
Lipid and Fatty Acids Accumulation Features of Entomoneis cf. paludosa during Exponential and Stationary Growth Phases in Laboratory Culture

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algae Cultivation
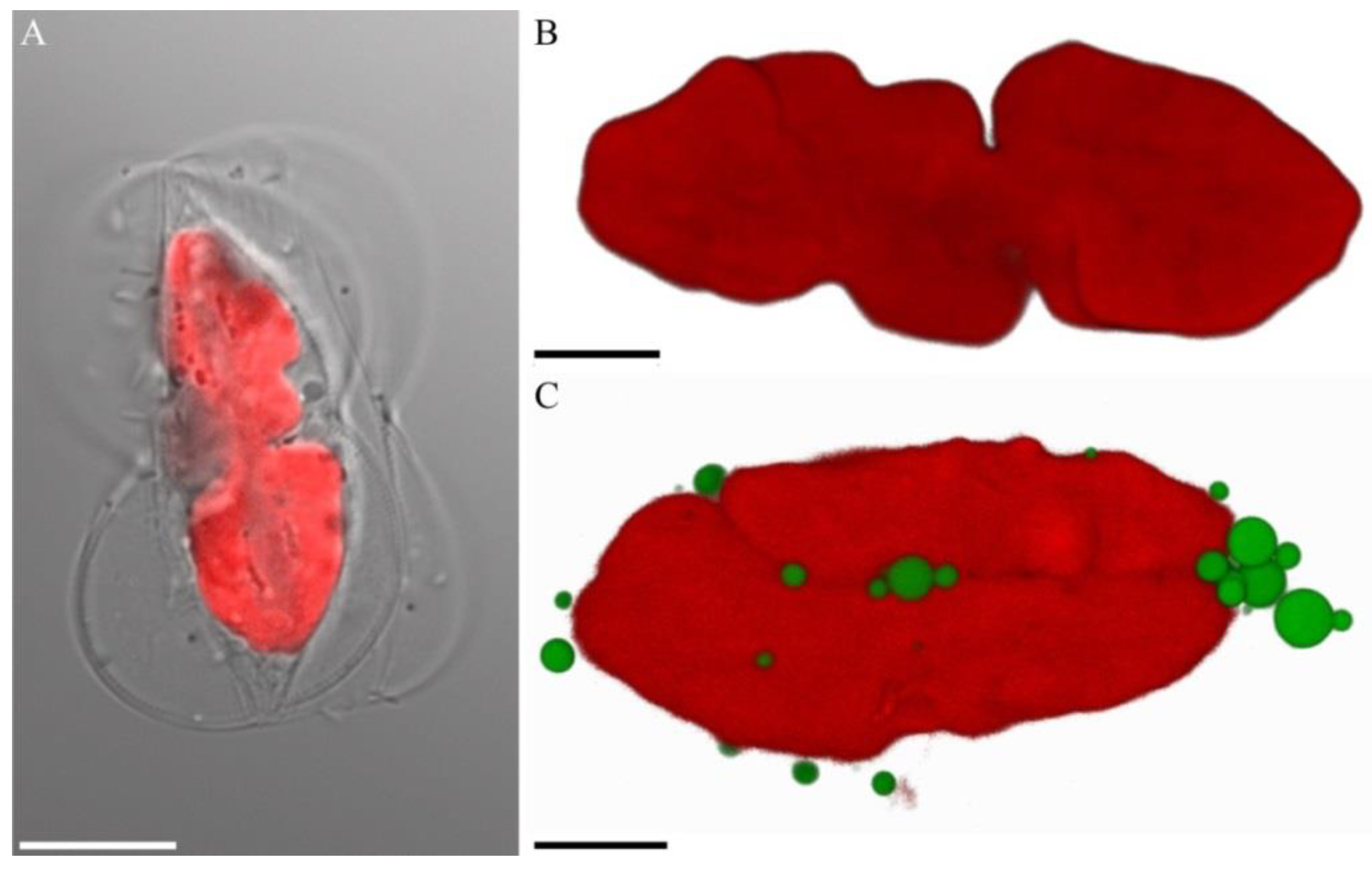
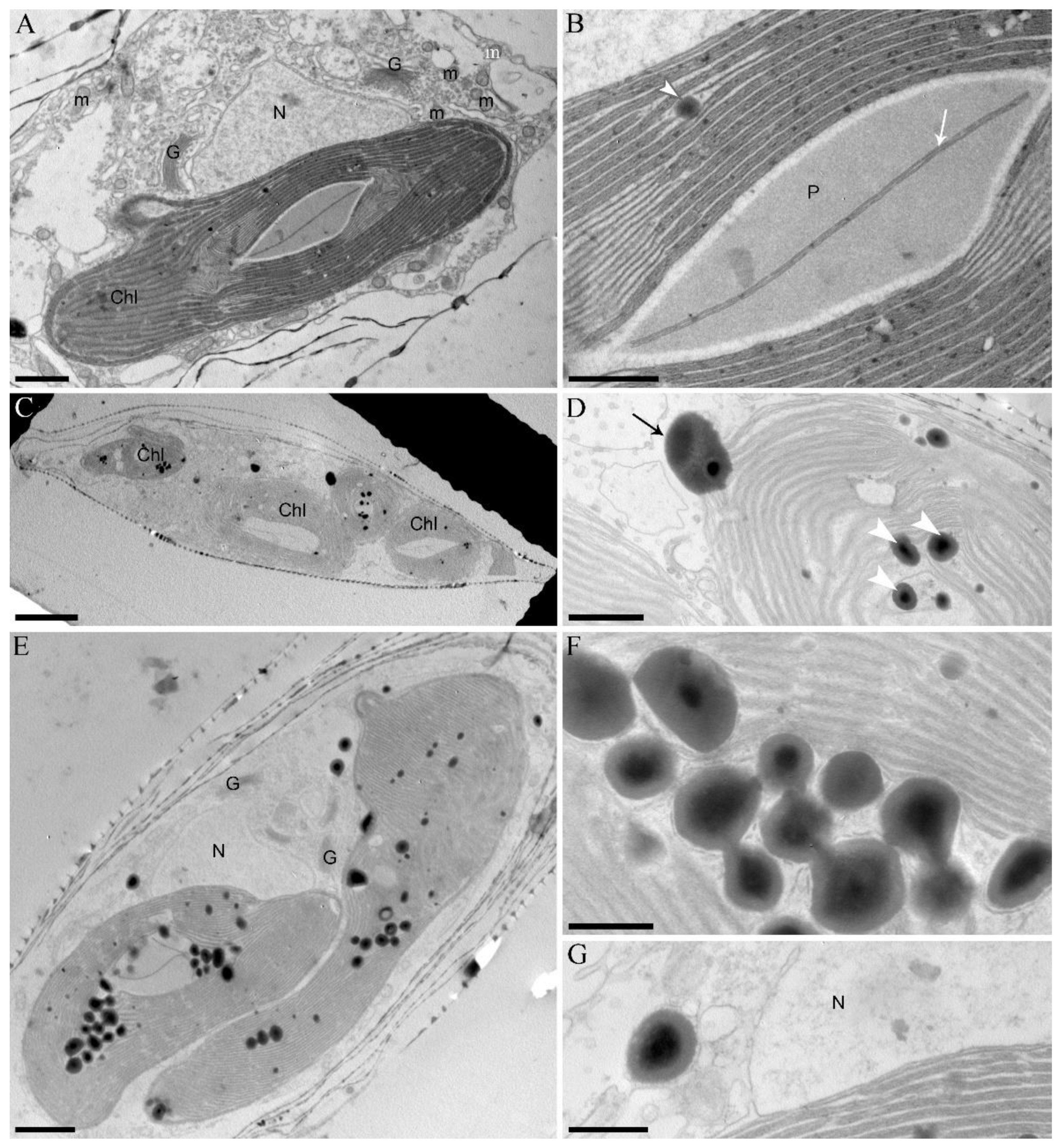
2.2. Microscopy
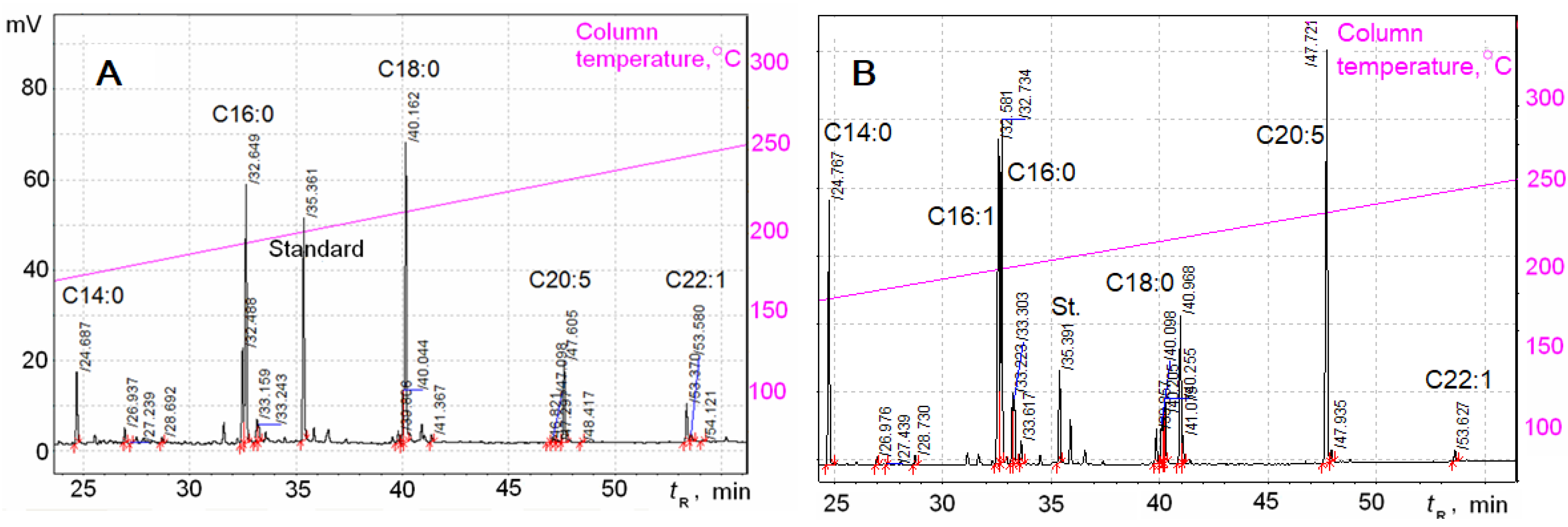
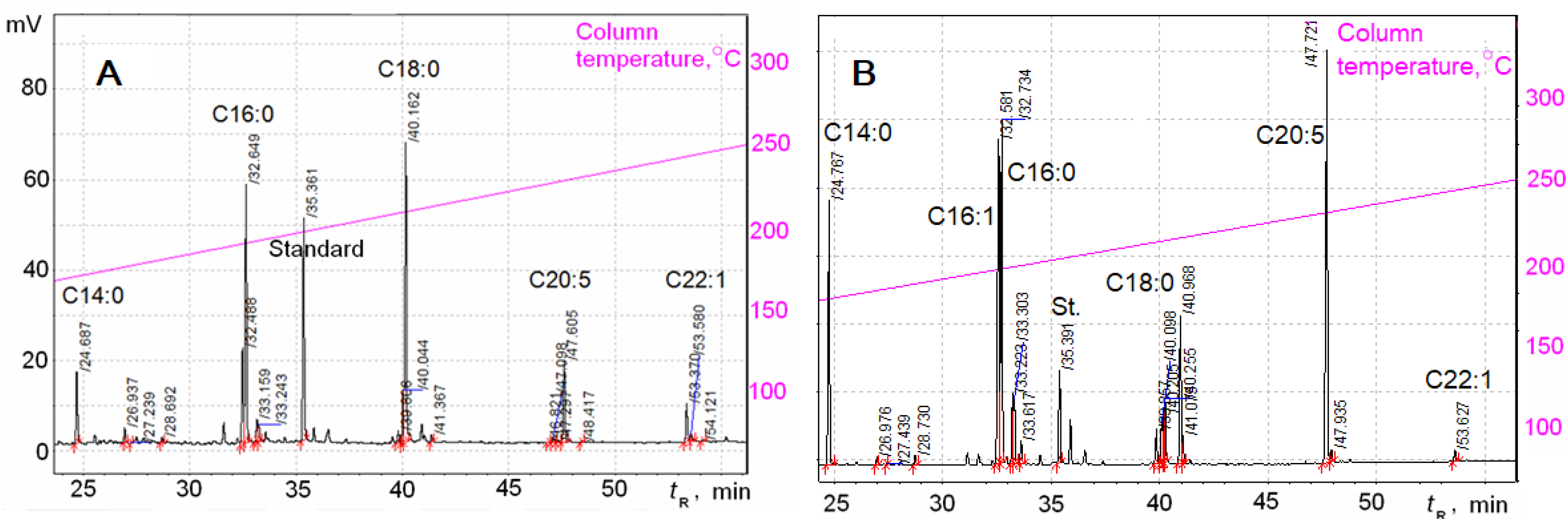
2.3. Fatty Acid Analysis
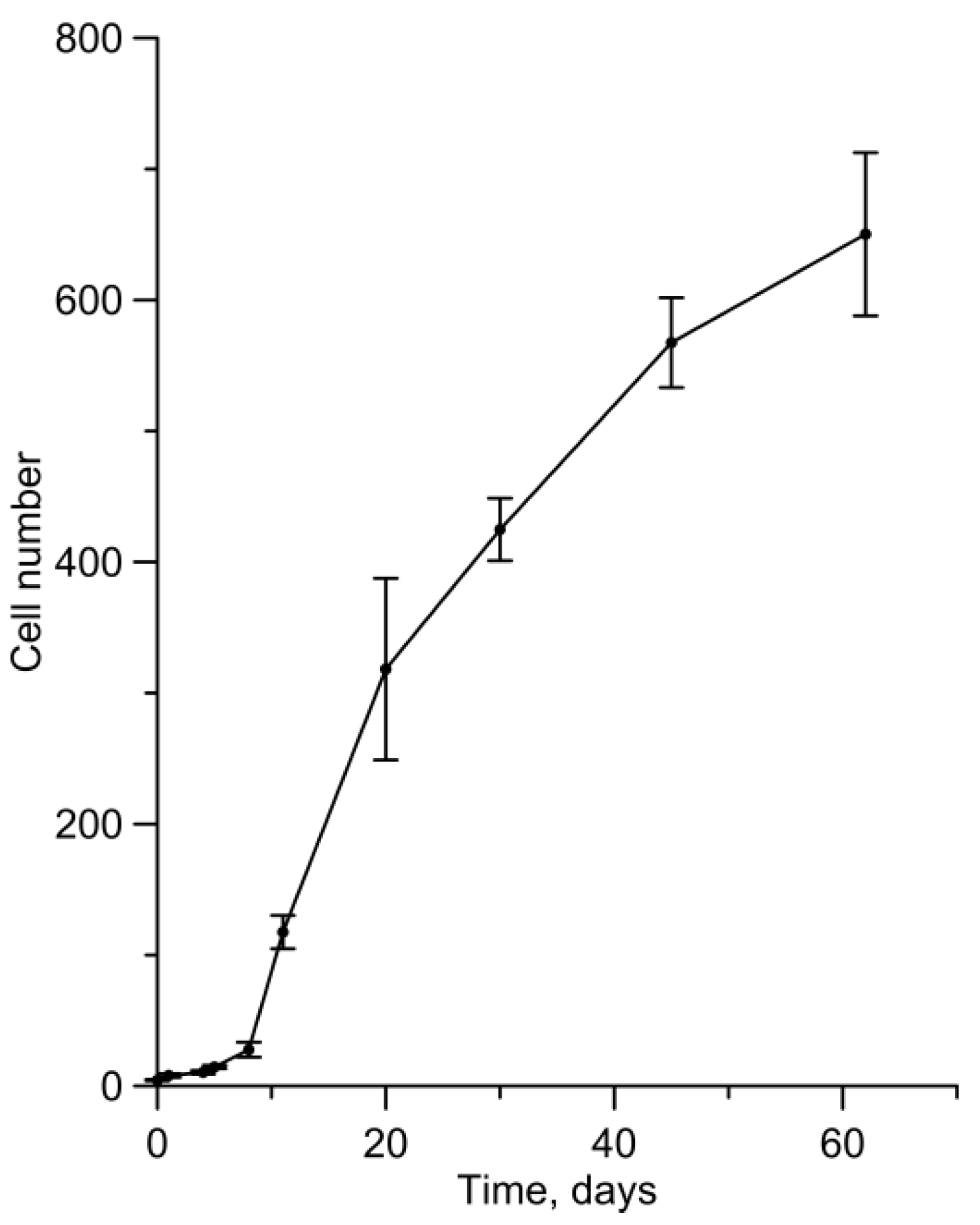
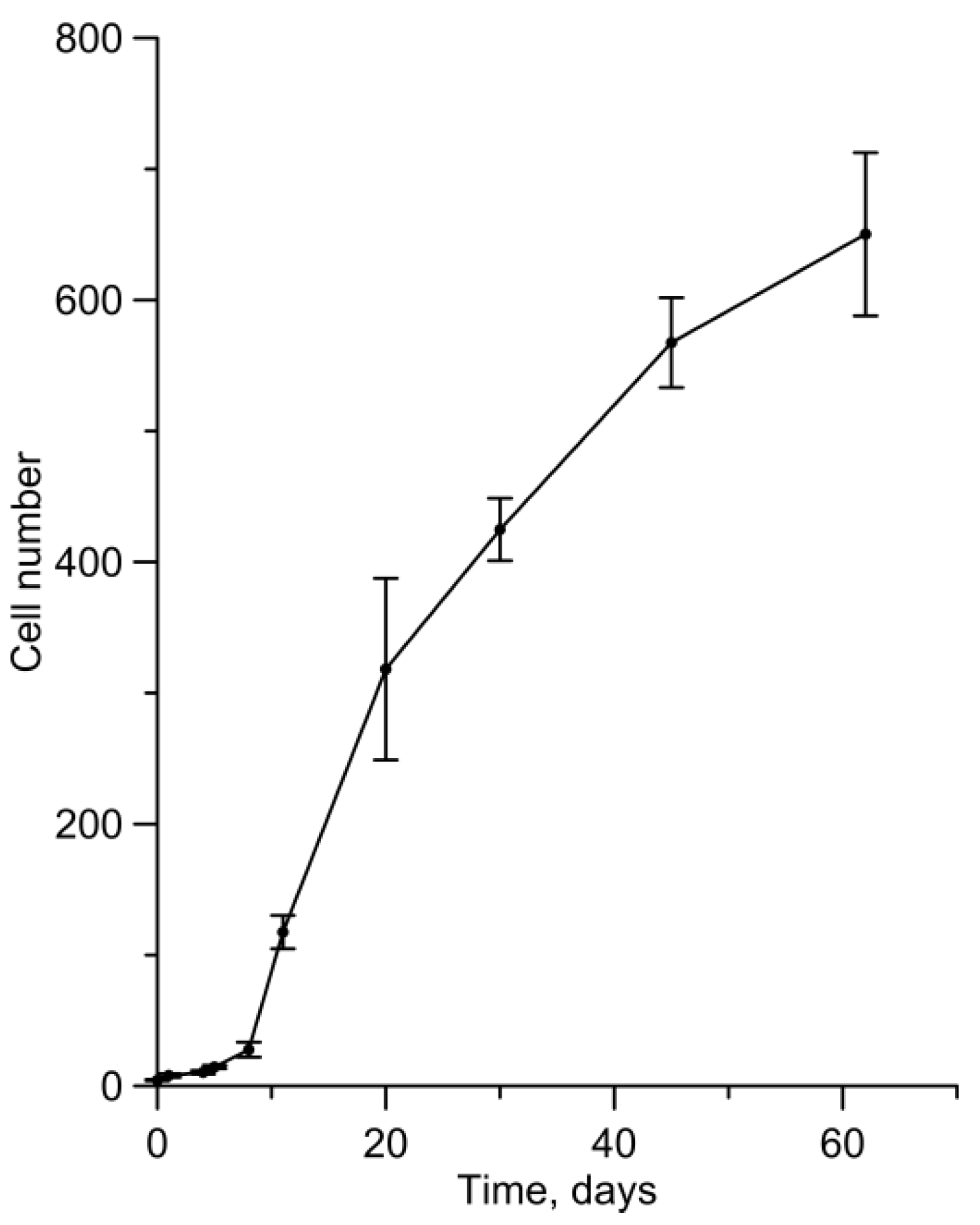
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ragueneau, O.; Treguer, P.; Leynaert, A.; Anderson, R.F.; Brzezinski, M.A.; DeMaster, D.J.; Dugdale, R.C.; Dymond, J.; Fischer, G.; Francois, R.; et al. A Review of the Si cycle in the modern ocean: Recent progress and missing gaps in the application of biogenic opal as a paleoproductivity proxy. Glob. Planet. Chang. 2000, 26, 317–365. [Google Scholar] [CrossRef]
- d’Ippolito, G.; Sardo, A.; Paris, D.; Vella, F.M.; Adelfi, M.G.; Botte, P.; Gallo, C.; Fontana, A. Potential of lipid metabolism in marine diatoms for biofuel production. Biotechnol. Biofuels 2015, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Maeda, Y.; Yoshino, T.; Matsumoto, M.; Tanaka, T. Profiling of fatty acid methyl esters from the oleaginous diatom Fistulifera sp. strain JPCC DA0580 under nutrition-sufficient and -deficient conditions. J. Appl. Phycol. 2014, 26, 2295–2302. [Google Scholar] [CrossRef]
- Zulu, N.N.; Zienkiewicz, K.; Vollheyde, K.; Feussner, I. Current trends to comprehend lipid metabolism in diatoms. Prog. Lipid Res. 2018, 70, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lupette, J.; Jaussaud, A.; Seddiki, K.; Morabito, C.; Brugière, S.; Schaller, H.; Kuntz, M.; Putaux, J.-L.; Jouneau, P.-H.; Rébeillé, F.; et al. The architecture of lipid droplets in the diatom Phaeodactylum tricornutum. Algal Res. 2019, 38, 101415. [Google Scholar] [CrossRef]
- Goold, H.; Beisson, F.; Peltier, G.; Li-Beisson, Y. Microalgal lipid droplets: Composition, diversity, biogenesis and functions. Plant Cell Rep. 2015, 34, 545–555. [Google Scholar] [CrossRef]
