Stable Isotope Dynamics of Herbivorous Reef Fishes and Their Ectoparasites



Abstract
1. Introduction
2. Materials and Methods
3. Results
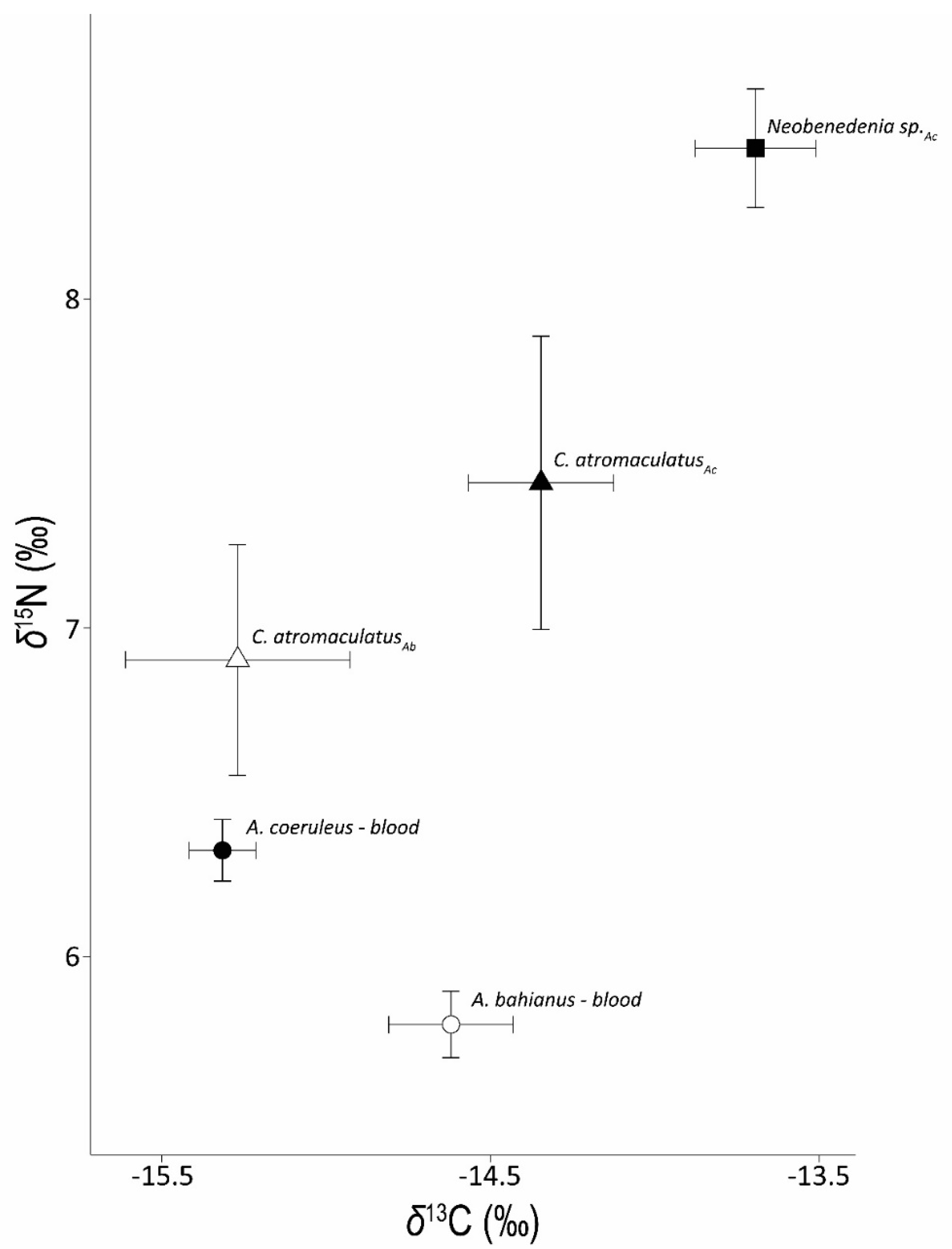
3.1. Carbon Stable Isotopes
3.2. Nitrogen Stable Isotopes
3.3. Stable Isotope Patterns over Time and with C:N
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reaka-Kudla, M.L. The global biodiversity of coral reefs: a comparison with rain forests. In Biodiversity II: Understanding and Protecting Our Biological Resources; Reaka-Kudla, M.L., Wilson, D.E., Wilson, E.O., Eds.; Joseph Henry Press: Washington, DC, USA, 1997; pp. 83–108. ISBN 0309052270. [Google Scholar]
- Roberts, C.; McClean, C.; Vernon, J.; Hawkins, J.; Allen, G.; McAllister, D.; Mittermeier, C.; Schueler, F.; Spalding, M.; Wells, F.; et al. Marine biodiversity hotspots and conservation priorities for tropical reefs. Science (80-) 2002, 295, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, N.; Brainard, R.E.; Fisher, R.; Moews, M.; Plaisance, L.; Caley, M.J. Coral reef biodiversity. In Life in the World’s Oceans; McIntyre, A.D., Ed.; John Wiley & Sons: West Sussex, UK, 2010. [Google Scholar]
- Dornelas, M.; Connolly, S.R.; Hughes, T.P. Coral reef diversity refutes the neutral theory of biodiversity. Nature 2006, 440, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Bellwood, D.R.; Streit, R.P.; Brandl, S.J.; Tebbett, S.B. The meaning of the term ‘function’ in ecology: a coral reef perspective. Funct. Ecol. 2019, 33, 948–961. [Google Scholar] [CrossRef]
- Brandl, S.J.; Rasher, D.B.; Côté, I.M.; Casey, J.M.; Darling, E.S.; Lefcheck, J.S.; Duffy, J.E. Coral reef ecosystem functioning: eight core processes and the role of biodiversity. Front. Ecol. Environ. 2019, 17, 445–454. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Dobson, A.P.; Kuris, A.M. Parasites dominate food web links. Proc. Natl. Acad. Sci. USA 2006, 103, 11211–11216. [Google Scholar] [CrossRef]
- Wood, C.L.; Byers, J.E.; Cottingham, K.L.; Altman, I.; Donahue, M.J.; Blakeslee, A.M.H. Parasites alter community structure. Proc. Natl. Acad. Sci. USA 2007, 104, 9335–9339. [Google Scholar] [CrossRef]
- Johnson, P.T.J.; Dobson, A.; Lafferty, K.D.; Marcogliese, D.J.; Memmott, J.; Orlofske, S.A.; Poulin, R.; Thieltges, D.W. When parasites become prey: ecological and epidemiological significance of eating parasites. Trends Ecol. Evol. 2010, 25, 362–371. [Google Scholar] [CrossRef]
- Rohde, K. Latitudinal gradients in species diversity: the search for the primary cause. Oikos 1992, 65, 514–527. [Google Scholar] [CrossRef]
- Rohde, K. Latitudinal gradients in species diversity and rapoport’s rule revisited: a review of recent work and what can parasites teach us about the causes of the gradients? Ecography (Cop.) 1999, 22, 593–613. [Google Scholar] [CrossRef]
- Poulin, R.; Morand, S. The diversity of parasites. Q. Rev. Biol. 2000, 75, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, G.; Grutter, A.S.; Cribb, T.H. Structure of the parasite communities of a coral reef fish assemblage (Labridae): testing ecological and phylogenetic factors. J. Parasitol. 2007, 93, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, K.D.; Allesina, S.; Arim, M.; Briggs, C.J.; De Leo, G.; Dobson, A.P.; Dunne, J.A.; Johnson, P.T.J.; Kuris, A.M.; Marcogliese, D.J.; et al. Parasites in food webs: the ultimate missing links. Ecol. Lett. 2008, 11, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Marcogliese, D.J.; Cone, D.K. Food webs: A plea for parasites. Trends Ecol. Evol. 1997, 12, 320–325. [Google Scholar] [CrossRef]
- Jephcott, T.G.; Sime-Ngando, T.; Gleason, F.H.; Macarthur, D.J. Host-parasite interactions in food webs: diversity, stability, and coevolution. Food Webs 2016, 6, 1–8. [Google Scholar] [CrossRef]
- De Meeûs, T.; Renaud, F. Parasites within the new phylogeny of eukaryotes. Trends Parasitol. 2002, 18, 247–250. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Kuris, A.M. Trophic strategies, animal diversity and body size. Trends Ecol. Evol. 2002, 17, 507–513. [Google Scholar] [CrossRef]
