Ichthyological Differentiation and Homogenization in the Pánuco Basin, Mexico

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
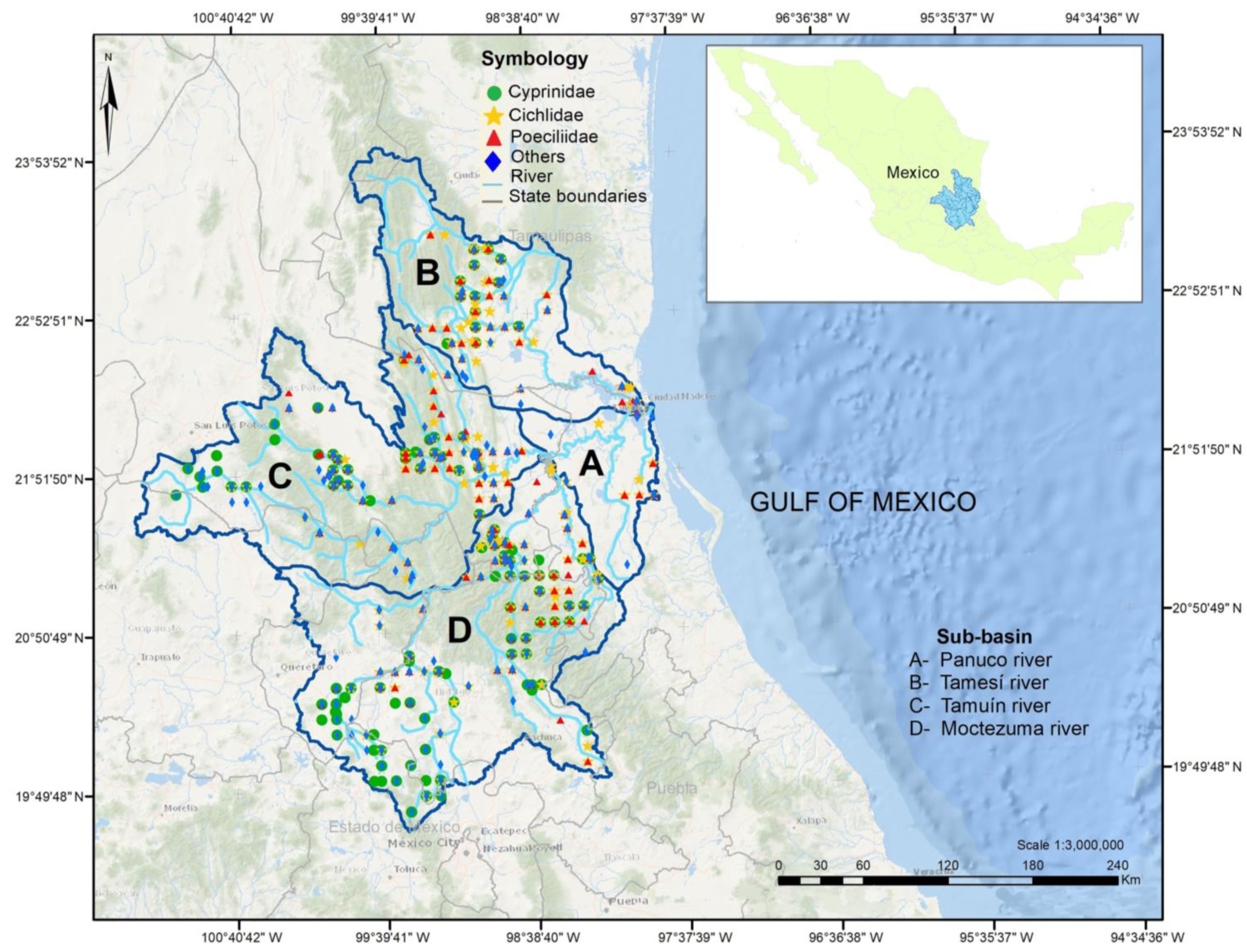
2.1. Units of Analysis
2.2. Database
2.3. Analyses
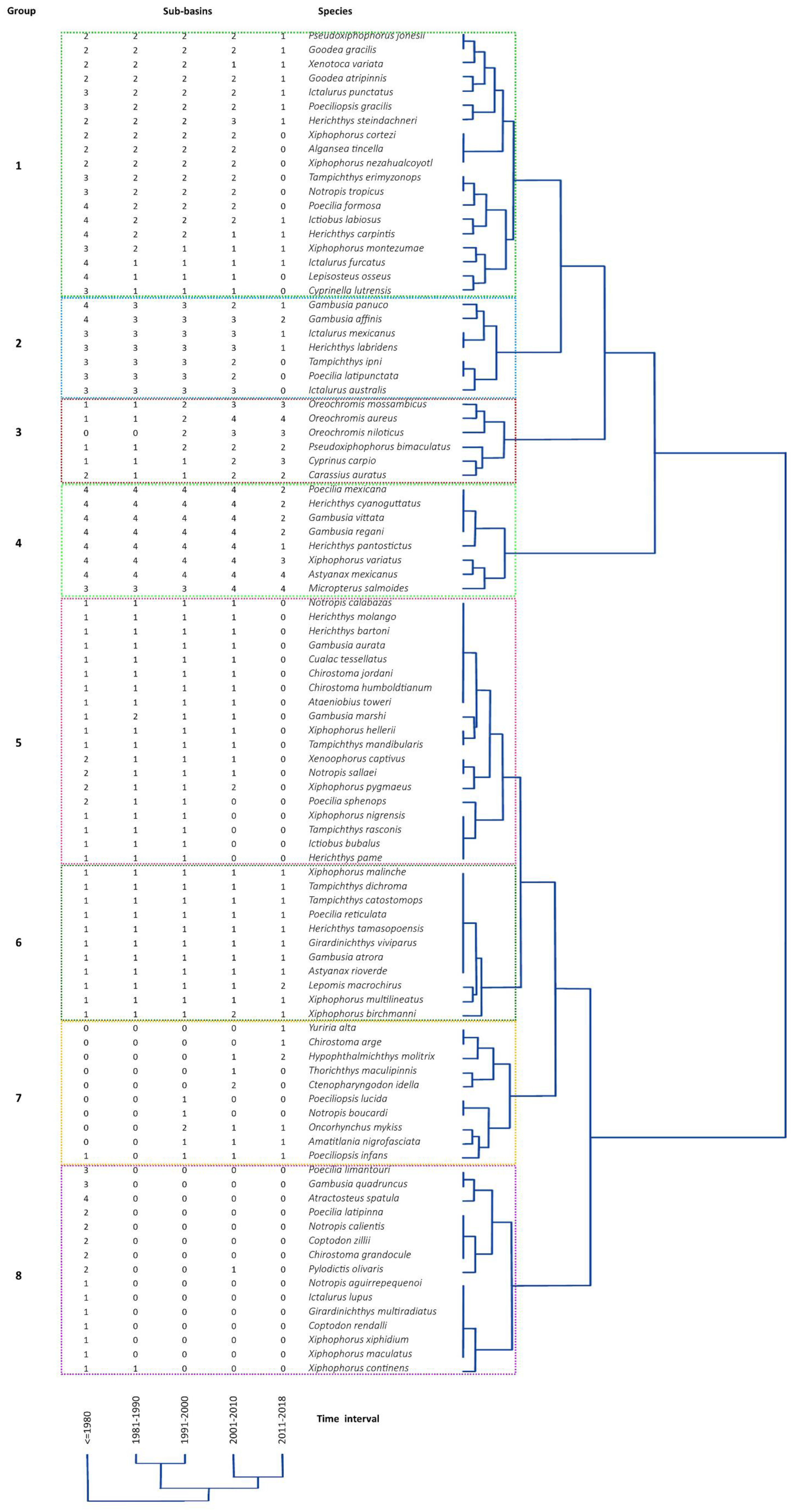
3. Results
4. Discussion
4.1. Compositional Changes
4.2. Assemblage Change among Time Intervals
4.3. Species Contribution
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| No. | Scientific Name | Interval 1 (≤1980) | Interval 2 (1981–1990) | Interval 3 (1991–2000) | Interval 4 (2001–2010) | Interval 5 (2011–2018) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | A | B | C | D | A | B | C | D | A | B | C | D | A | B | C | D | ||
| 1 | Algansea tincella | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| 2 | Amatitlania nigrofasciata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| 3 | Astyanax rioverde | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 4 | Astyanax mexicanus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 5 | Ataeniobius toweri | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 6 | Atractosteus spatula | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 | Carassius auratus | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 |
| 8 | Chirostoma arge | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| 9 | Chirostoma grandocule | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 10 | Chirostoma humboldtianum | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 11 | Chirostoma jordani | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 12 | Coptodon rendalli | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 13 | Coptodon zillii | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 14 | Ctenopharyngodon idella | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| 15 | Cualac tessellatus | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 16 | Cyprinella lutrensis | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 17 | Cyprinus carpio | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 |
| 18 | Gambusia affinis | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 |
| 19 | Gambusia atrora | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| 20 | Gambusia aurata | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 21 | Gambusia marshi | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 22 | Gambusia panuco | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 |
| 23 | Gambusia quadruncus | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 24 | Gambusia regani | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 25 | Gambusia vittata | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 |
| 26 | Girardinichthys multiradiatus | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 27 | Girardinichthys viviparus | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| 28 | Goodea atripinnis | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 |
| 29 | Goodea gracilis | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| 30 | Herichthys bartoni | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 31 | Herichthys carpintis | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 32 | Herichthys cyanoguttatus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 33 | Herichthys labridens | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 |
| 34 | Herichthys molango | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 35 | Herichthys pame | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 36 | Herichthys pantostictus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 |
| 37 | Herichthys steindachneri | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 |
| 38 | Herichthys tamasopoensis | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 39 | Hypophthalmichthys molitrix | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| 40 | Ictalurus australis | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
| 41 | Ictalurus furcatus | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| 42 | Ictalurus lupus | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 43 | Ictalurus mexicanus | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 |
| 44 | Ictalurus punctatus | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 |
| 45 | Ictiobus bubalus | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 46 | Ictiobus labiosus | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 |
| 47 | Lepisosteus osseus | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 48 | Lepomis macrochirus | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 |