- Zienkiewicz, K.; Du, Z.Y.; Ma, W.; Vollheyde, K.; Benning, C. Stress-induced neutral lipid biosynthesis in microalgae: Molecular, cellular and physiological insights. Biochim. Biophys. Acta 2016, 1861, 1269–1281. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.K.; Niu, Y.F.; Ma, Y.H.; Xue, J.; Zhang, M.H.; Yang, W.D.; Liu, J.S.; Lu, S.H.; Guan, Y.; Li, H.Y. Molecular and cellular mechanisms of neutral lipid accumulation in diatom following nitrogen deprivation. Biotechnol. Biofuels 2013, 6, 67. [Google Scholar] [CrossRef] [Green Version]
- Abida, H.; Dolch, L.J.; Meï, C.; Villanova, V.; Conte, M.; Block, M.A.; Finazzi, G.; Bastien, O.; Tirichine, L.; Bowler, C.; et al. Membrane glycerolipid remodeling triggered by nitrogen and phosphorus starvation in Phaeodactylum tricornutum. Plant Physiol. 2015, 167, 118–136. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.R.; Gle, C.; Abbriano, R.M.; Traller, J.C.; Davis, A.; Trentacoste, E.; Vernet, M.; Allen, A.E.; Hildebrand, M. Transcript level coordination of carbon pathways during silicon starvation-induced lipid accumulation in the diatom Thalassiosira pseudonana. New Phytol. 2016, 210, 890–904. [Google Scholar] [CrossRef]
- Cointet, E.; Wielgosz-Collin, G.; Bougaran, G.; Rabesaotra, V.; Gonçalves, O.; Méléder, V. Effects of light and nitrogen availability on photosynthetic efficiency and fatty acid content of three original benthic diatom strains. PLoS ONE 2019, 14, e0224701. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Nojima, D.; Yoshino, T.; Tanaka, T. Structure and properties of oil bodies in diatoms. Phil. Trans. R. Soc. B 2017, 372, 20160408. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, A.D.; Leech, R.M.; Williams, J.P. The osmiophilic globules of chloroplasts: I. Osmiophilic globules as a normal component of chloroplasts and their iso-lation and composition in Viciafaba L. Biochim. Biophys. Acta 1963, 78, 148–162. [Google Scholar] [CrossRef]
- Austin, J.R.I.; Frost, E.; Vidi, P.-A.; Kessler, F.; Staehelin, L.A. Plastoglobules are lipoprotein subcompartments of the chloroplast that are permanently coupled to thylakoid membranes and contain biosynthetic enzymes. Plant Cell 2006, 18, 1693–1703. [Google Scholar] [CrossRef] [Green Version]