- Poulin, R. Evolutionary Ecology of Parasites; Princeton University Press: Princeton, NJ, USA, 2011; ISBN 9781400840809. [Google Scholar]
- Kuris, A.M.; Hechinger, R.F.; Shaw, J.C.; Whitney, K.L.; Aguirre-Macedo, L.; Boch, C.A.; Dobson, A.P.; Dunham, E.J.; Fredensborg, B.L.; Huspeni, T.C.; et al. Ecosystem energetic implications of parasite and free-living biomass in three estuaries. Nature 2008, 454, 515–518. [Google Scholar] [CrossRef]
- Preston, D.L.; Orlofske, S.A.; Lambden, J.P.; Johnson, P.T.J. Biomass and productivity of trematode parasites in pond ecosystems. J. Anim. Ecol. 2013, 82, 509–517. [Google Scholar] [CrossRef]
- Byers, J.E. Including parasites in food webs. Trends Parasitol. 2009, 25, 55–57. [Google Scholar] [CrossRef]
- Sukhdeo, M.V.K. Food Webs for parasitologists: A Review. J. Parasitol. 2010, 96, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Amundsen, P.A.; Lafferty, K.D.; Knudsen, R.; Primicerio, R.; Klemetsen, A.; Kuris, A.M. Food web topology and parasites in the pelagic zone of a subarctic lake. J. Anim. Ecol. 2009, 78, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.D.; Sukhdeo, M.V.K. Parasites alter the topology of a stream food web across seasons. Oecologia 2008, 156, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Dunne, J.A.; Lafferty, K.D.; Dobson, A.P.; Hechinger, R.F.; Kuris, A.M.; Martinez, N.D.; McLaughlin, J.P.; Mouritsen, K.N.; Poulin, R.; Reise, K.; et al. Parasites affect food web structure primarily through increased diversity and complexity. PLoS Biol. 2013, 11, e1001579. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Kimo Morris, A. Altered behavior of parasitized killifish increases susceptibility to predation by bird final hosts. Ecology 1996, 77, 1390–1397. [Google Scholar] [CrossRef]
- Hay, M.E. Fish-seaweed interaction on coral reefs: effects of herbivorous fishes and adaptation of their prey. In The Ecology of Fishes on Coral Reef; Academic Press: San Diego, CA, USA, 1991; pp. 96–119. [Google Scholar]
- Choat, J.H.; Clements, K.D.; Robbins, W.D. The trophic status of herbivorous fishes on coral reefs. Mar. Biol. 2002, 140, 613–623. [Google Scholar] [CrossRef]
- Hixon, M.A. 60 Years of coral reef fish ecology: Past, present, future. Bull. Mar. Sci. 2011, 87, 727–765. [Google Scholar] [CrossRef]
- Lobel, P.S. Herbivory by damselfishes and their role in coral reef community ecology. Bull. Mar. Sci. 1980, 30, 273–289. [Google Scholar]
- Polunin, N.V.C.; Klumpp, D.W. Algal food supply and grazer demand in a very productive coral-reef zone. J. Exp. Mar. Bio. Ecol. 1992, 164, 1–15. [Google Scholar] [CrossRef]
- Goatley, C.H.R.; Bellwood, D.R. Biologically mediated sediment fluxes on coral reefs: sediment removal and off-reef transportation by the surgeonfish Ctenochaetus striatus. Mar. Ecol. Prog. Ser. 2010, 415, 237–245. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Goatley, C.H.R.; Brandl, S.J.; Bellwood, O. Fifty million years of herbivory on coral reefs: fossils, fish and functional innovations. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133046. [Google Scholar] [CrossRef] [PubMed]
- Marshell, A.; Mumby, P.J. The role of surgeonfish (Acanthuridae) in maintaining algal turf biomass on coral reefs. J. Exp. Mar. Bio. Ecol. 2015, 473, 152–160. [Google Scholar] [CrossRef]
- Tebbett, S.B.; Goatley, C.H.R.; Bellwood, D.R. Clarifying functional roles: algal removal by the surgeonfishes Ctenochaetus striatus and Acanthurus nigrofuscus. Coral Reefs 2017, 36, 803–813. [Google Scholar] [CrossRef]
- Randall, J.E. Surgeonfishes of the World; Bishop Museum Press: Honolulu, HI, USA, 2001; ISBN 1566475619. [Google Scholar]
- Tilghman, G.C.; Klinger-Bowen, R.E.; Francis-Floyd, R. Feeding electivity indices in surgeonfish (Acanthuridae) of the Florida keys. Aquarium Sci. Conserv. 2001, 3, 215–223. [Google Scholar] [CrossRef]
- Miyake, S.; Ngugi, D.K.; Stingl, U. Diet strongly influences the gut microbiota of surgeonfishes. Mol. Ecol. 2015, 24, 656–672. [Google Scholar] [CrossRef]
