Resource Partitioning of Sympatric African Wolves (Canis lupaster) and Side-Striped Jackals (Canis adustus) in an Arid Environment from West Africa


Abstract
:1. Introduction
2. Materials and Methods
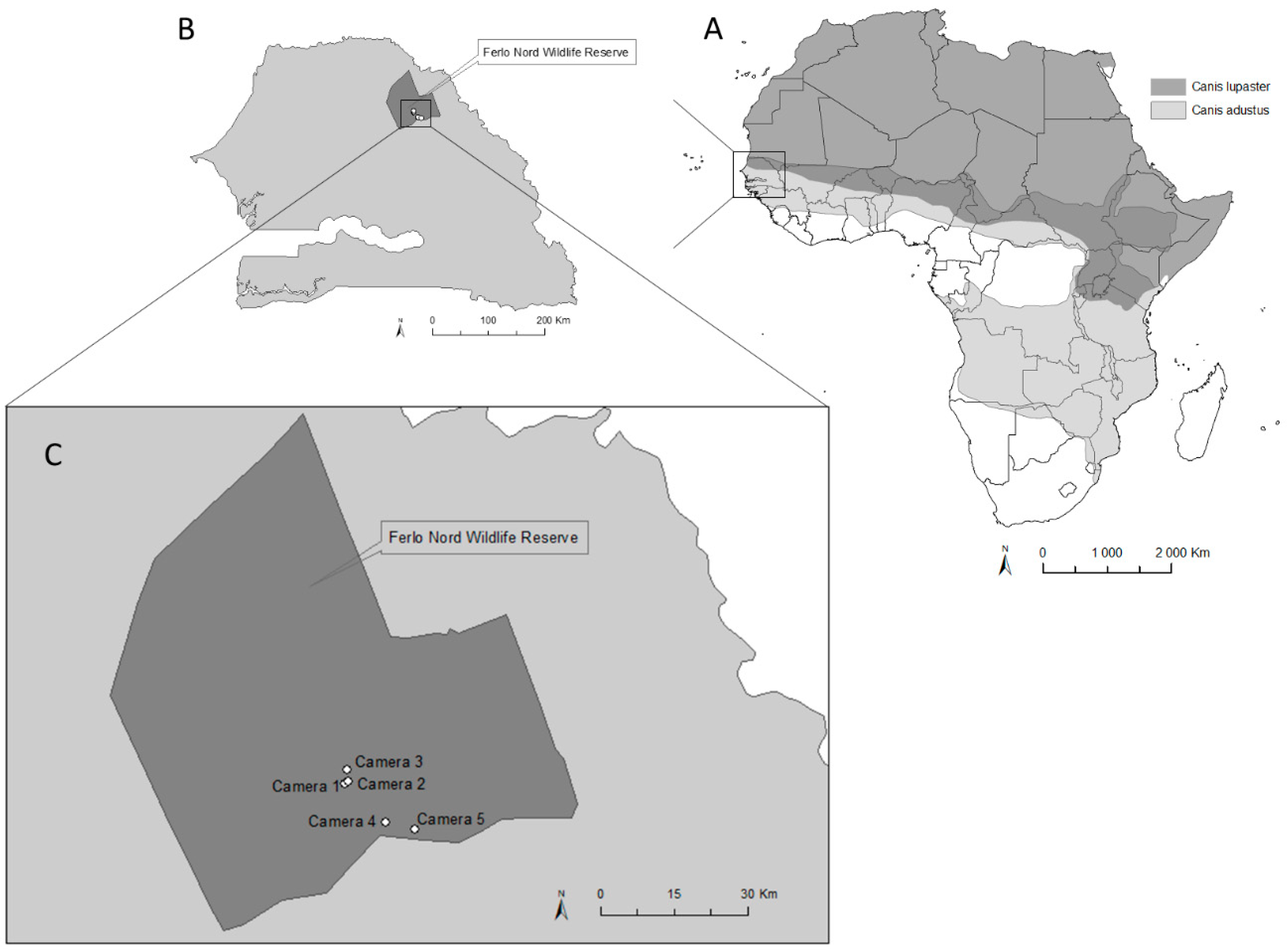
2.1. Study Area
2.2. Data Collection and Analysis
3. Results
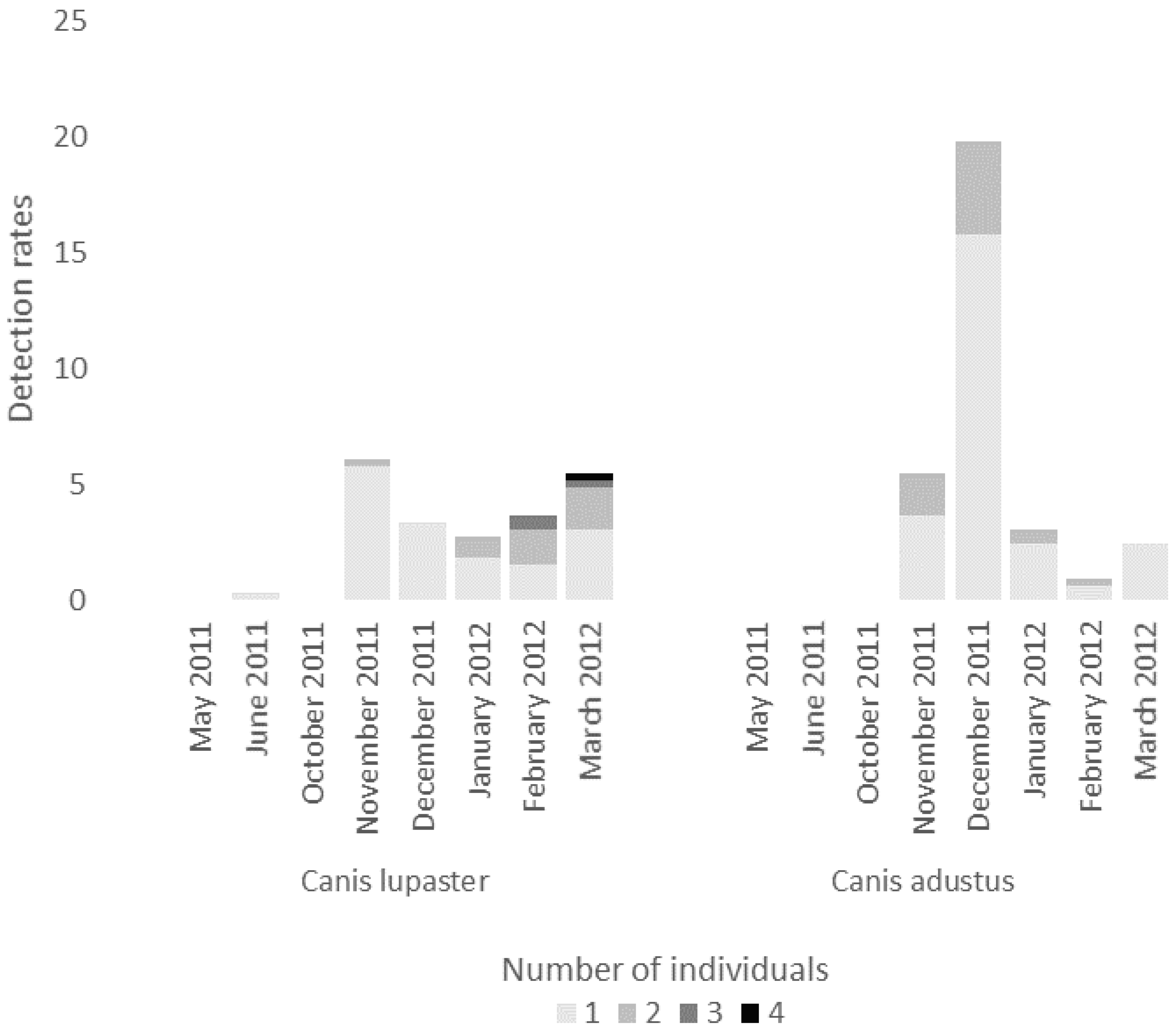
3.1. Detection Rates of Carnivore Species
3.2. Canid Group Size Near Waterholes
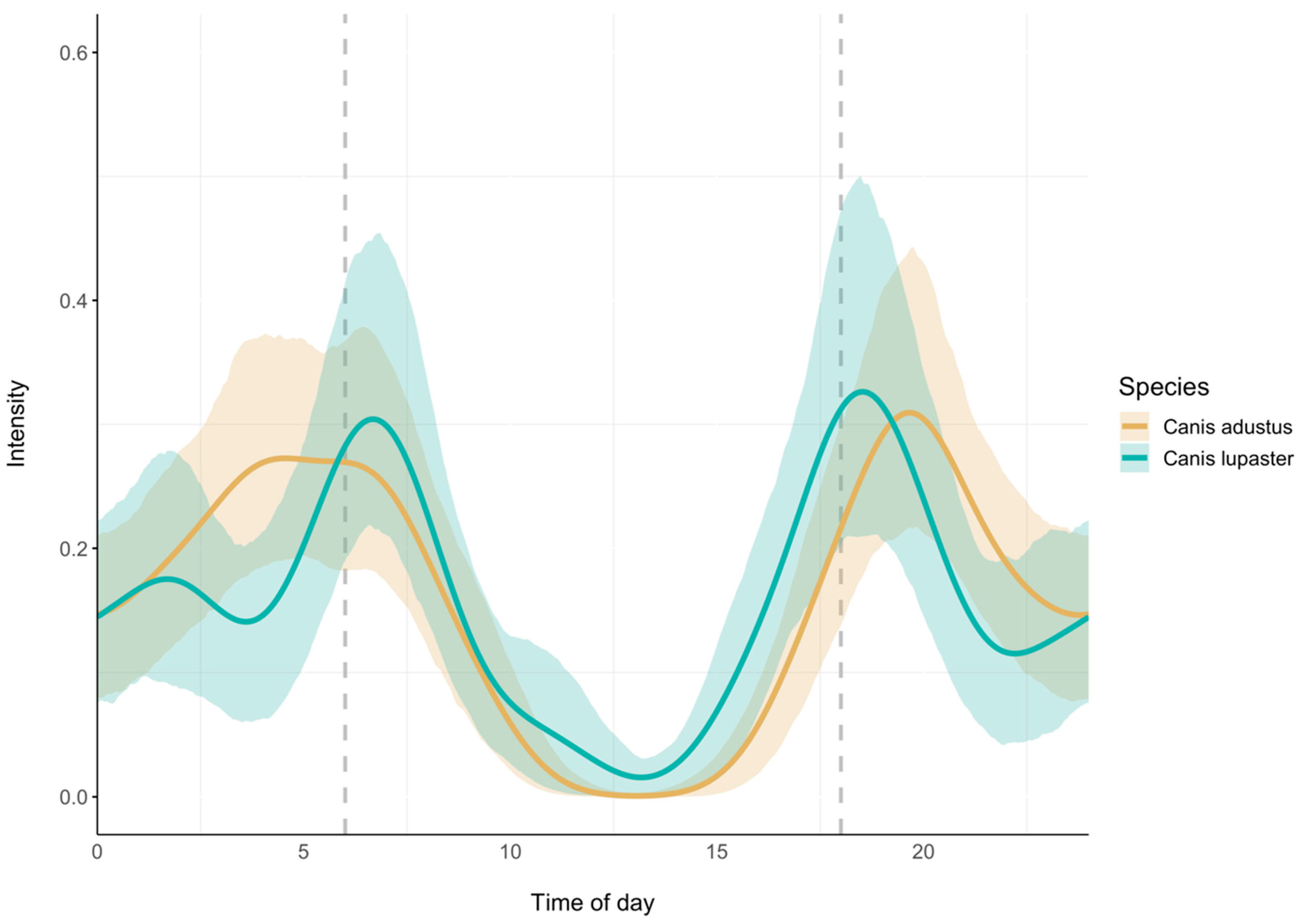
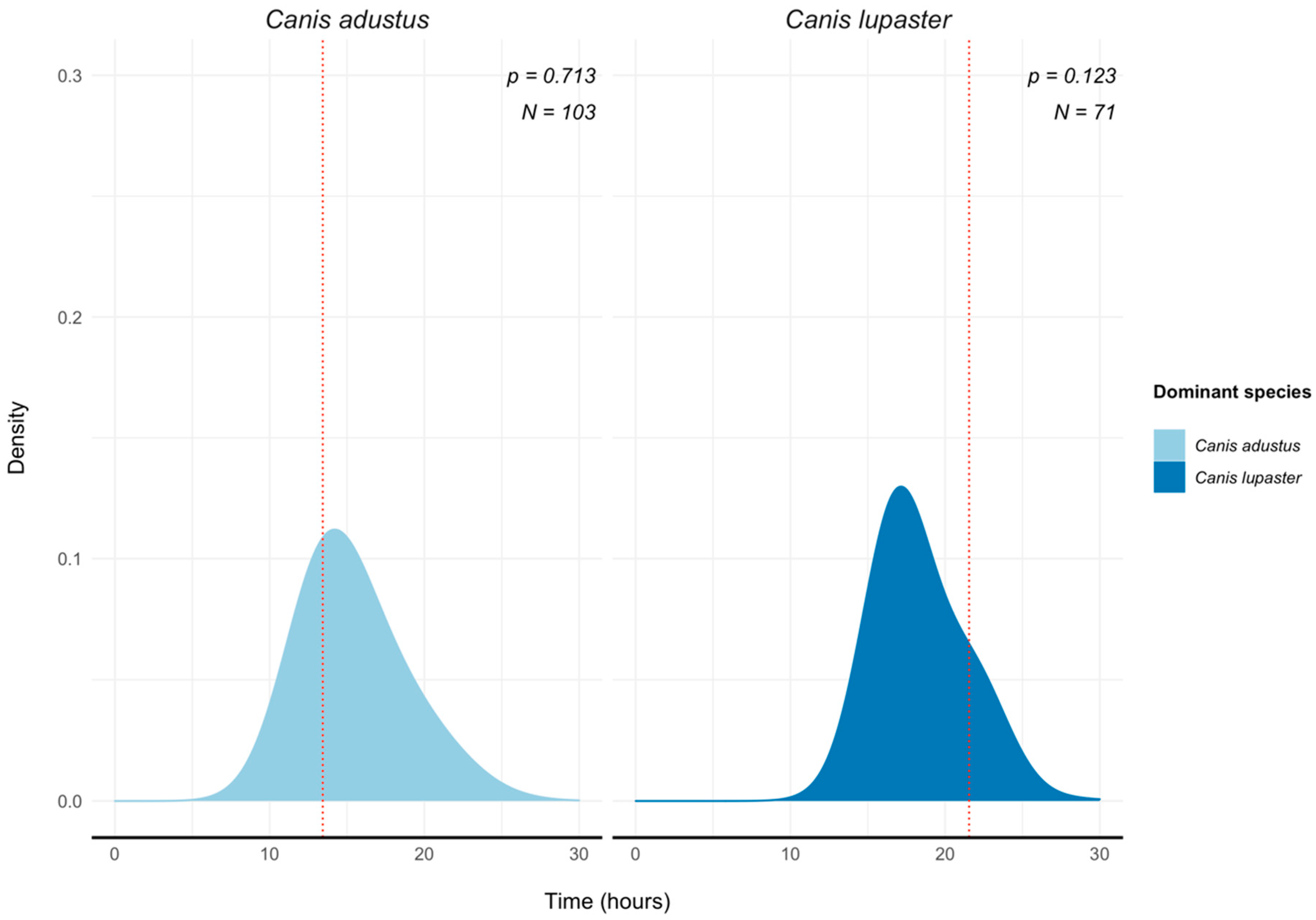
3.3. Canid Activity Patterns Near Waterholes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halle, S. Ecological relevance of daily activity patterns. In Activity Patterns in Small Mammals: An Ecological Approach; Halle, S., Stenseth, N.C., Eds.; Springer: New York, NY, USA, 2000; Volume 141, pp. 67–90. [Google Scholar]
- Kronfeld-Schor, N.; Dayan, T. Partitioning of Time as an Ecological Resource. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 153–181. [Google Scholar] [CrossRef] [Green Version]
- Linnell, J.D.C.; Strand, O. Interference interactions, co-existence and conservation of mammalian carnivores. Divers. Distrib. 2000, 6, 169–176. [Google Scholar] [CrossRef]
- Vahl, W.K.; Van Der Meer, J.; Weissing, F.J.; Van Dullemen, D.; Piersma, T. The mechanisms of interference competition: Two experiments on foraging waders. Behav. Ecol. 2005, 16, 845–855. [Google Scholar] [CrossRef] [Green Version]
