
Origin of the Chordate Notochord



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
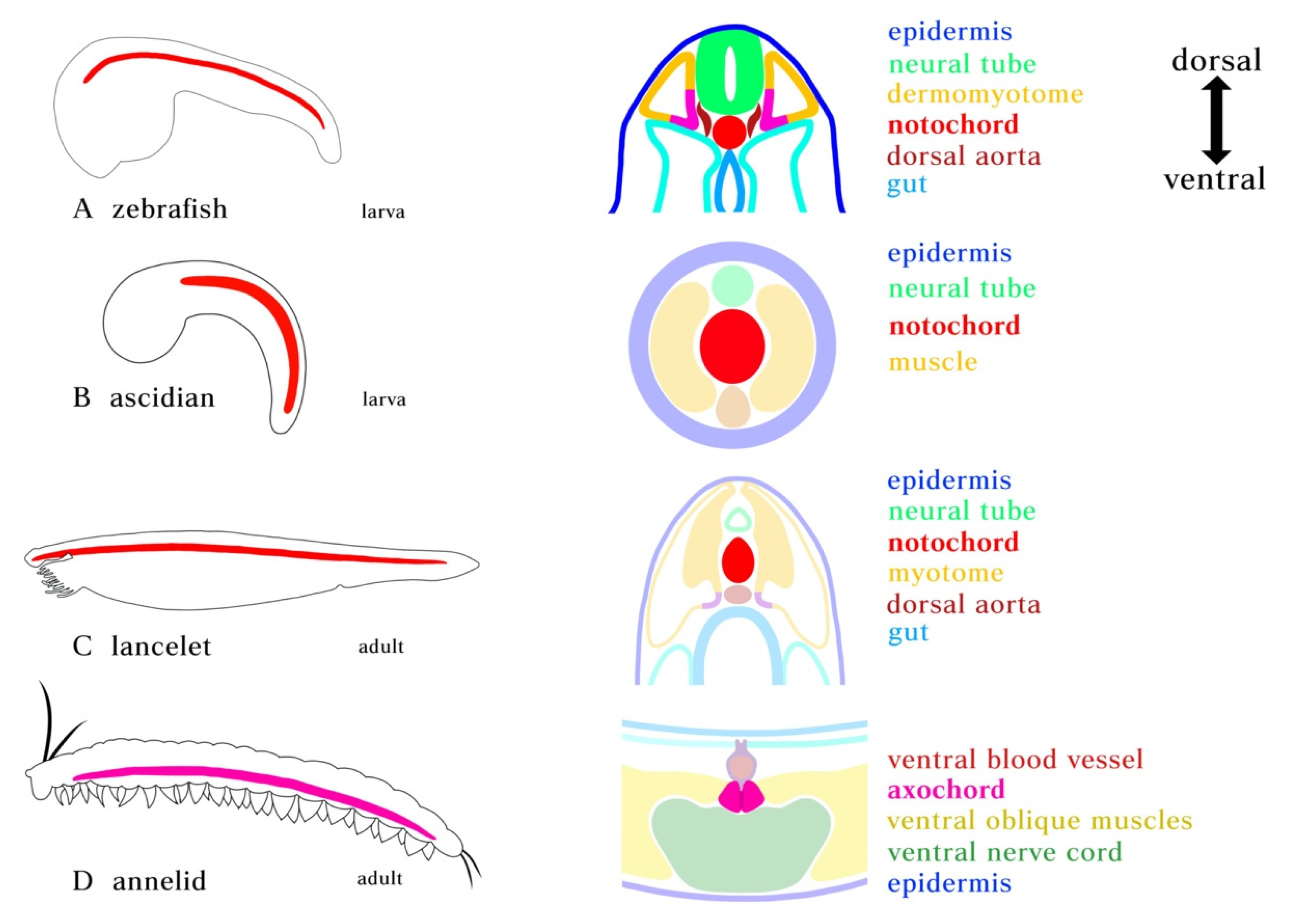
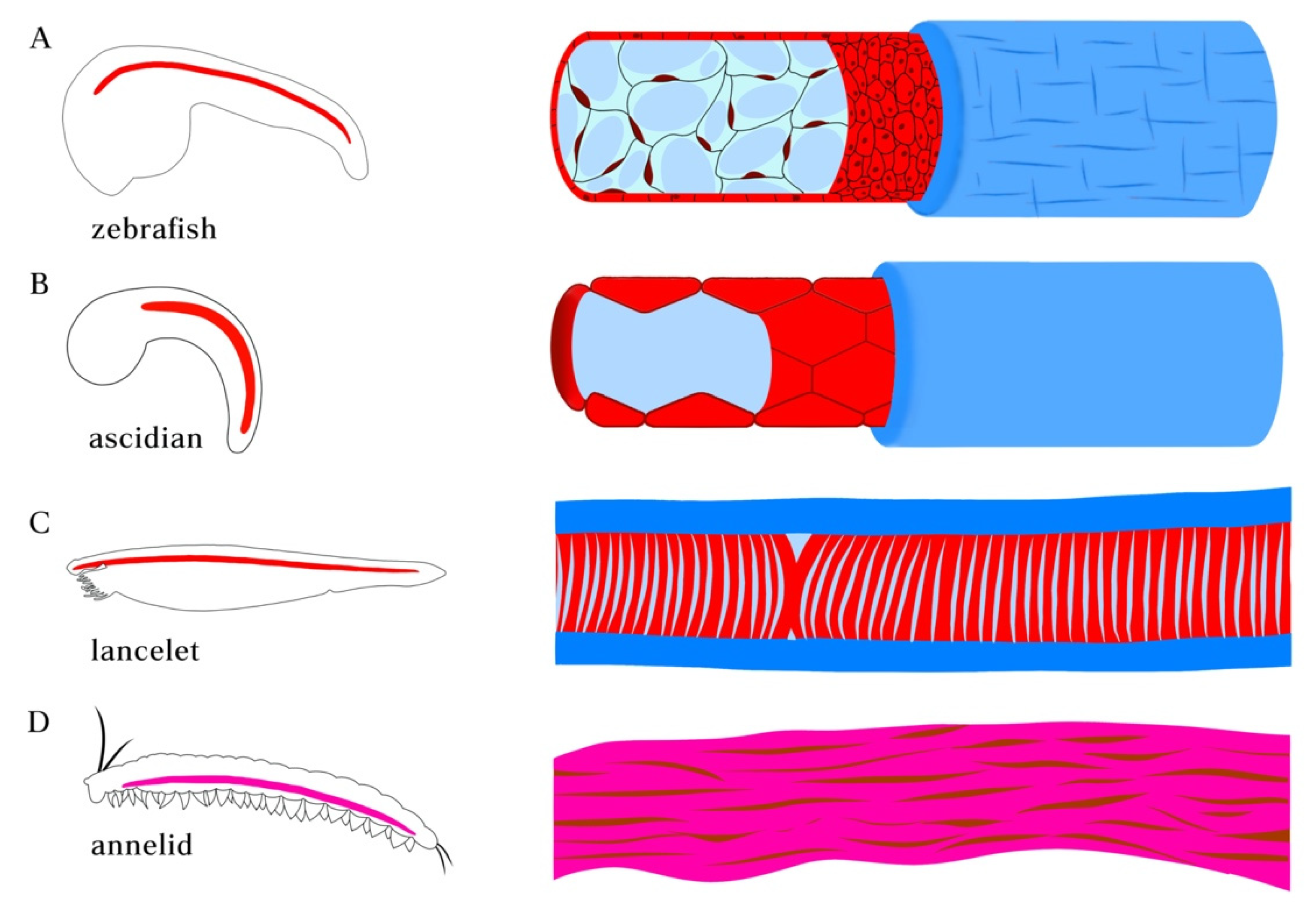
2. Embryonic Development of Notochord-like Structures
3. Molecular Profiles and Cell Types of Notochord-Associated Tissues
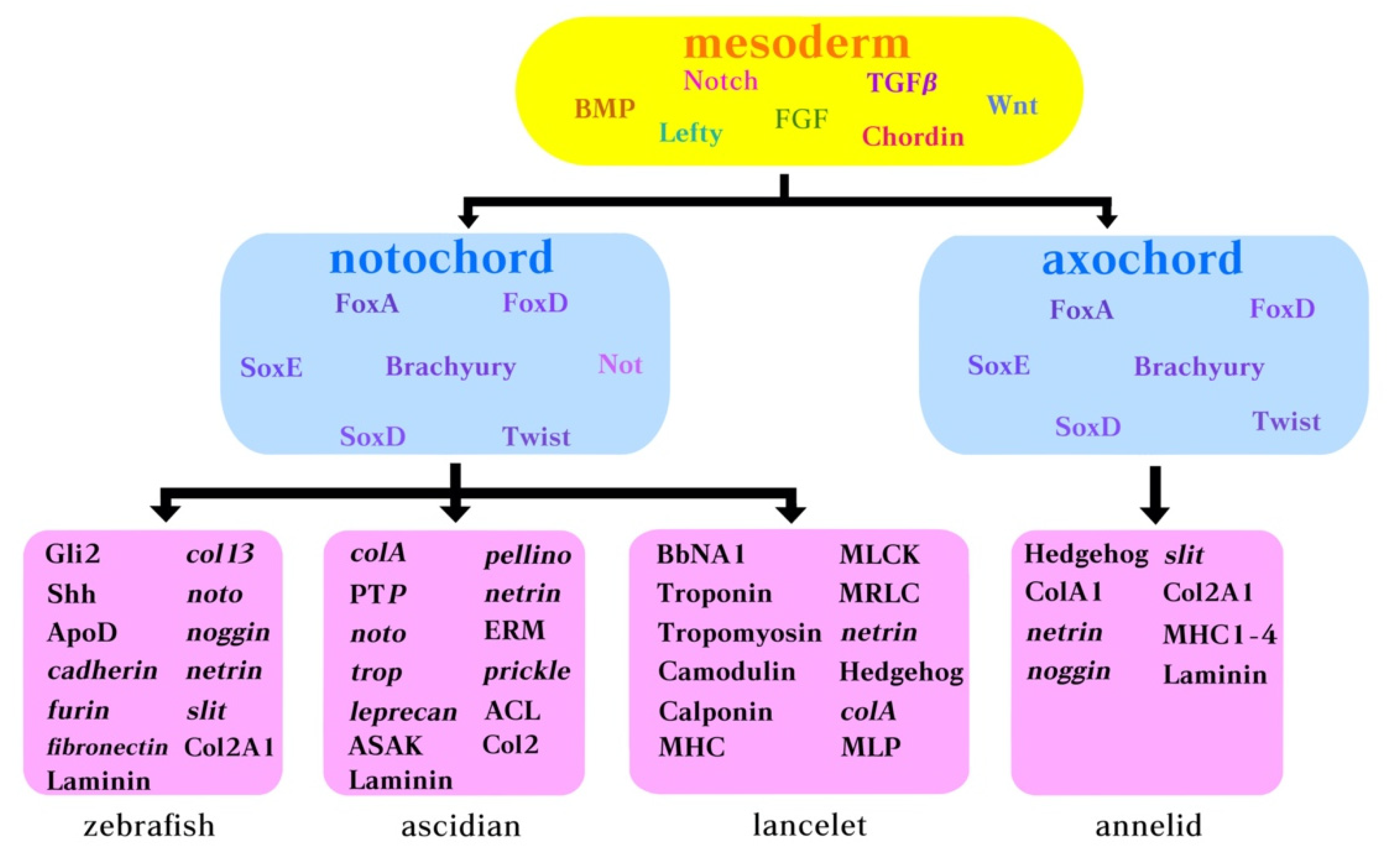
4. Molecular Components Belonging to the Notochord Gene Regulatory Networks (GRNs) in Ascidians, Amphioxus, and Annelids
4.1. Wnt/Beta-Catenin
4.2. FoxA
4.3. Brachyury
4.4. Tbx2/3
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Placzek, M.; Yamada, T.; Tessier-Lavigne, M.; Jessell, T.; Dodd, J. Control of dorsoventral pattern in vertebrate neural development: Induction and polarizing properties of the floor plate. Development 1991, 113 (Suppl. 2), 105–122. [Google Scholar] [CrossRef]
- Peng, H.; Qiao, R.; Dong, B. Polarity Establishment and Maintenance in Ascidian Notochord. Front. Cell Dev. Biol. 2020, 8, 1245. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, H.; Wessely, O.; De Robertis, E.M. Neural induction in Xenopus: Requirement for ectodermal and endomesodermal signals via Chordin, Noggin, β-Catenin, and Cerberus. PLoS Biol. 2004, 2, e92. [Google Scholar] [CrossRef]
- De Robertis, E.M.; Kuroda, H. Dorsal-ventral patterning and neural induction in Xenopus embryos. Annu. Rev. Cell Dev. Biol. 2004, 20, 285–308. [Google Scholar] [CrossRef]
- Smits, P.; Lefebvre, V. Sox5 and Sox6 are required for notochord extracellular matrix sheath formation, notochord cell survival and development of the nucleus pulposus of intervertebral discs. Development 2003, 130, 1135–1148. [Google Scholar] [CrossRef] [PubMed]
- Christ, B.; Huang, R.; Wilting, J. The development of the avian vertebral column. Anat. Embryol. 2000, 202, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Nibu, Y.; José-Edwards, D.S.; Di Gregorio, A. From notochord formation to hereditary chordoma: The many roles of brachyury. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Bateson, W. The Ancestry of the Chordata. Q. J. Microsc. Sci. 1886, 26, 535–571. [Google Scholar]
- Ruppert, E.E. Structure, Ultrastructure and Function of the Neural Gland Complex of Ascidia interrupta (Chordata, Ascidiacea): Clarification of Hypotheses Regarding the Evolution of the Vertebrate Anterior Pituitary. Acta Zool. 1990, 71, 135–149. [Google Scholar] [CrossRef]
- Miyamoto, N.; Wada, H. Hemichordate neurulation and the origin of the neural tube. Nat. Commun. 2013, 4, 2713. [Google Scholar] [CrossRef]
- Satoh, N.; Tagawa, K.; Lowe, C.J.; Yu, J.K.; Kawashima, T.; Takahashi, H.; Ogasawara, M.; Kirschner, M.; Hisata, K.; Su, Y.H.; et al. On a Possible Evolutionary Link of the Stomochord of Hemichordates to Pharyngeal Organs of Chordates. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Gerhart, J.; Lowe, C.; Kirschner, M. Hemichordates and the origin of chordates. Curr. Opin. Genet. Dev. 2005, 15, 461–467. [Google Scholar] [CrossRef]
