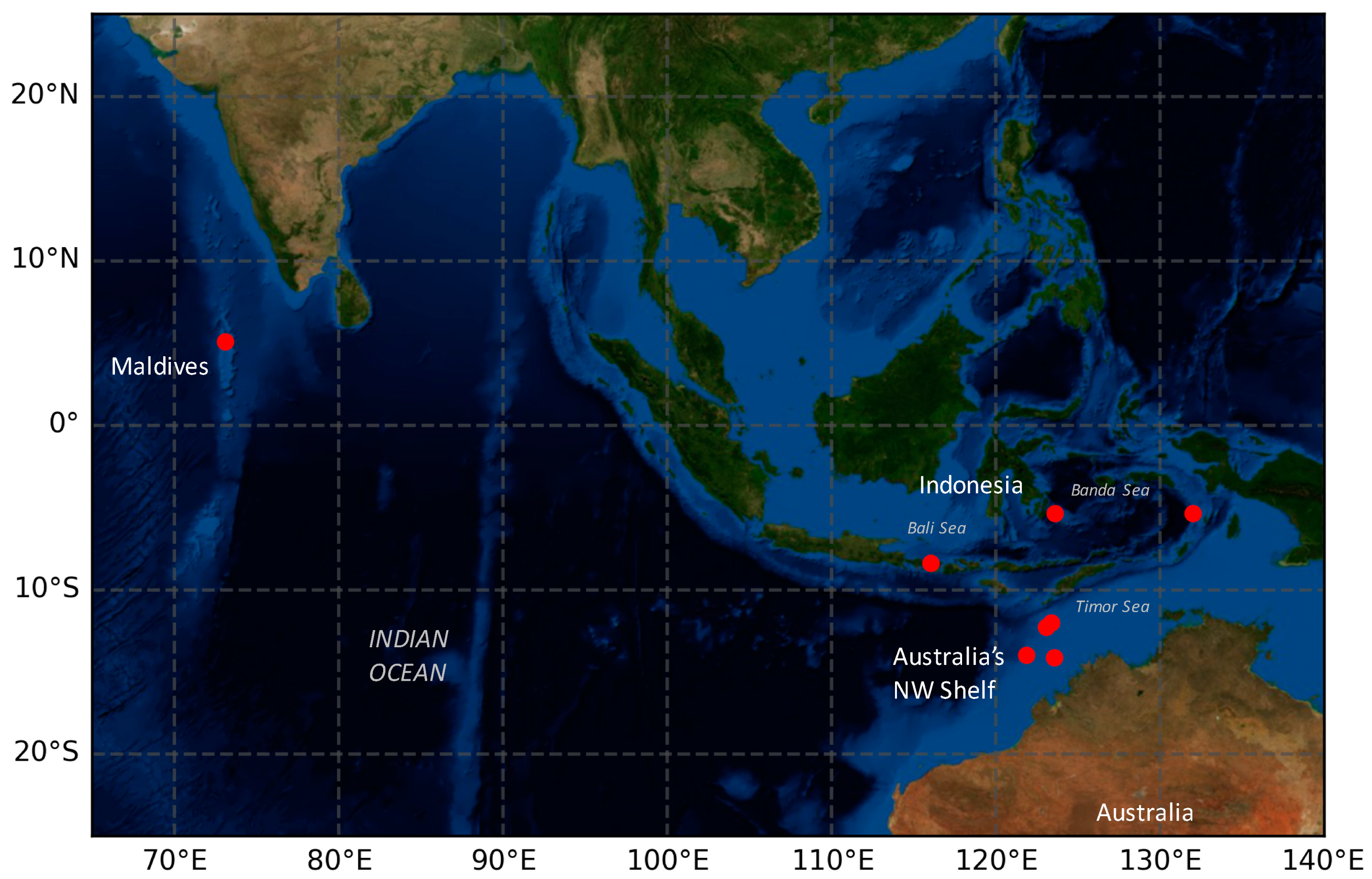
New Records of Heliopora hiberniana from SE Asia and the Central Indian Ocean




{kind=link}
{kind=link}
{kind=link}
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darling, E.S.; McClanahan, T.R.; Maina, J.; Gurney, G.G.; Graham, N.A.; Januchowski-Hartley, F.; Cinner, J.E.; Mora, C.; Hicks, C.C.; Maire, E.; et al. Social-environmental drivers inform strategic management of coral reefs in the Anthropocene. Nat. Ecol. Evol. 2019, 3, 1341–1350. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.T.; Alvarez-Filip, L.; Graham, N.A.; Mumby, P.J.; Wilson, S.K.; Kench, P.S.; Manzello, D.P.; Morgan, K.M.; Slangen, A.B.; Thomson, D.P.; et al. Loss of coral reef growth capacity to track future increases in sea level. Nature 2018, 396–400. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef]
- Graham, N.A.; Jennings, S.; MacNeil, M.A.; Mouillot, D.; Wilson, S.K. Predicting climate-driven regime shifts versus rebound potential in coral reefs. Nature 2015, 518, 94–97. [Google Scholar] [CrossRef]
- Inoue, S.; Kayanne, H.; Yamamoto, S.; Kurihara, H. Spatial community shift from hard to soft corals in acidified water. Nat. Clim. Chang. 2013, 3, 683–687. [Google Scholar] [CrossRef]
- Enochs, I.C.; Manzello, D.P.; Donham, E.M.; Kolodziej, G.; Okano, R.; Johnston, L.; Young, C.; Iguel, J.; Edwards, C.B.; Fox, M.D.; et al. Shift from coral to macroalgae dominance on a volcanically acidified reef. Nat. Clim. Chang. 2015, 5, 1083–1088. [Google Scholar] [CrossRef]
- Richards, Z.T.; Yasuda, N.; Kikuchi, T.; Foster, T.; Mitsuyuki, C.; Stat, M.; Suyama, Y.; Wilson, N.G. Integrated evidence reveals a new species in the ancient blue coral genus Heliopora (Octocorallia). Sci. Rep. 2018, 8, 15875. [Google Scholar] [CrossRef] [PubMed]
