
Systematics and Biogeography of the New World Genus Plumolepilius (Coleoptera: Curculionidae)



Abstract
:1. Introduction
2. Materials and Methods
- Apex of elytra shape: 0. rounded, 1. straight.
- Femora with single spine on inner margin: 0. absent, 1. present.
- Lateral expansions of pronotum contiguous with humeri: 0. absent, 1. present.
- Pronotum: 0. subquadrate (width: length 1:0.8), 1. transverse (width: length 1:0.65).
- Prosternal channel with plumose scales: 0. absent, 1. present.
- Shape of elytral declivity: 0. rounded, 1. angulate.
- Shape of elytral humeri in males: 0. rounded, 1. prominent.
- Shape of elytral humeri in females: 0. rounded, 1. acutangulate.
- Elytral tubercles: 0. absent, 1. present.
- Shape of tubercle on interstice 5 at elytral declivity: 0. absent, 1. moderately developed, 2. well–developed.
- Shape of interstice 4 at elytral declivity: 0. rounded, 1. angulate, 2. small tubercle present, 3. large tubercle present.
- Elytral tubercle in females: 0. absent, 1. several small, 2. two large tubercles on 4th interstice, 3. two large tubercles on 4th interstice but convergent.
- Elytral striae: 0. well-defined, 1. seriate–punctate.
- Shape of apex of ventrite 5: 0. flat, 1. with two protuberances, 2. depressed.
- Number of distinguishable genital sclerites: 0. two, 1. one.
- Shape of genital sclerites: 0. symmetric, 1. asymmetric.
- Shape of posterior genital sclerite: 0. hook-like, 1. flagellate, 2. sting-like, 3. enlarged-straight, 4. single plate, 5. not distinguishable, 6. karabiner-like.
