
Contributions of Distribution Modelling to the Ecological Study of Psittaciformes



Abstract
1. Parrots and Their Important Ecological Role
2. Literature Review of Distribution Modelling in Parrot Species
2.1. Sample of Scientific Literature
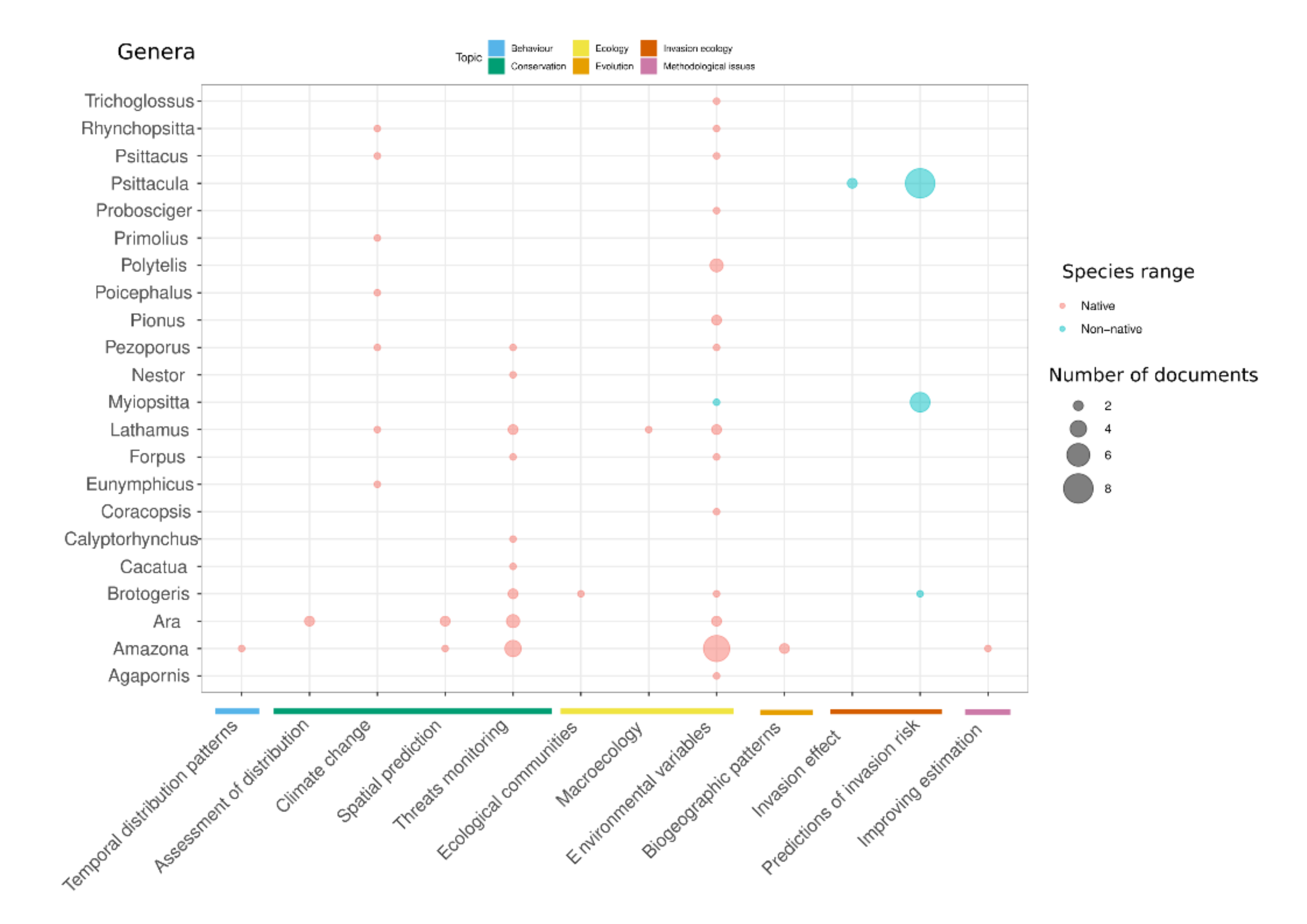
2.2. Document Classification

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Topics | General Application | Specific Issues | Number of Publications | Paradigm Used | Type of Data | Examples |
|---|---|---|---|---|---|---|
| Behavior | Habitat use related to behavior types | Occurrence of behavior types | 1 | ENM | Literature | [20] |
| Temporal distribution patterns | Movement related to environmental factors | 1 | ENM | Open access databases | [21] | |
| Conservation | Climate change | Change in distribution driven by climate change | 6 | ENM, SOM | Field work; Open access databases; Citizen science project; Literature; Museum collections | [22,23,24] |
| Combined effects of climate and habitat changes | 1 | ENM | Literature; Open access databases | [25] | ||
| Evaluating or forecasting the effect of environmental changes | 1 | ENM | Field work | [26] | ||
| Spatial prediction | Effect of conservation actions | 1 | ENM | Field work | [27] | |
| Identification of priority areas for conservation | 1 | ENM | Literature; Open access databases | [28] | ||
| Resource distribution | 2 | ENM | Field work; Open access databases | [29,30] | ||
| Threats monitoring | Change in distribution driven by habitat loss | 7 | ENM, SOM | Field work; Open access databases | [31,32,33,34,35,36] | |
| Effect of conservation actions | 1 | RSF | Field work | [37] | ||
| Fragmentation effect | 2 | ENM | Field work | [38,39] | ||
| Input for population models/population viability analysis | 1 | ENM | Field work | [40] | ||
| Threat distribution | 2 | ENM | Field work | [40,41] | ||
| Threat effect on distribution/occupancy | 1 | ENM | Field work | [42] | ||
| Ecology | Macroecology | Abundance-occupancy relationship | 1 | SOM | Field work | [43] |
| Effect of biotic interactions on distribution | 1 | ENM | Open access databases; Museum collections | [11] | ||
| Global distribution patterns of diet type | 1 | ENM | Open access databases | [44] | ||
| Relation with environmental variables | Determining areas for survey | 2 | ENM | Field work | [20] | |
| Identifying breeding habitat | 5 | ENM, RSF | Field work | [45,46] | ||
| Identifying potential habitat | 1 | ENM | Open access databases | [47] | ||
| Inter-annual variability in distribution | 2 | ENM, RSF | Field work | [48] | ||
| Variables affecting distribution | 8 | ENM, SOM, RSF | Field work; Open access databases | [49,50,51,52,53] | ||
| Ecological communities | Richness and alpha-diversity | 2 | SOM, RSF | Field work; Open access databases | [54,55] | |
| Evolution | Biogeographic patterns | Change in historical distribution | 2 | ENM | Open access databases; Museum collections; Field work; | [56,57] |
