
Comparative Floral and Pollen Micromorphology of Leonurus japonicus and L. macranthus (Lamiaceae)



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Microscopic Observations and Cell Type Classification
3. Results
3.1. Leonurus japonicus Houtt
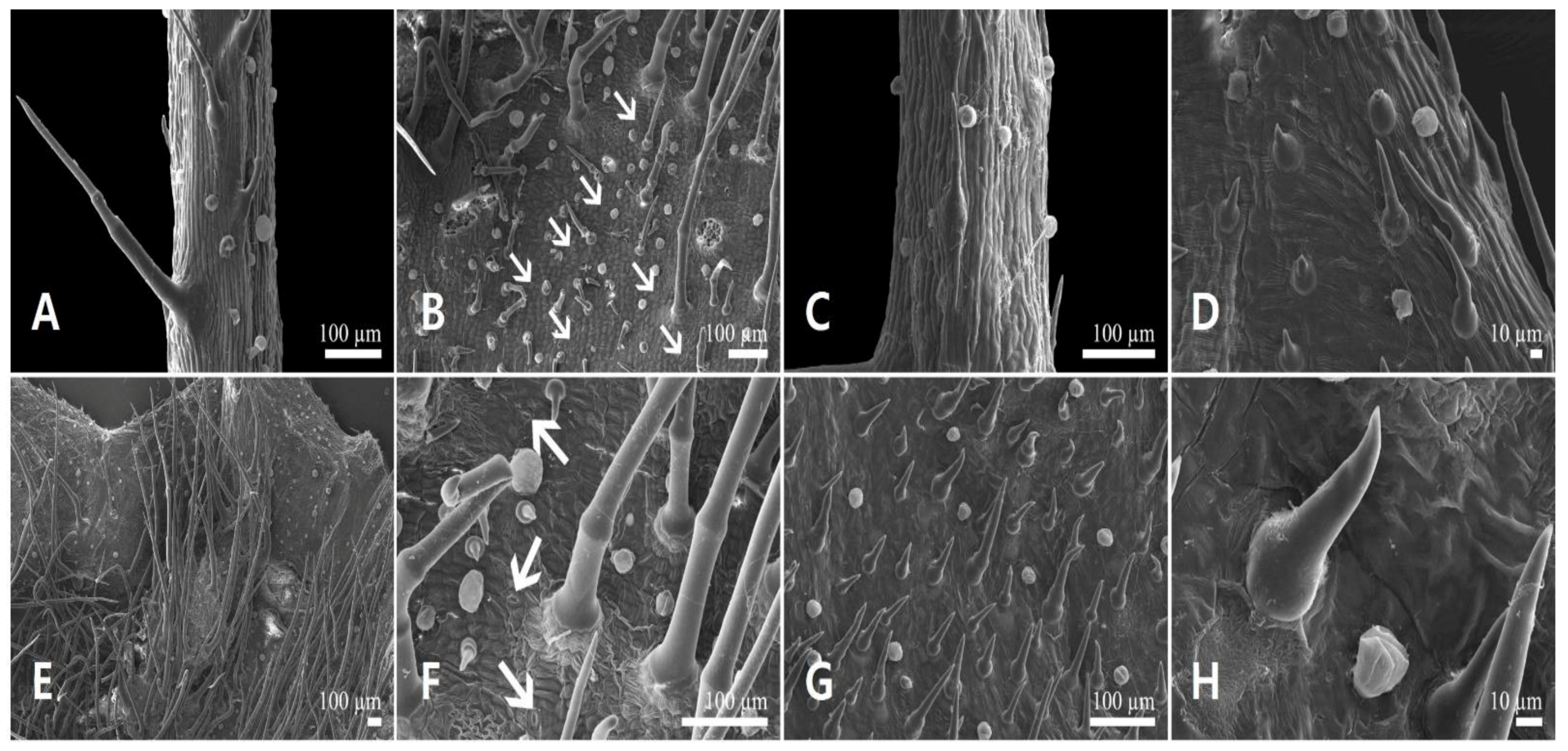
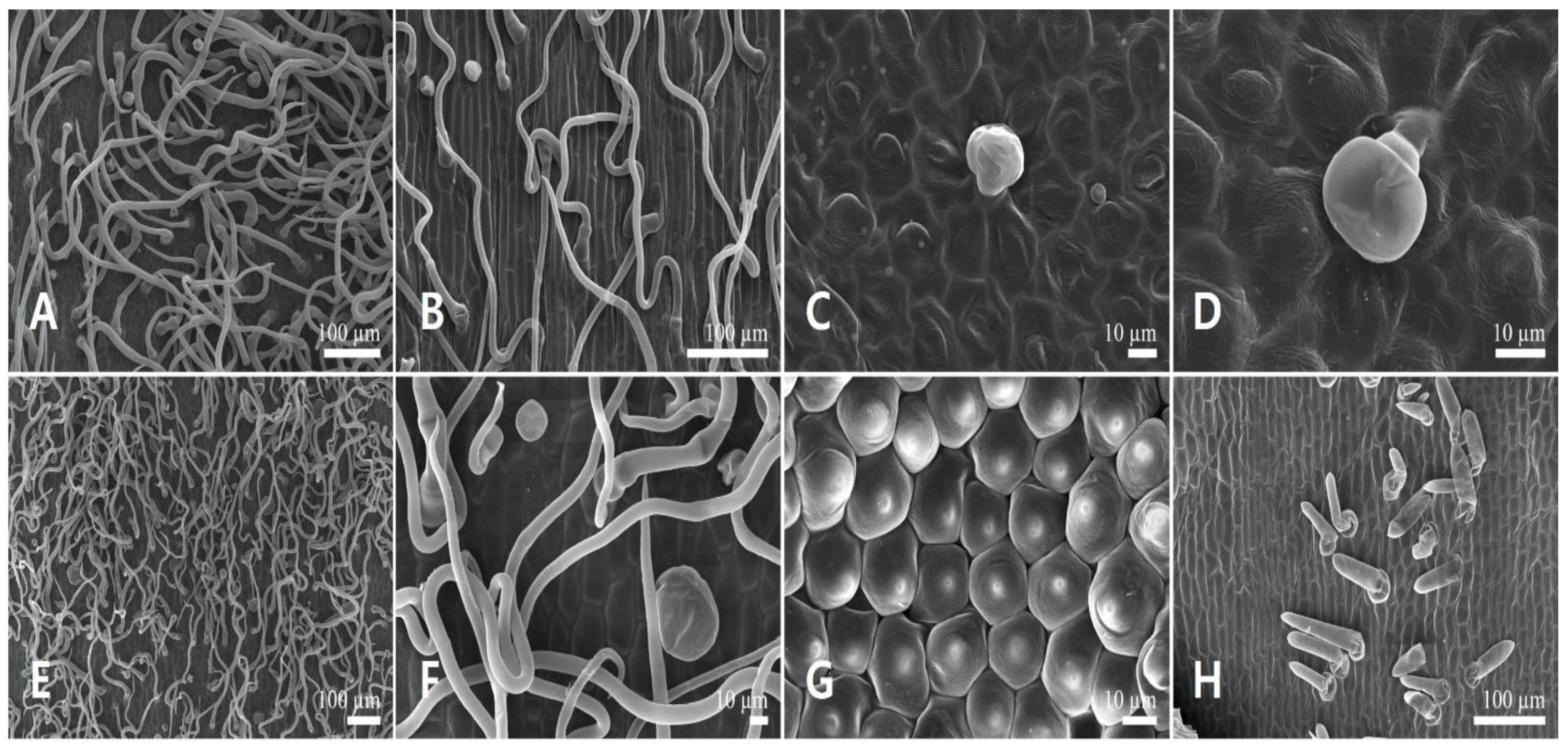
3.1.1. Floral Micromorphology
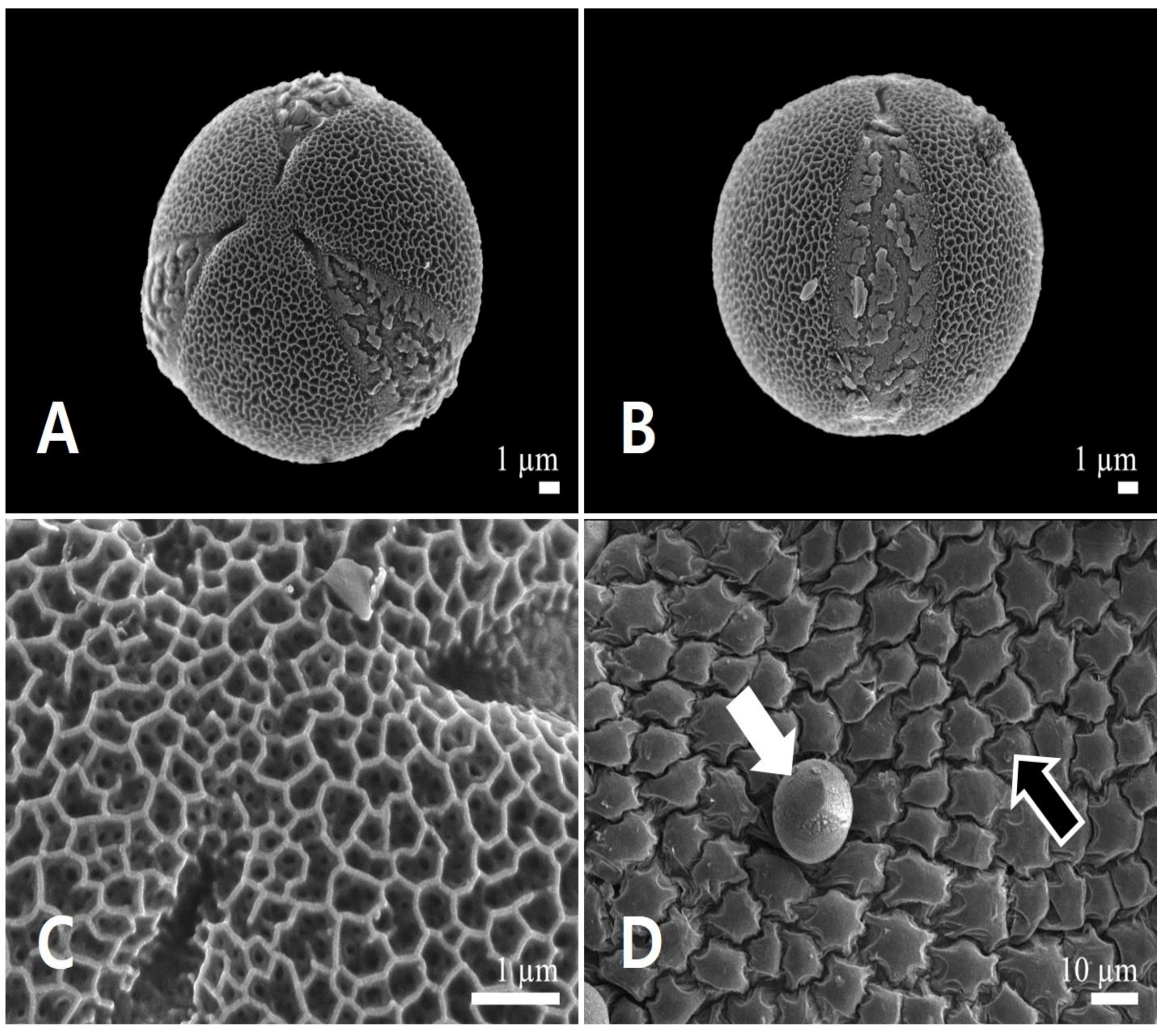
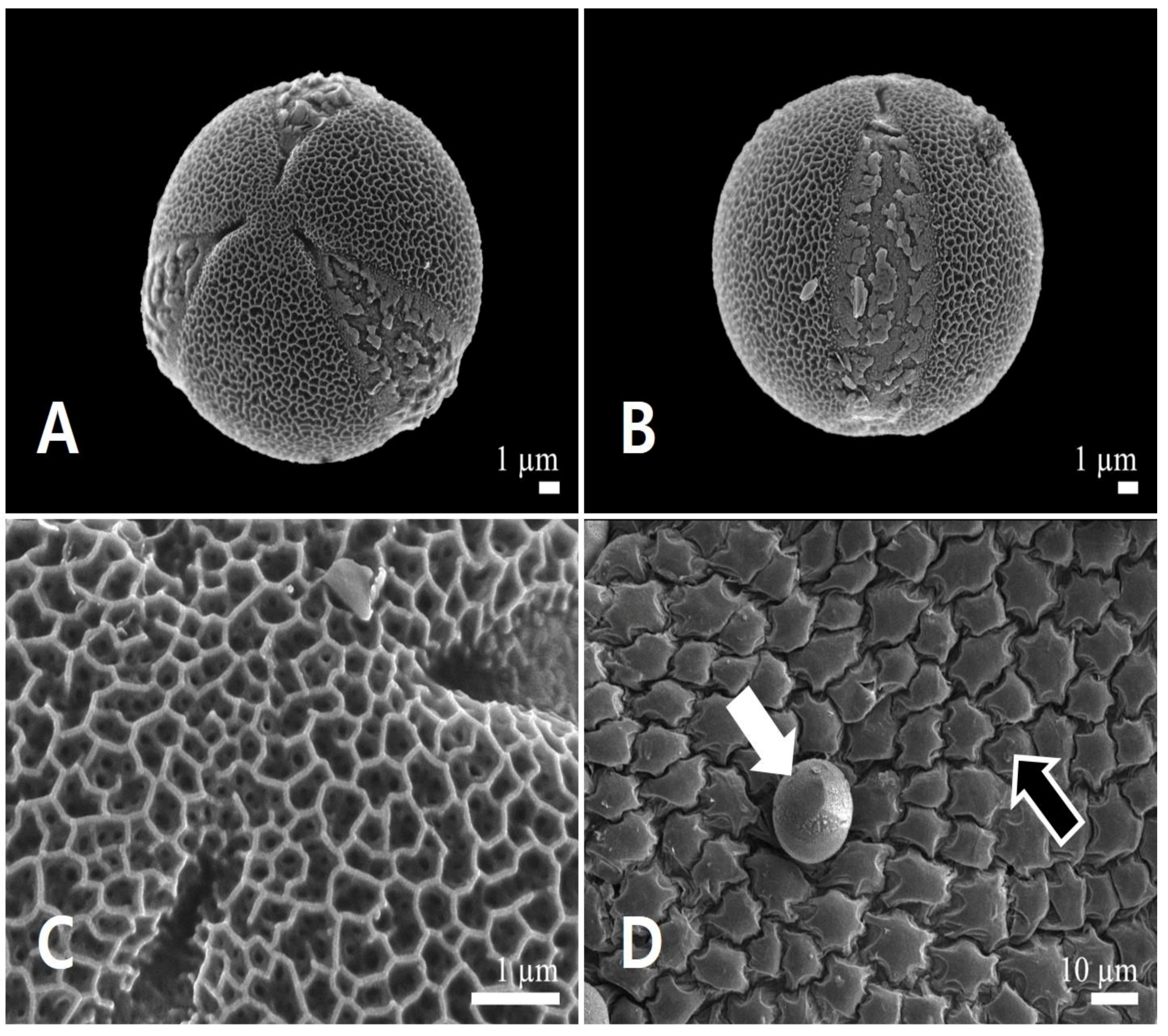
3.1.2. Pollen Micromorphology
3.2. Leonurus macranthus Maxim
3.2.1. Floral Micromorphology
3.2.2. Pollen Micromorphology
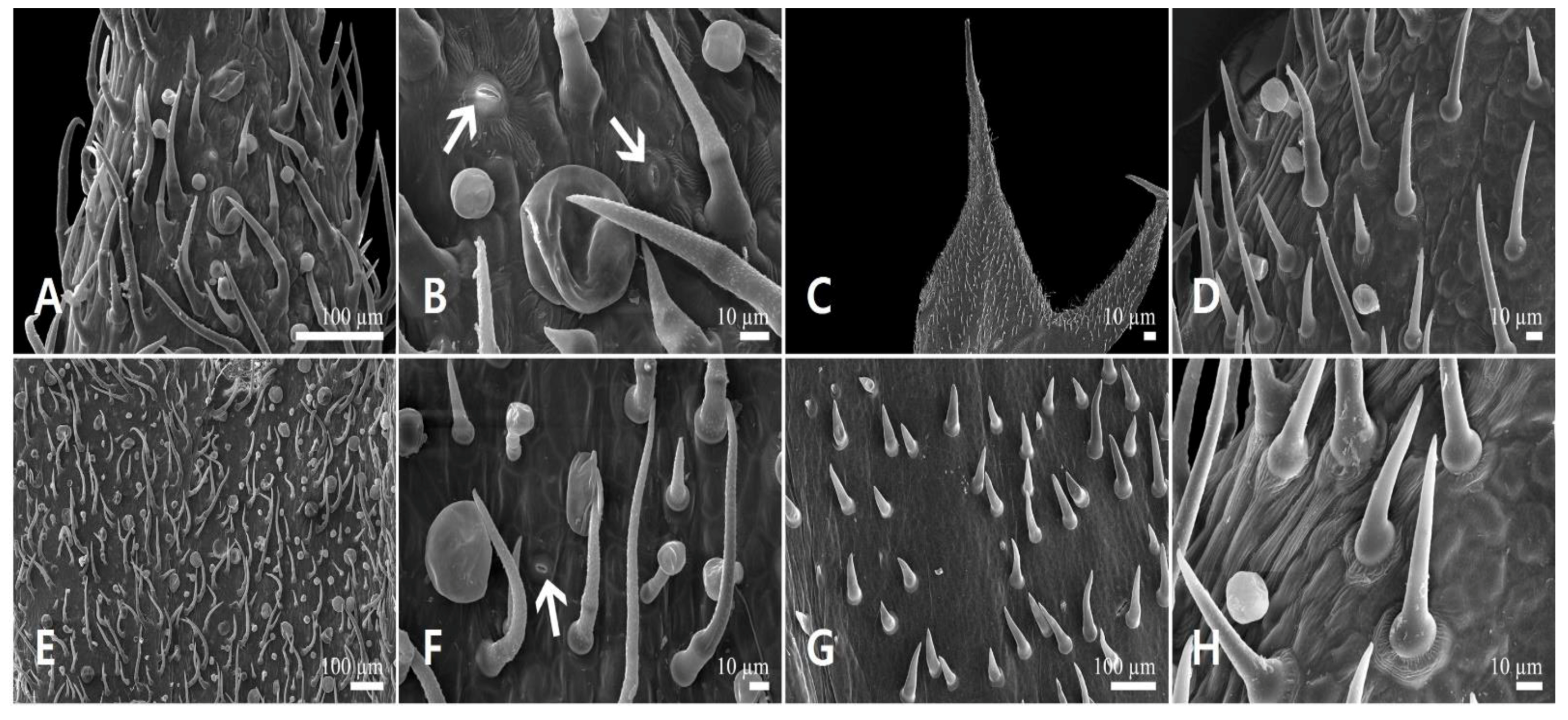
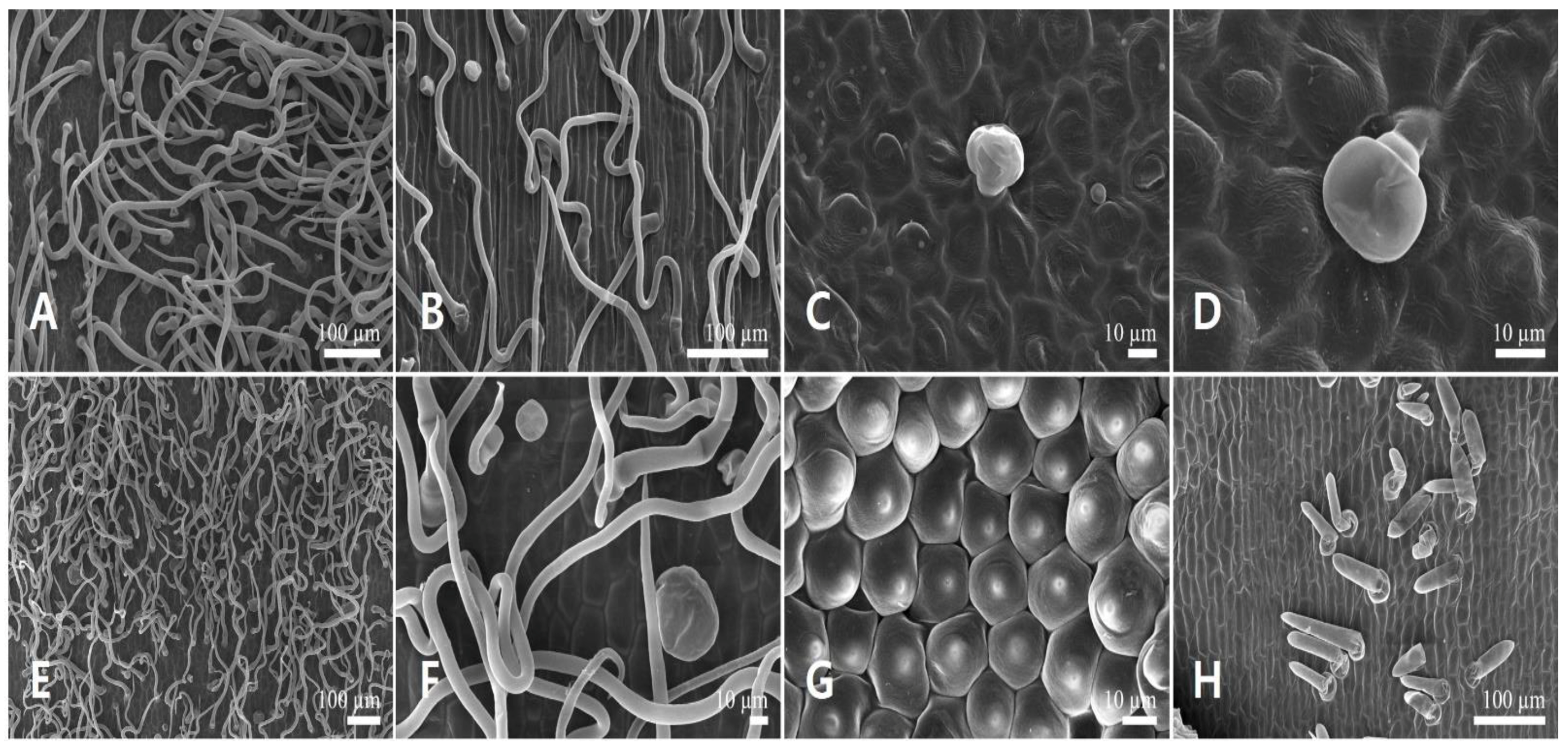
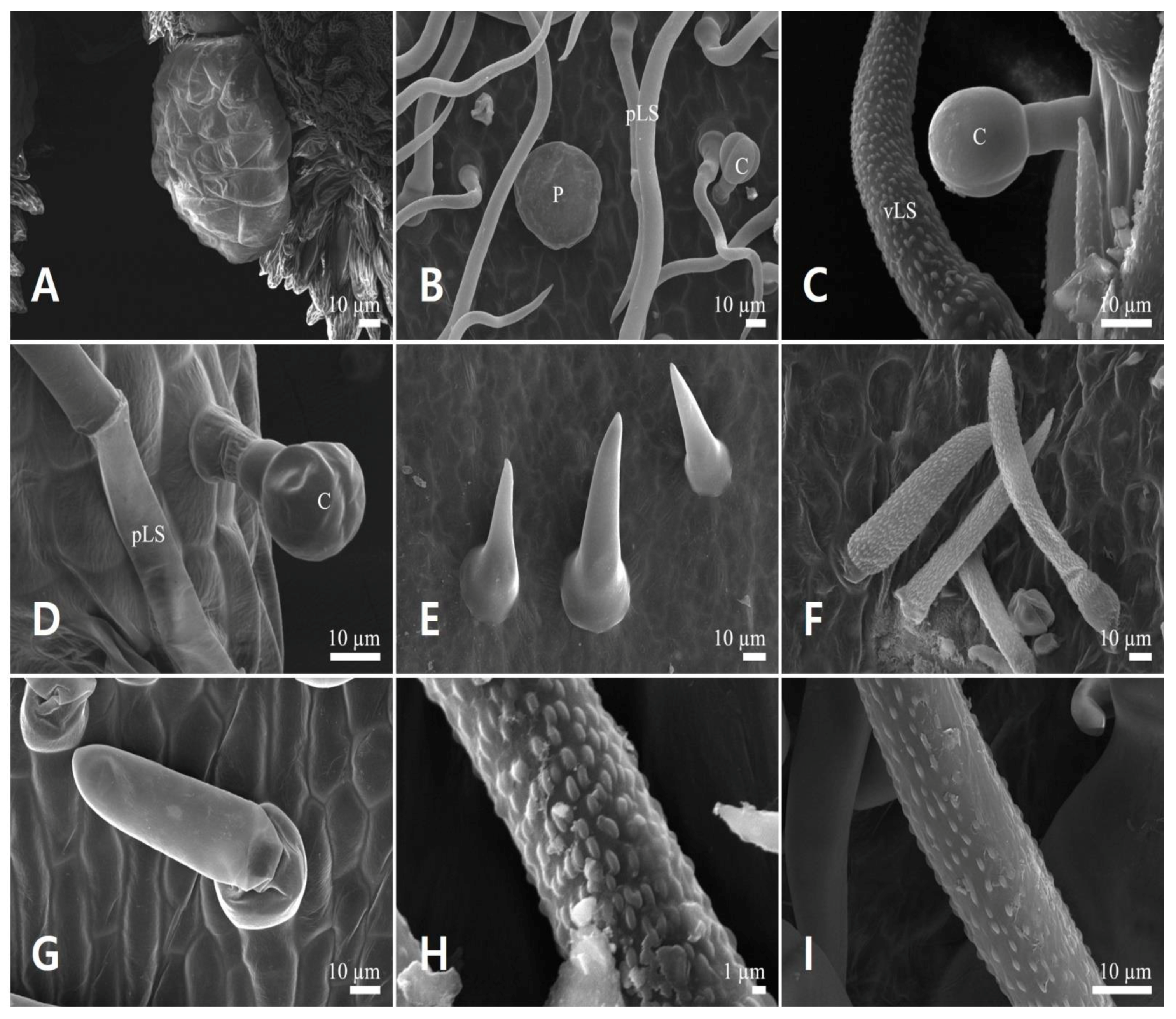
3.3. Floral Trichome Diversity
3.3.1. Glandular Trichomes
3.3.2. Non-Glandular Trichomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Whitney, H.M.; Bennett, K.V.; Dorling, M.; Sandbach, L.; Prince, D.; Chittka, L.; Glover, B.J. Why do so many petals have conical epidermal cells? Ann. Bot. 2011, 108, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Glover, B.J.; Martin, C. The role of petal cell shape and pigmentation in pollination success in Antirrhinum majus. Heredity 1998, 80, 778–784. [Google Scholar] [CrossRef]
- Costa, V.B.S.; Pimentel, R.M.M.; Chagas, M.G.S.; Alves, G.D.; Castro, C.C. Petal micromorphology and its relationship to pollination. Plant Biol. 2017, 19, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Kay, Q.O.N.; Daoud, H.S.; Stirton, C.H. Pigment distribution, light reflection and cell structure in petals. Bot. J. Linn. Soc. 1981, 83, 57–83. [Google Scholar] [CrossRef]
- Gkikas, D.; Argiropoulos, A.; Rhizopoulou, S. Epidermal focusing of light and modelling of reflectance in floral-petals with conically shaped epidermal cells. Flora 2015, 212, 38–45. [Google Scholar] [CrossRef]
