]
- Lambrecht, K. Handbuch der Palaeornithologie; Borntrager: Berlin, Germany, 1933. [Google Scholar]
- Leroy, P. L’Institut de Géobiologie à Pékin, 1940–1946. Les dernières années du P. Teilhard de Chardin en Chine. L Anthropologie 1965, 69, 360–367. [Google Scholar]
- Cai, B.Q.; Zheng, S.H.; Liddicoat, J.; Li, Q. Review of the litho-, bio-, and chronostratigraphy in the Nihewan Basin, Hebei, China. In Fossil Mammals of Asia; Wang, X., Flynn, L.J., Fortelius, M., Eds.; Columbia University Press: New York, NY, USA, 2013; pp. 218–242. [Google Scholar]
- Licent, E. Comptes-rendus de onze années (1923–1933) de séjour et d’exploration dans le bassin du Fleuve Jaune, du Pai Ho et des autres tributaires du golfe du Pai Tcheu ly. Pub. Mus. Hoang Ho Pai Ho 1935, 38, 1–1131. [Google Scholar]
- Campbell, K.E.; Marcus, L. The relationship of hindlimb bone dimensions to body weight in birds. Nat. Hist. Mus. Los Angeles County Sc. Ser. 1992, 36, 395–412. [Google Scholar]
- Angst, D.; Buffetaut, E. Paleobiology of Giant Flightless Birds; Elsevier & ISTE Press: Oxford/London, UK, 2017. [Google Scholar]
- Buffetaut, E.; Angst, D. How large was the giant ostrich of China? Evoluçao 2018, 2, 6–8. [Google Scholar]
- Shaw, T.H. Einige Bemerkungen zum Oberschenkelknochen des fossilen Strausses Struthio anderssoni Lowe von Chou Kou Tien in Nord-China. Ornithol. Monatsber. 1937, 45, 201–202. [Google Scholar]
- Hou, L. Avian fossils of Pleistocene from Zhoukoudian. Mem. Inst. Vert. Palaeont. Palaeoanthrop. Acad. Sin. 1993, 19, 165–297. [Google Scholar]
- Lowe, P.R. On some struthious remains: -1. Description of some pelvic remains of a large fossil ostrich, Struthio oldawayi, sp. n., from the Lower Pleistocene of of Oldaway (Tanganyika Territory); 2. Egg-shell fragments referable to Psammornis and other Struthiones collected by H. St. John Philby in southern Arabia. Ibis 1933, 75, 652–657. [Google Scholar]
- Leakey, L.S.B. Olduvai Gorge 1951–1961; Cambridge University Press: Cambridge, UK, 1967; Volume 1. [Google Scholar]
- Burchak-Abramovich, N.I.; Vekua, A. The fossil ostrich Struthio dmanisensis sp. n. from the Lower Pleistocene of eastern Georgia. Acta Zool. Cracov. 1990, 33, 121–132. [Google Scholar]
- Vekua, A. Giant ostrich in Dmanisi fauna. Bull. Georg. Nat. Acad. Sc. 2013, 7, 143–148. [Google Scholar]
- Zelenkov, N.V.; Lavrov, A.V.; Startsev, D.B.; Vislobokova, I.A.; Lopatin, A.V. A giant early Pleistocene bird from eastern Europe: Unexpected component of terrestrial faunas at the time of early Homo arrival. J. Vert. Paleont. 2019. [Google Scholar] [CrossRef]
- Kretzoi, M. Ostrich and camel remains from the Central Danube basin. Acta Geologica 1954, 2, 231–242. [Google Scholar]
- Dennell, R.W. The colonization of “Savannahstan”: Issues of timing(s) and patterns of dispersal across Asia in the Late Pliocene and Early Pleistocene. In Asian Paleoanthropology: From Africa to China and Beyond; Norton, C.J., Braun, D.R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 7–30. [Google Scholar]