- Yu, J.K.; Holland, L.Z.; Jamrich, M.; Blitz, I.L.; Holland, N.D. AmphiFoxE4, an amphioxus winged helix/forkhead gene encoding a protein closely related to vertebrate thyroid transcription factor-2: Expression during pharyngeal development. Evol. Dev. 2002, 4, 9–15. [Google Scholar] [CrossRef]
- Lowe, C.J.; Clarke, D.N.; Medeiros, D.M.; Rokhsar, D.S.; Gerhart, J. The deuterostome context of chordate origins. Nature 2015, 520, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Haeckel, E. Anthropogenie oder Entwickelungsgeschichte des Menschen; gemeinverständliche wissenschaftliche Vorträge. Nature 1874, 11, 22–24. [Google Scholar] [CrossRef][Green Version]
- Alexander, K. Embryologische Studien an Würmer und Arthropoden. Mém. Acad. Imp. Sci. St.-Pétersbg. 1871, 16, 1–70. [Google Scholar]
- Dohrn, A. Der Ursprung der Wirbelthiere und das Princip des Functionswechsels: Genealogische Skizzen; W. Engelmann: Leipzig, Germany, 1875. [Google Scholar]
- Semper, C. Die Stammverwandschaft der Wirbelthiere und Wirbellosen. Arb. Zool.-Zootom. Inst. Würzburg 1875, 2, 25–76. [Google Scholar]
- Lauri, A.; Brunet, T.; Handberg-Thorsager, M.; Fischer, A.H.; Simakov, O.; Steinmetz, P.R.; Tomer, R.; Keller, P.J.; Arendt, D. Development of the Annelid Axochord: Insights into notochord evolution. Science 2014, 345, 1365–1368. [Google Scholar] [CrossRef]
- Hejnol, A.; Lowe, C.J. Animal evolution: Stiff or squishy notochord origins? Curr. Biol. 2014, 24, R1131–R1133. [Google Scholar] [CrossRef] [PubMed]
- Brunet, T.; Lauri, A.; Arendt, D. Did the notochord evolve from an ancient axial muscle? The axochord hypothesis. BioEssays 2015, 37, 836–850. [Google Scholar] [CrossRef] [PubMed]
- Metchnikoff, E.; Kovalevsky, A. le développement d l’Amphioxus lanceolatus. Arch. Sci. Phys. Nat. 1866, 27, 193–195. [Google Scholar]
- Kauffman, S. Paramyosin muscle in the notochord of amphioxus. Nature 1969, 224, 177–178. Available online: http://adsabs.harvard.edu/abs/1969Natur.224..177K (accessed on 13 June 2021.). [CrossRef] [PubMed]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Flood, P.R. The connection between spinal cord and notochord in Amphioxus (Branchiostoma lanceolatum). Z. Zellforsch. Mikrosk. Anat. 1970, 103, 115–128. [Google Scholar] [CrossRef]
- Stach, T. The ontogeny of the notochord of Branchiostoma lanceolatum. Acta Zool. 1999, 80, 25–33. [Google Scholar] [CrossRef]
- Hubrecht, A.A.W. On the ancestral form of the Chordata. Q. J. Microsc. Sci. 1883, XXIII, 349–368. [Google Scholar]
- Struck, T.H.; Fisse, F. Phylogenetic position of nemertea derived from phylogenomic data. Mol. Biol. Evol. 2008, 25, 728–736. [Google Scholar] [CrossRef]