- Duran, A.; Adam, T.C.; Palma, L.; Moreno, S.; Collado-Vides, L.; Burkepile, D.E. Feeding behavior in Caribbean surgeonfishes varies across fish size, algal abundance, and habitat characteristics. Mar. Ecol. 2019, 40. [Google Scholar] [CrossRef]
- Lo, C.M.; Morgan, J.A.T.; Galzin, R.; Cribb, T.H. Identical digeneans in coral reef fishes from French Polynesia and the Great Barrier Reef (Australia) demonstrated by morphology and molecules. Int. J. Parasitol. 2001, 31, 1573–1578. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Nemeth, D.; McCammon, A.; Williams, E.H., Jr. Habitat and species differences in prevalence and intensity of Neobenedenia melleni (Monogenea: Capsalidae) on sympatric Caribbean surgeonfishes (Acanthuridae). J. Parasitol. 2009, 95, 63–68. [Google Scholar] [CrossRef]
- Bernal, M.A.; Floeter, S.R.; Gaither, M.R.; Longo, G.O.; Morais, R.; Ferreira, C.E.L.; Vermeij, M.J.A.; Rocha, L.A. High prevalence of dermal parasites among coral reef fishes of Curaçao. Mar. Biodivers. 2016, 46, 67–74. [Google Scholar] [CrossRef]
- Santos, T.R.N.; Sikkel, P.C. Habitat associations of fish-parasitic gnathiid isopods in a shallow reef system in the central Philippines. Mar. Biodivers. 2017, 49, 83–96. [Google Scholar] [CrossRef]
- Bshary, R.; Grutter, A.S. Parasite distribution on client reef fish determines cleaner fish foraging patterns. Mar. Ecol. Prog. Ser. 2002, 235, 217–222. [Google Scholar] [CrossRef]
- McCammon, A.; Sikkel, P.C.; Nemeth, D. Effects of three Caribbean cleaner shrimps on ectoparasitic monogeneans in a semi-natural environment. Coral Reefs 2010, 29, 419–426. [Google Scholar] [CrossRef]
- Huebner, L.K.; Chadwick, N.E. Patterns of cleaning behaviour on coral reef fish by the anemoneshrimp Ancylomenes pedersoni. J. Mar. Biol. Assoc. UK 2012, 92, 1557–1562. [Google Scholar] [CrossRef]
- Fry, B. Stable Isotope Ecology; Springer: New York, NY, USA, 2006. [Google Scholar]
- Boecklen, W.J.; Yarnes, C.T.; Cook, B.A.; James, A.C. On the use of stable isotopes in trophic ecology. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 411–440. [Google Scholar] [CrossRef]
- Deniro, M.J.; Epstein, S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Acta 1981, 45, 341–351. [Google Scholar] [CrossRef]
- McCutchan, J.H.; Lewis, W.M.; Kendall, C.; McGrath, C.C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 2003, 102, 378–390. [Google Scholar] [CrossRef]
- Minagawa, M.; Wada, E. Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochim. Cosmochim. Acta 1984, 48, 1135–1140. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: model, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Hatcher, M.J.; Dunn, A.M. Parasites in Ecological Communities: From Interactions to Ecosystems; Cambridge University Press: Cambridge, UK, 2011; ISBN 9780511987359. [Google Scholar]
- Pinnegar, J.K.; Campbell, N.; Polunin, N.V.C. Unusual stable isotope fractionation patterns observed for fish host-parasite trophic relationships. J. Fish Biol. 2001, 59, 494–503. [Google Scholar] [CrossRef]
- Deudero, S.; Pinnegar, J.K.; Polunin, N.V.C. Insights into fish host-parasite trophic relationships revealed by stable isotope analysis. Dis. Aquat. Organ. 2002, 52, 77–86. [Google Scholar] [CrossRef]
- Voigt, C.C.; Kelm, D.H. Host preferences of bat flies: following the bloody path of stable isotopes in a host–parasite food chain. Can. J. Zool. 2006, 84, 397–403. [Google Scholar] [CrossRef]
- Schmidt, O.; Dautel, H.; Newton, J.; Gray, J.S. Natural isotope signatures of host blood are replicated in moulted ticks. Ticks Tick. Borne. Dis. 2011, 2, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Demopoulos, A.W.J.; Sikkel, P.C. Enhanced understanding of ectoparasite-host trophic linkages on coral reefs through stable isotope analysis. Int. J. Parasitol. 2015, 4, 125–134. [Google Scholar] [CrossRef]
- Jenkins, W.G.; Demopoulos, A.W.J.; Sikkel, P.C. Host feeding ecology and trophic position significantly influence isotopic discrimination between a generalist ectoparasite and its hosts: implications for parasite-host trophic studies. Food Webs 2018, 16. [Google Scholar] [CrossRef]