- Shape of anterior genital sclerite: 0. hook-like 1. capsule, 2. sting-like, 3. enlarged-straight, 4. single plate, 5. folded plate, 6. flagellate, 7. absent.
- Length of temones: 0. subequal to pedon, 1. between 1.5 and 2.0X length of pedon, 2. between 2.0 and 2.9X length of pedon, 3. more than 3.0X length of pedon.
- Shape of pedon: 0. rectangular, 1. other shapes.
- Bifurcation distance of spiculum ventrale: 0. first quarter, 1. half.
- Shape of border of female tergite VII: 0. straight, 1. sinuated.
- Spermatheca shape: 0. wide (twice width of gonocoxites), 1. thin (as wide as gonocoxites).
- Appressed scales at mesanepisternum and mesepimeron: 0. present, 1. absent.
- Erect scales at elytra: 0. absent, 1. present.
- Elytral scales shape: 0. rounded, 1. slightly elongate, 2. clavate, 3. lanceolate.
- Angle of elytral scales: 0. appressed, 1. almost erect, 2. suberect.
- Density of erect or suberect scales: 0. absent, 1. sparse, 2. dense.
- Tuft of setae on apex of abdominal ventrite 5: 0. absent, 1. present.
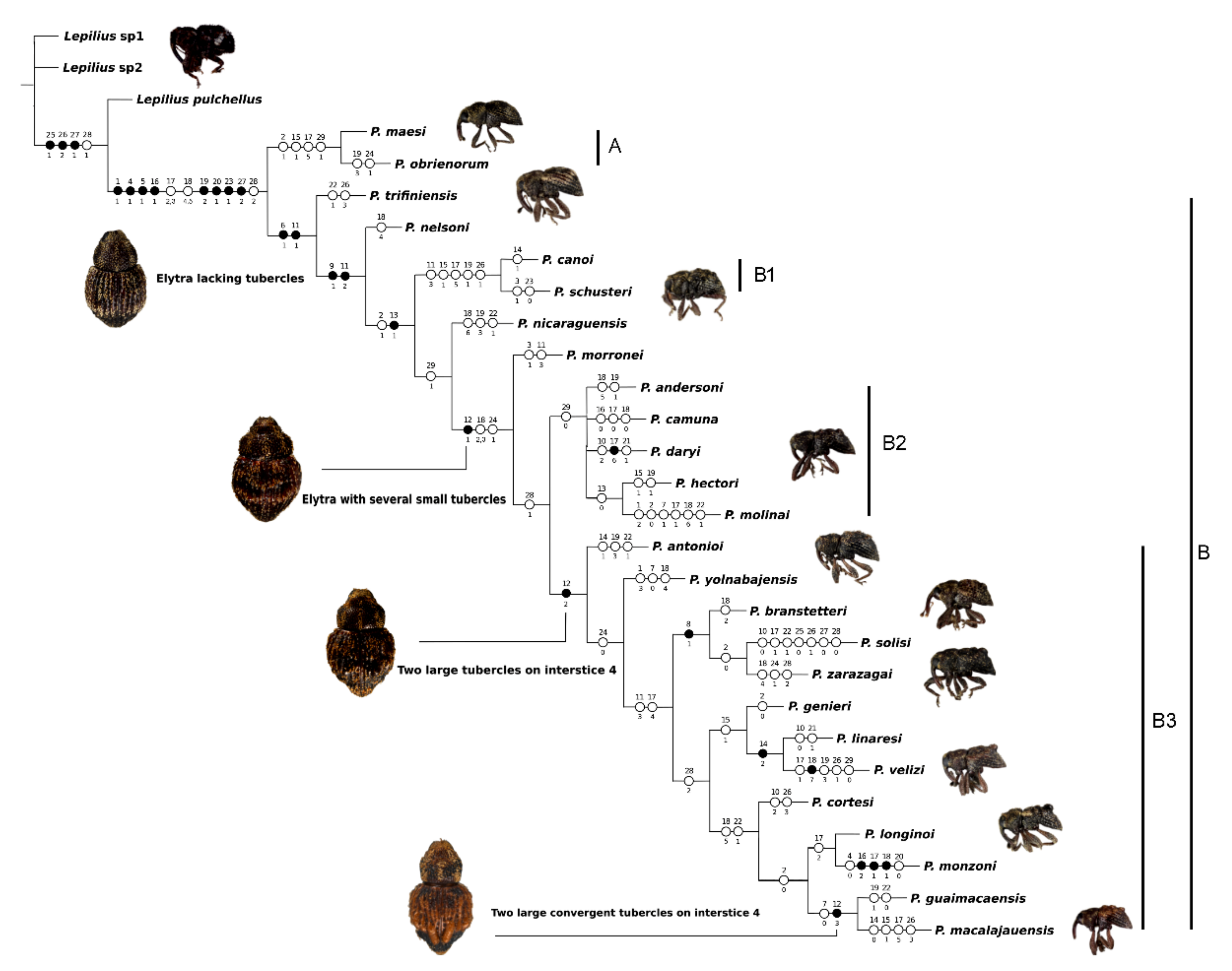
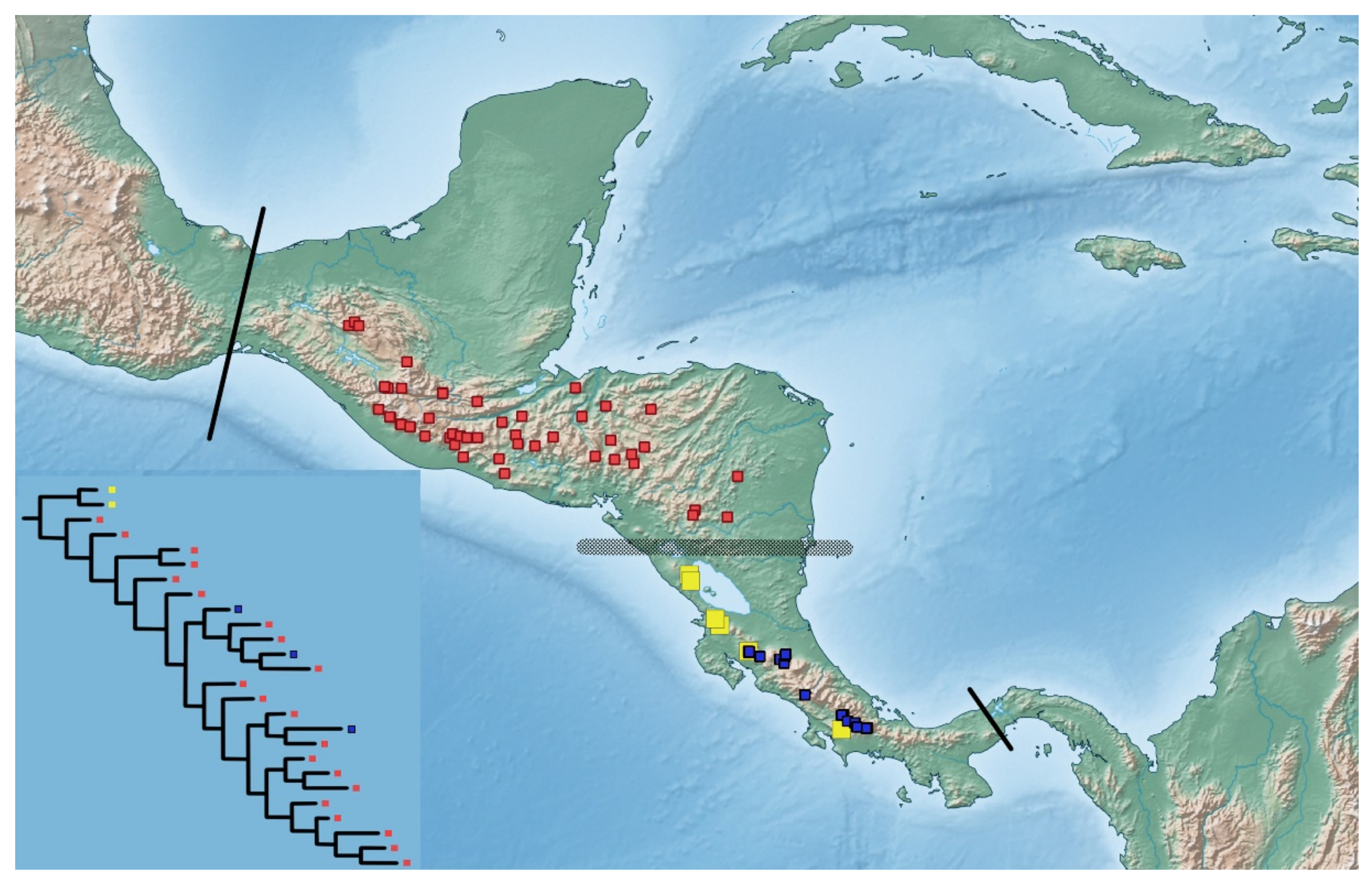
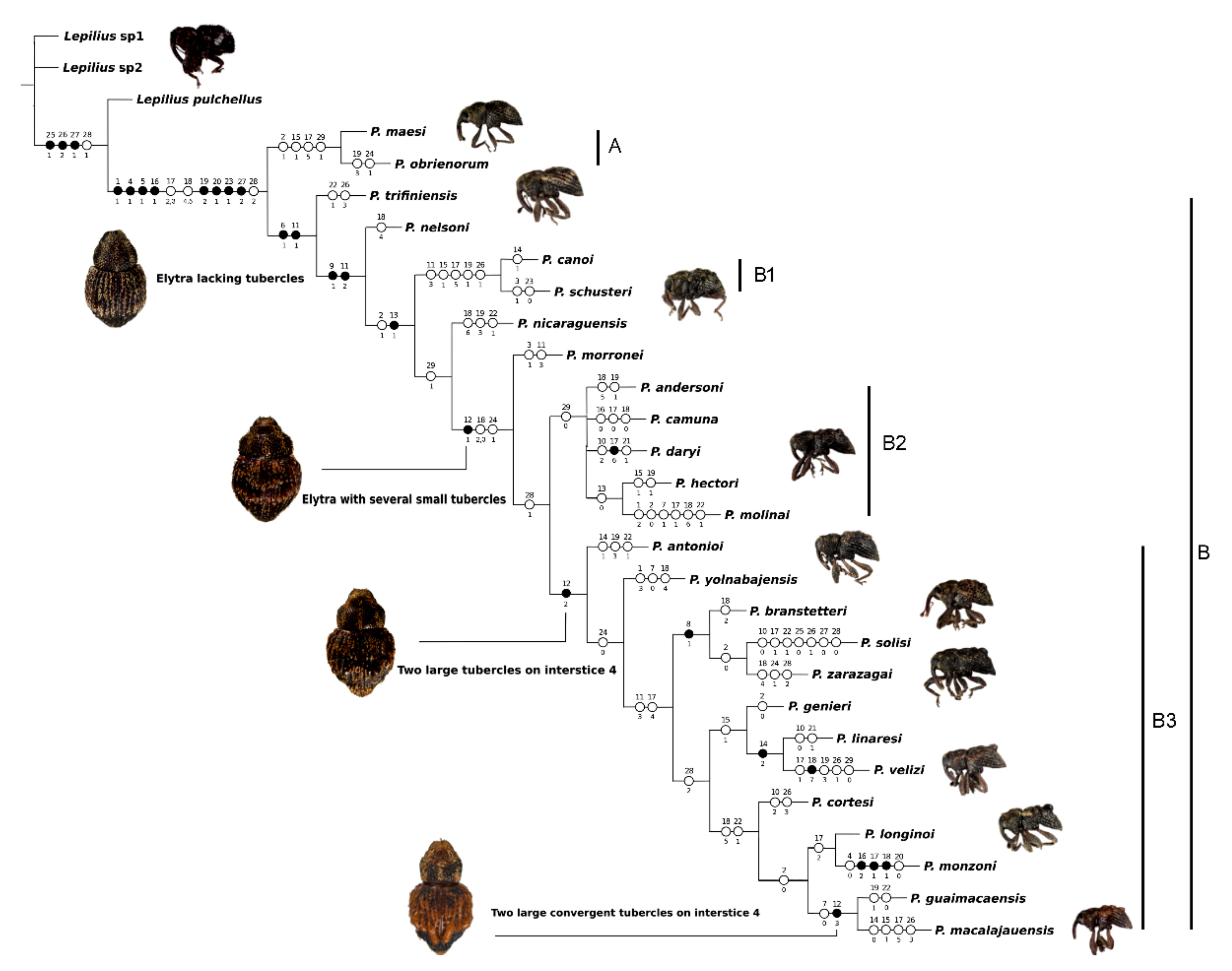
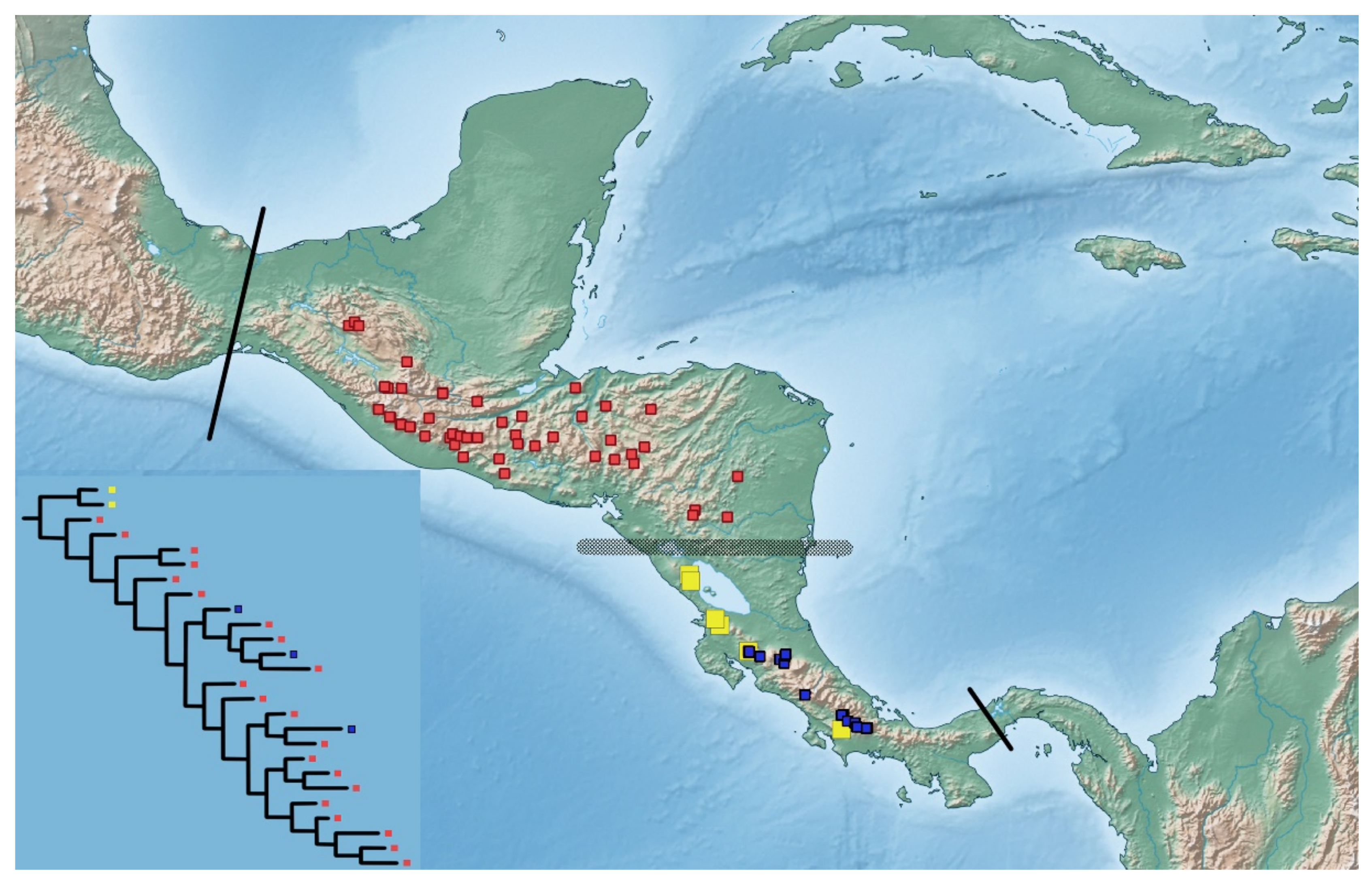
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrios-Izás, M.A.; Anderson, R.S.; Morrone, J.J. A taxonomic monograph of the leaf-litter inhabiting weevil genus Plumolepilius new genus (Coleoptera: Curculionidae: Molytinae: Conotrachelini) from Mexico, Guatemala, and El Salvador. Zootaxa 2016, 4168, 61–91. [Google Scholar] [CrossRef] [PubMed]