| 49 | Micropterus salmoides | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 50 | Notropis aguirrepequenoi | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 51 | Notropis boucardi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 52 | Notropis calabazas | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 53 | Notropis calientis | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 54 | Notropis sallaei | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 55 | Notropis tropicus | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| 56 | Oncorhynchus mykiss | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| 57 | Oreochromis aureus | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 58 | Oreochromis mossambicus | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 |
| 59 | Oreochromis niloticus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 |
| 60 | Poecilia formosa | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 61 | Poecilia latipinna | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 62 | Poecilia latipunctata | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| 63 | Poecilia limantouri | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 64 | Poecilia mexicana | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 65 | Poecilia reticulata | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| 66 | Poecilia sphenops | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 67 | Poeciliopsis gracilis | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 |
| 68 | Poeciliopsis infans | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 69 | Poeciliopsis lucida | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 70 | Pseudoxiphophorus bimaculatus | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 |
| 71 | Pseudoxiphophorus jonesii | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| 72 | Pylodictis olivaris | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 73 | Tampichthys catostomops | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 74 | Tampichthys dichroma | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 75 | Tampichthys erimyzonops | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| 76 | Tampichthys ipni | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| 77 | Tampichthys mandibularis | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 78 | Tampichthys rasconis | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 79 | Thorichthys maculipinnis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 80 | Xenoophorus captivus | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 81 | Xenotoca variata | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 82 | Xiphophorus birchmanni | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| 83 | Xiphophorus continens | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 84 | Xiphophorus cortezi | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| 85 | Xiphophorus hellerii | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 86 | Xiphophorus maculatus | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 87 | Xiphophorus malinche | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| 88 | Xiphophorus montezumae | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 89 | Xiphophorus multilineatus | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 90 | Xiphophorus nezahualcoyotl | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| 91 | Xiphophorus nigrensis | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 92 | Xiphophorus pygmaeus | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| 93 | Xiphophorus variatus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 |
| 94 | Xiphophorus xiphidium | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 95 | Yuriria alta | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| No. | Family | Scientific Name | ORIGIN | Invasive (Yes /No) | Red List Iucn | Nom 059 Semarnat 2010 | |
|---|---|---|---|---|---|---|---|
| (Native = N, Non Native = NN) | Sub-Basin (A,B,C,D) | ||||||
| 1 | Cyprinidae | Algansea tincella | N | C | No | LC | No |
| 2 | Cichlidae | Amatitlania nigrofasciata | NN | Yes | NE | No | |
| 3 | Characidae | Astyanax rioverde | N | C | No | LC | No |
| 4 | Characidae | Astyanax mexicanus * | N | ABCD | No | LC/VU | No/A |
| 5 | Goodeidae | Ataeniobius toweri | N | C | No | EN | P |
| 6 | Lepisosteidae | Atractosteus spatula | N | B | No | LC | No |
| 7 | Cyprinidae | Carassius auratus | NN | Yes | LC | No | |
| 8 | Atherinopsidae | Chirostoma arge | NN | No | DD | No | |