- Spindle, S.T. Confocal Microscopy Study of the Embryonic Development of the Viviparous Nemertean Prosorhochmus americanus Reveals Larval Features Supporting Indirect Development in Hoplonemerteans. Master’s Thesis, Virginia Commonwealth University, Richmond, VA, USA, 2013. [Google Scholar]
- Simakov, O.; Kawashima, T.; Marlétaz, F.; Jenkins, J.; Koyanagi, R.; Mitros, T.; Hisata, K.; Bredeson, J.; Shoguchi, E.; Gyoja, F.; et al. Hemichordate genomes and deuterostome origins. Nature 2015, 527, 459–465. [Google Scholar] [CrossRef]
- McMenamin, M.A.S. Cambrian chordates and vetulicolians. Geosciences 2019, 9, 354. [Google Scholar] [CrossRef]
- Ou, Q.; Morris, C.S.; Han, J.; Zhang, Z.; Liu, J.; Chen, A.; Zhang, X.; Shu, D. Evidence for gill slits and a pharynx in Cambrian vetulicolians: Implications for the early evolution of deuterostomes. BMC Biol. 2012, 10, 81. [Google Scholar] [CrossRef]
- Shu, D.G.; Morris, S.C.; Han, J.; Zhang, Z.F.; Liu, J.N. Ancestral echinoderms from the Chengjiang deposits of China. Nature 2004, 430, 422–428. [Google Scholar] [CrossRef]
- Gee, H. On being vetulicolian. Nature 2001, 414, 407–409. [Google Scholar] [CrossRef]
- Satoh, N. Molecular Phylogeny. In Chordate Origins and Evolution; Academic Press: Cambridge, UK, 2016; pp. 45–58. [Google Scholar] [CrossRef]
- Gerhart, J. Evolution of the organizer and the chordate body plan. Int. J. Dev. Biol. 2001, 45, 133–153. [Google Scholar] [CrossRef]
- Sulik, K.; Dehart, D.B.; Inagaki, T.; Carson, J.L.; Vrablic, T.; Gesteland, K.; Schoenwolf, G.C. Morphogenesis of the murine node and notochordal plate. Dev. Dyn. 1994, 201, 260–278. [Google Scholar] [CrossRef]
- Glickman, N.S.; Kimmel, C.B.; Jones, M.A.; Adams, R.J. Shaping the zebrafish notochord. Development 2003, 130, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Bagwell, J.; Norman, J.; Ellis, K.; Peskin, B.; Hwang, J.; Ge, X.; Nguyen, V.S.; McMenamin, K.S.; Stainier, Y.D.; Bagnat, M. Notochord vacuoles absorb compressive bone growth during zebrafish spine formation. Elife 2020, 9, e51221. [Google Scholar] [CrossRef]
- Wopat, S.; Bagwell, J.; Sumigray, K.D.; Dickson, A.L.; Huitema, L.F.; Poss, K.D.; Schulte-Merker, S.; Bagnat, M. Spine Patterning Is Guided by Segmentation of the Notochord Sheath. Cell Rep. 2018, 22, 2026–2038. [Google Scholar] [CrossRef]
- Ellis, K.; Bagwell, J.; Bagnat, M. Notochord vacuoles are lysosome-related organelles that function in axis and spine morphogenesis. J. Cell Biol. 2013, 200, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.; Harfe, B.D. Notochord to Nucleus Pulposus Transition. Curr. Osteoporos. Rep. 2015, 13, 336–341. [Google Scholar] [CrossRef]