| Understanding distribution of extincted species | 1 | ENM | Literature; Museum collections | [58] | ||
| Invasion ecology | Invasion effect | Impacts on native species | 2 | ENM, RSF | Field work; Open access databases | [59,60] |
| Predictions of invasion risk | Establishment of non-native specie | 3 | ENM | Open access databases | [27,61,62] | |
| Limitations into invasion risk | 1 | RSF | Field work | [63] | ||
| Niche shift | 3 | ENM | Field work | [21] | ||
| Potential range of invasive species | 5 | ENM | Literature; Field work; Open access databases | [64,65] | ||
| Methodological issues | Improving estimation | Factors affecting distribution estimation | 1 | SOM | Open access databases; Field work | [66] |
| Survey methodology biases | Evaluating citizen science data | 1 | ENM | Literature; Open access databases; Citizen science; Field work | [65] |
3. Contributions of Species Distribution Models to Parrots Research
3.1. Distribution Models to Study the Ecology of Parrots
3.2. The Biotic Component of Parrots’ Distribution
3.3. Applications of Distribution Models to Conservation Problems
3.4. Applications of Distribution Models to Invasion Ecology
4. Challenges and Opportunities for New Research on Parrots’ Distribution
4.1. Social Behavior and Geographic Variability
4.2. From Global to Local
4.3. Automatic Data Collection
4.4. Citizen-Science and Socially-Derived Data Collection
4.5. Cyber Infrastructure for Research
4.6. Supporting Decision Making
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- IUCN The International Union for Conservation of Nature’s Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 20 May 2021).
- Blanco, G.; Hiraldo, F.; Rojas, A.; Dénes, F.v.; Tella, J.L. Parrots as Key Multilinkers in Ecosystem Structure and Functioning. Ecol. Evol. 2015, 5, 4141–4160. [Google Scholar] [CrossRef] [PubMed]
- Blanco, G.; Bravo, C.; Pacifico, E.C.; Chamorro, D.; Speziale, K.L.; Lambertucci, S.A.; Hiraldo, F.; Tella, J.L. Internal Seed Dispersal by Parrots: An Overview of a Neglected Mutualism. PeerJ 2016, 2016, e1688. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Tabares, D.L.; Cordier, J.M.; Landi, M.A.; Olah, G.; Nori, J. Global Trends of Habitat Destruction and Consequences for Parrot Conservation. Glob. Chang. Biol. 2020, 26, 4251–4262. [Google Scholar] [CrossRef] [PubMed]
- Devarajan, K.; Morelli, T.L.; Tenan, S. Multi-Species Occupancy Models: Review, Roadmap, and Recommendations. Ecography 2020, 43, 1612–1624. [Google Scholar] [CrossRef]
- Berkunsky, I.; Quillfeldt, P.; Brightsmith, D.; Abbud, M.; Aguilar, J.; Alemán-Zelaya, U.; Aramburú, R.; Arce Arias, A.; Balas McNab, R.; Balsby, T.; et al. Current Threats Faced by Neotropical Parrot Populations. Biol. Conserv. 2017, 214, 278–287. [Google Scholar] [CrossRef]
- BirdLife International IUCN Red List for Birds. Available online: http://www.birdlife.org (accessed on 10 October 2019).