- Keven, P.G.; Lane, M.A. Flower petal microtexture is a tactile cue for bees. Proc. Natl. Acad. Sci. USA. 1985, 82, 4750–4752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitney, H.M.; Chittka, L.; Bruce, T.J.; Glover, B.J. Conical epidermal cells allow bees to grip flowers and increase foraging efficiency. Curr. Biol. 2009, 19, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Kolosova, N.; Sherman, D.; Karlson, D.; Dudareva, N. Cellular and subcellular localization of S-adenosyl-l-methionine: Benzoic acid carboxyl methyltransferase, the enzyme responsible for biosynthesis of the volatile ester methylbenzoate in snapdragon flowers. Plant Physiol. 2001, 126, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Fahn, A. Secretory tissues in plants. New Phytol. 1988, 108, 229–257. [Google Scholar] [CrossRef]
- Palo, R.T.; Robbins, C.T. Plant Defenses against Mammalian Herbivory; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Ferreira, J.F.; Janick, J. Floral morphology of Artemisia annua with special reference to trichomes. Int. J. Plant Sci. 1995, 156, 807–815. [Google Scholar] [CrossRef]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Annu. Rev. Plant Biol. 2012, 63, 431–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mithöfer, A.; Boland, W.; Maffei, M.E. Chemical ecology of plant–insect interactions. Annu. Plant Rev. 2018, 34, 261–291. [Google Scholar] [CrossRef]
- Endress, P.K. Floral structure and evolution of primitive angiosperms: Recent advances. Plant Syst. Evol. 1994, 192, 79–97. [Google Scholar] [CrossRef]