| 9 | Atherinopsidae | Chirostoma grandocule | NN | No | DD | No | |
| 10 | Atherinopsidae | Chirostoma humboldtianum | NN | No | NE | No | |
| 11 | Atherinopsidae | Chirostoma jordani | NN | No | LC | No | |
| 12 | Cichlidae | Coptodon rendalli | NN | No | LC | No | |
| 13 | Cichlidae | Coptodon zillii | NN | Yes | LC | No | |
| 14 | Cyprinidae | Ctenopharyngodon idella | NN | Yes | NE | No | |
| 15 | Cyprinodontidae | Cualac tessellatus | N | C | No | VU | P |
| 16 | Cyprinidae | Cyprinella lutrensis | N | C | No | LC | A |
| 17 | Cyprinidae | Cyprinus carpio | NN | Yes | VU | No | |
| 18 | Poeciliidae | Gambusia affinis | NN | No | LC | No | |
| 19 | Poeciliidae | Gambusia atrora | N | D | No | DD | No |
| 20 | Poeciliidae | Gambusia aurata | N | B | No | DD | No |
| 21 | Poeciliidae | Gambusia marshi | NN | No | LC | A | |
| 22 | Poeciliidae | Gambusia panuco | N | AB | No | DD | No |
| 23 | Poeciliidae | Gambusia quadruncus | N | AC | No | NE | No |
| 24 | Poeciliidae | Gambusia regani | N | ABCD | No | DD | No |
| 25 | Poeciliidae | Gambusia vittata | N | B | No | LC | No |
| 26 | Goodeidae | Girardinichthys multiradiatus | NN | No | EN | No | |
| 27 | Goodeidae | Girardinichthys viviparus | NN | No | EN | P | |
| 28 | Goodeidae | Goodea atripinnis | N | C | No | LC | No |
| 29 | Goodeidae | Goodea gracilis | N | C | No | LC | No |
| 30 | Cichlidae | Herichthys bartoni | N | C | No | EN | P |
| 31 | Cichlidae | Herichthys carpintis | N | AB | No | LC | No |
| 32 | Cichlidae | Herichthys cyanoguttatus | NN | No | LC | No | |
| 33 | Cichlidae | Herichthys labridens | N | B | No | EN | A |
| 34 | Cichlidae | Herichthys molango | N | D | No | LC | No |
| 35 | Cichlidae | Herichthys pame | N | C | No | NE | No |
| 36 | Cichlidae | Herichthys pantostictus | N | BA | No | LC | No |
| 37 | Cichlidae | Herichthys steindachneri | N | C | No | EN | P |
| 38 | Cichlidae | Herichthys tamasopoensis | N | C | No | VU | No |
| 39 | Cyprinidae | Hypophthalmichthys molitrix | NN | Yes | NT | No | |
| 40 | Ictaluridae | Ictalurus australis | N | B | No | DD | A |
| 41 | Ictaluridae | Ictalurus furcatus | N | CD | No | LC | No |
| 42 | Ictaluridae | Ictalurus lupus | NN | No | DD | No | |
| 43 | Ictaluridae | Ictalurus mexicanus | N | C | No | VU | A |
| 44 | Ictaluridae | Ictalurus punctatus | N | BCD | No | LC | No |
| 45 | Catostomidae | Ictiobus bubalus | NN | No | LC | A | |
| 46 | Catostomidae | Ictiobus labiosus | N | CD | No | DD | No |
| 47 | Lepisosteidae | Lepisosteus osseus | N | ABC | No | LC | No |
| 48 | Centrarchidae | Lepomis macrochirus | NN | No | LC | No | |
| 49 | Centrarchidae | Micropterus salmoides | NN | Yes | LC | No | |
| 50 | Cyprinidae | Notropis aguirrepequenoi | NN | No | VU | Pr | |
| 51 | Cyprinidae | Notropis boucardi | NN | No | EN | A | |
| 52 | Cyprinidae | Notropis calabazas | N | C | No | CR | No |
| 53 | Cyprinidae | Notropis calientis | N | C | No | CR | No |
| 54 | Cyprinidae | Notropis sallaei | N | D | No | LC | No |
| 55 | Cyprinidae | Notropis tropicus | N | BC | No | NT | No |
| 56 | Salmonidae | Oncorhynchus mykiss | NN | Yes | NE | Pr | |
| 57 | Cichlidae | Oreochromis aureus | NN | Yes | LC | No | |
| 58 | Cichlidae | Oreochromis mossambicus | NN | Yes | NT | No | |
| 59 | Cichlidae | Oreochromis niloticus | NN | Yes | LC | No | |
| 60 | Poeciliidae | Poecilia formosa | N | ABD | No | LC | No |
| 61 | Poeciliidae | Poecilia latipinna | NN | No | LC | No | |
| 62 | Poeciliidae | Poecilia latipunctata | N | B | No | DD | P |
| 63 | Poeciliidae | Poecilia limantouri | NN | No | NE | No | |
| 64 | Poeciliidae | Poecilia mexicana | N | ABCD | No | LC | No |
| 65 | Poeciliidae | Poecilia reticulata | NN | Yes | NE | No | |
| 66 | Poeciliidae | Poecilia sphenops | NN | No | LC | Pr | |
| 67 | Poeciliidae | Poeciliopsis gracilis | NN | No | LC | No | |
| 68 | Poeciliidae | Poeciliopsis infans | NN | No | LC | No | |
| 69 | Poeciliidae | Poeciliopsis lucida | NN | No | DD | No | |
| 70 | Poeciliidae | Pseudoxiphophorus bimaculatus | NN | No | LC | No | |
| 71 | Poeciliidae | Pseudoxiphophorus jonesii | NN | No | LC | No | |
| 72 | Ictaluridae | Pylodictis olivaris | NN | No | LC | No | |
| 73 | Cyprinidae | Tampichthys catostomops | N | C | No | NT | No |
| 74 | Cyprinidae | Tampichthys dichroma | N | C | No | NE | A |
| 75 | Cyprinidae | Tampichthys erimyzonops | N | BC | No | DD | No |
| 76 | Cyprinidae | Tampichthys ipni | N | BCD | No | LC | No |
| 77 | Cyprinidae | Tampichthys mandibularis | N | C | No | EN | P |
| 78 | Cyprinidae | Tampichthys rasconis | N | C | No | EN | No |