- Bréhélin, C.; Kessler, F.; Van Wijk, K.J. Plastoglobules: Versatile lipoprotein particles in plastids. Trends Plant Sci. 2007, 12, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Piller, E.L.; Abraham, M.; Dormann, P.; Kessler, F.; Besagni, C. Plastid lipid droplets at the crossroads of prenylquinone metabolism. J. Exp. Bot. 2012, 63, 1609–1618. [Google Scholar] [CrossRef]
- Rottet, S.; Besagni, C.; Kessler, F. The role of plastoglobules in thylakoid lipid remodeling during plant development. Biochim. Biophys. Acta 2015, 1847, 889–899. [Google Scholar] [CrossRef] [Green Version]
- Round, F.; Crawford, R.; Mann, D. The diatoms: Biology and Morphology of the Genera; Cambridge University Press: Bath, UK, 1990. [Google Scholar]
- Lohscheider, J.N.; Bártulos, C.R. Plastoglobules in algae: A comprehensive comparative study of the presence of major structural and functional components in complex plastids. Mar. Genom. 2016, 28, 127–136. [Google Scholar] [CrossRef]
- Podunay, Y.A.; Davidovich, N.A.; Davidovich, O.I.; Witkowski, A.; Gastineau, R.; Solak, C.N. The sexual reproduction and life cycle of the pennate diatom Entomoneis cf. paludosa (W. Smith) Reimer (Bacillariophyta). Russ. J. Mar. Biol. 2021, 47, 19–28. [Google Scholar] [CrossRef]
- Davidovich, N.A.; Davidovich, O.I.; Podunay, Y.A. Diatom culture collection of the Karadag scientific station (Crimea). Mar. Biol. J. 2017, 2, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Andersen, R.A.; Berges, J.A.; Harrison, P.J.; Watanabe, M.M. Appendix A—Recipes for Freshwater and Seawater Media. In Algal Culturing Techniques; Elsevier: Amsterdam, The Netherlands, 2005; pp. 429–538. [Google Scholar]
- Polyakova, S.L.; Davidovich, O.I.; Podunai, Y.A.; Davidovich, N.A. Modification of the ESAW culture medium used for cultivation of marine diatoms. Morsk. Biol. Zhurnal 2018, 3, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Safonova, T.A.; Aslamov, I.A.; Basharina, T.N.; Chenski, A.G.; Vereschagin, A.L.; Glyzina, O.Y.; Grachev, M.A. Cultivation and automatic counting of diatom algae cells in multi-well plastic plates. Diatom Res. 2007, 22, 189–195. [Google Scholar] [CrossRef]