- Luckow, M.; Grimes, J. A survey of anther glands in the mimosoid legume tribes Parkieae and Mimoseae. Am. J. Bot. 1997, 84, 285–297. [Google Scholar] [CrossRef] [PubMed]
- de Barros, T.C.; Teixeira, S.P. Revisited anatomy of anther glands in mimosoids (Leguminosae). Int. J. Plant Sci. 2016, 177, 18–33. [Google Scholar] [CrossRef]
- de Barros, T.C.; Pedersoli, G.D.; Teixeira, S.P. Anther glands in Mimosoideae (Leguminosae) are emergences with a conserved meristematic origin. Flora 2017, 226, 1–9. [Google Scholar] [CrossRef]
- Leite, V.G.; Mansano, V.F.; Pansarin, E.R.; Teixeira, S.P. Presence of the anther gland is a key feature in pollination of the early-branching papilionoids Dipteryx alata and Pterodon pubescens (Leguminosae). Plant Biol. 2019, 21, 1016–1023. [Google Scholar] [CrossRef]
- Davies, K.L.; Stpiczyńska, M.; Gregg, A. Nectar-secreting floral stomata in Maxillaria anceps Ames & C. Schweinf. (Orchidaceae). Ann. Bot. 2005, 96, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Subedi, A.; Chaudhary, R.P.; van Achterberg, C.; Heijerman, T.; Lens, F.; Van Dooren, T.J.; Gravendeel, B. Pollination and protection against herbivory of Nepalese Coelogyninae (Orchidaceae). Am. J. Bot. 2011, 98, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Hadacek, F.; Weber, M. Club-shaped organs as additional osmophores within the Sauromatum inflorescence: Odour analysis, ultrastructural changes and pollination aspects. Plant Biol. 2002, 4, 367–383. [Google Scholar] [CrossRef]
- Song, J.H.; Hong, S.P. Identity and localization of floral scent components in an androdioecious species, Chionanthus retusus (Oleaceae). J. Asia-Pac. Biodivers. 2020, 13, 288–294. [Google Scholar] [CrossRef]