- Andersson, J.G. Der Weg über die Steppen. Bull. Mus. Far East. Antiquit. 1929, 1, 143–165. [Google Scholar]
- Wang, S.; Hu, Y.; Wang, L. New ratite eggshell material from the Miocene of Inner Mongolia, China. Chinese Birds 2011, 2, 18–26. [Google Scholar] [CrossRef]
- Mikhailov, K.E.; Zelenkov, N. The late Cenozoic history of the ostriches (Aves: Struthionidae), as revealed by fossil eggshell and bone remains. Earth Sc. Rev. 2020, 208, 103270. [Google Scholar] [CrossRef]
- Hou, L.; Zhou, Z.; Zhang, F.; Wang, Z. A Miocene ostrich fossil from Gansu Province, northwest China. Chin. Sci. Bull. 2005, 50, 1808–1810. [Google Scholar] [CrossRef]
- Andersson, J.G. Essays on the Cenozoic of northern China. Mem. Geol. Surv. China Ser. A 1923, 3, 1–152. [Google Scholar]
- Qiu, Z.X.; Qiu, Z.D.; Deng, T.; Li, C.K.; Zhang, Z.Q.; Wang, B.Y.; Wang, X. Neogene Land Mammal Ages/Stages of China. In Fossil Mammals of Asia; Wang, X., Flynn, L.J., Fortelius, M., Eds.; Columbia University Press: New York, NY, USA, 2013; pp. 29–90. [Google Scholar]
- Young, C.C. On the new finds of fossil eggs of Struthio anderssoni Lowe in North China, with remarks on the egg remains found in Shansi, Shensi and in Choukoutien. Bull. Geol. Soc. China 1933, 12, 145–152. [Google Scholar] [CrossRef]
- Young, C.C.; Sun, A.L. New material of ostrich eggs and its stratigraphic significance. Paleovert. Paleoanthrop. 1960, 2, 115–119. (In Chinese) [Google Scholar]
- Shaw, T.H. Preliminary observations on the fossil birds from Chou-Kou-Tien. Bull. Geol. Soc. China 1935, 14, 77–81. [Google Scholar] [CrossRef]
- Qi, T. Choukoutienology; Zhejiang University Press and Archaeopress: Hangzhou, China, 2018. [Google Scholar]
- Li, F.; Bae, C.J.; Ramsey, C.B.; Chen, F.; Gao, X. Re-dating Zhoukoudian Upper Cave, northern China and its regional significance. J. Hum. Evol. 2018, 121, 170–177. [Google Scholar] [CrossRef]
- Zhao, Z.; Yuan, G.; Wang, J.; Zhong, Y. On the amino acid composition and microstructure of fossil ostrich eggshell from Sinanthropus site, Choukoutien. Vert. Palasiat. 1981, 19, 327–337. [Google Scholar]
- Janz, L.; Elston, R.G.; Burr, G.S. Dating North Asian surface assemblages with ostrich eggshell: Implications for palaeoecology and extirpation. J. Archaeol. Sci. 2009, 36, 1982–1989. [Google Scholar] [CrossRef]
- Kurochkin, E.N.; Kuzmin, Y.V.; Antoshchenko-Olenev, I.V.; Zabelin, V.I.; Krivonogov, S.K.; Nohrina, T.I.; Lbova, L.V.; Burr, G.S.; Cruz, R.J. The timing of ostrich existence in Central Asia: AMS 14C age of eggshells from Mongolia and southernSiberia (a pilot study). Nucl. Instr. Meth. Phys. Res. B 2010, 268, 1091–1093. [Google Scholar] [CrossRef]
- Khatsenovich, A.M.; Rybin, E.P.; Gunchinsuren, B.; Bolorbat, T.; Odsuren, D.; Angaragdulguun, G.; Margad-Erdene, G. Human and Struthio asiaticus: One page of Paleolithic art in the eastern part of Central Asia. Izv. Irkutsk. gosudarst. Univers. Ser. Geoarkheol. Etnol. Antrop. 2017, 21, 80–106. [Google Scholar]