- Valeix, M.; Chamaillé-Jammes, S.; Fritz, H. Interference competition and temporal niche shifts: Elephants and herbivore communities at waterholes. Oecologia 2007, 153, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Caro, T.M.; Stoner, C.J. The potential for interspecific competition among African carnivores. Biol. Conserv. 2003, 110, 67–75. [Google Scholar] [CrossRef]
- Gerber, B.D.; Karpanty, S.M.; Randrianantenaina, J. Activity patterns of carnivores in the rain forests of Madagascar: Implications for species coexistence. J. Mammal. 2012, 93, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Brito, J.C.; Godinho, R.; Martínez-Freiría, F.; Pleguezuelos, J.M.; Rebelo, H.; Santos, X.; Vale, C.G.; Velo-Antón, G.; Boratyński, Z.; Carvalho, S.B.; et al. Unravelling biodiversity, evolution and threats to conservation in the sahara-sahel. Biol. Rev. 2014, 89, 215–231. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.S.; Pitts, J.P. Identifying Pleistocene refugia in North American cold deserts using phylogeographic analyses and ecological niche modelling. Divers. Distrib. 2012, 18, 1139–1152. [Google Scholar] [CrossRef]
- Schulz, E.; Abichou, A.; Adamou, A.; Ousseïni, I.; Ballouche, A. The desert in the Sahara. Transitions and boundaries. In Palaeoecology of Africa and the Surrounding Islands; Baumhauer, R., Runge, J., Eds.; Taylor & Francis Group: London, UK, 2009; Volume 29, pp. 63–89. [Google Scholar]
- Thrash, I.; Theron, G.K.; Bothma, J.P. Dry season herbivore densities around drinking troughs in the Kruger National Park. J. Arid Environ. 1995, 29, 213–219. [Google Scholar] [CrossRef]
- Wilson, W.G.; Richards, S.A. Consuming and grouping: Recource-mediated animal aggregation. Ecol. Lett. 2000, 3, 175–180. [Google Scholar] [CrossRef]
- Macdonald, D.W.; Sillero-Zubiri, C. Biology and Conservation of Wild Canids; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Vieira, E.M.; Port, D. Niche overlap and resource partitioning between two sympatric fox species in southern Brazil. J. Zool. 2007, 272, 57–63. [Google Scholar] [CrossRef]
- Hoffmann, M.; Atickem, A. Canis lupaster. The IUCN Red List Threatened Species 2019: e.T118264888A118265889. Available online: https://dx.doi.org/10.2305/IUCN.UK.2019-1.RLTS.T118264888A118265889.en (accessed on 14 December 2020).
- Koepfli, K.P.; Pollinger, J.; Godinho, R.; Robinson, J.; Lea, A.; Hendricks, S.; Schweizer, R.M.; Thalmann, O.; Silva, P.; Fan, Z.; et al. Genome-wide evidence reveals that African and Eurasian golden