- Moon, H.K.; Hong, S.P. The taxonomic consideration of petal and sepal micromorphology in Lycopus L. (Mentheae-Lamiaceae). Korean J. Pl. Taxon. 2004, 34, 273–285. [Google Scholar] [CrossRef]
- Hong, S.P.; Moon, H.K. Gynodioecy in Lycopus maackianus Makino (Lamiaceae) in Korea: Floral dimorphism and nutlet production. Flora 2003, 198, 461–467. [Google Scholar] [CrossRef]
- Jang, T.S.; Lee, J.; Hong, S.P. A systematic study of Glechoma L. (Lamiaceae) based on micromorphological characters and nuclear ribosomal ITS sequences. Korean J. Pl. Taxon. 2014, 44, 22–32. [Google Scholar] [CrossRef]
- Jang, T.S.; Hong, S.P. Floral micromorphology and microsporogenesis of the gynodioecious herb Glechoma longituba (Lamiaceae). Nord. J. Bot. 2015, 33, 708–714. [Google Scholar] [CrossRef]
- Atalay, Z.; Celep, F.; Bara, F.; Doğan, M. Systematic significance of anatomy and trichome morphology in Lamium (Lamioideae; Lamiaceae). Flora 2016, 225, 60–75. [Google Scholar] [CrossRef]
- Santos Tozin, L.R.D.; de Melo Silva, S.C.; Rodrigues, T.M. Non-glandular trichomes in Lamiaceae and Verbenaceae species: Morphological and histochemical features indicate more than physical protection. N. Z. J. Bot. 2016, 54, 446–457. [Google Scholar] [CrossRef]
- Eiji, S.; Salmaki, Y. Evolution of trichomes and its systematic significance in Salvia (Mentheae; Nepetoideae; Lamiaceae). Bot. J. Linn. Soc. 2016, 180, 241–257. [Google Scholar] [CrossRef] [Green Version]
- Haratym, W.; Weryszko-Chmielewska, E. Ultrastructural and histochemical analysis of glandular trichomes of Marrubium vulgare L. (Lamiaceae). Flora 2017, 231, 11–20. [Google Scholar] [CrossRef]
- Mannethody, S.; Purayidathkandy, S. Trichome micromorphology and its systematic significance in Asian Leucas (Lamiaceae). Flora 2018, 242, 70–78. [Google Scholar] [CrossRef]