- Bedoshvili, Y.D.; Likhoshway, Y.V. The cell ultrastructure of diatoms—Implications for phylogeny? In The Transmission Electron Microscope; Maaz, K., Ed.; InTech: Rijeka, Chroatia, 2012; pp. 147–160. [Google Scholar]
- Nikonova, A.A.; Shishlyannikov, S.M.; Shishlyannikova, T.A.; Avezova, T.N.; Babenko, T.A.; Belykh, O.I.; Glyzina, O.Y.; Obolkin, V.A.; Pavlova, O.N.; Smagunova, A.N.; et al. Determination of free and esterified fatty acids in hydrocoles of different content of polyunsaturated fatty acids by gas–liquid chromatography. J. Anal. Chem. 2020, 75, 1310–1321. [Google Scholar] [CrossRef]
- Smagunova, A.N. Metody matematicheskoi statistiki v analiticheskoi khimii. In Methods of Mathematical Statistics in Analytical Chemistry; Gosudarstvennyi Universitet: Rostov-na-Donu, Russia, 2008. (In Russian) [Google Scholar]
- Bedoshvili, Y.D.; Popkova, T.P.; Likhoshway, Y.V. Chloroplast structure of diatoms of different classes. Cell Tissue Biol. 2009, 3, 297–310. [Google Scholar] [CrossRef]
- Flori, S.; Jouneau, P.H.; Bailleul, B.; Gallet, B.; Estrozi, L.F.; Moriscot, C.; Bastien, O.; Eicke, S.; Schober, A.; Bártulos, C.R.; et al. Plastid thylakoid architecture optimizes photosynthesis in diatoms. Nat. Commun. 2017, 8, 15885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonon, T.; Harvey, D.; Larson, T.R.; Graham, I.A. Long chain polyunsaturated fatty acid production and partitioning to triacylglycerols in four microalgae. Phytochemistry 2002, 61, 15–24. [Google Scholar] [CrossRef]
- Artamonova, E.Y.; Vasskog, T.; Eilertsen, H.C. Lipid content and fatty acid composition of Porosira glacialis and Attheya longicornis in response to carbon dioxide (CO2) aeration. PLoS ONE 2017, 12, e0177703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budge, S.M.; Parrish, C.C.; McKenzie, C.H. Fatty acid composition of phytoplankton, settling particulate matter and sediments at a sheltered bivalve aquaculture site. Mar. Chem. 2001, 76, 285–303. [Google Scholar] [CrossRef]