- Barrios-Izás, M.A. Taxonomy of the weevil genus Plumolepilius Barrios-Izás & Anderson (Coleoptera: Curculionidae: Molytinae: Conotrachelini): New species from Central America. Zootaxa 2020, 4768, 151–192. [Google Scholar] [CrossRef]
- Morrone, J.J. Biogeographical regionalisation of the Neotropical region. Zootaxa 2014, 3782, 1–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacVean, C.; Schuster, J.C. Altitudinal Distribution of Passalid Beetles (Coleoptera, Passalidae) and Pleistocene Dispersal on the Volcanic Chain of Northern Central America. Biotropica 1981, 13, 29. [Google Scholar] [CrossRef]
- Halffter, G.; Morrone, J.J. An analytical review of Halffter’s Mexican Transition Zone, and its relevance for evolutionary biogeography, ecology and biogeographical regionalization. Zootaxa 2017, 4226, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Steedman, H.F. Dimethyl Hydantoin Formaldehyde: A new Water-soluble Resin for Use as a Mounting Medium. J. Cell Sci. 1958, 3, 451–452. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Catalano, S.A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Goloboff, P.A. Estimating characters weights during tree search. Cladistics 1993, 9, 83–91. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Carpenter, J.M.; Arias, J.S.; Miranda-Esquivel, D.R. Weighting against homoplasy improves phylogenetic analysis of morphological data sets. Cladistics 2008, 24, 758–773. [Google Scholar] [CrossRef]