- Gul, S.; Ahmad, M.; Zafar, M.; Bahadur, S.; Sultana, S.; Begum, N.; Shah, S.N.; Zaman, W.; Ullah, F.; Ayaz, A.; et al. Taxonomic study of subfamily Nepetoideae (Lamiaceae) by palynomorphological approach. Microsc. Res. Tech. 2019, 82, 1021–1031. [Google Scholar] [CrossRef]
- de Almeida, V.P.; Raman, V.; Raeski, P.A.; Urban, A.M.; Swiech, J.N.; Miguel, M.D.; Farago, P.V.; Khan, I.A.; Budel, J.M. Anatomy, micromorphology, and histochemistry of leaves and stems of Cantinoa althaeifolia (Lamiaceae). Microsc. Res. Tech. 2020, 83, 551–557. [Google Scholar] [CrossRef]
- Siadati, S.; Salmaki, Y.; Bräuchler, C. Trichome morphology provides phylogenetically informative signal for generic delimitation in tribe Marrubieae (Lamiaceae). Flora 2020, 273, 151720. [Google Scholar] [CrossRef]
- Ma, Z.; Bramley, G.L.; Zhang, D. Pollen morphology of Callicarpa L. (Lamiaceae) from China and its systematic implications. Plant Syst. Evol. 2016, 302, 67–88. [Google Scholar] [CrossRef]
- Marzouk, R.I.; Salama, M.; Askar, A.B.M. Pollen morphology of Teucrium L. (Lamiaceae, Ajugoideae) in Libya. Bangladesh J. Plant Taxon. 2017, 24, 219–226. [Google Scholar] [CrossRef]
- Doaigey, A.R.; El-Zaidy, M.; Alfarhan, A.; Milagy, A.E.S.; Jacob, T. Pollen morphology of certain species of the family Lamiaceae in Saudi Arabia. Saudi J. Biol. Sci. 2018, 25, 354–360. [Google Scholar] [CrossRef]
- Bedolla-Garcia, B.Y.; Castro-Morales, M.; Cultid-Medina, C.A. Comparative assessment of pollen micromorphology of Salvia assurgens (Lamiaceae), an endemic sage from Mexico. Phytotaxa 2020, 458, 183–194. [Google Scholar] [CrossRef]