- Abedi, E.; Sahari, A.M. Long-chain polyunsaturated fatty acid sources and evaluation of their nutrition and functional properties. Review. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.C. Essential fatty acids in aquatic food webs. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M., Eds.; Springer: New York, NY, USA, 2009; pp. 309–326. [Google Scholar] [CrossRef]
- Le, P.N.T.; Desbois, A.P. Antibacterial effect of eicosapentaenoic acid against Bacillus cereus and Staphylococcus aureus: Killing kinetics, selection for resistance, and potential cellular target. Mar. Drugs 2017, 15, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, Z.; Xu, M.; Di, X.; Brynjolfsson, S.; Fu, W. Exploring valuable lipids in diatoms. Front. Mar. Sci. 2017, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Xu, J.; Yan, X.; Zhao, P.; Chen, J.; Zhou, C.; Zhao, F.; Li, S. Lipidomic changes during different growth stages of Nitzschia closterium f. minutissima. Metabolomics 2013, 9, 300–310. [Google Scholar] [CrossRef]
- Pasquet, V.; Ulmann, L.; Mimouni, V.; Guihéneuf, F.; Jacquette, B.; Morant-Manceau, A.; Tremblin, G. Fatty acids profile and temperature in the cultured marine diatom Odontella aurita. J. Appl. Phycol. 2014, 26, 2265–2271. [Google Scholar] [CrossRef]
- Tokushima, H.; Inoue-Kashino, N.; Nakazato, Y.; Masuda, A.; Ifuku, K.; Kashino, Y. Advantageous characteristics of the diatom Chaetoceros gracilis as a sustainable biofuel producer. Biotechnol. Biofuels 2016, 9, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Marella, T.K.; Tao, L.; Li, R.; Tiwari, A.; Li, G. Optimization of growth conditions and fatty acid analysis for three freshwater diatom isolates. Phycol. Res. 2017, 65, 177–187. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FAs Nos | FAs Structure | Retention Times (tR), Min | FAs Content at Two Phases of Growth | |||
|---|---|---|---|---|---|---|
| Stationary | Exponential | |||||
| mg g−1 | % | mg g−1 | % | |||
| 1 | C12:0 | 17.68 | 4.16 ± 0.4 | 10.47 | 0.143 ± 0.013 | 0.9 |
| 2 | C14:0 | 24.69 | 2.46 ± 0.22 | 6.27 | 2.36 ± 0.21 | 14.74 |
| 3 | iso-C15:0 | 26.94 | 0.49 ± 0.04 | 1.25 | 0.071 ± 0.006 | 0.45 |
| 4 | aiso-C15:0 | 27.24 | 0.064 ± 0.006 | 0.16 | 0.0047 ± 0.0004 | 0.03 |
| 5 | C15:0 | 28.69 | 0.198 ± 0.018 | 0.5 | 0.087 ± 0.008 | 0.54 |
| 6 | C16:1 (9) | 32.49 | 3.1 ± 0.3 | 7.9 | 3.04 ± 0.27 | 19.01 |