- Munro, E.M.; Odell, G. Morphogenetic pattern formation during ascidian notochord formation is regulative and highly robust. Development 2002, 129, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Smith, W.C. Ascidian notochord morphogenesis. Dev. Dyn. 2007, 236, 1748–1757. [Google Scholar] [CrossRef]
- Munro, E.; Robin, F.; Lemaire, P. Cellular morphogenesis in ascidians: How to shape a simple tadpole. Curr. Opin. Genet. Dev. 2006, 16, 399–405. [Google Scholar] [CrossRef]
- Lu, Q.; Bhattachan, P.; Dong, B. Ascidian notochord elongation. Dev. Biol. 2019, 448, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Holland, L.Z.; Onai, T. Early development of cephalochordates (amphioxus). Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Horie, T.; Denker, E.; Kusakabe, T.; Tsuda, M.; Smith, C.W.; Jiang, D. Tube formation by complex cellular processes in Ciona intestinalis notochord. Dev. Biol. 2009, 330, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Balser, E.J.; Ruppert, E.E. Structure, Ultrastructure, and Function of the Preoral Heart-Kidney in Saccoglossus kowalevskii (Hemichordata, Enteropneusta) Including New Data on the Stomochord. Acta Zool. 1990, 71, 235–249. [Google Scholar] [CrossRef]
- Annona, G.; Holland, N.D.; D’Aniello, S. Evolution of the notochord. Evodevo 2015, 6, 30. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.A.; Henrich, T.; Arendt, D. The normal development of Platynereis dumerilii (Nereididae, Annelida) Frontiers in Zoology The normal development of Platynereis dumerilii (Nereididae, Annelida). Front. Zool. 2010, 7, 31. Available online: http://www.frontiersinzoology.com/content/7/1/31%5Cnhttp://www.frontiersinzoology.com/info/instructions/%5Cnhttp://www.biomedcentral.com/ (accessed on 13 June 2021). [CrossRef]
- De Robertis, E.M.; Sasai, Y. A common plan for dorsoventral patterning in Bilateria. Nature 1996, 380, 37–40. [Google Scholar] [CrossRef]
- Arendt, D.; Nübler-Jung, K. Inversion of dorsoventral axis? Nature 1994, 371, 26. [Google Scholar] [CrossRef]
- Su, H.Y.; Chen, C.Y.; Ting, C.H.; Fan, P.T.; Lin, Y.C.; Wang, T.K.; Yu, K.J. BMP controls dorsoventral and neural patterning in indirect-developing hemichordates providing insight into a possible origin of chordates. Proc. Natl. Acad. Sci. USA 2019, 116, 12925–12932. [Google Scholar] [CrossRef]
- Lowe, C.J.; Terasaki, M.; Wu, M.; Freeman, R.M., Jr.; Runft, L.; Kwan, K.; Haigo, S.; Aronowicz, J.; Lander, E.; Gruber, C.; et al. Dorsoventral patterning in hemichordates: Insights into early chordate evolution. PLoS Biol. 2006, 4, 1603–1619. [Google Scholar] [CrossRef]
- Onai, T.; Yu, J.K.; Blitz, I.L.; Cho, K.W.; Holland, L.Z. Opposing Nodal/Vg1 and BMP signals mediate axial patterning in embryos of the basal chordate amphioxus. Dev. Biol. 2010, 344, 377–389. [Google Scholar] [CrossRef]