- Özler, H.; Kahraman, A.; Pehlivan, S.; Dogan, M.; Baser, B.; Fisne, A.Y.; Bagherpour, S. Contribution to the knowledge of the pollen morphology of the genus Salvia (Lamiaceae). Phytotaxa 2020, 428, 228–240. [Google Scholar] [CrossRef]
- Krestovskaja, T. Systematics and phytogeography of Leonurus L. In Advances in Labiate Science; Harley, R.M., Reynolds, R.M., Eds.; Royal Botanic Gardens, Kew: Richmond, UK, 1992; pp. 139–148. [Google Scholar]
- Harley, R.M.; Atkins, S.; Budantsey, A.L.; Cantino, P.D.; Conn, B.J.; Grayer, R.; Harley, M.M.; de Kok, R.; Krestovskaja, T.; Morales, R.; et al. Labiatae. In Families and Genera of Vascular Plants, Flowering Plants: Dicotyledons; Lamiales (except Acanthaceae including Avicenniaceae) Vol. 7; Kubitzki, K., Kadereit, J.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 167–275. [Google Scholar]
- Murata, G.; Yamazaki, T. Lamiaceae. In Flora of Japan Vol. 3a; Iwatsuki, K., Yamazaki, T., Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 1993; pp. 273–321. [Google Scholar]
- Li, X.W.; Hedge, I.C. Leonurus. In Flora of China Vol. 17; Wu, C.Y., Raven, P.H., Eds.; Science Press and Missouri Botanical Garden Press: St. Louis, UK; Beijing, China, 1994; pp. 162–165. [Google Scholar]
- Park, S.J. Leonurus. In Flora of Korea Asteridae: Loganiaceae to Oleaceae Vol. 6a; Flora of Korea Editorial Committee: Seoul, Korea, 2018; pp. 105–106. ISBN 978-89-6811-334-5. [Google Scholar]
- Korea Food and Drug Administration. The Korean Pharmacophoeia, 12th ed.; Korea Food and Drug Administration: Seoul, Korea, 2019. [Google Scholar]
- Korea Institute of Oriental Medicine. Defining Dictionary for Medicinal Herbs. 2021. Available online: http://https://oasis.kiom.re.kr/herblib/hminfo/hbmcod/hbmcodList.do (accessed on 21 February 2021).