- Sánchez-Mercado, A.; Ferrer-Paris, J.R.; Rodríguez, J.P.; Tella, J.L. A Literature Synthesis of Actions to Tackle Illegal Parrot Trade. Diversity 2021, 13, 191. [Google Scholar] [CrossRef]
- Simmonds, J.S.; Watson, J.E.M.; Salazar, A.; Maron, M. A Composite Measure of Habitat Loss for Entire Assemblages of Species. Conserv. Biol. 2019, 33, 1438–1447. [Google Scholar] [CrossRef]
- Carrete, M.; Tella, J.L. Wild-Bird Trade and Exotic Invasions: A New Link of Conservation Concern? Front. Ecol. Environ. 2008, 6, 207–211. [Google Scholar] [CrossRef]
- De Araújo, C.B.; Marcondes-Machado, L.O.; Costa, G.C. The Importance of Biotic Interactions in Species Distribution Models: A Test of the Eltonian Noise Hypothesis Using Parrots. J. Biogeogr. 2014, 41, 513–523. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Engler, J.O.; Stiels, D.; Schidelko, K.; Strubbe, D.; Quillfeldt, P.; Brambilla, M. Avian SDMs: Current State, Challenges, and Opportunities. J. Avian Biol. 2017, 48, 1483–1504. [Google Scholar] [CrossRef]
- Martín-Martín, A.; Orduna-Malea, E.; Delgado López-Cózar, E. Coverage of Highly-Cited Documents in Google Scholar, Web of Science, and Scopus: A Multidisciplinary Comparison. Scientometrics 2018, 116, 2175–2188. [Google Scholar] [CrossRef]
- Kumar, V.S.; Singh, P.; Karmakar, M.; Leta, J.; Mayr, P. The Journal Coverage of Web of Science, Scopus and Dimensions: A Comparative Analysis. Scienciometric 2021, 5113–5142. [Google Scholar] [CrossRef]
- Angulo, E.; Diagne, C.; Ballesteros-Mejia, L.; Adamjy, T.; Ahmed, D.A.; Akulov, E.; Banerjee, A.K.; Capinha, C.; Dia, C.A.K.M.; Dobigny, G.; et al. Non-English Languages Enrich Scientific Knowledge: The Example of Economic Costs of Biological Invasions. Sci. Total Environ. 2021, 775, 144441. [Google Scholar] [CrossRef]
- O’Neil, D. English as the Lingua Franca of International Publishing. World Engl. 2018, 37, 146–165. [Google Scholar] [CrossRef]
- ISO International Organization for Standardization—ISO 3166 Country Codes. Available online: https://www.iso.org/iso-3166-country-codes.html (accessed on 2 January 2021).
- BirdLife International Data Zone. Available online: http://datazone.birdlife.org/species/search (accessed on 3 May 2020).
- Mzumara, T.I.; Martin, R.O.; Tripathi, H.; Phiri, C.; Amar, A. Distribution of a Habitat Specialist: Mopane Woodland Structure Determines Occurrence of near Threatened Lilian’s Lovebird Agapornis Lilianae. Bird Conserv. Int. 2019, 29, 413–422. [Google Scholar] [CrossRef]
- Sánchez-Barradas, A.; Santiago-Jiménez, Q.J.; Rojas-Soto, O. Variación Temporal en la Distribución Geográfica y Ecológica de Amazona Finschi, (Psittaciformes: Psittacidae). Rev. Biol. Trop. 2017, 65, 1194–1207. [Google Scholar] [CrossRef]
- Porfirio, L.L.; Harris, R.M.B.; Stojanovic, D.; Webb, M.H.; Mackey, B. Projected Direct and Indirect Effects of Climate Change on the Swift Parrot, an Endangered Migratory Species. Emu 2016, 116, 273–283. [Google Scholar] [CrossRef]
- Monterrubio-Rico, T.C.; Charre-Medellin, J.F.; Sáenz-Romero, C. Current and Future Habitat Availability for Thick-Billed and Maroon-Fronted Parrots in Northern Mexican Forests. J. Field Ornithol. 2015, 86, 1–16. [Google Scholar] [CrossRef]
- Kalle, R.; Ramesh, T.; Downs, C.T. When and Where to Move: Dynamic Occupancy Models Explain the Range Dynamics of a Food Nomadic Bird under Climate and Land Cover Change. Glob. Chang. Biol. 2018, 24, e27–e39. [Google Scholar] [CrossRef]
- Peers, M.J.L.; Thornton, D.H.; Majchrzak, Y.N.; Bastille-Rousseau, G.; Murray, D.L. De-Extinction Potential under Climate Change: Extensive Mismatch between Historic and Future Habitat Suitability for Three Candidate Birds. Biol. Conserv. 2016, 197, 164–170. [Google Scholar] [CrossRef]
- Legault, A.; Theuerkauf, J.; Chartendrault, V.; Rouys, S.; Saoumoé, M.; Verfaille, L.; Desmoulins, F.; Barré, N.; Gula, R. Using Ecological Niche Models to Infer the Distribution and Population Size of Parakeets in New Caledonia. Biol. Conserv. 2013, 167, 149–160. [Google Scholar] [CrossRef]