jackals are distinct species. Curr. Biol. 2015, 25, 2158–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelló, J.R. Canids of the World: Wolves, Wild Dogs, Foxes, Jackals, Coyotes and their Relatives; Princeton University Press: Princetion, UK, 2018. [Google Scholar]
- Eddine, A.; Mostefai, N.; De Smet, K.; Klees, D.; Ansorge, H.; Karssene, Y.; Nowak, C.; Van Der Leer, P. Diet composition of a newly recognized canid species, the African Golden Wolf (Canis anthus), in Northern Algeria. Ann. Zool. Fennici. 2017, 54, 347–356. [Google Scholar] [CrossRef]
- Wilson, D.; Mittermeier, R.A. Handbook of the Mammals of the World. Vol. 1. Carnivores; Lynx Edicions: Barcelona, Spain, 2009. [Google Scholar]
- Karssene, Y.; Chammem, M.; Li, F.; Eddine, A.; Hermann, A.; Nouira, S. Spatial and temporal variability in the distribution, daily activity and diet of fennec fox (Vulpes zerda), red fox (Vulpes vulpes) and African golden wolf (Canis anthus) in southern Tunisia. Mamm. Biol. 2019, 95, 41–50. [Google Scholar] [CrossRef]
- Admasu, E.; Thirgood, S.J.; Bekele, A.; Laurenson, M.K. Spatial ecology of golden jackal in farmland in the Ethiopian Highlands. Afr. J. Ecol. 2004, 42, 144–152. [Google Scholar] [CrossRef]