- Galetto, L.; Torres, C. Nectar sugar composition and pollinators for the naturalized exotic Leonurus japonicus (Lamiaceae) in Central Argentina. Int. J. Plant Reprod. Biol. 2010, 2, 1–4. [Google Scholar]
- Moyano, F.; Cocucci, A.; Sersic, A. Accessory pollen adhesive from glandular trichomes on the anthers of Leonurus sibiricus L. (Lamiaceae). Plant Biol. 2003, 5, 411–418. [Google Scholar] [CrossRef]
- Borna, F.; Ahmad, N.M.; Luo, S.; Trethowan, R. Reproductive biology of a medicinally important plant Leonurus cardiaca (Lamiaceae). Aust. J. Bot. 2016, 64, 342–358. [Google Scholar] [CrossRef]
- Shekari, A.; Mahdipour, M.H.; Nazeri, V.; Shokrpour, M. The reproductive biology of motherwort (Leonurus cardiaca L.). J. Biodivers. Environ. Sci. 2018, 12, 109–116. [Google Scholar]
- Park, I.; Yang, S.; Song, J.-H.; Moon, B.C. Dissection for floral micromorphology and plastid genome of valuable medicinal borages Arnebia and Lithospermum (Boraginaceae). Front. Plant Sci. 2020, 11, 606463. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-H.; Hong, S.P. A taxonomic revision of the genus Sorbaria (Rosaceae) with a new infrageneric classification based on morphology, micromorphology, and palynology. Phytotaxa 2021, 487, 1–25. [Google Scholar] [CrossRef]
- Barthlott, W. Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects. Nord. J. Bot. 1981, 1, 345–355. [Google Scholar] [CrossRef]
- Barthlott, W. Scanning electron microscopy of the epidermal surface in plants. In Scanning Electron Microscopy in Taxonomy and Functional Morphology; Claugher, D., Ed.; Clarendon Press: Oxford, UK, 1990; pp. 69–94. [Google Scholar]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Millner, H.J.; Baldwin, T.C. Floral micromorphology of the genus Restrepia (Orchidaceae) and the potential consequences for pollination. Flora 2016, 225, 10–19. [Google Scholar] [CrossRef]
- Ojeda, D.I.; Valido, A.; Fernández de Castro, A.G.; Ortega-Olivencia, A.; Fuertes-Aguilar, J.; Carvalho, J.A.; Santos-Guerra, A. Pollinator shifts drive petal epidermal evolution on the Macaronesian Islands bird-flowered species. Biol. Lett. 2016, 12, 20160022. [Google Scholar] [CrossRef] [Green Version]
- Piwowarczyk, R.; Kasińska, J. Petal epidermal micromorphology in holoparasitic Orobanchaceae and its significance for systematics and pollination ecology. Aust. Syst. Bot. 2017, 30, 48–63. [Google Scholar] [CrossRef]
- Christensen, K.I.; Hansen, H.V. SEM studies of epidermal patterns of petals in the angiosperms. Opera Bot. 1998, 135, 5–91. [Google Scholar]
- Kevan, P.G.; Baker, H.G. Insects as flower visitors and pollinators. Annu. Rev. Entomol. 1983, 28, 407–453. [Google Scholar] [CrossRef]
- Rands, S.A.; Glover, B.J.; Whitney, H.M. Floral epidermal structure and flower orientation: Getting to grips with awkward flowers. Arthropod Plant Interact. 2011, 5, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Werker, E. Trichome diversity and development. Adv. Bot. Res. 2000, 31, 1–35. [Google Scholar] [CrossRef]
- Levin, D.A. The role of trichomes in plant defense. Q. Rev. Biol. 1973, 48, 3–15. [Google Scholar] [CrossRef]
- Dalin, P.; Ågren, J.; Björkman, C.; Huttunen, P.; Kärkkäinen, K. Leaf trichome formation and plant resistance to herbivory. In Induced Plant Resistance to Herbivory; Schaller, A., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 89–105. [Google Scholar]
- Pansarin, E.R.; Maciel, A.A. Evolution of pollination systems involving edible trichomes in orchids. AoB Plants 2017, 9, plx033. [Google Scholar] [CrossRef] [Green Version]
- Lustofin, K.; Świątek, P.; Stolarczyk, P.; Miranda, V.F.; Płachno, B.J. Do food trichomes occur in Pinguicula (Lentibulariaceae) flowers? Ann. Bot. 2020, 126, 1039–1048. [Google Scholar] [CrossRef]
- Kaya, A.; Demirci, B.; Baser, K.H.C. Micromorphology of glandular trichomes of Nepeta congesta Fisch. & Mey. var. congesta (Lamiaceae) and chemical analysis of the essential oils. S. Afr. J. Bot. 2007, 73, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Schmiderer, C.; Grassi, P.; Novak, J.; Weber, M.; Franz, C. Diversity of essential oil glands of clary sage (Salvia sclarea L., Lamiaceae). Plant Biol. 2008, 10, 433–440. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Tozin, L.R.; Rodrigues, T.M. Glandular trichomes in the tree-basil (Ocimum gratissimum L., Lamiaceae): Morphological features with emphasis on the cytoskeleton. Flora 2019, 259, 151459. [Google Scholar] [CrossRef]
- Konarska, A.; Chmielewski, P. Taxonomic traits in the microstructure of flowers of parasitic Orobanche picridis with particular emphasis on secretory structures. Protoplasma 2020, 257, 299–317. [Google Scholar] [CrossRef] [Green Version]
- Raguso, R.A. Functions of essential oils and natural volatiles in plant-insect interactions. In Handbook of Essential Oils. Science, Technology, and Applications, 3rd ed.; Hüsnü Can Başer, K., Buchbauer, G., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 481–496. ISBN 9781351246460. [Google Scholar]
- de Melo, M.C.; Borba, E.L.; Paiva, E.A.S. Morphological and histological characterization of the osmophores and nectaries of four species of Acianthera (Orchidaceae: Pleurothallidinae). Plant Syst. Evol. 2010, 286, 141–151. [Google Scholar] [CrossRef]
- Maiti, S.; Mitra, A. Morphological, physiological and ultrastructural changes in flowers explain the spatio-temporal emission of scent volatiles in Polianthes tuberosa L. Plant Cell Physiol. 2017, 58, 2095–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdtman, G. Pollen morphology and plant taxonomy. IV. Labiatae, Verbanaceae and Avicenniaceae. Sven. Bot. Tidskr. 1945, 39, 279–285. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons; The Clarendon Press: Oxford, UK, 1950; Volume 1. [Google Scholar]