| 79 | Cichlidae | Thorichthys maculipinnis | NN | No | NE | No | |
| 80 | Goodeidae | Xenoophorus captivus | N | C | No | EN | P |
| 81 | Goodeidae | Xenotoca variata | N | C | No | LC | No |
| 82 | Poeciliidae | Xiphophorus birchmanni | N | D | No | LC | No |
| 83 | Poeciliidae | Xiphophorus continens | N | C | No | DD | No |
| 84 | Poeciliidae | Xiphophorus cortezi | N | CD | No | DD | No |
| 85 | Poeciliidae | Xiphophorus hellerii | NN | No | LC | No | |
| 86 | Poeciliidae | Xiphophorus maculatus | NN | No | DD | No | |
| 87 | Poeciliidae | Xiphophorus malinche | N | D | No | DD | No |
| 88 | Poeciliidae | Xiphophorus montezumae | N | C | No | DD | No |
| 89 | Poeciliidae | Xiphophorus multilineatus | N | C | No | DD | No |
| 90 | Poeciliidae | Xiphophorus nezahualcoyotl | N | BC | No | DD | No |
| 91 | Poeciliidae | Xiphophorus nigrensis | N | C | No | DD | No |
| 92 | Poeciliidae | Xiphophorus pygmaeus | N | C | No | DD | No |
| 93 | Poeciliidae | Xiphophorus variatus | N | ABCD | No | LC | No |
| 94 | Poeciliidae | Xiphophorus xiphidium | NN | No | LC | No | |
| 95 | Cyprinidae | Yuriria alta | NN | No | EN | No | |
References
- Morrone, J.J.; Escalante, T. Introducción a la Biogeografía, 1st ed.; Prensas de Ciencias Universidad Nacional Autónoma de México: Mexico, DF, Mexico, 2016; p. 316. [Google Scholar]
- Petsch, D.K. Causes and consequences of biotic homogenization in freshwater ecosystems. Int. Rev. Hydrobiol. 2016, 101, 113–122. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Mckinney, M. Biotic Homogenization, 1st ed.; Springer: New York, NY, USA, 2001; p. 289. [Google Scholar]
- Olden, J.D. Biotic Homogenisation. In eLS Chichester SSX, ENG; John Wiley and Sons Ltd.: London, UK, 2016; p. 8. [Google Scholar]
- Olden, J.D.; Poff, N.L. Toward a mechanistic understanding and prediction of biotic homogenization. Am. Nat. 2003, 162, 442–460. [Google Scholar] [CrossRef]
- Hudson, P.F.; Hendrickson, D.A.; Benke, A.C.; Rodiles-Hernández, R.; Minckle, W.L. Rivers of Mexico. In Rivers of North America; Benke, A.C., Cushing, C.E., Eds.; Academic Press: Toronto, ON, Canada, 2005; pp. 1030–1084. [Google Scholar]
- Taylor, E.B. Changes in taxonomy and species distributions and their influence on estimates of faunal homogenization and differentiation in freshwater fishes. Divers. Distrib. 2010, 16, 676–689. [Google Scholar] [CrossRef]
- Rahel, F.J. Homogenization of fish faunas across the United States. Science 2000, 288, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Olden, J.D.; Poff, L.; McKinney, M.L. Forecasting faunal and floral homogenization associated with human population geography in North America. Biol. Conserv. 2006, 127, 261–271. [Google Scholar] [CrossRef]
- Leprieur, F.; Beauchard, O.; Hugueny, B.; Grenouillet, G.; Brosse, S. Null model of biotic homogenization: A test with the European freshwater fish fauna. Divers. Distrib. 2008, 14, 291–300. [Google Scholar] [CrossRef]
- Daga, V.S.; Skóra, F.; Padial, A.A.; Abilhoa, V.; Gubiani, É.A.; Vitule, J.R.S. Homogenization dynamics of the fish assemblages in Neotropical reservoirs: Comparing the roles of introduced species and their vectors. Hydrobiologia 2015, 746, 327–347. [Google Scholar] [CrossRef]
- Vitule, J.R.S.; Skóra, F.; Abilhoa, V. Homogenization of freshwater fish faunas after the elimination of a natural barrier by a dam in Neotropics. Divers. Distrib. 2012, 18, 111–120. [Google Scholar] [CrossRef]
- Vargas, P.V.; Arismendi, I.; Gomez-Uchida, D. Evaluating taxonomic homogenization of freshwater fish assemblages in Chile. Rev. Chil. Hist. Nat. 2015, 88, 16. [Google Scholar] [CrossRef]
- Miller, R.R.; Minckley, W.L.; Norris, S.M. Peces Dulceacuícolas de México, 1st ed.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Sociedad Ictiológica Mexicana A.C., El Colegio de la Frontera Sur y Consejo de los Peces del Desierto: Mexico City, Mexico, 2009; p. 559. [Google Scholar]
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. BioScience 2008, 58, 403–414. [Google Scholar] [CrossRef]
- Gutiérrez-Yurrita, P.J.; Morales-Ortiz, J.A.; Marín-García, L. Diversidad biológica, distribución y estrategias de conservación de la ictiofauna de la cuenca del río Moctezuma, centro de Mexico. Limnetica 2013, 32, 215–228. [Google Scholar]
- Espinosa Perez, H.; Ramírez, H.M. Exotic and invasive fishes in Mexico. Check List 2015, 11, 1–13. [Google Scholar] [CrossRef]
- Soto Galera, E. Actualización de la Colección Nacional de Peces Dulceacuícolas Mexicanos, ENCB. Available online: https://www.gbif.org/dataset/36b77d42-895c-4192-8872-8f234d58071a (accessed on 30 April 2019).