- Thieltges, D.W.; Goedknegt, M.A.; O’Dwyer, K.; Senior, A.M.; Kamiya, T. Parasites and stable isotopes: a comparative analysis of isotopic discrimination in parasitic trophic interactions. Oikos 2019, 128, 1329–1339. [Google Scholar] [CrossRef]
- Gilbert, B.M.; Nachev, M.; Jochmann, M.A.; Schmidt, T.C.; Köster, D.; Sures, B.; Avenant-Oldewage, A. You are how you eat: differences in trophic position of two parasite species infecting a single host according to stable isotopes. Parasitol. Res. 2020, 119, 1393–1400. [Google Scholar] [CrossRef]
- Weston, M.J. Investigating trophic interactions between parasites and their marine fish hosts using stable isotope analysis. Master’s Thesis, University of Cape Town, Cape Town, South Africa, 2017. [Google Scholar]
- Whittington, I.D. Diversity “down under”: monogeneans in the Antipodes (Australia) with a prediction of monogenean biodiversity worldwide. Int. J. Parasitol. 1998, 28, 1481–1493. [Google Scholar] [CrossRef]
- Whittington, I.D. The capsalidae (Monogenea: Monopisthocotylea): A review of diversity, classification and phylogeny with a note about species complexes. Folia Parasitologica 2004, 51, 109–122. [Google Scholar] [CrossRef]
- Montero, F.E.; Crespo, S.; Padrós, F.; De La Gándara, F.; García, A.; Raga, J.A. Effects of the gill parasite Zeuxapta seriolae (Monogenea: Heteraxinidae) on the amberjack Seriola dumerili Risso (Teleostei: Carangidae). Aquaculture 2004, 232, 153–163. [Google Scholar] [CrossRef]
- Thoney, D.A.; Hargis, W.J. Monogenea (platyhelminthes) as hazards for fish in confinement. Annu. Rev. Fish Dis. 1991, 1, 133–153. [Google Scholar] [CrossRef]
- Cowell, L.E.; Watanabe, W.O.; Head, W.D.; Grover, J.J.; Shenker, J.M. Use of tropical cleaner fish to control the ectoparasite Neobenedenia melleni (Monogenea: Capsalidae) on seawater-cultured Florida red tilapia. Aquaculture 1993, 113, 189–200. [Google Scholar] [CrossRef]
- Grutter, A.S.; Deveney, M.R.; Whittington, I.D.; Lester, R.J.G. The effect of the cleaner fish Labroides dimidiatus on the capsalid monogenean Benedenia lolo parasite of the labrid fish Hemigymnus melapterus. J. Fish Biol. 2002, 61, 1098–1108. [Google Scholar] [CrossRef]
- Becker, J.H.; Grutter, A.S. Cleaner shrimp do clean. Coral Reefs 2004, 23, 515–520. [Google Scholar] [CrossRef]
- Sures, B.; Nachev, M.; Gilbert, B.M.; Dos Santos, Q.M.; Jochmann, M.A.; Köster, D.; Schmidt, T.C.; Avenant-Oldewage, A. The monogenean Paradiplozoon ichthyoxanthon behaves like a micropredator on two of its hosts, as indicated by stable isotopes. J. Helminthol. 2019, 93, 71–75. [Google Scholar] [CrossRef]
- Avenant-Oldewage, A.; Le Roux, L.E.; Mashego, S.N.; Van Vuuren, B.J. Paradiplozoon ichthyoxanthon n. sp. (Monogenea: Diplozoidae) from Labeobarbus aeneus (Cyprinidae) in the Vaal River, South Africa. J. Helminthol. 2014, 88, 166–172. [Google Scholar] [CrossRef]
- Humes, A.G. How many copepods? In Ecology and Morphology of Copepods; Ferrari, F.D., Bradley, B.P., Eds.; Springer: Dordrecht, Netherlands, 1994; pp. 1–7. [Google Scholar]
- Sikkel, P.C.; Welicky, R.L. The Ecological Significance of Parasitic Crustaceans. In Parasitic Crustacea. Zoological Monographs, vol 3; Smit, N., Bruce, N., Hadfield, K., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2019; pp. 421–477. [Google Scholar]
- Gilbert, B.M.; Nachev, M.; Jochmann, M.A.; Schmidt, T.C.; Köster, D.; Sures, B.; Avenant-Oldewage, A. Stable isotope analysis spills the beans about spatial variance in trophic structure in a fish host – parasite system from the Vaal River System, South Africa. Int. J. Parasitol. Parasites Wildl. 2020, 12, 134–141. [Google Scholar] [CrossRef]
- Whittington, I.D.; Horton, M.A. A revision of neobenedenia yamaguti, 1963 (Monogenea: Capsalidae) including a redescription of N. melleni (maccallum, 1927) yamaguti, 1963. J. Nat. Hist. 1996, 30, 1113–1156. [Google Scholar] [CrossRef]