- Cantino, P.D. The phylogenetic significance of stomata and trichomes in the Labiatae and Verbenaceae. J. Arnold Arbor. 1990, 71, 323–370. [Google Scholar]
- Harley, M.M.; Paton, A.; Harley, R.M.; Cade, P.G. Pollen morphological studies in tribe Ocimeae (Nepetoideae: Labiatae): I. Ocimum L. Grana 1992, 31, 161–176. [Google Scholar] [CrossRef]
- Abu-Asab, M.S.; Cantino, P.D. Systematic implications of pollen morphology in subfamilies Lamioideae and Pogostemonoideae (Labiatae). Ann. Missouri Bot. Gard. 1994, 81, 653–686. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leonurus japonicus | Leonurus macranthus | |||||||
|---|---|---|---|---|---|---|---|---|
| Floral Organs | SP | AW | PW | FR | SP | AW | PW | FR |
| Calyx lobe abaxial side | iso, elo to pol | dep, str to cur | cov | stri | iso, elo to pol | dep, str to cur | cov | stri |
| Calyx lobe adaxial side | iso, elo to pol | dep, str to cur | cov | stri | iso, elo to pol | dep, str to cur | cov | stri |
| Calyx tube abaxial side | iso, pol | dep, str to cur | cov | stri | iso, pol | dep, str to cur | cov | stri |
| Calyx tube adaxial side | iso, pol | dep, str to cur | cov | stri | iso, pol | dep, str to cur | cov | stri |
| Upper lipabaxial side | iso, elo to pol | dep, str to cur | cov | smo to stri | iso, pol | dep, str to cur | cov | smo |
| Upper lipadaxial side | iso, rou to pol | dep, cur | cov to central-con | stri | iso, rou to pol | dep, cur | cov | stri |
| Lower lipabaxial side | iso, pol | dep, str to cur | cov to central-con | smo to stri | iso, pol | dep, str to cur | cov | smo |
| Lower lip adaxial side | iso, rou to pol | dep, str to cur | con to central-con | rug | iso, rou to pol | dep, str to cur | con to central con | rug |
| Anther abaxial side | iso, pol | dep, sin | cov | stri | iso, pol | dep, sin | cov | stri |
| Anther apical part | iso, pol | dep, sin | con | stri | iso, pol | dep, sin | con | stri |
| Anther adaxial side | iso, pol | dep, sin | cov | smo | iso, pol | dep, sin | cov | smo |
| Filament | iso, elo to tet | dep, str | cov | smo to stri | iso, elo to tet | dep, sin | cov | smo to stri |
| Stigma | irr, pol | rai, str | flt to cov | stri | irr, pol | dep, str | flt to cov | stri |
| Style | iso, elo | rai, str | flt to cov | stri | iso, elo | dep, str | flt to cov | stri |
| Leonurus japonicus | Leonurus macranthus | t-Value P | |
|---|---|---|---|
| Polar axis length (μm) | 14.17–(16.07)–17.43 | 20.46–(23.47)–24.79 | −24.44 *** |
| Equatorial axis diameter (μm) | 15.11–(17.10)–18.19 | 18.49–(20.66)–22.74 | −12.35 *** |
| P/E | 0.89–(0.94)–1.03 | 1.06–(1.14)–1.22 | N/A |
| Shape | oblate- to prolate-spheroidal | spherical to subprolate | N/A |
| Colpus length (μm) | 13.58–(15.31)–18.75 | 19.48–(21.38)–24.40 | −15.24 *** |
| Exine ornamentation | bi-reticulate | bi-reticulate | N/A |
| Diameter of primary lumina (μm) | 0.30–(0.39)–0.56 | 0.44–(0.66)–0.99 | −7.70 *** |
| Thickness of primary muri (μm) | 0.05–(0.08)–0.09 | 0.10–(0.14)–0.19 | −11.97 *** |
| Shape of primary lumina | angular | angular | N/A |
| Diameter of secondary lumina (μm) | 0.05–(0.09)–0.12 | 0.08–(0.12)–0.19 | −4.13 *** |
| Thickness of secondary muri (μm) | 0.07–(0.12)–0.15 | 0.08–(0.14)–0.20 | −2.46 * |
| Shape of secondary lumina | rounded | rounded | N/A |
| Number of secondary lumina per primary lumen | 3–(4.77)–8 | 3–(6.47)–9 | −4.41 *** |
| Leonurus japonicus | Leonurus macranthus | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Floral Organs | pSS | pLS | vSS | vLS | C | P | SG | FS | pSS | pLS | vSS | vLS | C | P | SG | FS |
| Calyx lobe abaxial | ◎ | ○ | ◎ | ● | ◎ | ◎ | ○ | ◎ | ◎ | ○ | ◎ | ● | ◎ | ◎ | ○ | ◎ |
| Calyx tube abaxial | ○ | ○ | ◎ | ● | ● | ● | ○ | ◎ | ◎ | ○ | ○ | ● | ◎ | ◎ | ○ | ◎ |
| Calyx lobe adaxial | ◎ | ◎ | ○ | ○ | ◎ | ◎ | ○ | ○ | ◎ | ○ | ◎ | ○ | ◎ | ○ | ○ | ○ |
| Calyx tube adaxial | ◎ | ◎ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| Upper lip abaxial | ○ | ○ | ○ | ● | ◎ | ● | ○ | ○ | ○ | ○ | ○ | ● | ◎ | ● | ○ | ○ |
| Lower lip abaxial | ○ | ● | ○ | ○ | ● | ● | ○ | ○ | ○ | ● | ○ | ○ | ● | ● | ○ | ○ |
| Upper lip adaxial | ○ | ○ | ○ | ○ | ◎ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| Lower lip adaxial | ◎ | ◎ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ◎ | ◎ | ○ | ○ | ○ | ○ |
| Filament | ○ | ◎ | ○ | ○ | ◎ | ○ | ○ | ○ | ○ | ◎ | ○ | ○ | ◎ | ○ | ○ | ○ |
| Anther | ○ | ○ | ○ | ○ | ○ | ○ | ◎ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ◎ | ○ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oak, M.-K.; Yang, S.; Choi, G.; Song, J.-H. Comparative Floral and Pollen Micromorphology of Leonurus japonicus and L. macranthus (Lamiaceae). Diversity 2021, 13, 533. https://doi.org/10.3390/d13110533
Oak M-K, Yang S, Choi G, Song J-H. Comparative Floral and Pollen Micromorphology of Leonurus japonicus and L. macranthus (Lamiaceae). Diversity. 2021; 13(11):533. https://doi.org/10.3390/d13110533
Chicago/Turabian StyleOak, Min-Kyeong, Sungyu Yang, Goya Choi, and Jun-Ho Song. 2021. "Comparative Floral and Pollen Micromorphology of Leonurus japonicus and L. macranthus (Lamiaceae)" Diversity 13, no. 11: 533. https://doi.org/10.3390/d13110533
APA StyleOak, M.-K., Yang, S., Choi, G., & Song, J.-H. (2021). Comparative Floral and Pollen Micromorphology of Leonurus japonicus and L. macranthus (Lamiaceae). Diversity, 13(11), 533. https://doi.org/10.3390/d13110533