- Darnell, R.M. Fishes of the Rio Tamesi and related coastal lagoons in East-Central Mexico. Inst. Mar. Sci. 1962, 8, 290–365. [Google Scholar]
- Contreras-Balderas, S.; Ramírez-Flores, M. Inventario nacional de la ictiofauna dulceacuícola mexicana. In Estado de Salud de la Acuacultura en Mexico; Instituto Nacional de la Pesca/Secretaría de Medio Ambiente, Recursos Naturales y Pesca: Mexico, DF, Mexico, 2000; Volume 5, pp. 47–70. [Google Scholar]
- García de León, F.J.; Hernández Sandoval, A.I.; Contreras-Catala, F.; Sánchez-Velasco, L.; Ruiz-Campos, G. Distribution of fishes in the Río Guayalejo-Río Tamesí system and relationships with environmental factors in northeastern Mexico. Environ. Biol. Fishes 2018, 167–180. [Google Scholar] [CrossRef]
- Contreras-Balderas, S.; Almada-Villela, P.; de Lozano-Vilano, M.L.; García-Ramírez, M. Freshwater fish at risk or extinct in Mexico. Rev. Fish Biol. Fish. 2002, 12, 241–251. [Google Scholar] [CrossRef]
- DOF-Diario Oficial de la Federación. Acuerdo por el que se dan a Conocer los Resultados del Estudio Técnico de Aguas Nacionales Superficiales en las Cuencas Hidrológicas Río Amajac, Río Verde 1, Río Verde 2, Río Verde 3, Río Tamasopo 1, Río Tamasopo 2, Río Gallinas, Río El Salto, Río Valles, Río Tampaón 1, Río Choy, Río Coy 2, Río Tampaón 2, Río Huichihuayan, Río Moctezuma 3, Río Moctezuma 5 y Río Pánuco 1, Mismas que Forman Parte de la Subregión Hidrológica Río Pánuco, de la Región Hidrológica Número 26 Pánuco. Available online: http://www.dof.gob.mx/nota_detalle.php?codigo=5512516&fecha=07/02/2018 (accessed on 10 December 2019).
- INEGI-Instituto Nacional de Estadística y Geografía. Mapas Hidrológicos. Available online: https://www.inegi.org.mx/temas/hidrologia/ (accessed on 11 May 2018).
- Myers, G.S. Salt-tolerance of fresh-water fish groups in relation to zoogeographical problems. Bijdr. Tot Dierkd. 1949, 28, 315–322. [Google Scholar]
- FishNet2 Search. Available online: http://www.fishnet2.net/search.aspx (accessed on 4 January 2019).
- GBIF-Global Biodiversity Information Facility. Available online: https://www.gbif.org/ (accessed on 30 April 2019).
- Arce-H, M.; Lundberg, J.G.; O’Leary, M.A. Phylogeny of the North American catfish family Ictaluridae (Teleostei: Siluriformes) combining morphology, genes and fossils. Cladistics 2017, 33, 406–428. [Google Scholar] [CrossRef]
- Ávila, E.G.; Valero, A.; Macías-Garcia, C. Componentes conductuales del cortejo incrementan la exposición a parásitos en el pez vivíparo xenoophorus captivus. TIP Rev. Espec. En Cienc. Quím.-Biológicas 2011, 14, 83–89. [Google Scholar]
- Barbour, C.D. The systematics and evolution of the genus Chirostoma Swainson (Pisces, Atherinidae). Tulane Stud. Zool. Bot. 1973, 18, 97–141. [Google Scholar]
- Bart, H.L.; Clements, M.D.; Blanton, R.E.; Piller, K.R.; Hurley, D.L. Discordant molecular and morphological evolution in buffalofishes (Actinopterygii: Catostomidae). Mol. Phylogenet. Evol. 2010, 56, 808–820. [Google Scholar] [CrossRef]
- Bloom, D.D.; Weir, J.T.; Piller, K.R.; Lovejoy, N.R. Do freshwater fishes diversify faster than marine fishes? A test using state-dependent diversification analyses and molecular phylogenetics of new world silversides (atherinopsidae). Evolution 2013, 67, 2040–2057. [Google Scholar] [CrossRef] [PubMed]
- Bono, L.M.; Rios-Cardenas, O.; Morris, M.R. Alternative life histories in Xiphophorus multilineatus: evidence for different ages at sexual maturity and growth responses in the wild. J. Fish Biol. 2011, 78, 1311–1322. [Google Scholar] [CrossRef]
- Castañeda-Rivera, M.; Grijalva-Chon, J.M.; Gutiérrez-Millán, L.E.; Ruiz-Campos, G.; Varela-Romero, A. Analysis of the Ictalurus pricei complex (Teleostei: Ictaluridae) in northwest Mexico based on mitochondrial DNA. Southwest. Nat. 2014, 59, 434–438. [Google Scholar] [CrossRef]
- Chernoff, B.; Miller, R.R. Fishes of the Notropis calientis complex with a key to the southern shiners of Mexico. Copeia 1986, 1986, 170–183. [Google Scholar] [CrossRef]
- Culumber, Z.W.; Rosenthal, G.G. Mating preferences do not maintain the tailspot polymorphism in the platyfish, Xiphophorus variatus. Behav. Ecol. 2013, 24, 1286–1291. [Google Scholar] [CrossRef][Green Version]
- Culumber, Z.W.; Tobler, M. Ecological divergence and conservatism: spatiotemporal patterns of niche evolution in a genus of livebearing fishes (Poeciliidae: Xiphophorus). BMC Evol. Biol. 2016, 16, 44. [Google Scholar] [CrossRef]
- Doadrio, I.; Domínguez, O. Phylogenetic relationships within the fish family Goodeidae based on cytochrome b sequence data. Mol. Phylogenet. Evol. 2004, 31, 416–430. [Google Scholar] [CrossRef]
- Domínguez-Domí, O.; Pedraza-Lara, C.; Gurrola-Sánchez, N.; Perea, S.; Pérez-Rodríguez, R.; Israde-Alcántara, I.; Garduño-Monroy, V.H.; Doadrio, I.; Pérez-Ponce de León, G.; Brooks, D.R. Historical biogeography of the Goodeinae (Cyprinodontiformes). In Viviparous Fishes II; Uribe, M.C., Grier, H.J., Eds.; New Life Publication: Homestead, FL, USA, 2010; pp. 19–61. [Google Scholar]
- Domínguez-Domínguez, O.; Bernal-Zuñiga, D.M.; Piller, K.R. Two new species of the genus Xenotoca Hubbs and Turner, 1939 (Teleostei, Goodeidae) from central-western Mexico. Zootaxa 2016, 4189, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Echelle, A.A.; Carson, E.W.; Echelle, A.F.; Bussche, R.A.V.D.; Dowling, T.E.; Meyer, A. Historical biogeography of the new-world pupfish Genus Cyprinodon (Teleostei: Cyprinodontidae). Copeia 2005, 2005, 320–339. [Google Scholar] [CrossRef]
