Critical Thermal Limits Do Not Vary between Wild-caught and Captive-bred Tadpoles of Agalychnis spurrelli (Anura: Hylidae)

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Populations and Thermal Variability
2.2. Estimates of Critical Thermal Limits
2.3. Statistical Analyses
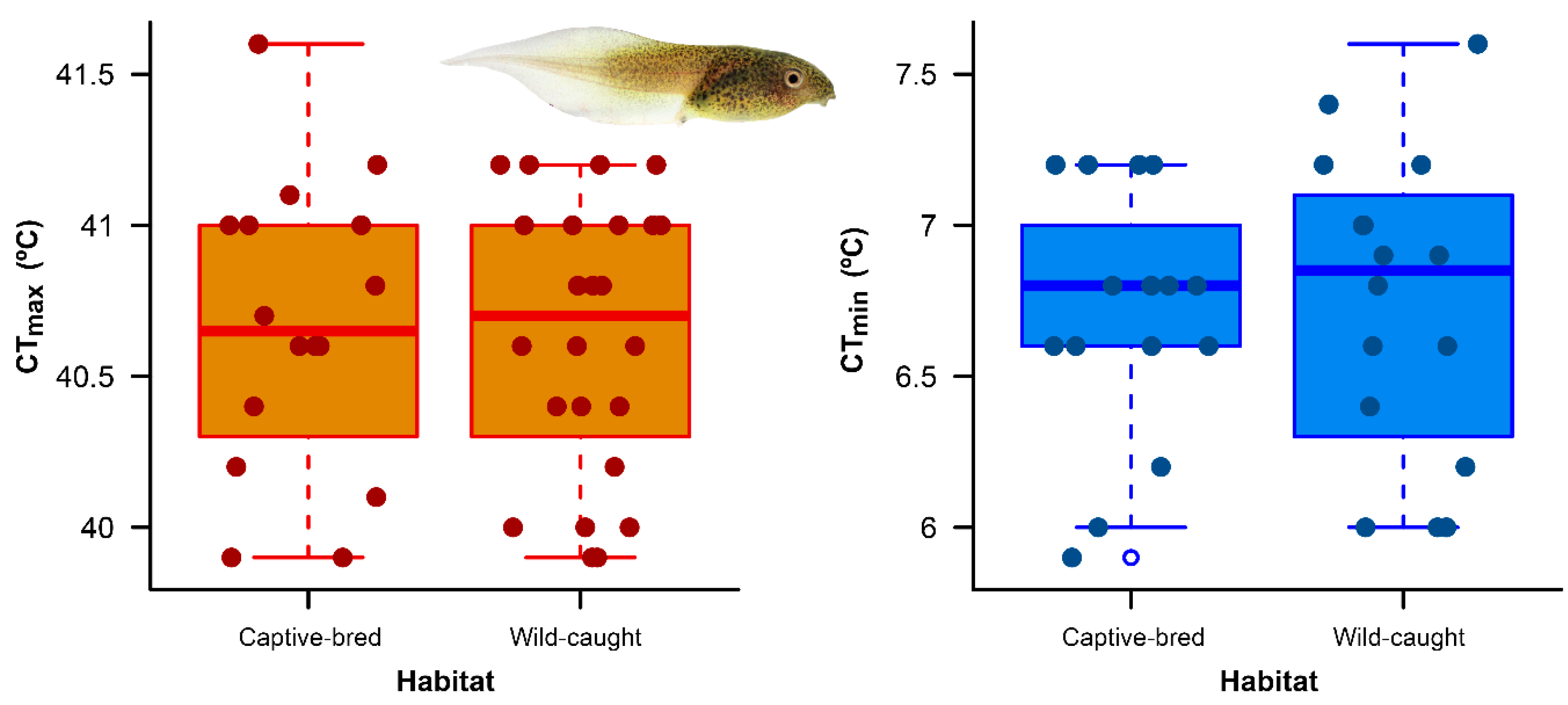
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chown, S.L.; Gaston, K.J. Macrophysiology-progress and prospects. Funct. Ecol. 2016, 30, 330–344. [Google Scholar] [CrossRef]
- Chown, S.L.; Gaston, K.J.; Robinson, D. Macrophysiology: Large-scale patterns in physiological traits and their ecological implications. Funct. Ecol. 2004, 18, 159–167. [Google Scholar] [CrossRef]
- Pintanel, P.; Tejedo, M.; Ron, S.R.; Llorente, G.A.; Merino-Viteri, A. Elevational and microclimatic drivers of thermal tolerance in Andean Pristimantis frogs. J. Biogeogr. 2019, 46, 1664–1675. [Google Scholar] [CrossRef]
- Kearney, M.; Porter, W. Mechanistic niche modelling: Combining physiological and spatial data to predict species’ ranges. Ecol. Lett. 2009, 12, 334–350. [Google Scholar] [CrossRef]
- Duarte, H.; Tejedo, M.; Katzenberger, M.; Marangoni, F.; Baldo, D.; Beltrán, J.F.; Martí, D.A.; Richter-Boix, A.; Gonzalez-Voyer, A. Can amphibians take the heat? Vulnerability to climate warming in subtropical and temperate larval amphibian communities. Glob. Chang. Biol. 2012, 18, 412–421. [Google Scholar] [CrossRef]
- Pincebourde, S.; Casas, J. Narrow safety margin in the phyllosphere during thermal extremes. Proc. Natl. Acad. Sci. USA 2019. [Google Scholar] [CrossRef]
- Floyd, R.B. Ontogenetic change in the temperature tolerance of larval Bufo marinus (Anura: Bufonidae). Comp. Biochem. Physiol. A 1983, 75, 267–271. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Woods, H.A.; Buckley, L.B.; Potter, K.A.; MacLean, H.J.; Higgins, J.K. Complex life cycles and the responses of insects to climate change. Integr. Comp. Biol. 2011, 51, 719–732. [Google Scholar] [CrossRef]
- Agudelo-Cantero, G.A.; Navas, C.A. Interactive effects of experimental heating rates, ontogeny and body mass on the upper thermal limits of anuran larvae. J. Therm. Biol. 2019, 82, 43–51. [Google Scholar] [CrossRef]
- Tüzün, N.; Op de Beeck, L.; Brans, K.I.; Janssens, L.; Stoks, R. Microgeographic differentiation in thermal performance curves between rural and urban populations of an aquatic insect. Evol. Appl. 2017, 10, 1067–1075. [Google Scholar] [CrossRef]
- Herrando-Pérez, S.; Ferri-Yáñez, F.; Monasterio, C.; Beukema, W.; Gomes, V.; Belliure, J.; Chown, S.L.; Vieites, D.R.; Araújo, M.B. Intraspecific variation in lizard heat tolerance alters estimates of climate impact. J. Anim. Ecol. 2019, 88, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.A.; Hoffmann, A.A. Thermal ramping rate influences evolutionary potential and species differences for upper thermal limits in Drosophila. Funct. Ecol. 2010, 24, 694–700. [Google Scholar] [CrossRef]
- Brattstrom, B.H. Thermal acclimation in Anuran amphibians as a function of latitude and altitude. Comp. Biochem. Physiol. 1968, 24, 93–111. [Google Scholar] [CrossRef]