- Cobden, M.; Alves, F.; Robinson, S.; Heinsohn, R.; Stojanovic, D. Impact of Removal on Occupancy Patterns of the Invasive Rainbow Lorikeet (Trichoglossus Moluccanus) in Tasmania. Austral Ecol. 2021, 46, 31–38. [Google Scholar] [CrossRef]
- Botero-Delgadillo, E.; Páez, C.A.; Bayly, N. Biogeography and Conservation of Andean and Trans-Andean Populations of Pyrrhura Parakeets in Colombia: Modelling Geographic Distributions to Identify Independent Conservation Units. Bird Conserv. Int. 2012, 22, 445–461. [Google Scholar] [CrossRef]
- Baños-Villalba, A.; Blanco, G.; Díaz-Luque, J.A.; Dénes, F.v.; Hiraldo, F.; Tella, J.L. Seed Dispersal by Macaws Shapes the Landscape of an Amazonian Ecosystem. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Valentine, L.E.; Fisher, R.; Wilson, B.A.; Sonneman, T.; Stock, W.D.; Fleming, P.A.; Hobbs, R.J. Time since Fire Influences Food Resources for an Endangered Species, Carnaby’s Cockatoo, in a Fire-Prone Landscape. Biol. Conserv. 2014, 175, 1–9. [Google Scholar] [CrossRef]
- Marín-Togo, M.C.; Monterrubio-Rico, T.C.; Renton, K.; Rubio-Rocha, Y.; Macías-Caballero, C.; Ortega-Rodríguez, J.M.; Cancino-Murillo, R. Reduced Current Distribution of Psittacidae on the Mexican Pacific Coast: Potential Impacts of Habitat Loss and Capture for Trade. Biodivers. Conserv. 2011, 21, 451–473. [Google Scholar] [CrossRef]
- Monterrubio-Rico, T.C.; Renton, K.; Ortega-Rodrguez, J.M.; Pérez-Arteaga, A.; Cancino-Murillo, R. The Endangered Yellow-Headed Parrot Amazona Oratrix along the Pacific Coast of Mexico. ORYX 2010, 44, 602–609. [Google Scholar] [CrossRef]
- Gibson, L.; Barrett, B.; Burbidge, A. Dealing with Uncertain Absences in Habitat Modelling: A Case Study of a Rare Ground-Dwelling Parrot. Divers. Distrib. 2007, 13, 704–713. [Google Scholar] [CrossRef]
- Cooper, N.W.; Hallworth, M.T.; Marra, P.P. Light-Level Geolocation Reveals Wintering Distribution, Migration Routes, and Primary Stopover Locations of an Endangered Long-Distance Migratory Songbird. J. Avian Ecol. 2017, 48, 1–11. [Google Scholar] [CrossRef]
- Laranjeiras, T.O.; Cohn-Haft, M. Where Is the Symbol of Brazilian Ornithology? The Geographic Distribution of the Golden Parakeet (Guarouba Guarouba - Psittacidae). Rev. Bras. Ornitol. 2009, 17, 1–19. [Google Scholar]
- Ferrer-Paris, J.R.; Sánchez-Mercado, A.; Rodríguez-Clark, K.M.; Rodríguez, J.P.; Rodríguez, G.A. Using Limited Data to Detect Changes in Species Distributions: Insights from Amazon Parrots in Venezuela. Biol. Conserv. 2014, 173, 133–143. [Google Scholar] [CrossRef]
- Recio, M.R.; Payne, K.; Seddon, P.J. Emblematic Forest Dwellers Reintroduced into Cities: Resource Selection by Translocated Juvenile Kaka. Curr. Zool. 2016, 62, 15–22. [Google Scholar] [CrossRef]
- Berkunsky, I.; Simoy, M.v.; Cepeda, R.E.; Marinelli, C.; Kacoliris, F.P.; Daniele, G.; Cortelezzi, A.; Díaz-Luque, J.A.; Friedman, J.M.; Aramburú, R.M. Assessing the Use of Forest Islands by Parrot Species in a Neotropical Savanna. Avian Conserv. Ecol. 2015, 10, 11–15. [Google Scholar] [CrossRef]
- Plasencia-Vázquez, A.H.; Escalona Segura, G. Characterization of the Potential Geographical Distribution Area of Parrot Species in Yucatan Peninsula, Mexico. Rev. Biol. Trop. 2014, 62, 1509–1522. [Google Scholar]
- Stojanovic, D.; Webb, M.H.; Alderman, R.; Porfirio, L.L.; Heinsohn, R. Discovery of a Novel Predator Reveals Extreme but Highly Variable Mortality for an Endangered Migratory Bird. Divers. Distrib. 2014, 20, 1200–1207. [Google Scholar] [CrossRef]
- Allen, M.; Webb, M.H.; Alves, F.; Heinsohn, R.; Stojanovic, D. Occupancy Patterns of the Introduced, Predatory Sugar Glider in Tasmanian Forests. Austral Ecol. 2018, 43, 470–475. [Google Scholar] [CrossRef]
- Bluff, L.A. Ground Parrots and Fire in East Gippsland, Victoria: Habitat Occupancy Modelling from Automated Sound Recordings. Emu 2016, 116, 402–410. [Google Scholar] [CrossRef]
- Webb, M.H.; Heinsohn, R.; Sutherland, W.J.; Stojanovic, D.; Terauds, A. An Empirical and Mechanistic Explanation of Abundance-Occupancy Relationships for a Critically Endangered Nomadic Migrant. Am. Nat. 2018, 193, 59–69. [Google Scholar] [CrossRef]
- Kissling, W.D.; Böhning–Gaese, K.; Jetz, W. The Global Distribution of Frugivory in Birds. Glob. Ecol. Biogeogr. 2009, 18, 150–162. [Google Scholar] [CrossRef]