- Kang, J.H.; Schartl, M.; Walter, R.B.; Meyer, A. Comprehensive phylogenetic analysis of all species of swordtails and platies (Pisces: Genus Xiphophorus) uncovers a hybrid origin of a swordtail fish, Xiphophorus monticolus, and demonstrates that the sexually selected sword originated in the ancestral lineage of the genus, but was lost again secondarily. BMC Evol. Biol. 2013, 13, 1–19. [Google Scholar]
- Lundberg, J.L. The phylogeny of ictalurid catfishes: A synthesis of recent work. In Systematics, Historical Ecology, and North American Freshwater Fishes; Mayden, R.L., Ed.; Stanford University Press: Stanford, CA, USA, 1992; pp. 392–420. [Google Scholar]
- Lyons, J.; Mercado-Silva, N. Notropis calabazas (Teleostei; Cyprinidae): new species from the Río Pánuco Basin of Central México. Copeia 2004, 2004, 868–875. [Google Scholar] [CrossRef]
- Lyons, J.; Piller, K.R.; Artigas-Azas, J.M.; Dominguez-Dominguez, O.; Gesundheit, P.; Köck, M.; Medina-Nava, M.; Mercado-Silva, N.; García, A.R.; Findley, K.M. Distribution and current conservation status of the Mexican Goodeidae (Actinopterygii, Cyprinodontiformes). ZooKeys 2019, 885, 115–158. [Google Scholar] [CrossRef]
- MacLaren, R.D.; Fontaine, A. Incongruence between the sexes in preferences for body and dorsal fin size in Xiphophorus variatus. Behav. Proc. 2013, 92, 99–106. [Google Scholar] [CrossRef]
- Miranda, R.; Galicia, D.; Monks, S.; Pulido-Flores, G. First record of Goodea atripinnis (Cyprinodontiformes: Goodeidae) in the state of Hidalgo (Mexico) and some considerations about its taxonomic position. Hidrobiológica 2010, 20, 185–190. [Google Scholar]
- Ornelas-García, C.P.; Martínez-Ramírez, E.; Doadrio, I. A new species of killifish of the family Profundulidae from the highlands of the Mixteca region, Mexico. Rev. Mex. Biodivers. 2015, 86, 926–933. [Google Scholar] [CrossRef]
- Palacios, M.; Voelker, G.; Arias Rodriguez, L.; Mateos, M.; Tobler, M. Phylogenetic analyses of the subgenus Mollienesia (Poecilia, Poeciliidae, Teleostei) reveal taxonomic inconsistencies, cryptic biodiversity, and spatio-temporal aspects of diversification in Middle America. Mol. Phylogenet. Evol. 2016, 103, 230–244. [Google Scholar] [CrossRef]
- Pérez-Miranda, F.; Mejía, O.; Soto-Galera, E.; Espinosa-Pérez, H.; Piálek, L.; Říčan, O. Phylogeny and species diversity of the genus Herichthys (Teleostei: Cichlidae). J. Zool. Syst. Evol. Res. 2018, 56, 223–247. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, R.; Domínguez-Domínguez, O.; Pérez Ponce de León, G.; Doadrio, I. Phylogenetic relationships and biogeography of the genus Algansea Girard (Cypriniformes: Cyprinidae) of central Mexico inferred from molecular data. BMC Evol. Biol. 2009, 9, 223. [Google Scholar] [CrossRef] [PubMed]
- Reznick, D.N.; Furness, A.I.; Meredith, R.W.; Springer, M.S. The origin and biogeographic diversification of fishes in the family Poeciliidae. PLoS ONE 2017, 12, e0172546. [Google Scholar] [CrossRef]
- Říčan, O.; Piálek, L.; Dragová, K.; Novák, J. Diversity and Evolution of the Middle American Cichlid Fishes (Teleostei: Cichlidae) with Revised Classification; Senckenberg Gesellschaft für Naturforschung: Frankfurt, Germany, 2016; p. 102. [Google Scholar]
- Rodiles-Hernández, R.; Lundberg, J.G.; Sullivan, J.P. Taxonomic discrimination and identification of extant blue catfishes (Siluriformes: Ictaluridae: Ictalurus furcatus Group). Proc. Acad. Nat. Sci. Phila. 2010, 159, 67–82. [Google Scholar] [CrossRef]
- Rosen, D.E. Middle-american poeciliid fishes of the genus Xiphophorus. Bull. Fla. State Mus. Biol. Sci. 1960, 5, 57–242. [Google Scholar]
- Rosen, D.E.; Bailey, R.M. The poeciliid fishes (Cyprinodontiformes): their structure, zoogeography, and systematics. Bull. AMNH 1963, 126, 1–176. [Google Scholar]
- Schönhuth, S.; Doadrio, I. Phylogenetic relationships of Mexican minnows of the genus Notropis (Actinopterygii, Cyprinidae). Biol. J. Linn. Soc. 2003, 80, 323–337. [Google Scholar] [CrossRef]
- Schönhuth, S.; Doadrio, I.; Dominguez-Dominguez, O.; Hillis, D.M.; Mayden, R.L. Molecular evolution of southern North American Cyprinidae (Actinopterygii), with the description of the new genus Tampichthys from central Mexico. Mol. Phylogenet. Evol. 2008, 47, 729–756. [Google Scholar] [CrossRef]
- Schönhuth, S.; Mayden, R.L. Phylogenetic relationships in the genus Cyprinella (Actinopterygii: Cyprinidae) based on mitochondrial and nuclear gene sequences. Mol. Phylogenet. Evol. 2010, 55, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.A. Molecular systematics of the genus Allodontichthys (Cyprinodontiformes: Goodeidae). Rev. Fish Biol. Fish. 2002, 12, 193–205. [Google Scholar] [CrossRef]
- Wright, J.J.; David, S.R.; Near, T.J. Gene trees, species trees, and morphology converge on a similar phylogeny of living gars (Actinopterygii: Holostei: Lepisosteidae), an ancient clade of ray-finned fishes. Mol. Phylogenet. Evol. 2012, 63, 848–856. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; Wiley: Hoboken, NJ, USA, 2016; p. 707. [Google Scholar]
- Eschmeyer’s Catalog of Fishes. Available online: https://www.calacademy.org/scientists/projects/eschmeyers-catalog-of-fishes (accessed on 16 December 2019).
- CONABIO Especies Exóticas Invasoras. Available online: http://enciclovida.mx/exoticas-invasoras?grupo=Peces (accessed on 16 December 2019).
- Jiménez-Valverde, A.; Hortal, J. Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Rev. Ibérica Aracnol. 2003, 8, 151–161. [Google Scholar]
- EstimateS: Stistical Estimation of Species Richness and Shared Species from Samples. Version 9.1. Available online: http://viceroy.eeb.uconn.edu/estimates/ (accessed on 9 May 2019).