- Loeschcke, V.; Sørensen, J.G. Acclimation, heat shock and hardening-a response from evolutionary biology. J. Therm. Biol. 2005, 30, 255–257. [Google Scholar] [CrossRef]
- Floyd, R.B. Effects of photoperiod and starvation on the temperature tolerance of larvae of the giant toad, Bufo marinus. Copeia 1985, 1985, 625–631. [Google Scholar] [CrossRef]
- Lutterschmidt, W.I.; Hutchison, V.H. The critical thermal maximum: Data to support the onset of spasms as the definitive end point. Can. J. Zool. 1997, 75, 1553–1560. [Google Scholar] [CrossRef]
- Chown, S.L.; Jumbam, K.R.; Sørensen, J.G.; Terblanche, J.S. Phenotypic variance, plasticity and heritability estimates of critical thermal limits depend on methodological context. Funct. Ecol. 2009, 23, 133–140. [Google Scholar] [CrossRef]
- Morgan, R.; Sundin, J.; Finnøen, M.H.; Dresler, G.; Vendrell, M.M.; Dey, A.; Sarkar, K.; Jutfelt, F. Are model organisms representative for climate change research? Testing thermal tolerance in wild and laboratory zebrafish populations. Conserv. Physiol. 2019, 7, 1–11. [Google Scholar]
- Maclean, H.J.; Kristensen, T.N.; Sørensen, J.G.; Overgaard, J. Laboratory maintenance does not alter ecological and physiological patterns among species: A Drosophila case study. J. Evol. Biol. 2018, 31, 530–542. [Google Scholar] [CrossRef]
- Kellermann, V.; Overgaard, J.; Hoffmann, A.A.; Fløjgaard, C.; Svenning, J.-C.; Loeschcke, V. Upper thermal limits of Drosophila are linked to species distributions and strongly constrained phylogenetically. Proc. Natl. Acad. Sci. USA 2012, 109, 16228–16233. [Google Scholar] [CrossRef]
- Paaijmans, K.P.; Heinig, R.L.; Seliga, R.A.; Blanford, J.I.; Blanford, S.; Murdock, C.C.; Thomas, M.B. Temperature variation makes ectotherms more sensitive to climate change. Glob. Chang. Biol. 2013, 19, 2373–2380. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, J.; Ryan, A. Developmental plasticity in the thermal tolerance of zebrafish Danio rerio. J. Fish Biol. 2006, 69, 722–734. [Google Scholar] [CrossRef]
- Jensen, A.; Alemu, T.; Alemneh, T.; Pertoldi, C.; Bahrndorff, S. Thermal acclimation and adaptation across populations in a broadly distributed soil arthropod. Funct. Ecol. 2019, 33, 833–845. [Google Scholar] [CrossRef]
- Huey, R.B.; Partridge, L.; Fowler, K. Thermal sensitivity of Drosophila melanogaster responds rapidly to laboratory natural selection. Evolution 1991, 45, 751–756. [Google Scholar] [CrossRef]
- Harshman, L.G.; Hoffmann, A.A. Laboratory selection experiments using Drosophila: What do they really tell us? Trends Ecol. Evol. 2000, 15, 32–36. [Google Scholar] [CrossRef]
- Tejedo, M.; Marangoni, F.; Pertoldi, C.; Richter-Boix, À.; Laurila, A.; Orizaola, G.; Nicieza, A.G.; Álvarez, D.; Gomez-Mestre, I. Contrasting effects of environmental factors during larval stage on morphological plasticity in post-metamorphic frogs. Clim. Res. 2010, 43, 31–39. [Google Scholar] [CrossRef]
- Enriquez-Urzelai, U.; Sacco, M.; Palacio, A.S.; Pintanel, P.; Tejedo, M.; Nicieza, A.G. Ontogenetic reduction in thermal tolerance is not alleviated by earlier developmental acclimation in Rana temporaria. Oecologia 2019, 189, 385–394. [Google Scholar] [CrossRef]