- Saunders, D.L.; Heinsohn, R. Winter Habitat Use by the Endangered, Migratory Swift Parrot (Lathamus Discolor) in New South Wales. Emu 2008, 108, 81–89. [Google Scholar] [CrossRef]
- Webb, M.H.; Terauds, A.; Tulloch, A.; Bell, P.; Stojanovic, D.; Heinsohn, R. The Importance of Incorporating Functional Habitats into Conservation Planning for Highly Mobile Species in Dynamic Systems. Conserv. Biol. 2017, 31, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Monterrubio-Rico, T.C.; Álvarez-Jara, M.; Téllez-García, L.; Tena-Morelos, C. Nesting Habitat Characterization for Amazona Oratrix (Psittaciformes: Psittacidae) in the Central Pacific, Mexico. Rev. Biol. Trop. 2014, 62, 1053–1072. [Google Scholar] [CrossRef]
- Ortiz-Maciel, S.G.; Hori-Ochoa, C.; Enkerlin-Hoeflich, E. Maroon-Fronted Parrot (Rhynchopsitta Terrisi) Breeding Home Range and Habitat Selection in the Northern Sierra Madre Oriental, Mexico. Wilson J. Ornithol. 2010, 122, 513–517. [Google Scholar] [CrossRef]
- Keighley, M.v.; Langmore, N.E.; Peñalba, J.v.; Heinsohn, R. Modelling Dispersal in a Large Parrot: A Comparison of Landscape Resistance Models with Population Genetics and Vocal Dialect Patterns. Landsc. Ecol. 2020, 35, 129–144. [Google Scholar] [CrossRef]
- Shukuroglou, P.; McCarthy, M.A. Modelling the Occurrence of Rainbow Lorikeets (Trichoglossus Haematodus) in Melbourne. Austral Ecol. 2006, 31, 240–253. [Google Scholar] [CrossRef]
- Rodríguez–Pastor, R.; Senar, J.C.; Ortega, A.; Faus, J.; Uribe, F.; Montalvo, T. Distribution Patterns of Invasive Monk Parakeets (Myiopsitta Monachus) in an Urban Habitat. Anim. Biodivers. Conserv. 2012, 35, 107–117. [Google Scholar] [CrossRef]
- Watson, S.J.; Watson, D.M.; Luck, G.W.; Spooner, P.G. Effects of Landscape Composition and Connectivity on the Distribution of an Endangered Parrot in Agricultural Landscapes. Landsc. Ecol. 2014, 29, 1249–1259. [Google Scholar] [CrossRef]
- Plasencia-Vázquez, A.H.; Escalona-Segura, G.; Esparza-Olguín, L.G. Interaction of Landscape Variables on the Potential Geographical Distribution of Parrots in the Yucatan Peninsula, Mexico. Anim. Biodivers. Conserv. 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Kavanagh, R.P.; Stanton, M.A. Bird Population Recovery 22 Years after Intensive Logging near Eden, New South Wales. Emu 2003, 103, 221–231. [Google Scholar] [CrossRef]
- Koleff, P.; Gaston, K.J. Latitudinal Gradients in Diversity: Real Patterns and Random Models. Ecography 2001, 24, 341–351. [Google Scholar] [CrossRef]
- Monterrubio-Rico, T.C.; Villaseñor-Gómez, L.E.; Marin-Togo, M.C.; López-Cordova, E.A.; Fabian-Turja, B.; Sorani-Dalbon, V. Distribución Historica y Actual Del Loro Cabeza Amarilla (Amazona Oratrix) En La Costa Central Del Pacífico Mexicano: Ventajas y Limitaciones En El Uso de GARP En Especies Bajo Fuerte Presión de Trafico. Ornitol. Neotrop. 2007, 18, 263–276. [Google Scholar]
- Mota-Vargas, C.; Parra-Noguez, K.P.; Rojas-Soto, O. Análisis Del Conocimiento Histórico de la Distribución Geográfica y Ecológica del Loro Frente Blanca, Amazona Albifrons, Con Evidencia de Colonización Reciente. Rev. Mex. Biodivers. 2020, 91, 912708. [Google Scholar] [CrossRef]
- Burgio, K.R.; Carlson, C.J.; Tingley, M.W. Lazarus Ecology: Recovering the Distribution and Migratory Patterns of the Extinct Carolina Parakeet. Ecol. Evol. 2017, 7, 5467–5475. [Google Scholar] [CrossRef]
- Strubbe, D.; Matthysen, E.; Graham, C.H. Assessing the Potential Impact of Invasive Ring-Necked Parakeets Psittacula Krameri on Native Nuthatches Sitta Europeae in Belgium. J. Appl. Ecol. 2010, 47, 549–557. [Google Scholar] [CrossRef]
- Mori, E.; Ancillotto, L.; Menchetti, M.; Strubbe, D. ‘The Early Bird Catches the Nest’: Possible Competition between Scops Owls and Ring-Necked Parakeets. Anim. Conserv. 2017, 20, 463–470. [Google Scholar] [CrossRef]
- Mori, E.; Cardador, L.; Reino, L.; White, R.L.; Hernández-Brito, D.; le Louarn, M.; Mentil, L.; Edelaar, P.; Pârâu, L.G.; Nikolov, B.P.; et al. Lovebirds in the Air: Trade Patterns, Establishment Success and Niche Shifts of Agapornis Parrots within Their Non-Native Range. Biol. Invasions 2020, 22, 421–435. [Google Scholar] [CrossRef]