- Hirazawa, N.; Mitsuboshi, T.; Hirata, T.; Shirasu, K. Susceptibility of spotted halibut Verasper variegatus (Pleuronectidae) to infection by the monogenean Neobenedenia girellae (Capsalidae) and oral therapy trials using praziquantel. Aquaculture 2004, 238, 83–95. [Google Scholar] [CrossRef]
- Hirazawa, N.; Takano, R.; Hagiwara, H.; Noguchi, M.; Narita, M. The influence of different water temperatures on Neobenedenia girellae (Monogenea) infection, parasite growth, egg production and emerging second generation on amberjack Seriola dumerili (Carangidae) and the histopathological effect of this parasite on fish skin. Aquaculture 2010, 299, 2–7. [Google Scholar] [CrossRef]
- Dyer, W.G.; Williams, E.H.; Bunkley-Williams, L. Neobenedenia pargueraensis n. sp. (Monogenea: Capsalidae) from the Red Hind, Epinephelus guttatus, and comments about Neobenedenia melleni. J. Parasitol. 1992, 78, 399. [Google Scholar] [CrossRef]
- Bullard, S.; Benz, G.; Overstreet, R.; Williams, E.; Hemdal, J. Six new host records and an updated list of wild hosts for Neobenedenia melleni (MacCallum) (Monogenea: Capsalidae). Comp. Parasitol. 2000, 67, 190–196. [Google Scholar]
- Loerch, S.M.; McCammon, A.M.; Sikkel, P.C. Low susceptibility of invasive Indo-Pacific lionfish Pterois volitans to ectoparasitic Neobenedenia in the eastern Caribbean. Environ. Biol. Fishes 2015, 98, 1979–1985. [Google Scholar] [CrossRef]
- Cressey, R.F. Parasitic copepods from the Gulf of Mexico and Caribbean Sea, III: Caligus. Smithson. Contrib. Zool. 1991, 1–53. [Google Scholar] [CrossRef]
- Dojiri, M.; Ho, J.S. Systematics of the Caligidae, Copepods Parasitic on Marine Fishes; Brill Academic Publishers: Leiden, Netherlands, 2013; Volume 18, ISBN 9789004204249. [Google Scholar]
- Morales-Serna, F.N.; Medina-Guerrero, R.M.; Fajer-Avila, E.J. Sea lice (Copepoda: Caligidae) parasitic on fishes reported from the Neotropical region. Neotrop. Biodivers. 2016, 2, 141–150. [Google Scholar] [CrossRef]
- Johnson, S.C.; Treasurer, J.W.; Bravo, S.; Nagasawa, K.; Kabata, Z. A Review of the impact of parasitic copepods on marine aquaculture. Zool. Stud. 2004, 43, 229–243. [Google Scholar]
- Gonçalves, A.T.; Farlora, R.; Gallardo-Escárate, C. Transcriptome survey of the lipid metabolic pathways involved in energy production and ecdysteroid synthesis in the salmon louse Caligus rogercresseyi (Crustacea: Copepoda). Comp. Biochem. Physiol. Part B 2014, 176, 9–17. [Google Scholar] [CrossRef]
- Rocha, L.A.; Bass, A.L.; Robertson, D.R.; Bowen, B.W. Adult habitat preferences, larval dispersal, and the comparative phylogeography of three Atlantic surgeonfishes (Teleostei: Acanthuridae). Mol. Ecol. 2002, 11, 243–251. [Google Scholar] [CrossRef]
- Hobson, K.A.; Gloutney, M.L.; Gibbs, H.L. Preservation of blood and tissue samples for stable-carbon and stable-nitrogen isotope analysis. Can. J. Zool. 1997, 75, 1720–1723. [Google Scholar] [CrossRef]
- Syväranta, J.; Vesala, S.; Rask, M.; Ruuhijärvi, J.; Jones, R.I. Evaluating the utility of stable isotope analyses of archived freshwater sample materials. Hydrobiologia 2008, 600, 121–130. [Google Scholar] [CrossRef]
- Gloutney, M.L.; Hobson, K.A. Field preservation techniques for the analysis of stable-carbon and nitrogen isotope ratios in eggs. J. Field Ornithol. 1998, 69, 223–227. [Google Scholar]
- Demopoulos, A.W.J.; Gualtieri, D.; Kovacs, K. Food-web structure of seep sediment macrobenthos from the Gulf of Mexico. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 1972–1981. [Google Scholar] [CrossRef]
- Polissar, P.J.; Fulton, J.M.; Junium, C.K.; Turich, C.C.; Freeman, K.H. Measurement of 13 C and 15 N isotopic composition on nanomolar quantities of C and N. Anal. Chem. 2009, 81, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montaña, C.G. Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using {lme4}. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 August 2020).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd edition; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. 2020. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 15 August 2020).
- Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 15 August 2020).
- Fanelli, E.; Cartes, J.E.; Rumolo, P.; Sprovieri, M. Food-web structure and trophodynamics of mesopelagic-suprabenthic bathyal macrofauna of the Algerian Basin based on stable isotopes of carbon and nitrogen. Deep. Res. Part I Oceanogr. Res. Pap. 2009, 56, 1504–1520. [Google Scholar] [CrossRef]
- Welicky, R.L.; Demopoulos, A.W.J.; Sikkel, P.C. Host-dependent differences in resource use associated with Anilocra spp. parasitism in two coral reef fishes, as revealed by stable carbon and nitrogen isotope analyses. Mar. Ecol. 2017, 38. [Google Scholar] [CrossRef]
- Stapp, P.; Salkeld, D.J. Inferring host-parasite relationships using stable isotopes: implications for disease transmission and host specificity. Ecology 2009, 90, 3268–3273. [Google Scholar] [CrossRef]
- DeNiro, M.J.; Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 1978, 42, 495–506. [Google Scholar] [CrossRef]
- Carter, W.A.; Bauchinger, U.; McWilliams, S.R. The importance of isotopic turnover for understanding key aspects of animal ecology and nutrition. Diversity 2019, 11, 84. [Google Scholar] [CrossRef]
- Buchheister, A.; Latour, R.J. Turnover and fractionation of carbon and nitrogen stable isotopes in tissues of a migratory coastal predator, summer flounder (Paralichthys dentatus). Can. J. Fish. Aquat. Sci. 2010, 67, 445–461. [Google Scholar] [CrossRef]
- Yohannes, E.; Grimm, C.; Rothhaupt, K.-O.; Behrmann-Godel, J. The effect of parasite infection on stable isotope turnover rates of δ15n, δ13c and δ34s in multiple tissues of eurasian perch perca fluviatilis. PLoS ONE 2017, 12, e0169058. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, P.B.; Flecker, A.S. Rapid turnover of tissue nitrogen of primary consumers in tropical freshwaters. Oecologia 2006, 148, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Matley, J.K.; Fisk, A.T.; Tobin, A.J.; Heupel, M.R.; Simpfendorfer, C.A. Diet-tissue discrimination factors and turnover of carbon and nitrogen stable isotopes in tissues of an adult predatory coral reef fish, Plectropomus leopardus. Rapid Commun. Mass Spectrom. 2016, 30, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Brazenor, A.K.; Hutson, K.S. Effects of temperature and salinity on the life cycle of Neobenedenia sp. (Monogenea: Capsalidae) infecting farmed barramundi (Lates calcarifer). Parasitol. Res. 2015, 114, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- González, L.; Carvajal, J. Life cycle of Caligus rogercresseyi, (Copepoda: Caligidae) parasite of Chilean reared salmonids. Aquaculture 2003, 220, 101–117. [Google Scholar] [CrossRef]
- Hobson, K.A.; Alisauskas, R.T.; Clark, R.G. Stable-nitrogen isotope enrichment in avian tissues due to fasting and nutritional stress: implications for isotopic analyses of diet. Condor 1993, 95, 394. [Google Scholar] [CrossRef]
- Hobson, K.A.; Clark, R.G. Assessing avian diets using sable isotopes II: factors influencing diet-tissue fractionation. Condor 1992, 94, 189–197. [Google Scholar] [CrossRef]
- Sarakinos, H.C.; Johnson, M.L.; Vander Zanden, M.J. A synthesis of tissue-preservation effects on carbon and nitrogen stable isotope signatures. Can. J. Zool. 2002, 80, 381–387. [Google Scholar] [CrossRef]
- Chouvelon, T.; Chappuis, A.; Bustamante, P.; Lefebvre, S.; Mornet, F.; Guillou, G.; Violamer, L.; Dupuy, C. Trophic ecology of European sardine Sardina pilchardus and European anchovy Engraulis encrasicolus in the Bay of Biscay (north-east Atlantic) inferred from δ13C and δ15N values of fish and identified mesozooplanktonic organisms. J. Sea Res. 2014, 85, 277–291. [Google Scholar] [CrossRef]