- Anderson, R.S. A taxonomic monograph of the Middle American leaf-litter inhabiting weevil genus Theognete Champion (Coleoptera: Curculionidae; Molytinae; Lymantini). Zootaxa 2010, 2458, 1–127. [Google Scholar] [CrossRef] [Green Version]
- Luna-Cozar, J.; Anderson, R.S.; Jones, R.W.; León-Corté, J.L. A taxonomic monograph of the genus Tylodinus Champion (Coleoptera: Curculionidae: Cryptorhynchinae: Tylodina) of Chiapas, Mexico. Zootaxa 2014, 3788, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.S. The genus Lepilius Champion (Coleoptera: Curculionidae: Molytinae: Conotrachelini) in North America, with description of a new species, Lepilius chisosensis Anderson, from Big Bend National Park, Texas, U.S.A. Coleopts. Bull. 2012, 66, 67–69. [Google Scholar] [CrossRef]
- Gilbert, E.E.; Howden, A.T. Neoubychia mexicana, a new genus and new species of Raymondionyminae, with a discussion of the characters of blind weevils (Coleoptera: Curculionidae). Coleopts. Bull. 1987, 41, 141–149. [Google Scholar]
- Morrone, J.J.; Hlaváč, P. Checklist of the micro- and anophthalmic soil-dwelling weevils of the world (Coleoptera: Curculionidae). Zootaxa 2017, 4239, 1–102. [Google Scholar] [CrossRef] [PubMed]
- Halffter, G. Un nuevo patrón de dispersión en la zona de transición mexicana: El mesoamericano de montaña. Folia Entomol. Mex. 1978, 39, 219–222. [Google Scholar]