- Cardador, L.; Carrete, M.; Gallardo, B.; Tella, J.L. Combining Trade Data and Niche Modelling Improves Predictions of the Origin and Distribution of Non-Native European Populations of a Globally Invasive Species. J. Biogeogr. 2016, 43, 967–978. [Google Scholar] [CrossRef]
- Strubbe, D.; Matthysen, E. Invasive Ring-Necked Parakeets Psittacula Krameri in Belgium: Habitat Selection and Impact on Native Birds. Ecography 2007, 30, 578–588. [Google Scholar] [CrossRef]
- Vall-llosera, M.; Woolnough, A.P.; Anderson, D.; Cassey, P. Improved Surveillance for Early Detection of a Potential Invasive Species: The Alien Rose-Ringed Parakeet Psittacula Krameri in Australia. Biol. Invasions 2017, 19, 1273–1284. [Google Scholar] [CrossRef]
- Falcón, W.; Tremblay, R.L. From the Cage to the Wild: Introductions of Psittaciformes to Puerto Rico. PeerJ 2018, 6, e5669. [Google Scholar] [CrossRef]
- Ferrer-Paris, J.R.; Sánchez-Mercado, A. Making Inferences about Non-Detection Observations to Improve Occurrence Predictions in Venezuelan Psittacidae. Bird Conserv. Int. 2020, 30. [Google Scholar] [CrossRef]
- GBIF.org. Psittaciformes Occurrences. Available online: https://www.gbif.org/occurrence/download/0009968-210914110416597 (accessed on 10 September 2021).
- Thuiller, W. BIOMOD—Optimizing Predictions of Species Distributions and Projecting Potential Future Shifts under Global Change. Glob. Chang. Biol. 2003, 9, 1353–1362. [Google Scholar] [CrossRef]
- Boyce, M.S.; Vernier, P.R.; Nielsen, S.E.; Schmiegelow, F.K.A. Evaluating Resource Selection Functions. Ecol. Model. 2002, 157, 281–300. [Google Scholar] [CrossRef]
- Manning, A.D.; Lindenmayer, D.B.; Nix, H.A.; Barry, S.C. A Bioclimatic Analysis for the Highly Mobile Superb Parrot of South-Eastern Australia. Emu 2005, 105, 193–201. [Google Scholar] [CrossRef]
- Manning, A.D.; Lindenmayer, D.B.; Barry, S.C.; Nix, H.A. Multi-Scale Site and Landscape Effects on the Vulnerable Superb Parrot of South-Eastern Australia during the Breeding Season. Landsc. Ecol. 2006, 21, 1119–1133. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Heines, J.E. Occupancy Estimation and Modeling. Inferring Patterns and Dynamics of Species Occurence; Academic Press: London, UK, 2006. [Google Scholar]
- Rodrigues, P.O.; Borges, M.R.; Melo, C. Richness, Composition and Detectability of Psittacidae (Aves) in Three Palm Swamps of the Cerrado Sensu Lato in Central Brazil. Rev. Chil. Hist. Nat. 2012, 85, 171–178. [Google Scholar] [CrossRef][Green Version]
- Renton, K.; Salinas-Melgoza, A.; de Labra-Hernández, M.Á.; de la Parra-Martínez, S.M. Resource Requirements of Parrots: Nest Site Selectivity and Dietary Plasticity of Psittaciformes. J. Ornithol. 2015, 156, 73–90. [Google Scholar] [CrossRef]
- Engeman, R.; Whisson, D.; Quinn, J.; Cano, F.; Quiñones, P.; White, T.H. Monitoring Invasive Mammalian Predator Populations Sharing Habitat with the Critically Endangered Puerto Rican Parrot Amazona Vittata. ORYX 2006, 40, 95–102. [Google Scholar] [CrossRef]
- Berkunsky, I.; Cepeda, R.E.; Marinelli, C.; Simoy, M.V.; Daniele, G.; Kacoliris, F.P.; Díaz Luque, J.A.; Gandoy, F.; Aramburú, R.M.; Gilardi, J.D. Occupancy and Abundance of Large Macaws in the Beni Savannahs, Bolivia. Oryx 2016, 50, 113–120. [Google Scholar] [CrossRef]
- Wintle, B.A.; Walshe, T.v.; Parris, K.M.; McCarthy, M.A. Designing Occupancy Surveys and Interpreting Non-Detection When Observations Are Imperfect. Divers. Distrib. 2011, 18, 417–424. [Google Scholar] [CrossRef]
- Rivera-Ortíz, F.A.; Oyama, K.; Ríos-Muñoz, C.A.; Solórzano, S.; Navarro-Sigüenza, A.G.; del Coro Arizmendi, M. Habitat Characterization and Modeling of the Potential Distribution of the Military Macaw (Ara Militaris) in Mexico. Rev. Mex. Biodivers. 2013, 84, 1200–1215. [Google Scholar] [CrossRef]
- Rios-Muñoz, C.A.; Navarro-Sigüenza, A.G. Efectos Del Cambio de Uso de Suelo En La Disponibilidad Hipotética de Hábitat Para Los Psitácidos de México. Ornitol. Neotrop. 2009, 20, 491–509. [Google Scholar]