- Vázquez, D.P.; Gianoli, E.; Morris, W.F.; Bozinovic, F. Ecological and evolutionary impacts of changing climatic variability. Biol. Rev. 2017, 92, 22–42. [Google Scholar] [CrossRef]
- Kellermann, V.; Heerwaarden, B.v.; Sgrò, C.M. How important is thermal history? Evidence for lasting effects of developmental temperature on upper thermal limits in Drosophila melanogaster. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170447. [Google Scholar] [CrossRef]
- Jenkins, N.L.; Hoffmann, A.A. Genetic and maternal variation for heat resistance in Drosophila from the field. Genetics 1994, 137, 783–789. [Google Scholar]
- Ortega-Andrade, H.M. Agalychnis spurrelli Boulenger (Anura, Hylidae): Variación, distribución y sinonimia. Pap. Avulsos Zool. 2008, 48, 103–117. [Google Scholar] [CrossRef]
- Gutiérrez-Pesquera, L.M.; Tejedo, M.; Olalla-Tárraga, M.Á.; Duarte, H.; Nicieza, A.; Solé, M. Testing the climate variability hypothesis in thermal tolerance limits of tropical and temperate tadpoles. J. Biogeogr. 2016, 43, 1166–1178. [Google Scholar] [CrossRef]
- Lutterschmidt, W.I.; Hutchison, V.H. The critical thermal maximum: History and critique. Can. J. Zool. 1997, 75, 1561–1574. [Google Scholar] [CrossRef]
- Gosner, K.L. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 1960, 16, 183–190. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Vincent, R.E. Some influences of domestication upon three stocks of brook trout (Salvelinus fontinalis Mitchill). Trans. Am. Fish. Soc. 1960, 89, 35–52. [Google Scholar] [CrossRef]
- Carline, R.F.; Machung, J.F. Critical thermal maxima of wild and domestic strains of trout. Trans. Am. Fish. Soc. 2001, 130, 1211–1216. [Google Scholar] [CrossRef]
- Krebs, R.A.; Roberts, S.P.; Bettencourt, B.R.; Feder, M.E. Changes in thermotolerance and Hsp70 expression with domestication in Drosophila melanogaster. J. Evol. Biol. 2001, 14, 75–82. [Google Scholar] [CrossRef]
- Gilchrist, G.W.; Huey, R.B. The direct response of Drosophila melanogaster to selection on knockdown temperature. Heredity 1999, 83, 15–29. [Google Scholar] [CrossRef]
- Donaldson, L.R.; Olson, P.R. Development of rainbow trout brood stock by selective breeding. Trans. Am. Fish. Soc. 1957, 85, 93–101. [Google Scholar] [CrossRef]
- Santos, M.; Castañeda, L.E.; Rezende, E.L. Keeping pace with climate change: What is wrong with the evolutionary potential of upper thermal limits? Ecol. Evol. 2012, 2, 2866–2880. [Google Scholar] [CrossRef]
- Lewis, C.H.R.; Richards-Zawacki, C.L.; Ibáñez, R.; Luedtke, J.; Voyles, J.; Houser, P.; Gratwicke, B. Conserving Panamanian harlequin frogs by integrating captive-breeding and research programs. Biol. Conserv. 2019, 236, 180–187. [Google Scholar] [CrossRef]
- Huey, R.B.; Kingsolver, J.G. Evolution of thermal sensitivity of ectotherm performance. Trends Ecol. Evol. 1989, 4, 131–135. [Google Scholar] [CrossRef]
