1. Introduction
Rare plant species have ecological, political, and intrinsic value, leading land managers and conservationists to advocate for their protection. Reserves are established in areas with a high concentration of rare and sensitive listed species [
1,
2] to help protect the biodiversity contributions of rare species [
3]. However, managing reserves with multiple rare species can prove challenging. Evaluating the types of rarity can help identify appropriate actions to protect populations. Rare species can be locally rare but have large geographic ranges, locally abundant with small ranges, or locally rare with large ranges, and can thus have wide or narrow habitat specificity [
4]. Rare species may also be rare because of their life history characteristics, such as their dependency on a disturbance regime or a specific pollinator [
5,
6,
7]. With diverse life histories, species ranges may be at the periphery of protected land, with only one population or maybe even one individual [
8]. These species are subject to inbreeding or extinction just from their small population sizes alone [
9,
10]. It’s important that conservation efforts cover the diversity of rare species characteristics when establishing reserves.
Protected areas are increasingly fragmented by urban or agricultural development, which is a barrier for rare species movement and leads to anthropogenic threats. These threats at the edge of a reserve include activities around unauthorized trail usage, a high density of invasive species, and a higher susceptibility to human-caused fires [
11]. Roads and trails can spread invasive species into rare species habitat and have negatively impacted the recruitment and persistence of rare plants [
12,
13]. Unfortunately, there are some reserves with high species richness near high-density human populations [
14]. These areas are also more vulnerable to increased fire return intervals [
15], which negatively impacts some rare species which are sensitive to increased fire regimes [
5,
6].
Considering the threats and challenges of managing rare plants, it is important that land managers curate and reflect on existing data to make management decisions [
16]. Scientists and botanical groups motivated to conserve rare species have curated lists of species that help to rank species based on the type of rarity and the threats to those species within specific geographic areas. Species have been ranked at many spatial scales: globally, through the International Union for Conservation of Nature and Natural Resources [
17]; regionally, like the California Native Plant Societies Inventory of Rare Plants [
18]; and locally, like a curated list developed by scientists for locally rare species in Napa County [
19]. These lists can then be used to develop databases with the occurrence of rare species to inform conservation planning [
20].
Incident data within databases can be used to further understand species characteristics or predict the potential presence of new populations [
21,
22,
23]. However, sampling bias in spatial data can negatively impact rare plant conservation by failing to accurately predict rare species distributions [
24]. Spatial sampling biases along roads negatively impact distribution models and reserve planning [
25,
26]. To account for sampling bias, sampling for rare species should be methodical, covering suitable and unsuitable habitat to create reliable presence-absence data [
27], and adequately capture annual variability [
28]. Instead, sampling for rare plants is opportunistic considering funding and disturbances like fire or a good rain year following many years of drought. These disturbances only capture part of the environmental variability and leave us questioning whether these rare species are present only after disturbance. While it would be nice to sample more frequently, it is not always feasible for reserve managers to systematically sample an entire reserve annually.
Here, we examine the occurrence and distribution of rare plant species within a natural reserve system in the western United States of America, specifically the Natural Communities Coalition (NCC) Reserve System in Orange County, California. The NCC Reserve System was established under California’s Natural Communities Conservation Plan/Habitat Conservation Plan (NCCP/HCP) to broadly protect sensitive species and their habitats adjacent to urban areas in a Mediterranean ecosystem. Within the NCC Reserve System, there are multiple stakeholders and land managers all collecting rare plant data on their individual management units. We seek to understand the distribution of rare plants within the NCC Reserve System, the survey efforts for rare species, their locations in relation to potential threats to their populations, and the vegetation communities these species occur in. Potential threats to rare plant species populations within our study area include disturbances associated with the urban–wildland interface (e.g., increased fire return interval, habitat edge, and habitat patchiness) and changes to habitat type from sea level rise. We expected that rare plant observations would focus on more charismatic species and be near roads and trails due to observer biases and access restrictions for surveyors. We also expected that rare species would be present within all vegetation communities. Still, some vegetation communities such as coastal sage scrub could have higher occurrence records of rare species due to the relative percent coverage of this community type within the protected area. Identifying other vegetation communities that are home to rare plant species will help clarify vegetation types that should be priorities for conservation and restoration efforts. This research aims to further the considerations for rare plant species prioritization and monitoring efforts.