| 7 | C16:0 | 32.65 | 10.2 ± 0.9 | 26 | 3.25 ± 0.29 | 20.33 |
| 8 | C16:3 (n-6,9,12) | 33.16 | 0.50 ± 0.04 | 1.29 | 0.37 ± 0.03 | 2.28 |
| 9 | C16:2 (n-9,12) | 33.24 | 0.37 ± 0.03 | 0.95 | 0.32 ± 0.03 | 2.02 |
| 10 | C16:4 (n-6,9,12,15) | 33.56 | 0.13 ± 0.12 | 0.34 | 0.12 ± 0.011 | 0.75 |
| 11 | C18:1 (13) | 39.81 | 0.86 ± 0.08 | 2.13 | 0.293 ± 0.026 | 1.83 |
| 12 | C18:1 (9) | 40.04 | 1.30 ± 0.12 | 3.34 | 0.36 ± 0.03 | 2.23 |
| 13 | C18:2 (9, 12) | 40.06 | n.d. | n.d. | 0.27 ± 0.024 | 1.7 |
| 14 | C18:0 | 40.16 | 11.0 ± 1.0 | 28.19 | 0.54 ± 0.05 | 3.41 |
| 15 | C18:4 (6,9,12,15) | 40.90 | 0.34 ± 0.03 | 0.86 | 0.83 ± 0.07 | 5.19 |
| 16 | C18:3 (9,12,15) | 41.04 | 0.24 ± 0.02 | 0.6 | 0.157 ± 0.014 | 1.0 |
| 17 | C20:5 (5,8,11,14,17) | 47.61 | 2.21 ± 0.20 | 5.7 | 3.5 ± 0.3 | 21.84 |
| 18 | C20:4 (8,11,14,17) | 47.89 | 0.036 ± 0.003 | 0.09 | 0.052 ± 0.005 | 0.33 |
| 19 | C22:0 | 53.37 | 1.21 ± 0.11 | 3.14 | 0.115 ± 0.010 | 0.72 |
| 20 | C24:1(9) | 59.48 | 0.190 ± 0.017 | 0.19 | 0.063 ± 0.006 | 0.39 |
| 21 | C24:0 | 59.60 | 0.125 ± 0.011 | 0.33 | 0.054 ± 0.005 | 0.43 |
| ΣFAs | 39 | 16 | ||||
| Saturated FAs (SFA) | 30 | 76 | 6.6 | 41 | ||
| Monounsaturated FAs (MUFA) | 5.4 | 14 | 3.8 | 23 | ||
| Polyunsaturated FAs (PUFA) | 3.8 | 10 | 5.3 | 33 | ||
| Diatom Species | Fatty Acid Content (% of the FAs Total) | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C12:0 | C14:0 | C14:1 (n − 5) | C15:0 | C16:0 | C16:1 | C16:2 (n − 9,12) | C16:3 (n − 6,9,12) | C16:4 (n − 6,9,12,15) | C17:0 | C18:0 | C18:1 (9c) | C18:1 (n − 3) | C18:2 | C18:3 (n − 9,12,15) or (6,9,12) | C18:4 (n − 6,9,12,15) | C20:0 | C20:4 (n − 8,11,14,17) | C20:5 (n − 5,8,11,14,17) | C22:0 | C22:1 | C22:6 (n − 4,7,10,13,16,19) | C24:0 | |
| Entomoneis paludosa stationary phase of growth (this publication) | 10.5 | 6.3 | – | 0.5 | 26 | 10.1 | 0.95 | 1.3 | 0.34 | – | 28 | 3.34 | 2.13 | 0 | 0.6 | 0.86 | – | 0.09 | 5.7 | 3.14 | – | – | 0.33 |
| Entomoneis paludosa exponential phase of growth (this publication) | 0.9 | 14.7 | – | 0.54 | 20 | 19 | 2.02 | 2.3 | 0.75 | – | 3.4 | 2.23 | 1.83 | 1.69 | 0.98 | 5.2 | – | 0.33 | 21.9 | 0.72 | – | – | 0.34 |