- Demey, J.R.; Pla, L.; Vicente-Villardón, J.L.; Di Rienzo, J.; Casanoves, F. Medidas de distancia y similitud. In Valoración y Análisis de la Diversidad Funcional y su Relación con los Servicios Ecosistémicos; Casanoves, F., Pla, L., Di Rienzo, J.A., Eds.; Centro Agronómico Tropical de Investigación y Enseñanza: Turrialba, CR, USA, 2011; Volume 5, pp. 47–59. [Google Scholar]
- Anderson, M.J. Permutational multivariate analysis of variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; American Cancer Society: Atlanta, GA, USA, 2017; pp. 1–15. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER (Version 7). Windows; PRIMER-EPlymouth: Devon, UK, 2015. [Google Scholar]
- Murtagh, F.; Legendre, P. Ward’s hierarchical agglomerative clustering method: Which algorithms implement Ward’s criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- StatSoft STATISTICA (Version 10). Available online: https://www.bioz.com/result/statistica%2010%20software/product/StatSoft%20inc (accessed on 9 April 2020).
- Domínguez, O. Notropis calientis. IUCN Red List Threat Species; IUCN: Gland, Switzerland, 2019; p. e.T14881A546437. [Google Scholar]
- Mercado Silva, N. Xiphophorus continens. IUCN Red List Threat Species; IUCN: Gland, Switzerland, 2019; p. e.T191777A2002890. [Google Scholar]
- Ibáñez, A.L.; Pérez-Ramírez, M.; García-Calderón, J.L. Institutional development of freshwater fish stocking in Mexico. J. Fish Biol. 2014, 85, 2003–2009. [Google Scholar] [CrossRef]
- Mejía-Mojica, H.; de Rodríguez-Romero, F.J.; Díaz-Pardo, E. Recurrencia histórica de peces invasores en la Reserva de la Biósfera Sierra de Huautla, Mexico. Rev. Biol. Trop. 2012, 60, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Amador del Angel, L.E.; Wakida-Kusunoki, A.T. Peces invasores en el Sureste de México. In Especies Acuáticas Invasoras en Mexico; Mendoza, R., Koleff, P., Eds.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico, DF, Mexico, 2014; pp. 425–433. [Google Scholar]
- Tampichthys rasconis. IUCN Red List Threat. Species. 2019. Available online: https://www.iucnredlist.org/species/191296/1975296 (accessed on 2 May 2018).
- Dextrase, A.J.; Mandrak, N.E. Impacts of alien invasive species on freshwater fauna at risk in Canada. Biol. Invasions 2006, 8, 13–24. [Google Scholar] [CrossRef]
- Light, T.; Marchetti, M.P. Distinguishing between invasions and habitat changes as drivers of diversity loss among California’s freshwater fishes. Conserv. Biol. J. Soc. Conserv. Biol. 2007, 21, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, H.R.; Teimori, A.; Owfi, F.; Abbasi, K.; Coad, B.W. Alien and invasive freshwater fish species in Iran: Diversity, environmental impacts and management. Iran. J. Ichthyol. 2015, 1, 61–72. [Google Scholar]
- Witte, F.; Goldschmidt, T.; Wanink, J.; van Oijen, M.; Goudswaard, K.; Witte-Maas, E.; Bouton, N. The destruction of an endemic species flock: Quantitative data on the decline of the haplochromine cichlids of Lake Victoria. Environ. Biol. Fishes 1992, 34, 1–28. [Google Scholar] [CrossRef]
- Guzman, A.F.; Barragan, S.J. Presencia de bagre sudamericano (Osteichthyes: Loricariidae) en el rio Mezcala, Guerrero, Mexico. Vertebr. Mex. 1997, 3, 1–4. [Google Scholar]
- Contreras-Balderas, S.; Ruiz-Campos, G.; Schmitter-Soto, J.J.; Díaz-Pardo, E.; Contreras-McBeath, T.; Medina-Soto, M.; Zambrano-González, L.; Varela-Romero, A.; Mendoza-Alfaro, R.; Ramírez-Martínez, C.; et al. Freshwater fishes and water status in México: A country-wide appraisal. Aquat. Ecosyst. Health Manag. 2008, 11, 246–256. [Google Scholar] [CrossRef]
- Olds, A.A.; Smith, M.K.S.; Weyl, O.L.F.; Russell, I.A. Invasive alien freshwater fishes in the Wilderness lakes system, a wetland of international importance in the Western Cape province, South Africa. Afr. Zool. 2011, 46, 179–184. [Google Scholar] [CrossRef]
- Marchetti, M.P.; Moyle, P.B.; Levine, R. Invasive species profiling? Exploring the characteristics of non-native fishes across invasion stages in California. Freshw. Biol. 2004, 49, 646–661. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The population biology of invasive species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef]
- Gozlan, R.E.; Britton, J.R.; Cowx, I.; Copp, G.H. Current knowledge on non-native freshwater fish introductions. J. Fish Biol. 2010, 76, 751–786. [Google Scholar] [CrossRef]
- Taylor, E.B. An analysis of homogenization and differentiation of Canadian freshwater fish faunas with an emphasis on British Columbia. Can. J. Fish. Aquat. Sci. 2004, 61, 68–79. [Google Scholar] [CrossRef]
- Clavero, M.; García-Berthou, E. Homogenization dynamics and introduction routes of invasive freshwater fish in the Iberian Peninsula. Ecol. Appl. 2006, 16, 2313–2324. [Google Scholar] [CrossRef]
- Marchetti, M.P.; Lockwood, J.L.; Light, T. Effects of urbanization on California’s fish diversity: Differentiation, homogenization and the influence of spatial scale. Biol. Conserv. 2006, 127, 310–318. [Google Scholar] [CrossRef]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef]
- Contreras-MacBeath, T.; Mejia-Mojica, H.; Carrillo-Wilson, R.C. Negative impact on the aquatic ecosystems of the state of Morelos, Mexico from introduced aquarium and other commercial fish. Aquar. Sci. Conserv. 1998, 2, 67–78. [Google Scholar] [CrossRef]
- Trujillo-Jiménez, P. Trophic spectrum of the cichlids Cichlasoma (Parapetenia) istlanum and Cichlasoma (Arconcentrus) nigrofasciatum in the Amacuzac River, Morelos, Mexico. J. Freshw. Ecol. 1998, 13, 465–473. [Google Scholar] [CrossRef][Green Version]
- Varela-Romero, A.; Ruiz-Campos, G.; Yépiz-Velázquez, L.M.; Alaníz-García, J. Distribution, habitat and conservation status of desert pupfish (Cyprinodon macularius) in the Lower Colorado River Basin, Mexico. Rev. Fish Biol. Fish. 2002, 12, 157–165. [Google Scholar] [CrossRef]
- Cohen, A.N.; Carlton, J.T. Accelerating invasion rate in a highly invaded estuary. Science 1998, 279, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Región hidrológica Num. 26 Río Pánuco. Cuenca del Río Guayalejo-Tamesí. Available online: https://agua.org.mx/biblioteca/region-hidrologica-num-26-rio-panuco-cuenca-del-rio-guayalejo-tamesi/ (accessed on 2 January 2020).