- Gaubert, P.; Bloch, C.; Benyacoub, S.; Abdelhamid, A.; Pagani, P.; Djagoun, C.A.M.S.; Couloux, A.; Dufour, S. Reviving the African Wolf Canis lupus lupaster in North and West Africa: A Mitochondrial Lineage Ranging More than 6000 km Wide. PLoS ONE 2012, 7, e42740. [Google Scholar] [CrossRef]
- Rueness, E.K.; Asmyhr, M.G.; Sillero-Zubiri, C.; Macdonald, D.W.; Bekele, A.; Atickem, A.; Stenseth, N.C. The cryptic African wolf: Canis aureus lupaster is not a golden jackal and is not endemic to Egypt. PLoS ONE 2011, 6, e16385. [Google Scholar] [CrossRef]
- Fuller, T.K.; Biknevicius, A.R.; Kat, P.W.; Van Valkenburgh, B.; Wayne, R.K. The ecology of three sympatric jackal species in the Rift Valley of Kenya. Afr. J. Ecol. 1989, 27, 313–323. [Google Scholar] [CrossRef]
- Hall, L.K.; Larsen, R.T.; Knight, R.N.; McMillan, B.R. Feral horses influence both spatial and temporal patterns of water use by native ungulates in a semi-arid environment. Ecosphere 2018, 9, e02096. [Google Scholar] [CrossRef]
- Hein, L.; Metzger, M.J.; Leemans, R. The local impacts of climate change in the Ferlo, Western Sahel. Clim. Chang. 2009, 93, 465–483. [Google Scholar] [CrossRef] [Green Version]
- Becker, B. The contribution of wild plants to human nutrition in the Ferlo (Northern Senegal). Agrofor. Syst. 1983, 1, 257–267. [Google Scholar] [CrossRef]