- Stemple, D.L.; Solnica-Krezel, L.; Zwartkruis, F.; Neuhauss, S.C.; Schier, A.F.; Malicki, J.; Stainier, D.Y.; Abdelilah, S.; Rangini, Z.; Mountcastle-Shah, E.; et al. Mutations affecting development of the notochord in zebrafish. Development 1996, 123, 117–128. [Google Scholar] [CrossRef]
- Dheen, T.; Sleptsova-Friedrich, I.; Xu, Y.; Clark, M.; Lehrach, H.; Gong, Z.; Korzh, V. Zebrafish tbx-c functions during formation of midline structures. Development 1999, 126, 2703–2713. [Google Scholar] [CrossRef]
- Schulte-Merker, S.; Ho, R.K.; Herrmann, B.G.; Nusslein-Volhard, C. The protein product of the zebrafish homologue of the mouse T gene is expressed in nuclei of the germ ring and the notochord of the early embryo. Development 1992, 116, 1021–1032. [Google Scholar] [CrossRef]
- Halpern, M.; Thisse, C.; Ho, K.R.; Thisse, B.; Riggleman, B.; Trevarrow, B.; Weinberg, S.E.; Postlethwait, H.J.; Kimmel, B.C. Cell-autonomous shift from axial to paraxial mesodermal development in zebrafish floating head mutants. Development 1995, 121, 4257–4264. [Google Scholar] [CrossRef]
- Schier, A.F.; Neuhauss, S.C.; Helde, K.A.; Talbot, W.S.; Driever, W. The one-eyed pinhead gene functions in mesoderm and endoderm formation in zebrafish and interacts with no tail. Development 1997, 124, 327–342. [Google Scholar] [CrossRef]
- Stemple, D.L. Structure and function of the notochord: An essential organ for chordate development. Development 2005, 132, 2503–2512. [Google Scholar] [CrossRef]
- Miyamoto, D.M.; Crowther, R.J. Formation of the notochord in living ascidian embryos. J. Embryol. Exp. Morphol. 1985, 86, 1–17. [Google Scholar] [CrossRef]
- Eakin, R.M.; Westfall, J.A. Fine structure of the notochord of amphioxus. J. Cell Biol. 1962, 12, 646–651. [Google Scholar] [CrossRef]
- Mansfield, J.H.; Haller, E.; Holland, N.D.; Brent, A.E. Development of somites and their derivatives in amphioxus, and implications for the evolution of vertebrate somites. Evodevo 2015, 6, 21. [Google Scholar] [CrossRef]
- Suzuki, M.M.; Satoh, N. Genes expressed in the amphioxus notochord revealed by EST analysis. Dev. Biol. 2000, 224, 168–177. [Google Scholar] [CrossRef]
- Bishop, J.J.; Vandergon, T.L.; Green, D.B.; Doeller, J.E.; Kraus, D.W. A High-Affinity Hemoglobin Is Expressed in the Notochord of Amphioxus, Branchiostoma californiense. Biol. Bull. 1998, 195, 255–259. [Google Scholar] [CrossRef]
- Nishida, H. Cell lineage analysis in ascidian embryos by intracellular injection of a tracer enzyme. Dev. Biol. 1987, 121, 526–541. [Google Scholar] [CrossRef]
- Reeves, W.; Thayer, R.; Veeman, M. Anterior-posterior regionalized gene expression in the Ciona notochord. Dev. Dyn. 2014, 243, 612–620. [Google Scholar] [CrossRef]
- Harder, M.J.; Hix, J.; Reeves, W.M.; Veeman, M.T. Ciona Brachyury proximal and distal enhancers have different FGF dose-response relationships. PLoS Genet. 2021, 17, e1009305. [Google Scholar] [CrossRef]
