
Genetic Diversity of a Rising Invasive Pest in the Native Range: Population Genetic Structure of Aromia bungii (Coleoptera: Cerambycidae) in South Korea


Abstract
1. Introduction
2. Materials and Methods
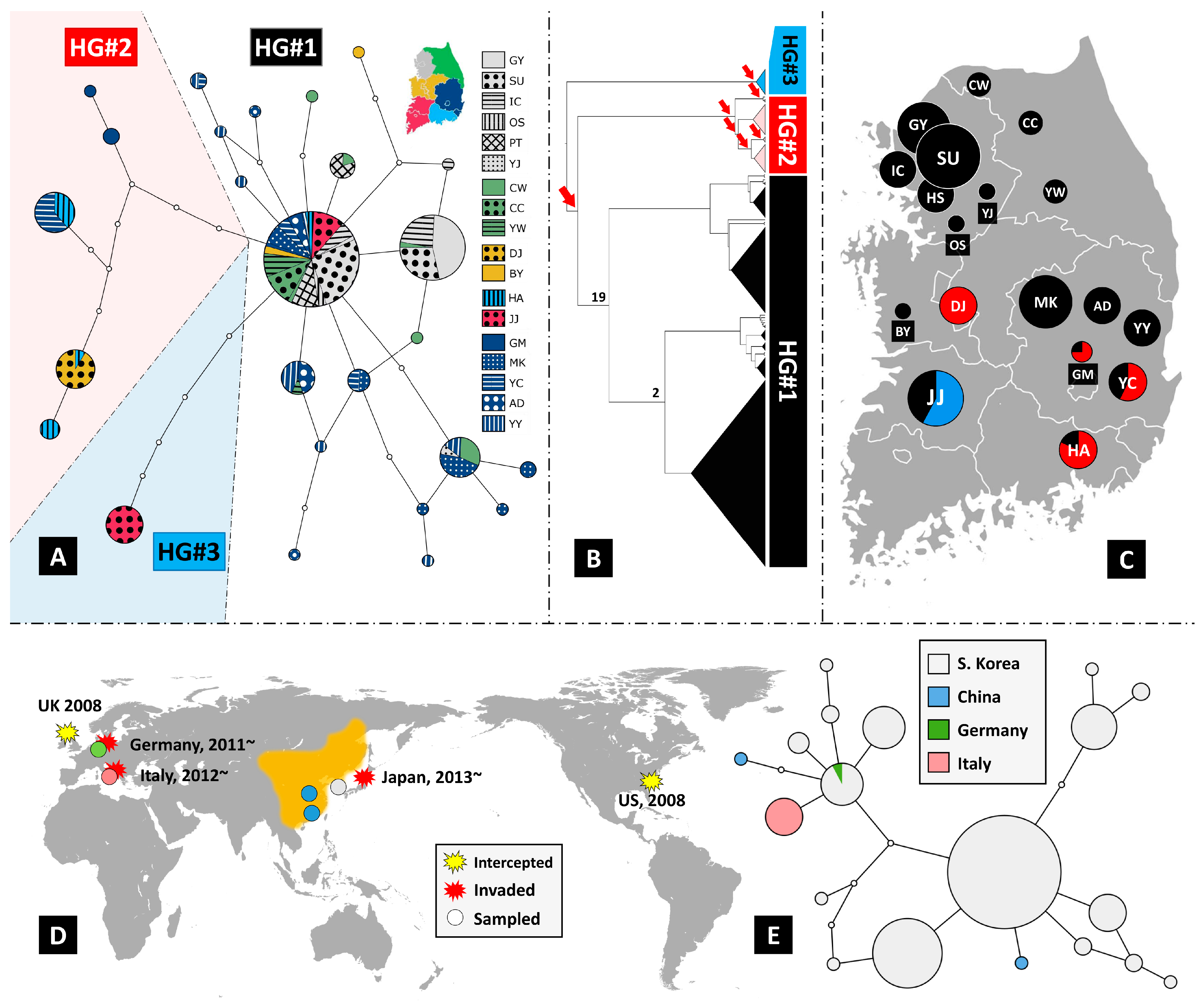
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization. Glossary of Phytosanitary Terms; International Plant Protection Convention (IPPC): Rome, Italy, 2019; 35p. [Google Scholar]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2010; Volume 12. [Google Scholar]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Entomol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Li, Y.; Johnson, A.J.; Gao, L.; Wu, C.; Hulcr, J. Two new invasive Ips bark beetles (Coleoptera: Curculionidae) in mainland China and their potential distribution in Asia. Pest Manag. Sci. 2021, 77, 4000–4008. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.; Nugnes, F.; Vicinanza, F.; Garonna, A.P.; Bernardo, U. Biological and molecular characterization of Aromia bungii (Faldermann, 1835) (Coleoptera: Cerambycidae), an emerging pest of stone fruits in Europe. Sci. Rep. 2020, 10, 7112. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.K.; Lee, S.H.; Choi, W. Cerambycidae of Korea; Geobook: Seoul, Korea, 2015; 400p. [Google Scholar]
- Hwang, S.H. Long-horned beetles in Korea. In Checklist of Organisms in Korea 14; Nature &Ecology: Seoul, Korea, 2015; 551p. [Google Scholar]