- Zhu, R.X.; Potts, H.; Xie, F.; Hoffman, K.A.; Deng, C.L.; Shi, C.D.; Pan, Y.X.; Wang, H.Q.; Shi, G.H.; Wu, N.Q. New evidence on the earliest human presence at high northern latitudes in northeast Asia. Nature 2004, 431, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Zhu, R.; Zhang, R.; Ao, H.; Pan, Y. Timing of the Nihewan formation and faunas. Quat. Res. 2008, 69, 77–90. [Google Scholar] [CrossRef]
- Shen, C.; Gao, X.; Wei, Q. The earliest hominin occupations in the Nihewan Basin of Northern China: Recent progress in field investigations. In Asian Paleoanthropology: From Africa to China and Beyond; Norton, C.J., Braun, D.R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 169–180. [Google Scholar]
- Dennell, R.W. The Nihewan Basin of North China in the Early Pleistocene: Continuous and flourishing, or discontinuous, infrequent and ephemeral occupation? Quat. Intern. 2012, 295, 223–236. [Google Scholar] [CrossRef]
- Liu, P.; Deng, C.; Li, S.; Cai, S.; Cheng, H.; Yuan, B.; Wei, Q.; Zhu, R. Magnetostratigraphic dating of the Xiashagou Fauna and implication for sequencing the mammalian faunas in the Nihewan Basin, North China. Palaeogeogr. Palaeoclimat. Palaeoecol. 2012, 315, 75–85. [Google Scholar] [CrossRef]
- Pei, S.; Gao, X.; Wang, H.; Kuman, K.; Bae, C.J.; Chen, F.; Guan, Y.; Zhang, Y.; Zhang, X.; Peng, F.; et al. Early Pleistocene archaeological occurrences at the Feiliang site, and the archaeology of human origins in the Nihewan Basin, North China. PLoS ONE 2017, 12, e0187251. [Google Scholar] [CrossRef]
- Murray, P.F.; Vickers-Rich, P. Magnificent Mihirungs: The Colossal Flightless Birds of the Australian Dreamtime; Indiana University Press: Bloomington/Indianapolis, IN, USA, 2004. [Google Scholar]
- Bonilauri, S.; Boëda, E.; Griggo, C.; Al-Sakhel, H.; Muhesen, S. Un éclat de silex moustérien coincé dans un bassin d’autruche (Struthio camelus) à Umm el Tlel (Syrie centrale). Paléorient 2007, 33, 39–46. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen Number | Species | Age | Geological Location | LCF (mm) | Body Mass Estimated (kg) | Total Length (mm) | Minimum Shaft Width (mm) | Stoutness Index (%) | Source |
|---|---|---|---|---|---|---|---|---|---|
| NIH008 | Pachystruthio indet. | Early Pleistocene | Nihewan Formation (China) | 199 | 300 | 340.0 | 74.0 | 21.76 | Boule et al. 1928; this paper |
| D70 | Pachystruthio dmanisensis | Early Pleistocene | Dmanisi (Georgia) | 220 | 382 | 380.0 | 76.0 | 20.00 | Burchak–Abramovich & Vekua 1990; Vekua 2013 |
| D5768 | Pachystruthio dmanisensis | Early Pleistocene | Dmanisi (Georgia) | 220 | 382 | 385.0 | 76.0 | 19.74 | Burchak–Abramovich & Vekua 1990; Vekua 2013 |
| PIN 5644/56 | Pachystruthio dmanisensis | Early Pleistocene | Taurida Cave (Crimea) | 240 | 472 | 390.0 | 74.7 | 19.15 | Zelenkov et al. 2019 |
| IVPP V6943 | Struthio anderssoni | Late Pleistocene | Zhoukoudian (China) | 190 | 269 | / | / | / | this paper |