- Ndiaye, I. Caracterisation Du Noyau Central De La Reserve De Faune Du Ferlo Nord.; Thecnical Report; Projet de Gestion Integree des Ecosystemes: Dakar, Senegal, 2006. [Google Scholar]
- Linkie, M.; Ridout, M.S. Assessing tiger-prey interactions in Sumatran rainforests. J. Zool. 2011, 284, 224–229. [Google Scholar] [CrossRef]
- Monterroso, P.; Alves, P.C.; Ferreras, P. Catch Me If You Can: Diel Activity Patterns of Mammalian Prey and Predators. Ethology 2013, 119, 1044–1056. [Google Scholar] [CrossRef] [Green Version]
- Ridout, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2009, 14, 322–337. [Google Scholar] [CrossRef]
- Rowcliffe, J.M.; Kays, R.; Kranstauber, B.; Carbone, C.; Jansen, P.A. Quantifying levels of animal activity using camera trap data. Methods Ecol. Evol. 2014, 5, 1170–1179. [Google Scholar] [CrossRef] [Green Version]
- Monterroso, P.; Alves, P.C.; Ferreras, P. Plasticity in circadian activity patterns of mesocarnivores in Southwestern Europe: Implications for species coexistence. Behav. Ecol. Sociobiol. 2014, 68, 1403–1417. [Google Scholar] [CrossRef] [Green Version]
- Batschelet, E. Circular Statistics in Ecology; Academic Press: London, UK, 1981. [Google Scholar]
- Rowcliffe, M. Activity: Animal Activity Statistics, Version 1.3. Available online: https://cran.r-project.org/web/packages/activity/index.html (accessed on 14 December 2020).