- CABI. Invasive Species Compendium; CAB International: Wallingford, UK, 2021; Available online: www.cabi.org/isc (accessed on 1 August 2021).
- Guérin-Méneville, F.E. Iconographie du règne animal de G. Cuvier: ou, Représentation d’après nature de l’une des espèces les plus et souvent non encore figurées de chaque genre d’animaux; J. B. Baillière: Paris, France, 1844; Volume 7, 576p. [Google Scholar] [CrossRef]
- Bates, H.W. On a collection of Coleoptera from Korea (Tribe Geodephaga, Lamellicornia, and Longicornia), made by Mr. J. H. Leech, F.Z.S. In Proceedings of the Scientific Meetings of the Zoological Society of London; Blackwell Publishing Ltd.: Oxford, UK, 1888; Volume 25–26, pp. 378–380. [Google Scholar]
- Faldermann, F. Coleopterorum ab illustrissimo Bungio in China boreali, Mongolia et montibus Altaicis collectorum, nec non ab illustrissimo Turczaninoffio et Stschukino e provinci Irkutzk missorum illustrationes; Caesareae Academiae Scientiarum Impress: Petrópolis, Brazil, 1835; Volume 2, pp. 337–464. [Google Scholar]
- Yu, C.; Nara, H.; Chu, Y. The Longicorn beetles of Taiwan; Muh Sheng Muh Sheng Museum of Entomology: Taipei, Taiwan, 2002; 151p. [Google Scholar]
- EPPO. EPPO Global Database. 2021. Available online: https://gd.eppo.int (accessed on 1 August 2021).