- Halffter, G. Biogeography of the montane entomofauna of Mexico and Central America. Annu. Rev. Entomol. 1987, 32, 95–114. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorites. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Taxa | ||||||
|---|---|---|---|---|---|---|
| Lepilius spp. | 00000 | 00000 | 00000 | 00000 | 00000 | 0000 |
| Lepilius pulchellus | 00000 | 00000 | 00000 | 00000 | 00001 | 2110 |
| P. andersoni | 11011 | 11011 | 21100 | 14511 | 00111 | 2210 |
| P. antonioi | 11011 | 11010 | 22110 | 12231 | 01111 | 2211 |
| P. branstetteri | 11011 | 11111 | 32100 | 14251 | 00101 | 2211 |
| P. camuna | 11011 | 10?11 | 2?100 | 00021 | ???11 | 2210 |
| P. canoi | 11011 | 11010 | 30111 | 15511 | 00101 | 1220 |
| P. cortesi | 11011 | 11012 | 32100 | 14521 | 01101 | 3220 |
| P. daryi | 11011 | 10012 | 21100 | 16221 | 10111 | 2220 |
| P. genieri | 10011 | 11011 | 32101 | 15321 | 00101 | 2221 |
| P. guaimacaensis | 10011 | 10011 | 33110 | 14511 | 00101 | 2221 |
| P. hectori | 11011 | 10011 | 21001 | 15211 | 00111 | 2210 |
| P. linaresi | 11011 | 11010 | 32121 | 15321 | 10101 | 2221 |
| P. longinoi | 10011 | 11011 | 32110 | 12521 | 01101 | 2220 |
| P. macalajauensis | 10011 | 10011 | 33101 | 15521 | 01101 | 3221 |
| P. maesi | 11011 | 00000 | 00001 | 15521 | 00101 | 2221 |
| P. molinai | 10011 | 11011 | 21000 | 11621 | 01111 | 2210 |
| P. morronei | 11111 | 11010 | 31100 | 12221 | 00111 | 2221 |