- Krueger, H.W.; Sullivan, C.H. Models for carbon isotope fractionation between diet and bone. In Stable Isotopes in Nutrition; Turnlund, J.R., Johnson, P.E., Eds.; American Chemical Society: Washington, D.C., USA, 1984; pp. 205–220. [Google Scholar]
- Vanderklift, M.A.; Ponsard, S. Sources of variation in consumer-diet δ15N enrichment: a meta-analysis. Oecologia 2003, 136, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Robbins, C.T.; Felicetti, L.A.; Sponheimer, M. The effect of dietary protein quality on nitrogen isotope discrimination in mammals and birds. Oecologia 2005, 144, 534–540. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Response Variable | Variables Tested | X2 | df | p-Value |
|---|---|---|---|---|
| δ13C | Sample type (blood, monogenean, copepod) | 70.7032 | 2 | <0.001 |
| Host Species (A. coeruleus, A. bahianus) | 1.9597 | 1 | 0.162 | |
| Sample type X host species | 24.6094 | 1 | <0.001 | |
| δ15N | Sample type (blood, monogenean, copepod) | 16.206 | 2 | <0.001 |
| Host Species (A. coeruleus, A. bahianus) | 6.3899 | 1 | 0.011 | |
| Sample type X host species | 0.4252 | 1 | 0.514 | |
| ∆13C parasite-host blood | Parasite type (monogenean, copepod) | 25.071 | 1 | <0.001 |
| Host Species (A. coeruleus, A. bahianus) | 18.68 | 1 | <0.002 | |
| Parasite mass | 0.036 | 1 | 0.849 | |
| Host blood δ13C | 2.751 | 1 | 0.097 | |
| Host species X parasite mass | 0.85 | 1 | 0.357 | |
| Parasite type X parasite mass | 8.917 | 1 | 0.003 | |
| Host Species X host blood δ13C | 1.811 | 1 | 0.178 | |
| Parasite type X host blood δ13C | 0.4 | 1 | 0.527 | |
| ∆15N parasite-host blood | Parasite type (monogenean, copepod) | 0.247 | 1 | 0.619 |
| Host Species (A. coeruleus, A. bahianus) | 8.106 | 1 | 0.004 | |
| Parasite mass | 10.855 | 1 | 0.001 | |
| Host blood δ15N | 0.641 | 1 | 0.423 | |
| Host species X parasite mass | 0.792 | 1 | 0.373 | |
| Parasite type X parasite mass | 6.577 | 1 | 0.010 | |
| Host Species X host blood δ15N | 1.928 | 1 | 0.165 | |
| Parasite type X host blood δ15N | 11.769 | 1 | 0.001 |
| Host | δ13C | δ15N | C:N | Parasite | δ13C | ∆13C | δ15N | ∆15N | C:N |
|---|---|---|---|---|---|---|---|---|---|
| A. coeruleus | −15.3 ± 0.1 | 6.3 ± 0.1 | 3.7 ± 0.0 | C. atromaculatus | −14.4 ± 0.2 | 1.0 ± 0.2 | 7.4 ± 0.4 | 0.9 ± 0.4 | 3.0 ± 0.1 |
| (−17.4 to −13.5, n = 63) | (4.5 to 7.9, n = 64) | (3.3 to 4.4, n = 64) | (−16.3 to −12.2, n = 23) | (−0.2 to 3.7, n = 22) | (3.1 to 10.9, n = 24) | (−4.5 to 4.2, n = 23) | (2.5 to 3.4, n = 14) | ||
| Neobenedenia sp. | −13.7 ± 0.2 | 1.9 ± 0.1 | 8.5 ± 0.2 | 2.0 ± 0.1 | 3.3 ± 0.0 | ||||
| (−17.1 to −10.5, n = 59) | (0.6 to 3.4, n = 44) | (4.2 to 12.6, n = 85) | (−0.1 to 5.2, n = 70) | (3.1 to 3.8, n = 18) | |||||
| A. bahianus | −14.6 ± 0.2 | 5.8 ± 0.1 | 3.7 ± 0.0 | C. atromaculatus | −15.3 ± 0.3 | −0.6 ± 0.1 | 6.9 ± 0.3 | 1.4 ± 0.3 | 3.1 ± 0.2 |
| (−16.3 to −12.4, n = 33) | (4.7 to 7.0, n = 31) | (3.5 to 4.0, n = 33) | (−17.9 to −12.2, n = 21) | (−1.8 to 0.1, n = 15) | (3.4 to 10.0, n = 22) | (−1.0 to 3.6, n = 17) | (2.47 to 4.0, n = 6) |
| Response Variable | Predictor Variable | Estimate ± SE | t-Value | p-Value |
|---|---|---|---|---|
| δ13C | (Intercept) | −15.4 ± 0.2 | −93.579 | <0.001 |
| C. atromaculatus | 0.9 ± 0.2 | 3.982 | 0.009 | |
| Neobenedenia sp. | 1.6 ± 0.2 | 9.103 | 0.002 | |
| A. bahianus | 0.9 ± 0.3 | 3.214 | 0.004 | |
| C. atromaculatus X A. bahianus | −1.6 ± 0.3 | −4.961 | <0.001 | |
| δ15N | (Intercept) | 6.7 ± 0.4 | 18.213 | <0.001 |
| C. atromaculatus | 0.6 ± 0.5 | 1.344 | 0.197 | |
| Neobenedenia sp. | 1.4 ± 0.4 | 3.467 | 0.003 | |