- Burmeister, E.G.; Hendrich, L.; Balke, M. Der asiatische moschusbock Aromia bungii (Faldermann, 1835)—Erstfund für Deutschland (Coleoptera: Cerambycidae). Nachr. Bl. Bay. Ent. 2012, 61, 29–31. [Google Scholar]
- Yasui, H.; Fujiwara-Tsujii, N.; Yasuda, T.; Fukaya, M.; Kiriyama, S.; Nakano, A.; Watanabe, T.; Mori, K. Electroantennographic responses and field attraction of an emerging invader, the red-necked longicorn beetle Aromia bungii (Coleoptera: Cerambycidae), to the chiral and racemic forms of its male-produced aggregation-sex pheromone. Appl. Entomol. Zool. 2019, 54, 109–114. [Google Scholar] [CrossRef]
- Kiriyama, S.; Iwata, R.; Kagaya, E. Newly discovered populations of Aromia bungii (Faldermann), an invasive cerambycid infesting cherry and Japanese apricot trees in Tatebayashi, Gunma Pref. and Fussa, Tokyo Pref. Plant. Prot. 2015, 69, 807–809. [Google Scholar]
- Lee, S.; Lee, Y.; Lee, S. Population genetic structure of Anoplophora glabripennis in South Korea: Invasive populations in the native range? J. Pest Sci. 2020, 93, 1181–1196. [Google Scholar] [CrossRef]
- Hendrich, L.; Morinière, J.; Haszprunar, G.; Hebert, P.D.; Hausmann, A.; Köhler, F.; Balke, M. A comprehensive DNA barcode database for Central European beetles with a focus on Germany: Adding more than 3500 identified species to BOLD. Mol. Ecol. Resour. 2015, 15, 795–818. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 7, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Múrias dos Santos, A.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2015, 32, 627–628. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, J.; Kim, H.; Jung, J. Comparison of Population Genetic Structure of Two Seashore-Dwelling Animal Species, Periwinkle Littorina brevicula and Acorn Barnacle Fistulobalanus albicostatus from Korea. Anim. Syst. Evol. Divers. 2016, 32, 105–111. [Google Scholar] [CrossRef][Green Version]
- Phillips, J.D.; Gillis, D.J.; Hanner, R.H. Incomplete estimates of genetic diversity within species: Implications for DNA barcoding. Ecol. Evol. 2019, 9, 2996–3010. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Manzoor, A.; Wang, X. Mitochondrial DNA analysis reveals spatial genetic structure and high genetic diversity of Massicus raddei (Blessig)(Coleoptera: Cerambycidae) in China. Ecol. Evol. 2020, 10, 11657–11670. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Locality | Abb | Prov | GPS-N | GPS-E | No. | Coll. | Genbank Accession | |
|---|---|---|---|---|---|---|---|---|
| 1 | Goyang | GY | GG | 37.65 | 126.89 | 17 | 2019.08.08 | OK428971~OK428987 |
| 2 | Hwaseong | PT | GG | 37.16 | 126.90 | 11 | 2020.07.15 | OK429054~OK429064 |
| 3 | Incheon | IC | GG | 37.43 | 126.71 | 13 | 2020.07.08 | OK428999~OK429011 |
| 4 | Osan | OS | GG | 37.14 | 127.08 | 2 | 2020.07.15 | OK429052~OK429053 |
| 5 | Seoul-NS | SU | GG | 37.55 | 126.98 | 4 | 2020.07.09 | OK429048~OK429051 |