- Plasencia Vázquez, A.H.; Escalona Segura, G.; Ferrer Sánchez, Y. The Relationship between Forest Fragmentation and the Potential Geographical Distribution of Psittacids (Psittaciformes: Psittacidae) in the Yucatan Peninsula, Mexico. Rev. Biol. Trop. 2017, 65, 1470. [Google Scholar] [CrossRef]
- Freeman, B.; Sunnarborg, J.; Peterson, A.T. Effects of Climate Change on the Distributional Potential of Three Range-Restricted West African Bird Species. Condor 2019, 121, 1–10. [Google Scholar] [CrossRef]
- Le Louarn, M.; Clergeau, P.; Strubbe, D.; Deschamps-Cottin, M. Dynamic Species Distribution Models Reveal Spatiotemporal Habitat Shifts in Native Range-Expanding versus Non-Native Invasive Birds in an Urban Area. J. Avian Biol. 2018, 49, jav-01527. [Google Scholar] [CrossRef]
- Shivambu, T.C.; Shivambu, N.; Downs, C.T. Impact Assessment of Seven Alien Invasive Bird Species Already Introduced to South Africa. Biol. Invasions 2020, 22, 1829–1847. [Google Scholar] [CrossRef]
- Muñoz, A.R.; Real, R. Assessing the Potential Range Expansion of the Exotic Monk Parakeet in Spain. Divers. Distrib. 2006, 12, 656–665. [Google Scholar] [CrossRef]
- Strubbe, D.; Matthysen, E. Predicting the Potential Distribution of Invasive Ring-Necked Parakeets Psittacula Krameri in Northern Belgium Using an Ecological Niche Modelling Approach. Biol. Invasions 2009, 11, 497–513. [Google Scholar] [CrossRef]
- Salinas-Melgoza, A.; Salinas-Melgoza, V.; Wright, T.F. Behavioral Plasticity of a Threatened Parrot in Human-Modified Landscapes. Biol. Conserv. 2013, 159, 303–312. [Google Scholar] [CrossRef]
- Montoya, D.; Purves, D.W.; Urbieta, I.R.; Zavala, M.A. Do Species Distribution Models Explain Spatial Structure within Tree Species Ranges? Glob. Ecol. Biogeogr. 2009, 18, 662–673. [Google Scholar] [CrossRef]
- Robinson, L.M.; Elith, J.; Hobday, A.J.; Pearson, R.G.; Kendall, B.E.; Possingham, H.P.; Richardson, A.J. Pushing the Limits in Marine Species Distribution Modelling: Lessons from the Land Present Challenges and Opportunities. Glob. Ecol. Biogeogr. 2011, 20, 789–802. [Google Scholar] [CrossRef]
- Ranasinghe, R. On the Need for a New Generation of Coastal Change Models for the 21st Century. Sci. Rep. 2020, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Morato, R.G.; Stabach, J.A.; Fleming, C.H.; Calabrese, J.M.; Paula, C.d.; Ferraz, M.P.M.; Kantek, D.L.Z.; Miyazaki, S.S.; Pereira, T.D.C.; Araujo, G.R.; et al. Space Use and Movement of a Neotropical Top Predator: The Endangered Jaguar. PLoS ONE 2016, 11, e0168176. [Google Scholar] [CrossRef] [PubMed]
- Caravaggi, A.; Banks, P.B.; Burton, A.C.; Finlay, C.M.V.; Haswell, P.M.; Hayward, M.W.; Rowcliffe, M.J.; Wood, M.D. A Review of Camera Trapping for Conservation Behaviour Research. Remote Sens. Ecol. Conserv. 2017, 3, 109–122. [Google Scholar] [CrossRef]
- Sugai, L.S.M.; Silva, T.S.F.; Ribeiro, J.W.; Llusia, D. Terrestrial Passive Acoustic Monitoring: Review and Perspectives. BioScience 2019, 69, 5–11. [Google Scholar] [CrossRef]
- Shonfield, J.; Bayne, E.M. Autonomous Recording Units in Avian Ecological Research: Current Use and Future Applications. Avian Conserv. Ecol. 2017, 12, 14. [Google Scholar] [CrossRef]
- Wäldchen, J.; Mäder, P. Machine Learning for Image Based Species Identification. Methods Ecol. Evol. 2018, 9, 2216–2225. [Google Scholar] [CrossRef]
- Farley, S.S.; Dawson, A.; Goring, S.J.; Williams, J.W. Situating Ecology as a Big-Data Science: Current Advances, Challenges, and Solutions. BioScience 2018, 68, 563–576. [Google Scholar] [CrossRef]
- Thomson, R.; Potgieter, G.C.; Bahaa-el-din, L. Closing the Gap between Camera Trap Software Development and the User Community. Afr. J. Ecol. 2018, 56, 721–739. [Google Scholar] [CrossRef]
- Lee, K.A.; Lee, J.R.; Bell, P. A Review of Citizen Science within the Earth Sciences: Potential Benefits and Obstacles. Proc. Geol. Assoc. 2020, 131, 605–617. [Google Scholar] [CrossRef]
- Sullivan, B.L.; Wood, C.L.; Iliff, M.J.; Bonney, R.E.; Fink, D.; Kelling, S. EBird: A Citizen-Based Bird Observation Network in the Biological Sciences. Biol. Conserv. 2009, 142, 2282–2292. [Google Scholar] [CrossRef]
- iNaturalist INaturalist. Available online: https://www.inaturalist.org/ (accessed on 10 November 2021).