- Zann, L.P.; Bolton, L. The distribution, abundance and ecology of the blue coral Heliopora coerulea (Pallas) in the Pacific. Coral Reefs 1985, 4, 125–134. [Google Scholar] [CrossRef]
- Yasuda, N.; Taquet, C.; Nagai, S.; Fortes, M.; Fan, T.Y.; Phongsuwan, N.; Nadaoka, K. Genetic structure and cryptic speciation in the threatened reef-building coral Heliopora coerulea along Kuroshio Current. Bull. Mar. Sci. 2014, 90, 233–255. [Google Scholar] [CrossRef]
- Villanueva, R.D. Cryptic speciation in the stony octocoral Heliopora coerulea: Temporal reproductive isolation between two growth forms. Mar. Biodivers. 2016, 46, 503–507. [Google Scholar] [CrossRef]
- Williams, D.E.; Bright, A.J. White rings on the threatened coral, Acropora palmata, associated with foraging activity of the honeycomb cowfish, Acanthostracion polygonius (Ostraciidae). Coral Reefs 2013, 32, 651. [Google Scholar] [CrossRef][Green Version]
- Denis, V.; Ribas Deulofeu, L.; De Palmas, S.; Chen, C.A. First record of the scleractinian coral Psammocora albopicta from Korean waters. Mar. Biodivers. 2014, 44, 157–158. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richards, Z.T.; Haines, L.; Scaps, P.; Ader, D. New Records of Heliopora hiberniana from SE Asia and the Central Indian Ocean. Diversity 2020, 12, 328. https://doi.org/10.3390/d12090328
Richards ZT, Haines L, Scaps P, Ader D. New Records of Heliopora hiberniana from SE Asia and the Central Indian Ocean. Diversity. 2020; 12(9):328. https://doi.org/10.3390/d12090328
Chicago/Turabian StyleRichards, Zoe T., Leon Haines, Patrick Scaps, and Denis Ader. 2020. "New Records of Heliopora hiberniana from SE Asia and the Central Indian Ocean" Diversity 12, no. 9: 328. https://doi.org/10.3390/d12090328
APA StyleRichards, Z. T., Haines, L., Scaps, P., & Ader, D. (2020). New Records of Heliopora hiberniana from SE Asia and the Central Indian Ocean. Diversity, 12(9), 328. https://doi.org/10.3390/d12090328