| 6 | Seoul-YED | SU | GG | 37.53 | 126.91 | 14 | 2020.07.09 | OK429091~OK429104 |
| 7 | Seoul-YED | SU | GG | 37.53 | 126.91 | 12 | 2019.09.23 | OK429065~OK429076 |
| 8 | Yeoju | YJ | GG | 37.24 | 127.65 | 1 (1 E) | 2020.07.15 | OK429105 |
| 9 | Buyeo | BY | CN | 36.28 | 126.91 | 3 (2 L) | 2020.07.10 | OK428936~OK428938 |
| 10 | Daejeon | DJ | CN | 36.32 | 127.43 | 11(1 L) | 2020.07.08 | OK428956~OK428966 |
| 11 | Cheorwon | CW | GW | 38.14 | 127.30 | 9 | 2020.07.07 | OK428947~OK428955 |
| 12 | Chuncheon | CC | GW | 37.92 | 127.78 | 8 | 2020.07.21 | OK428939~OK428946 |
| 13 | Yeongwol | YW | GW | 37.18 | 128.47 | 7 | 2020.07.05–08 | OK429106~OK429112 |
| 14 | Jeonju-BJW | JJ | JB | 35.85 | 127.14 | 9 | 2020.07.22 | OK429012~OK429020 |
| 15 | Jeonju-YYE | JJ | JB | 35.84 | 127.14 | 10 | 2020.07.21 | OK429021~OK429030 |
| 16 | Andong | AD | GB | 36.57 | 128.76 | 10 | 2020.07.05–08 | OK428926~OK428935 |
| 17 | Gumi | GM | GB | 36.10 | 128.39 | 4 | 2020.08.20 | OK428967~OK428970 |
| 18 | Mungyeong | MK | GB | 36.59 | 128.19 | 17 | 2020.07.21 | OK429031~OK429047 |
| 19 | Yeongcheon | YC | GB | 35.97 | 128.91 | 14 | 2020.08.20 | OK429077~OK429090 |
| 20 | Yeongyang | YY | GB | 36.66 | 129.12 | 12 | 2020.08.20 | OK429113~OK429124 |
| 21 | Haman | HA | GN | 35.36 | 128.48 | 11 (10 L) | 2019.09.26 | OK428988~OK428998 |
| Locality | Abbreviation | N | Nh | Gd | Nd | TD | TDp | |
|---|---|---|---|---|---|---|---|---|
| 1 | Goyang | GY | 17 | 2 | 0.1176 ± 0.1012 | 0.000094 ± 0.000175 | −1.164 | 0.140 |
| 2 | Hwaseong | PT | 11 | 2 | 0.5091 ± 0.1008 | 0.000408 ± 0.000420 | 1.186 | 0.879 |
| 3 | Incheon | IC | 13 | 3 | 0.5641 ± 0.1117 | 0.000493 ± 0.000467 | −0.127 | 0.420 |
| 4 | Osan | OS | 2 | 1 | 0.0000 ± 0.0000 | 0.000000 ± 0.000000 | 0.000 | 1.000 |
| 5 | Seoul | SU | 30 | 2 | 0.4598 ± 0.0612 | 0.000368 ± 0.000371 | 1.280 | 0.910 |
| 6 | Yeoju | YJ | 1 | 1 | 1.0000 ± 0.0000 | 0.000000 ± 0.000000 | 0.000 | 1.000 |
| 7 | Buyeo | BY | 3 | 2 | 0.6667 ± 0.3143 | 0.001603 ± 0.001511 | 0.000 | 0.894 |
| 8 | Daejeon | DJ | 11 | 1 | 0.0000 ± 0.0000 | 0.000000 ± 0.000000 | 0.000 | 1.000 |
| 9 | Cheorwon | CW | 9 | 6 | 0.8333 ± 0.1265 | 0.001914 ± 0.001300 | −0.323 | 0.402 |
| 10 | Chuncheon | CC | 8 | 1 | 0.0000 ± 0.0000 | 0.000000 ± 0.000000 | 0.000 | 1.000 |
| 11 | Yeongwol | YW | 7 | 2 | 0.2857 ± 0.1964 | 0.000229 ± 0.000313 | −1.006 | 0.254 |
| 12 | Jeonju | JJ | 19 | 2 | 0.5146 ± 0.0517 | 0.002062 ± 0.001292 | 2.484 | 0.999 |
| 13 | Andong | AD | 10 | 4 | 0.7333 ± 0.1005 | 0.001211 ± 0.000897 | −0.582 | 0.310 |
| 14 | Gumi | GM | 4 | 3 | 0.8333 ± 0.2224 | 0.002003 ± 0.001613 | −0.797 | 0.182 |
| 15 | Mungyeong | MK | 17 | 6 | 0.8015 ± 0.0646 | 0.001343 ± 0.000927 | 0.431 | 0.697 |
| 16 | Yeongcheon | YC | 14 | 4 | 0.6484 ± 0.1163 | 0.003513 ± 0.002072 | 1.055 | 0.880 |
| 17 | Yeongyang | YY | 12 | 8 | 0.8939 ± 0.0777 | 0.001967 ± 0.001287 | 0.898 | 0.817 |
| 18 | Haman | HA | 11 | 4 | 0.7455 ± 0.0978 | 0.003351 ± 0.002034 | 0.957 | 0.855 |