- Arimoto, A.; Tagawa, K. Hedgehog expression during development and regeneration in the hemichordate, Ptychodera flava. Zoolog. Sci. 2015, 32, 33–37. [Google Scholar] [CrossRef]
- Tagawa, K.; Humphreys, T.; Satoh, N. Novel pattern of Brachyury gene expression in hemichordate embryos. Mech. Dev. 1998, 75, 139–143. [Google Scholar] [CrossRef]
- Satoh, N.; Tagawa, K.; Takahashi, H. How was the notochord born? Evol. Dev. 2012, 14, 56–75. [Google Scholar] [CrossRef]
- Di Gregorio, A. The Notochord Gene Regulatory Network in Chordate Evolution: Conservation and Divergence from Ciona to Vertebrates, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 139. [Google Scholar]
- Valenta, T.; Hausmann, G.; Basler, K. The many faces and functions of beta-catenin. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef]
- Guger, K.A.; Gumbiner, B.M. β-Catenin Has Wnt-like Activity and Mimics the Nieuwkoop Signaling Center in Xenopus Dorsal—Ventral Patterning. Dev. Biol. 1995, 125, 115–125. [Google Scholar] [CrossRef]
- Reintsch, W.E.; Habring-Mueller, A.; Wang, R.W.; Schohl, A.; Fagotto, F. β-Catenin controls cell sorting at the notochord-somite boundary independently of cadherin-mediated adhesion. J. Cell Biol. 2005, 170, 675–686. [Google Scholar] [CrossRef]
- Heasman, J.; Crawford, A.; Goldstone, K.; Garner-Hamrick, P.; Gumbiner, B.; McCrea, P.; Kintner, C.; Noro, C.Y.; Wylie, C. Overexpression of Cadherins and Underexpression of B-Catenin Inhibit Dorsal Mesoderm Induction in Early Xenopus Embryos. Cell 1994, 79, 791–803. [Google Scholar] [CrossRef]
- Huelsken, J.; Vogel, R.; Brinkmann, V.; Erdmann, B.; Birchmeier, C.; Birchmeier, W. Requirement for β-Catenin in Anterior-Posterior Axis Formation in Mice. J. Cell Biol. 2000, 148, 567–578. [Google Scholar] [CrossRef]
- Haegel, H.; Larue, L.; Ohsugi, M.; Fedorov, L.; Herrenknecht, K.; Kemler, R. Lack of β-catenin affects mouse development at gastrulation. Development 1995, 121, 3529–3537. [Google Scholar] [CrossRef] [PubMed]
- Onai, T. Canonical Wnt/β-catenin and Notch signaling regulate animal/vegetal axial patterning in the cephalochordate amphioxus. Evol. Dev. 2019, 21, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.J.; Stappert, J.; Bauer, A.; Kispert, A.; Herrmann, B.G.; Kemler, R. Brachyury is a target gene of the Wnt/β-catenin signaling pathway. Mech. Dev. 2000, 91, 249–258. [Google Scholar] [CrossRef]
- Ikeda, T.; Satou, Y. Differential temporal control of Foxa.a and Zic-r.b specifies brain versus notochord fate in the ascidian embryo. Development 2017, 144, 38–43. [Google Scholar] [CrossRef]
- José-Edwards, D.S.; Oda-Ishii, I.; Nibu, Y.; di Gregorio, A. Tbx2/3 is an essential mediator within the brachyury gene network during Ciona notochord development. Development 2012, 140, 2422–2433. [Google Scholar] [CrossRef]