- Callaghan, C.T.; Rowley, J.J.L.; Cornwell, W.K.; Poore, A.G.B.; Major, R.E. Improving Big Citizen Science Data: Moving beyond Haphazard Sampling. PLoS Biol. 2019, 17, 1–11. [Google Scholar] [CrossRef]
- Ellwood, E.R.; Crimmins, T.M.; Miller-Rushing, A.J. Citizen Science and Conservation: Recommendations for a Rapidly Moving Field. Biol. Conserv. 2017, 208, 1–4. [Google Scholar] [CrossRef]
- Sánchez-Mercado, A.; Asmüssen, M.; Rodríguez-Clark, K.M.; Rodríguez, J.P.; Jedrzejewski, W. Using Spatial Patterns in Illegal Wildlife Uses to Reveal Connections between Subsistence Hunting and Trade. Conserv. Biol. 2016, 30, 1222–1232. [Google Scholar] [CrossRef]
- Sánchez-Mercado, A.; Cardozo-Urdaneta, A.; Moran, L.; Ovalle, L.; Arvelo, M.; Morales-Campo, J.; Coyle, B.; Braun, M.J.; Rodriguez-Clark, K.M. Social Network Analysis Reveals Specialized Trade in an Endangered Songbird. Anim. Conserv. 2019, 23, 132–144. [Google Scholar] [CrossRef]
- Hallgren, W.; Beaumont, L.; Bowness, A.; Chambers, L.; Graham, E.; Holewa, H.; Laffan, S.; Mackey, B.; Nix, H.; Price, J.; et al. The Biodiversity and Climate Change Virtual Laboratory: Where Ecology Meets Big Data. Environ. Model. Softw. 2016, 76, 182–186. [Google Scholar] [CrossRef]
- Velásquez-Tibatá, J.; Olaya-Rodríguez, M.H.; López-Lozano, D.; Gutiérrez, C.; González, I.; Londoño-Murcia, M.C. Biomodelos: A Collaborative Online System to Map Species Distributions. PLoS ONE 2019, 14, e0214522. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; Mcdonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting Species Distributions for Conservation Decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Monterrubio-Rico, T.C.; Charre-Medellín, J.F.; Pacheco-Figueroa, C.; Arriaga-Weiss, S.; de Dios Valdez-Leal, J.; Cancino-Murillo, R.; Escalona-Segura, G.; Bonilla-Ruz, C.; Rubio-Rocha, Y. Distribución Potencial Histórica y Contemporánea de La Familia Psittacidae En México. Rev. Mex. Biodivers. 2016, 87, 1103–1117. [Google Scholar] [CrossRef]
- Marini, M.Â.; Barbet-Massin, M.; Martinez, J.; Prestes, N.P.; Jiguet, F. Applying Ecological Niche Modelling to Plan Conservation Actions for the Red-Spectacled Amazon (Amazona Pretrei). Biol. Conserv. 2010, 143, 102–112. [Google Scholar] [CrossRef]
- Pidgeon, A.M.; Rivera, L.; Martinuzzi, S.; Politi, N.; Bateman, B. Will Representation Targets Based on Area Protect Critical Resources for the Conservation of the Tucuman Parrot? Condor 2015, 117, 503–517. [Google Scholar] [CrossRef]
- Hambuckers, A.; de Harenne, S.; Ledezma, E.R.; Zeballos, L.Z.; François, L. Predicting the Future Distribution of Ara Rubrogenys, an Endemic Endangered Bird Species of the Andes, Taking into Account Trophic Interactions. Diversity 2021, 13, 94. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrer-Paris, J.R.; Sánchez-Mercado, A. Contributions of Distribution Modelling to the Ecological Study of Psittaciformes. Diversity 2021, 13, 611. https://doi.org/10.3390/d13120611
Ferrer-Paris JR, Sánchez-Mercado A. Contributions of Distribution Modelling to the Ecological Study of Psittaciformes. Diversity. 2021; 13(12):611. https://doi.org/10.3390/d13120611
Chicago/Turabian StyleFerrer-Paris, José R., and Ada Sánchez-Mercado. 2021. "Contributions of Distribution Modelling to the Ecological Study of Psittaciformes" Diversity 13, no. 12: 611. https://doi.org/10.3390/d13120611
APA StyleFerrer-Paris, J. R., & Sánchez-Mercado, A. (2021). Contributions of Distribution Modelling to the Ecological Study of Psittaciformes. Diversity, 13(12), 611. https://doi.org/10.3390/d13120611