| AD | BY | CC | CW | DJ | GM | GY | HA | IC | JJ | MK | SU | OS | PT | YC | YY | YW | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AD | |||||||||||||||||
| BY | 0.161 | ||||||||||||||||
| CC | 0.209 * | 0.342 | |||||||||||||||
| CW | 0.234 * | 0.119 | 0.212 | ||||||||||||||
| DJ | 0.908 * | 0.952 * | 1 * | 0.873 * | |||||||||||||
| GM | 0.541 * | 0.391 | 0.743 * | 0.495 * | 0.891 * | ||||||||||||
| GY | 0.651 * | 0.779 * | 0.915 * | 0.503 * | 0.991 * | 0.863 * | |||||||||||
| HA | 0.577 * | 0.446 | 0.609 * | 0.551 * | 0.378 * | 0.354 | 0.743 * | ||||||||||
| IC | 0.418 * | 0.425 * | 0.534 * | 0.292 * | 0.957 * | 0.726 * | 0.121 | 0.663 * | |||||||||
| JJ | 0.451 * | 0.377 * | 0.449 * | 0.438 * | 0.817 * | 0.584 * | 0.637 * | 0.594 * | 0.533 * | ||||||||
| MK | 0.357 * | 0.266 | 0.352 * | −0.007 | 0.878 * | 0.59 * | 0.637 * | 0.628 * | 0.481 * | 0.516 * | |||||||
| SU | 0.351 * | 0.379 | 0.181 | 0.31 * | 0.954 * | 0.782 * | 0.511 * | 0.745 * | 0.175 * | 0.577 * | 0.476 * | ||||||
| OS | −0.069 | −0.2 | 0 | −0.079 | 1 * | 0.462 | 0.885 | 0.439 | 0.361 * | 0.291 | 0.16 | −0.017 | |||||
| PT | 0.267 * | 0.287 | 0.25 | 0.233 * | 0.964 * | 0.702 * | 0.788 * | 0.613 * | 0.501 * | 0.477 * | 0.389 * | 0.312 * | −0.01 | ||||
| YC | 0.398 * | 0.263 | 0.417 * | 0.397 * | 0.579 * | 0.255 | 0.602 * | 0.077 | 0.504 * | 0.484 * | 0.482 * | 0.583 * | 0.221 | 0.429 * | |||
| YY | 0.036 | 0.137 | 0.204 * | 0.116 * | 0.852 * | 0.489 * | 0.558* | 0.571 * | 0.378 * | 0.447 * | 0.198 * | 0.371 * | −0.054 | 0.261 * | 0.407 * | ||
| YW | 0.067 | 0.205 | 0.02 | 0.179 * | 0.985 * | 0.676 * | 0.845 * | 0.583 * | 0.471 * | 0.429 * | 0.327 * | 0.179 | −0.313 | 0.195 | 0.392 * | 0.119 | |
| YJ | 0.496 | 0.333 | 1 | −0.344 | 1 | 0.524 | 0.96 | 0.477 | 0.778 | 0.474 | −0.425 | 0.803 | 1 | 0.785 | 0.31 | 0.08 | 0.867 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Cha, D.; Nam, Y.; Jung, J. Genetic Diversity of a Rising Invasive Pest in the Native Range: Population Genetic Structure of Aromia bungii (Coleoptera: Cerambycidae) in South Korea. Diversity 2021, 13, 582. https://doi.org/10.3390/d13110582
Lee S, Cha D, Nam Y, Jung J. Genetic Diversity of a Rising Invasive Pest in the Native Range: Population Genetic Structure of Aromia bungii (Coleoptera: Cerambycidae) in South Korea. Diversity. 2021; 13(11):582. https://doi.org/10.3390/d13110582
Chicago/Turabian StyleLee, Seunghyun, Deokjea Cha, Yongwoo Nam, and Jonkook Jung. 2021. "Genetic Diversity of a Rising Invasive Pest in the Native Range: Population Genetic Structure of Aromia bungii (Coleoptera: Cerambycidae) in South Korea" Diversity 13, no. 11: 582. https://doi.org/10.3390/d13110582
APA StyleLee, S., Cha, D., Nam, Y., & Jung, J. (2021). Genetic Diversity of a Rising Invasive Pest in the Native Range: Population Genetic Structure of Aromia bungii (Coleoptera: Cerambycidae) in South Korea. Diversity, 13(11), 582. https://doi.org/10.3390/d13110582