- Imai, K.S.; Satoh, N.; Satou, Y. An essential role of a FoxD gene in notochord induction in Ciona embryos. Development 2002, 129, 3441–3453. [Google Scholar] [CrossRef] [PubMed]
- Yagi, K.; Satou, Y.; Satoh, N. A zinc finger transcription factor, ZicL, is a direct activator of Brachyury in the notochord specification of Ciona intestinalis. Development 2004, 131, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Ettensohn, C.A. Encoding anatomy: Developmental gene regulatory networks and morphogenesis. Genesis 2013, 51, 383–409. [Google Scholar] [CrossRef] [PubMed]
- Passamaneck, Y.J.; Katikala, L.; Perrone, L.; Dunn, M.P.; Oda-Ishii, I.; di Gregorio, A. Direct activation of a notochord cis-regulatory module by Brachyury and FoxA in the ascidian Ciona intestinalis. Development 2009, 136, 3679–3689. [Google Scholar] [CrossRef]
- Dal-Pra, S.; Thisse, C.; Thisse, B. FoxA transcription factors are essential for the development of dorsal axial structures. Dev. Biol. 2011, 350, 484–495. [Google Scholar] [CrossRef]
- Aldea, D.; Leon, A.; Bertrand, S.; Escriva, H. Expression of Fox genes in the cephalochordate Branchiostoma lanceolatum. Front. Ecol. Evol. 2015, 3, 80. [Google Scholar] [CrossRef]
- Hiruta, J.; Mazet, F.; Yasui, K.; Zhang, P.; Ogasawara, M. Comparative expression analysis of transcription factor genes in the endostyle of invertebrate chordates. Dev. Dyn. 2005, 233, 1031–1037. [Google Scholar] [CrossRef]
- Fritzenwanker, J.H.; Gerhart, J.; Freeman, R.M.; Lowe, C.J. The Fox/Forkhead transcription factor family of the hemichordate Saccoglossus kowalevskii. Evodevo 2014, 5, 17. [Google Scholar] [CrossRef]
- Kostyuchenko, R.P.; Kozin, V.V.; Filippova, N.A.; Sorokina, E.V. FoxA expression pattern in two polychaete species, Alitta virens and Platynereis dumerilii: Examination of the conserved key regulator of the gut development from cleavage through larval life, postlarval growth, and regeneration. Dev. Dyn. 2019, 248, 728–743. [Google Scholar] [CrossRef]
- Yasuo, H.; Satoh, N. Conservation of the developmental role of Brachyury in notochord formation in a Urochordate, the ascidian Halocynthia roretzi. Dev. Biol. 1998, 200, 158–170. [Google Scholar] [CrossRef]
- Scholz, C.B.; Technau, U. The ancestral role of Brachyury: Expression of NemBra1 in the basal cnidarian Nematostella vectensis (Anthozoa). Dev. Genes Evol. 2003, 212, 563–570. [Google Scholar] [CrossRef]
- Satoh, G.; Harada, Y.; Satoh, N. The expression of nonchordate deuterostome Brachyury genes in the ascidian Ciona embryo can promote the differentiation of extra notochord cells. Mech. Dev. 2000, 96, 155–163. [Google Scholar] [CrossRef]