| Entomoneis paludosa MLN+ [11] | – | 2.6 ± 13.4 | – | – | 30 ± 2 | 12 ± 9 | – | – | – | – | 1.7 ± 8.7 | 8.9 ± 2.3 | – | 1.1 ± 1.0 | – | – | – | – | – | – | – | – | – |
| Entomoneis paludosa MLN− [11] | – | 9.9 ± 2.1 | – | 0.7 ± 0.1 | 37 ± 7 | 4.0 ± 0.6 | – | – | – | – | 14 ± 2 | 20 ± 3 | – | 1.9 ± 1.8 | – | – | 1.2 ± 0.6 | – | – | – | – | – | – |
| Fistulifera sp. under nutrition sufficient [3] | – | 5.0 | – | – | 34 | 38 | 1.6 | 1.7 | – | – | 0.4 | 0.7 | – | – | 0.01 | 0.01 | 0.2 | – | – | – | – | – | – |
| Fistulifera sp under nutrition deficient [3] | – | 5.0 | – | – | 36 | 47 | 0.3 | 0.4 | – | – | 0.4 | 0.4 | – | – | 0.01 | 0.01 | 0.2 | – | – | – | – | – | – |
| Nitzschia alexandrina [11] MLN+ | – | 1.5 ± 0.5 | – | 0. 5± 0.1 | 24 ± 1 | 26 ± 3 | 2.9 ± 1.6 | – | – | – | 11 ± 4 | 9.4 ± 3.6 | – | – | – | – | 0.9 ± 0.2 | 1.7 ± 0.2 | 0.8 ± 0.1 | 3.7 ± 1.9 | – | 1.7 ± 0.3 | |
| Nitzschia alexandrina MLN− [11] | – | 3.7 ± 1 | – | 0.7 ± 0.1 | 39 ± 4 | 21 ± 4 | 9.5 ± 2 | – | – | – | 6.6 ± 0.9 | 5.7 ± 2.2 | – | 4.7 ± 1.3 | – | – | 0.6 ± 0.2 | – | – | – | 0.8 ± 0.3 | – | 0.9 ± 0.1 |
| Staurosira sp. MLN+ [11] | – | 1.7 ± 0.8 | – | 0.5 ± 0.2 | 41.8 ± 2.4 | 35.1 ± 10 | 4.6 ± 2.9 | – | – | – | 9.0 ± 10.2 | 3.3 ± 0.4 | – | – | – | – | 0.6 ± 0.2 | – | – | – | – | – | 0.3 ± 0.3 |
| Staurosira sp. MLN− [11] | – | 3.7 ± 1.3 | – | 0.3 ± 0.3 | 64.6 ± 6.0 | 6.9 ± 2.1 | 9.5 ± 0.7 | – | – | – | 4.6 ± 1.6 | 3.0 ± 2.0 | – | – | – | – | 0.2 ± 0.3 | – | – | – | – | – | – |
| Nitzschia closterium [37] | – | 5.4 | – | – | 21.8 | 47.5 | 2.2 | 2.4 | – | – | 0.9 | 7.1 | – | 0.31 | 0.25 | – | – | – | 10.4 | – | – | 0.22 | – |
| Odontella aurita [38] | – | 9.6 ± 2.8 | – | – | 11 ± 3 | 16 ± 1 | – | – | – | – | 1.4 ± 0.2 | 9.1 ± 0.8 | – | – | – | 1.1 ± 0.2 | – | – | 34 ± 2 | 4.1 ± 0.1 | – | ||
| Phaeodactylum tricornutum [12,30] | – | 6.7 ± 11 | – | – | 31 | 52 | – | – | – | – | – | 3.6 | – | – | – | – | – | – | 5.4 | – | – | – | – |
| Thalassiosira pseudonana [12,30] | – | 5.98 | – | – | 30 | 29 | – | – | – | – | 0.36 | 2.0 | – | – | – | – | – | – | 7.5 | – | – | 0.69 | – |
| Chaetoceros gracilis [39] | – | 6.7 | – | – | 42 | 41 | 2.7 | 1.5 | – | – | 1.1 | – | – | – | – | – | – | – | 3.9 | – | – | – | – |