| P. nelsoni | 10011 | 10?10 | 2?000 | 14421 | ???01 | 2220 |
| P. nicaraguensis | 11011 | 11011 | 20100 | 12631 | 01101 | 2221 |
| P. obrienorum | 11011 | 01000 | 00001 | 15531 | 00111 | 2221 |
| P. schusteri | 11111 | 11010 | 30101 | ?5511 | 00001 | 1220 |
| P. solisi | 10011 | 11110 | 32100 | 11331 | 01100 | 1001 |
| P. trifiniensis | 10011 | 11000 | 10000 | 12521 | 01101 | 3220 |
| P. velizi | 11011 | 11011 | 32121 | 11731 | 00101 | 1220 |
| P. yolnabajensis | 11011 | 10011 | 22100 | 12421 | 00101 | 2211 |
| P. zarazagai | 10011 | 11111 | 32100 | 14421 | 00111 | 2221 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrios-Izás, M.A.; Morrone, J.J. Systematics and Biogeography of the New World Genus Plumolepilius (Coleoptera: Curculionidae). Diversity 2021, 13, 596. https://doi.org/10.3390/d13110596
Barrios-Izás MA, Morrone JJ. Systematics and Biogeography of the New World Genus Plumolepilius (Coleoptera: Curculionidae). Diversity. 2021; 13(11):596. https://doi.org/10.3390/d13110596
Chicago/Turabian StyleBarrios-Izás, Manuel A., and Juan J. Morrone. 2021. "Systematics and Biogeography of the New World Genus Plumolepilius (Coleoptera: Curculionidae)" Diversity 13, no. 11: 596. https://doi.org/10.3390/d13110596
APA StyleBarrios-Izás, M. A., & Morrone, J. J. (2021). Systematics and Biogeography of the New World Genus Plumolepilius (Coleoptera: Curculionidae). Diversity, 13(11), 596. https://doi.org/10.3390/d13110596