| A. bahianus | −0.8 ± 0.3 | −2.524 | 0.013 | |
| ∆13C | (Intercept) | −2.1 ± 1.5 | −1.412 | 0.164 |
| A. bahianus | −1.5 ± 0.3 | −5.075 | <0.001 | |
| Neobenedenia sp. | 1.5 ± 0.4 | 4.073 | 0.001 | |
| Host blood δ13C | −0.2 ± 0.1 | −1.792 | 0.08 | |
| Parasite mass | 34.7 ± 15.6 | 2.226 | 0.029 | |
| Neobenedenia sp. X Parasite mass | −40.3 ± 18.6 | −2.168 | 0.037 | |
| ∆15N | (Intercept) | 3.5 ± 1.6 | 2.161 | 0.033 |
| A. bahianus | 0.2 ± 0.4 | 0.379 | 0.705 | |
| Neobenedenia sp. | −3 ± 2 | −1.55 | 0.124 | |
| Parasite mass | 80.7 ± 19.4 | 4.152 | <0.001 | |
| Host blood δ15N | −0.6 ± 0.2 | −2.37 | 0.02 | |
| Neobenedenia sp. X Parasite mass | −70.6 ± 25 | −2.825 | 0.006 | |
| Neobenedenia sp. X Host blood δ15N | 0.8 ± 0.3 | 2.542 | 0.013 |
| Host | Host Tissue | Parasite Type | Parasite Species | Family | n | ∆13C | ∆15N | Authors |
|---|---|---|---|---|---|---|---|---|
| Acanthurus coeruleus | Blood | Monogenean | Neobenedenia sp. | Capsalidae | 44, 70 | 1.9 | 2.0 | Present study |
| Labeobarbus aeneus | Muscle | Monogenean | P. ichthyoxanthon | Diplozoidae | 7 | 0.09 | 2.05 | Sures et al., 2018 |
| Labeobarbus kimberleyensis | Muscle | Monogenean | P. ichthyoxanthon | Diplozoidae | 2 | 0.22 | 2.31 | Sures et al., 2018 |
| Acanthurus coeruleus | Blood | Copepod | Caligus atromaculatus | Caligidae | 22, 23 | 1.0 | 0.9 | Present study |
| Acanthurus bahianus | Blood | Copepod | Caligus atromaculatus | Caligidae | 15, 17 | −0.6 | 1.4 | Present study |
| Gadus morhua | Gill | Copepod | Clavella adunca | Lernaeopodidae | 1 | −2.19 | −1.29 | Deudero et al., 2002 |
| Merlangius merlangus | Gill | Copepod | Clavella adunca | Lernaeopodidae | 5 | −4.06 | −4.23 | Deudero et al., 2002 |
| Clarias gariepinus | Muscle | Copepod | Lamproglena clariae | Lernaeidae | 44 | −0.5 | 0.24 | Gilbert et al., 2020 |
| Platichthys flesus | Skin | Copepod | Lepeophtheirus pectoralis | Caligidae | 2 | 0.11 | −0.22 | Deudero et al., 2002 |
| Sprattus sprattus | Eye | Copepod | Lernaeenicus sprattae | Pennellidae | 1 | −1.39 | 0.60 | Deudero et al., 2002 |
| Gadus morhua | Gill | Copepod | Lernaeocera branchialis | Pennellidae | 2 | −2.39 | −0.82 | Deudero et al., 2002 |
| Melanogrammus aeglefinus | Gill | Copepod | Lernaeocera branchialis | Pennellidae | 3 | −0.62 | 1.10 | Deudero et al., 2002 |
| Merlangius merlangus | Gill | Copepod | Lernaeocera branchialis | Pennellidae | 6 | −1.55 | −2.61 | Deudero et al., 2002 |
| Platichthys flesus | Muscle | Copepod | Lernaeocera branchialis | Pennellidae | 3 | −1.63 | −0.81 | Pinnegar et al., 2001 |
| Merlangius merlangus | Gill | Copepod | unknown | unknown | 2 | −1.97 | −5.82 | Deudero et al., 2002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jenkins, W.G.; Demopoulos, A.W.J.; Nicholson, M.D.; Sikkel, P.C. Stable Isotope Dynamics of Herbivorous Reef Fishes and Their Ectoparasites. Diversity 2020, 12, 429. https://doi.org/10.3390/d12110429
Jenkins WG, Demopoulos AWJ, Nicholson MD, Sikkel PC. Stable Isotope Dynamics of Herbivorous Reef Fishes and Their Ectoparasites. Diversity. 2020; 12(11):429. https://doi.org/10.3390/d12110429
Chicago/Turabian StyleJenkins, William G., Amanda W. J. Demopoulos, Matthew D. Nicholson, and Paul C. Sikkel. 2020. "Stable Isotope Dynamics of Herbivorous Reef Fishes and Their Ectoparasites" Diversity 12, no. 11: 429. https://doi.org/10.3390/d12110429
APA StyleJenkins, W. G., Demopoulos, A. W. J., Nicholson, M. D., & Sikkel, P. C. (2020). Stable Isotope Dynamics of Herbivorous Reef Fishes and Their Ectoparasites. Diversity, 12(11), 429. https://doi.org/10.3390/d12110429