- Agostinelli, C.; Lund, U. R package “circular”: Circular Statistics, Version 0.4-93. Available online: https://cran.r-project.org/web/packages/circular/index.html (accessed on 14 December 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Karanth, K.U.; Srivathsa, A.; Vasudev, D.; Puri, M.; Parameshwaran, R.; Kumar, N.S. Spatio-temporal interactions facilitate large carnivore sympatry across a resource gradient. Proc. R. Soc. B Biol. Sci. 2017, 284, 20161860. [Google Scholar] [CrossRef] [Green Version]
- Moehlman, P.D. Intraspecific Variation in Canid Social Systems. In Carnivore Behavior, Ecology, and Evolution; Cornell University Press: Ithaca, NY, USA, 1989; pp. 143–163. [Google Scholar]
- Moehlman, P.D.; Hofer, H. Cooperative Breeding, Reproductive Suppression, and Body Mass in Canids. In Cooperative Breeding in Mammals; Cambridge University Press: Cambridge, UK, 1997; pp. 76–128. [Google Scholar]
- Moehlman, P.D. Jackal helpers and pup survival. Nature 1979, 277, 382–383. [Google Scholar] [CrossRef]
- Ferguson, J.W.H.; Nel, J.A.J.; Wet, M.J. Social organization and movement patterns of Black- backed jackals Canis mesomelas in South Africa. J. Zool. 1983, 199, 487–502. [Google Scholar] [CrossRef]
- Loveridge, A.J.; Macdonald, D.W. Seasonality in spatial organization and dispersal of sympatric jackals (Canis mesomelas and C. adustus): Implications for rabies management. J. Zool. 2001, 253, 101–111. [Google Scholar] [CrossRef]
- Estes, R.D. The Behavior Guide to African Mammals: Including Hoofed Mammals, Carnivores, Primates; University of California Press: Berkeley, CA, USA, 1992. [Google Scholar]
- Bingham, J.; Purchase, G.K. Reproduction in the jackals Canis adustus Sundevall, 1846, and Canis mesomelas Schreber, 1778 (Carnivora: Canidae), in Zimbabwe. Afr. Zool. 2002, 37, 21–26. [Google Scholar] [CrossRef]
- Edwards, S.; Al Awaji, M.; Eid, E.; Attum, O. Mammalian activity at artificial water sources in Dana Biosphere Reserve, southern Jordan. J. Arid Environ. 2017, 141, 5255. [Google Scholar] [CrossRef]
- Ochoa, G.V.; Chou, P.P.; Hall, L.K.; Knight, R.N.; Larsen, R.T.; McMillan, B.R. Spatial and temporal interactions between top carnivores at water sources in two deserts of western North America. J. Arid Environ. 2021, 184, 104303. [Google Scholar] [CrossRef]
- Sutherland, K.; Ndlovu, M.; Pérez-Rodríguez, A. Use of Artificial Waterholes by Animals in the Southern Region of the Kruger National Park, South Africa. Afr. J. Wildl. Res. 2018, 48, 023003. [Google Scholar] [CrossRef]
- Lucherini, M.; Reppucci, J.I.; Walker, R.S.; Villalba, M.L.; Wurstten, A.; Gallardo, G.; Iriarte, A.; Villalobos, R.; Perovic, P. Activity Pattern Segregation of Carnivores in the High Andes. J. Mammal. 2009, 90, 1404–1409. [Google Scholar] [CrossRef]
- Cook, B.I.; Smerdon, J.E.; Seager, R.; Coats, S. Global warming and 21st century drying. Clim. Dyn. 2014, 43, 2607–2627. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Activity Level (h) | 95% CI | W | p-Value |
|---|---|---|---|---|
| Canis lupaster | 11.7 ± 1.6 | 7.5–13.5 | ||
| Canis adustus | 12.3 ± 1.5 | 8.4–14.1 | ||
| Difference | −0.7 ± 2.2 | 0.09 | 0.77 | |
| Activity overlap (Δ) | 0.85 | 67.05 | 2.75 × 10−15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paúl, M.J.; Layna, J.F.; Monterroso, P.; Álvares, F. Resource Partitioning of Sympatric African Wolves (Canis lupaster) and Side-Striped Jackals (Canis adustus) in an Arid Environment from West Africa. Diversity 2020, 12, 477. https://doi.org/10.3390/d12120477
Paúl MJ, Layna JF, Monterroso P, Álvares F. Resource Partitioning of Sympatric African Wolves (Canis lupaster) and Side-Striped Jackals (Canis adustus) in an Arid Environment from West Africa. Diversity. 2020; 12(12):477. https://doi.org/10.3390/d12120477
Chicago/Turabian StylePaúl, Maria João, Jorge F. Layna, Pedro Monterroso, and Francisco Álvares. 2020. "Resource Partitioning of Sympatric African Wolves (Canis lupaster) and Side-Striped Jackals (Canis adustus) in an Arid Environment from West Africa" Diversity 12, no. 12: 477. https://doi.org/10.3390/d12120477
APA StylePaúl, M. J., Layna, J. F., Monterroso, P., & Álvares, F. (2020). Resource Partitioning of Sympatric African Wolves (Canis lupaster) and Side-Striped Jackals (Canis adustus) in an Arid Environment from West Africa. Diversity, 12(12), 477. https://doi.org/10.3390/d12120477