| Atthey alongicornis [31] | – | 8.8 | – | – | 8.5 ± 0.3 | 25 | 8.8 ± 0.1 | 6.0 ± 0.1 | 3.8 ± 0.1 | – | 3.5 ± 0.3 | 4.2 ± 0.2 | – | 3.4 | 2.63 | 1.0 ± 0.1 | – | 0.07 | 19.1 | – | 0.13 | 4.0 ± 0.3 | – |
| Porosira glacialis [31] | – | 3.9 ± 0.4 | – | – | 12.0 ± 1.0 | 17 ± 2 | 5.2 ± 0.6 | 5.7 ± 0.2 | 9.6 ± 1.0 | – | 2.7 ± 0.6 | 1.7 ± 0.1 | – | 0.25 | 4.8 ± 0.2 | 4.2 ± 0.6 | – | – | 27 ± 2 | – | – | 3.9 ± 0.4 | – |
| Synedra sp. [40] | 1.8 | 5.2 | 6.5 | 0.2 | 31 | 10.1 | – | – | 6.4 | 22.1 | – | – | 0 | 3.6 | – | 0.9 | – | 11.8 | – | – | – | – | |
| Navicula sp. [40] | 2.8 | 4.1 | 2.2 | 0.1 | 33 | 24 | – | – | – | 1.2 | 11.3 | – | – | 1.2 | 3.1 | – | 1.5 | – | 15.2 | – | – | – | – |
| Cyclotella sp. [40] | 0.9 | 1.3 | 3.9 | 0.1 | 32 | 29 | – | – | – | 0.1 | 17.9 | – | – | 2.4 | 2.1 | – | 1.1 | – | 8.7 | – | – | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bedoshvili, Y.; Podunay, Y.; Nikonova, A.; Marchenkov, A.; Bairamova, E.; Davidovich, N.; Likhoshway, Y. Lipid and Fatty Acids Accumulation Features of Entomoneis cf. paludosa during Exponential and Stationary Growth Phases in Laboratory Culture. Diversity 2021, 13, 459. https://doi.org/10.3390/d13100459
Bedoshvili Y, Podunay Y, Nikonova A, Marchenkov A, Bairamova E, Davidovich N, Likhoshway Y. Lipid and Fatty Acids Accumulation Features of Entomoneis cf. paludosa during Exponential and Stationary Growth Phases in Laboratory Culture. Diversity. 2021; 13(10):459. https://doi.org/10.3390/d13100459
Chicago/Turabian StyleBedoshvili, Yekaterina, Yulia Podunay, Alyona Nikonova, Artyom Marchenkov, Elvira Bairamova, Nikolai Davidovich, and Yelena Likhoshway. 2021. "Lipid and Fatty Acids Accumulation Features of Entomoneis cf. paludosa during Exponential and Stationary Growth Phases in Laboratory Culture" Diversity 13, no. 10: 459. https://doi.org/10.3390/d13100459
APA StyleBedoshvili, Y., Podunay, Y., Nikonova, A., Marchenkov, A., Bairamova, E., Davidovich, N., & Likhoshway, Y. (2021). Lipid and Fatty Acids Accumulation Features of Entomoneis cf. paludosa during Exponential and Stationary Growth Phases in Laboratory Culture. Diversity, 13(10), 459. https://doi.org/10.3390/d13100459