- Frazier, M.R.; Huey, R.B.; Berrigan, D. Thermodynamics constrains the evolution of insect population growth rates: “Warmer is better”. Am. Nat. 2006, 168, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.F. Evolution of the control of body temperature: Is warmer better. In Comparative Physiology: Life in Water and on Land; Dejours, P., Bolis, L., Taylor, C., Weibel, E., Eds.; Liviana: Padova, Italia, 1987; pp. 421–431. [Google Scholar]
- Becker, M.H.; Walke, J.B.; Cikanek, S.; Savage, A.E.; Mattheus, N.; Santiago, C.N.; Minbiole, K.P.C.; Harris, R.N.; Belden, L.K.; Gratwicke, B. Composition of symbiotic bacteria predicts survival in Panamanian golden frogs infected with a lethal fungus. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142881. [Google Scholar] [CrossRef]
- Bustamante, H.M.; Livo, L.J.; Carey, C. Effects of temperature and hydric environment on survival of the Panamanian Golden Frog infected with a pathogenic chytrid fungus. Integr. Zool. 2010, 5, 143–153. [Google Scholar] [CrossRef]
- Ellison, A.R.; Savage, A.E.; DiRenzo, G.V.; Langhammer, P.; Lips, K.R.; Zamudio, K.R. Fighting a losing battle: Vigorous immune response countered by pathogen suppression of host defenses in the chytridiomycosis-susceptible frog Atelopus zeteki. G3 (Bethesda) 2014, 4, 1275–1289. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Critical Thermal Limit | Model | df | AIC | BIC | LogLik | p-Value |
|---|---|---|---|---|---|---|
| CTmax | * CTmax ~ 1 | 2 | 54.026 | 57.404 | −25.013 | |
| CTmax ~ mass | 3 | 55.991 | 61.058 | −24.996 | 0.855 | |
| CTmax ~ population | 3 | 55.967 | 61.034 | −24.984 | 0.813 | |
| CTmax ~ mass + population | 4 | 57.953 | 64.708 | −24.976 | 0.967 | |
| CTmax ~ mass * population | 5 | 59.581 | 68.026 | −24.791 | 0.940 | |
| CTmin | * CTmin ~ 1 | 2 | 43.083 | 46.014 | −19.541 | |
| CTmin ~ mass | 3 | 44.747 | 49.144 | −19.373 | 0.574 | |
| CTmin ~ population | 3 | 45.043 | 49.441 | −19.522 | 0.848 | |
| CTmin ~ mass + population | 4 | 46.203 | 52.066 | −19.102 | 0.668 | |
| CTmin ~ mass * population | 5 | 44.862 | 52.191 | −17.431 | 0.267 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pintanel, P.; Tejedo, M.; Almeida-Reinoso, F.; Merino-Viteri, A.; Gutiérrez-Pesquera, L.M. Critical Thermal Limits Do Not Vary between Wild-caught and Captive-bred Tadpoles of Agalychnis spurrelli (Anura: Hylidae). Diversity 2020, 12, 43. https://doi.org/10.3390/d12020043
Pintanel P, Tejedo M, Almeida-Reinoso F, Merino-Viteri A, Gutiérrez-Pesquera LM. Critical Thermal Limits Do Not Vary between Wild-caught and Captive-bred Tadpoles of Agalychnis spurrelli (Anura: Hylidae). Diversity. 2020; 12(2):43. https://doi.org/10.3390/d12020043
Chicago/Turabian StylePintanel, Pol, Miguel Tejedo, Freddy Almeida-Reinoso, Andrés Merino-Viteri, and Luis Miguel Gutiérrez-Pesquera. 2020. "Critical Thermal Limits Do Not Vary between Wild-caught and Captive-bred Tadpoles of Agalychnis spurrelli (Anura: Hylidae)" Diversity 12, no. 2: 43. https://doi.org/10.3390/d12020043
APA StylePintanel, P., Tejedo, M., Almeida-Reinoso, F., Merino-Viteri, A., & Gutiérrez-Pesquera, L. M. (2020). Critical Thermal Limits Do Not Vary between Wild-caught and Captive-bred Tadpoles of Agalychnis spurrelli (Anura: Hylidae). Diversity, 12(2), 43. https://doi.org/10.3390/d12020043