- Buenfil Friedman, J. Adaptación a los Impactos del Cambio Climático en los Humedales Costeros del Golfo de Mexico; Instituto Nacional de Ecología: Mexico, DF, Mexico, 2009; p. 485. [Google Scholar]
- Edwards, P.J.; Abivardi, C. The value of biodiversity: Where ecology and economy blend. Biol. Conserv. 1998, 83, 239–246. [Google Scholar] [CrossRef]
- Guo, Q.; Olden, J.D. Spatial scaling of non-native fish richness across the United States. PLoS ONE 2014, 9, 1–6. [Google Scholar] [CrossRef]
- Contreras-Balderas, S.; Almada-Villela, P. Astyanax mexicanus ssp. jordani. IUCN Red List Threat. Species 2019. [Google Scholar]
- Schmitter-Soto, J.J. A revision of Astyanax (Characiformes: Characidae) in Central and North America, with the description of nine new species. J. Nat. Hist. 2017, 51, 1331–1424. [Google Scholar] [CrossRef]



| Group | Species in Group | Native | Non Native | Invasive | IUCN * | NOM 059 SEMARNAT 2010 ** | Group Attributes |
|---|---|---|---|---|---|---|---|
| 1 | 19 | 17 | 2 | 0 | 2 | 2 | Eight species were not found in later time intervals. |
| 2 | 7 | 6 | 1 | 0 | 2 | 4 | Three species were not found in later time intervals. |
| 3 | 6 | 0 | 6 | 5 | 2 | 0 | All species present through time intervals. |
| 4 | 8 | 6 | 2 | 1 | 0 | 0 | All species present through time intervals. |
| 5 | 19 | 13 | 6 | 0 | 7 | 8 | Species were lost between time intervals 4 and 5. |
| 6 | 11 | 8 | 3 | 1 | 3 | 2 | All species present through time intervals. |
| 7 | 10 | 0 | 10 | 4 | 3 | 2 | Nine species were introduced after time interval 3, but four were not found in the last time interval. |
| 8 | 15 | 4 | 11 | 1 | 3 | 1 | Species were not reported after time interval 1. |
| Species | DF1 | DF2 |
|---|---|---|
| Algansea tincella | 0.038516 | 0.030990 |
| Amatitlania nigrofasciata | −0.006545 | −0.114626 |
| Ataeniobius toweri | 0.022237 | 0.017892 |
| Carassius auratus | −0.007613 | 0.028727 |
| Chirostoma grandocule | 0.008891 | 0.310992 |
| Coptodon rendalli | 0.005133 | 0.179551 |
| Coptodon zillii | 0.008891 | 0.310992 |
| Ctenopharyngodon idella | 0.035840 | −0.127134 |
| Cyprinus carpio | −0.029556 | −0.062774 |
| Gambusia affinis | 0.023831 | 0.103444 |
| Gambusia marshi | 0.024345 | 0.026311 |
| Ictiobus bubalus | 0.013731 | 0.063038 |
| Notropis boucardi | 0.012446 | −0.089352 |
| Eigenvalue | 126.94 | 7.05 |
| % of variance | 92.79 | 5.1 |
| Species | DF1 | DF2 |
|---|---|---|
| Algansea tincella | 0.103136 | −0.044257 |
| Amatitlania nigrofasciata | 0.056162 | 0.165854 |
| Ataeniobius toweri | 0.054143 | −0.333427 |
| Carassius auratus | 0.080127 | 0.099953 |
| Chirostoma grandocule | 0.001050 | 0.068697 |
| Coptodon rendalli | 0.022928 | 0.067710 |
| Coptodon zillii | 0.005592 | 0.038124 |
| Ctenopharyngodon idella | −0.001050 | −0.068697 |
| Cyprinus carpio | 0.062067 | 0.143948 |
| Gambusia atrora | 0.049687 | 0.212331 |
| Gambusia marshi | −0.023292 | 0.029809 |
| Gambusia panuco | 0.029366 | −0.127961 |
| Ictiobus bubalus | −0.010438 | −0.070169 |
| Notropis boucardi | 0.013536 | −0.083357 |
| Eigenvalue | 179.29 | 19.86 |
| % of variance | 87.67 | 9.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Lendech, N.; Martínez-Falcón, A.P.; Schmitter-Soto, J.J.; Mejía-Mojica, H.; Sorani-Dalbón, V.; Cruz-Ruíz, G.I.; Mercado-Silva, N. Ichthyological Differentiation and Homogenization in the Pánuco Basin, Mexico. Diversity 2020, 12, 187. https://doi.org/10.3390/d12050187
Martínez-Lendech N, Martínez-Falcón AP, Schmitter-Soto JJ, Mejía-Mojica H, Sorani-Dalbón V, Cruz-Ruíz GI, Mercado-Silva N. Ichthyological Differentiation and Homogenization in the Pánuco Basin, Mexico. Diversity. 2020; 12(5):187. https://doi.org/10.3390/d12050187
Chicago/Turabian StyleMartínez-Lendech, Norma, Ana P. Martínez-Falcón, Juan Jacobo Schmitter-Soto, Humberto Mejía-Mojica, Valentino Sorani-Dalbón, Gabriel I. Cruz-Ruíz, and Norman Mercado-Silva. 2020. "Ichthyological Differentiation and Homogenization in the Pánuco Basin, Mexico" Diversity 12, no. 5: 187. https://doi.org/10.3390/d12050187
APA StyleMartínez-Lendech, N., Martínez-Falcón, A. P., Schmitter-Soto, J. J., Mejía-Mojica, H., Sorani-Dalbón, V., Cruz-Ruíz, G. I., & Mercado-Silva, N. (2020). Ichthyological Differentiation and Homogenization in the Pánuco Basin, Mexico. Diversity, 12(5), 187. https://doi.org/10.3390/d12050187