| / | Struthio anderssoni | Late Pleistocene | Zhoukoudian (China) | / | / | 355.0 | 69.0 | 19.44 | Shaw 1937 |
| / | Struthio camelus | Modern | / | 140 | 129 | 301.0 | 38.4 | 12.75 | this paper |
| / | Struthio camelus | Modern | / | 140.5 | 130 | 327.0 | 36.9 | 11.29 | this paper |
| 1888.377 | Struthio camelus | Modern | / | 146 | 142 | 300.0 | 49.0 | 16.33 | this paper |
| 1888.377 | Struthio camelus | Modern | / | 147 | 145 | 300.0 | 50.0 | 16.67 | this paper |
| 1889.171 | Struthio camelus | Modern | / | 144 | 138 | 330.0 | 45.0 | 13.64 | this paper |
| 1889.171 | Struthio camelus | Modern | / | 145 | 140 | 307.0 | 40.0 | 13.03 | this paper |
| 1889.31 | Struthio camelus | Modern | / | 135 | 118 | 310.0 | 49.0 | 15.81 | this paper |
| 1889.31 | Struthio camelus | Modern | / | 136 | 120 | 330.0 | 45.0 | 13.64 | this paper |
| 1912.49 | Struthio camelus | Modern | / | 155 | 164 | 315.0 | 52.0 | 16.51 | this paper |
| 1922.105 | Struthio camelus | Modern | / | 142 | 133 | 286.0 | 32.0 | 11.19 | this paper |
| 1922.105 | Struthio camelus | Modern | / | 157 | 170 | 320.0 | 43.0 | 13.44 | this paper |
| 1923.2163 | Struthio camelus | Modern | / | 152 | 157 | 320.0 | 42.0 | 13.13 | this paper |
| 1923.2163 | Struthio camelus | Modern | / | 150 | 152 | 315.0 | 46.0 | 14.60 | this paper |
| 1937.114 | Struthio camelus | Modern | / | 139 | 126 | 325.0 | 45.0 | 13.85 | this paper |
| / | Struthio camelus | Modern | Baku | 156 | 167 | 310.0 | 51.0 | 16.45 | Burchak–Abramovich and Vekua 1990 |
| / | Struthio camelus | Modern | Moscow | 143 | 135 | 314.0 | 46.0 | 14.65 | Burchak–Abramovich and Vekua 1990 |
| / | Struthio camelus | Modern | Krakow | 163 | 186 | 325.0 | 51.0 | 15.69 | Burchak–Abramovich and Vekua 1990 |
| / | Struthio camelus | Modern | Krakow | 161 | 180 | 318.0 | 49.0 | 15.41 | Burchak–Abramovich and Vekua 1990 |
| / | Struthio oldawayi | Early Pleistocene | Olduvai Gorge, Tanzania | / | / | 400.0 | 64.0 | 16.00 | Leakey 1967; Vekua 2013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buffetaut, E.; Angst, D. A Giant Ostrich from the Lower Pleistocene Nihewan Formation of North China, with a Review of the Fossil Ostriches of China. Diversity 2021, 13, 47. https://doi.org/10.3390/d13020047
Buffetaut E, Angst D. A Giant Ostrich from the Lower Pleistocene Nihewan Formation of North China, with a Review of the Fossil Ostriches of China. Diversity. 2021; 13(2):47. https://doi.org/10.3390/d13020047
Chicago/Turabian StyleBuffetaut, Eric, and Delphine Angst. 2021. "A Giant Ostrich from the Lower Pleistocene Nihewan Formation of North China, with a Review of the Fossil Ostriches of China" Diversity 13, no. 2: 47. https://doi.org/10.3390/d13020047
APA StyleBuffetaut, E., & Angst, D. (2021). A Giant Ostrich from the Lower Pleistocene Nihewan Formation of North China, with a Review of the Fossil Ostriches of China. Diversity, 13(2), 47. https://doi.org/10.3390/d13020047