- Technau, U. Brachyury, the blastopore and the evolution of the mesoderm. BioEssays 2001, 23, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Nieuwkoop, P.D. The formation of the mesoderm in urodelean amphibians—I. Induction by the endoderm. Wilhelm Roux. Arch. Entwickl. Mech. Org. 1969, 162, 341–373. [Google Scholar] [CrossRef]
- Cunliffe, V.; Smith, J.C. Ectopic mesoderm formation in Xenopus embryos caused by widespread expression of a Brachyury homologue. Nature 1992, 358, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Hotta, K.; Takahashi, H.; Asakura, T.; Saitoh, B.; Takatori, N.; Satou, Y.; Satoh, N. Characterization of Brachyury-downstream notochord genes in the Ciona intestinalis embryo. Dev. Biol. 2000, 224, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.M.; Wu, Y.; Harder, M.J.; Veeman, M.T. Functional and evolutionary insights from the Ciona notochord transcriptome. Development 2017, 144, 3375–3387. [Google Scholar] [CrossRef] [PubMed]
- Arendt, D.; Technau, U.; Wittbrodt, J. Evolution of the bilaterian larval foregut. Nature 2001, 409, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Arenas-Mena, C. Brachyury, Tbx2/3 and sall expression during embryogenesis of the indirectly developing polychaete Hydroides elegans. Int. J. Dev. Biol. 2013, 57, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Horton, A.C.; Mahadevan, N.R.; Minguillon, C.; Osoegawa, K.; Rokhsar, D.S.; Ruvinsky, I.; de Jong, P.J.; Logan, M.P.; Gibson-Brown, J.J. Conservation of linkage and evolution of developmental function within the Tbx2/3/4/5 subfamily of T-box genes: Implications for the origin of vertebrate limbs. Dev. Genes Evol. 2008, 218, 613–628. [Google Scholar] [CrossRef] [PubMed]
- He, M.L.; Wen, L.; Campbell, C.E.; Wu, J.Y.; Rao, Y. Transcription repression by Xenopus ET and its human ortholog TBX3, a gene involved in ulnar-mammary syndrome. Proc. Natl. Acad. Sci. USA 1999, 96, 10212–10217. [Google Scholar] [CrossRef] [PubMed]
- Hayata, T.; Eisaki, A.; Kuroda, H.; Asashima, M. Expression of Brachyury-like T-box transcription factor Xbra3 in Xenopus embryo. Dev. Genes Evol. 1999, 209, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Takabatake, Y.; Takabatake, T.; Takeshima, K. Conserved and divergent expression of T-box genes Tbx2-Tbx5 in Xenopus. Mech. Dev. 2000, 91, 433–437. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sui, Z.; Zhao, Z.; Dong, B. Origin of the Chordate Notochord. Diversity 2021, 13, 462. https://doi.org/10.3390/d13100462
Sui Z, Zhao Z, Dong B. Origin of the Chordate Notochord. Diversity. 2021; 13(10):462. https://doi.org/10.3390/d13100462
Chicago/Turabian StyleSui, Zihao, Zhihan Zhao, and Bo Dong. 2021. "Origin of the Chordate Notochord" Diversity 13, no. 10: 462. https://doi.org/10.3390/d13100462
APA StyleSui, Z., Zhao, Z., & Dong, B. (2021). Origin of the Chordate Notochord. Diversity, 13(10), 462. https://doi.org/10.3390/d13100462