2. Materials and Methods
2.1. Location Description
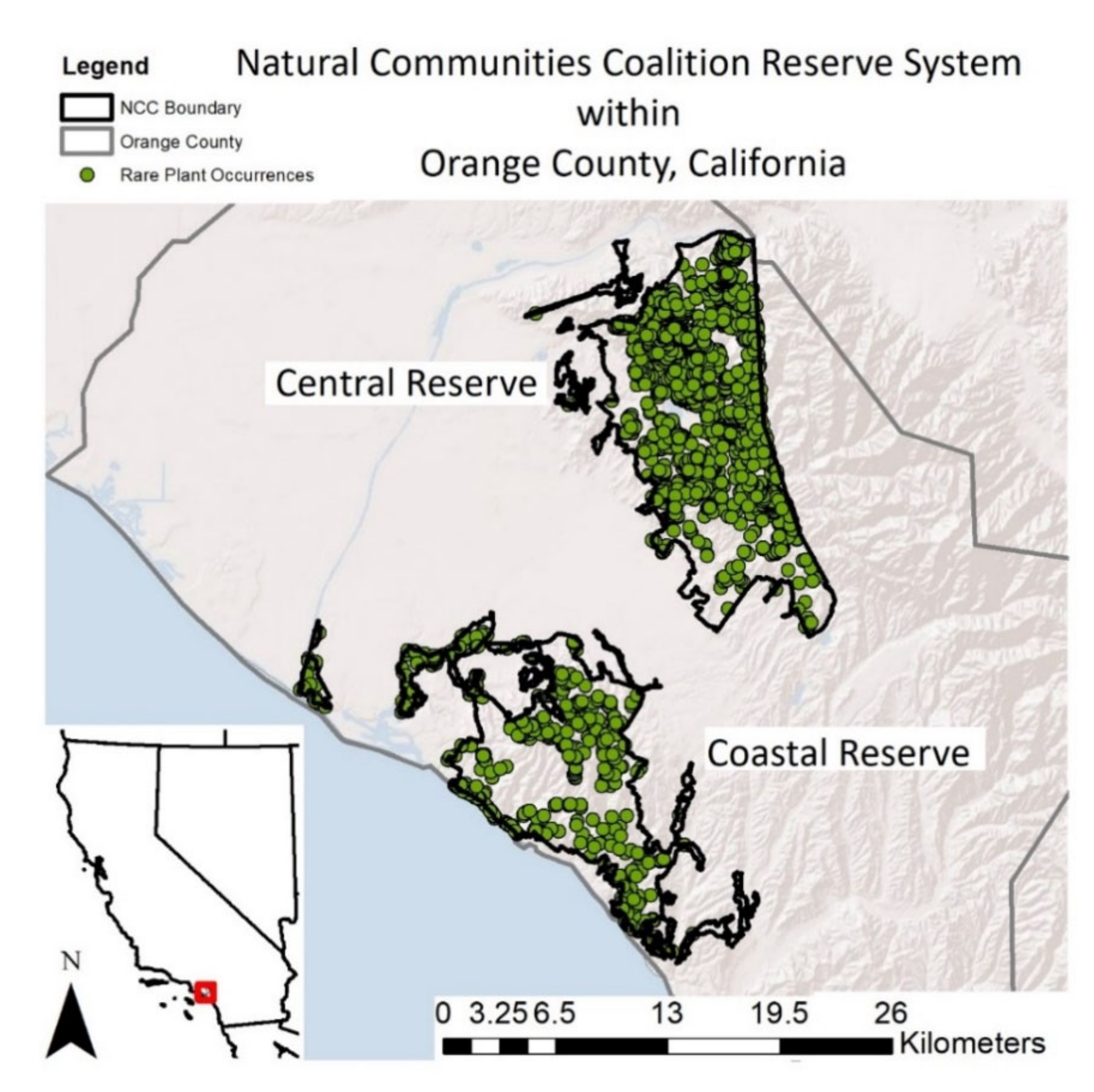
The rare plant species within this study occur in Orange County, California, USA and many are protected within the 38,000-acre (15,384 hectares). The NCC Reserve System is dominated by coastal sage scrub habitat and is composed of a Coastal Reserve and a Central Reserve, which are separated by urbanized areas. The land is owned and managed by many different organizations, including County, State, and non-profit entities (
Figure 1).
2.2. Species Selection
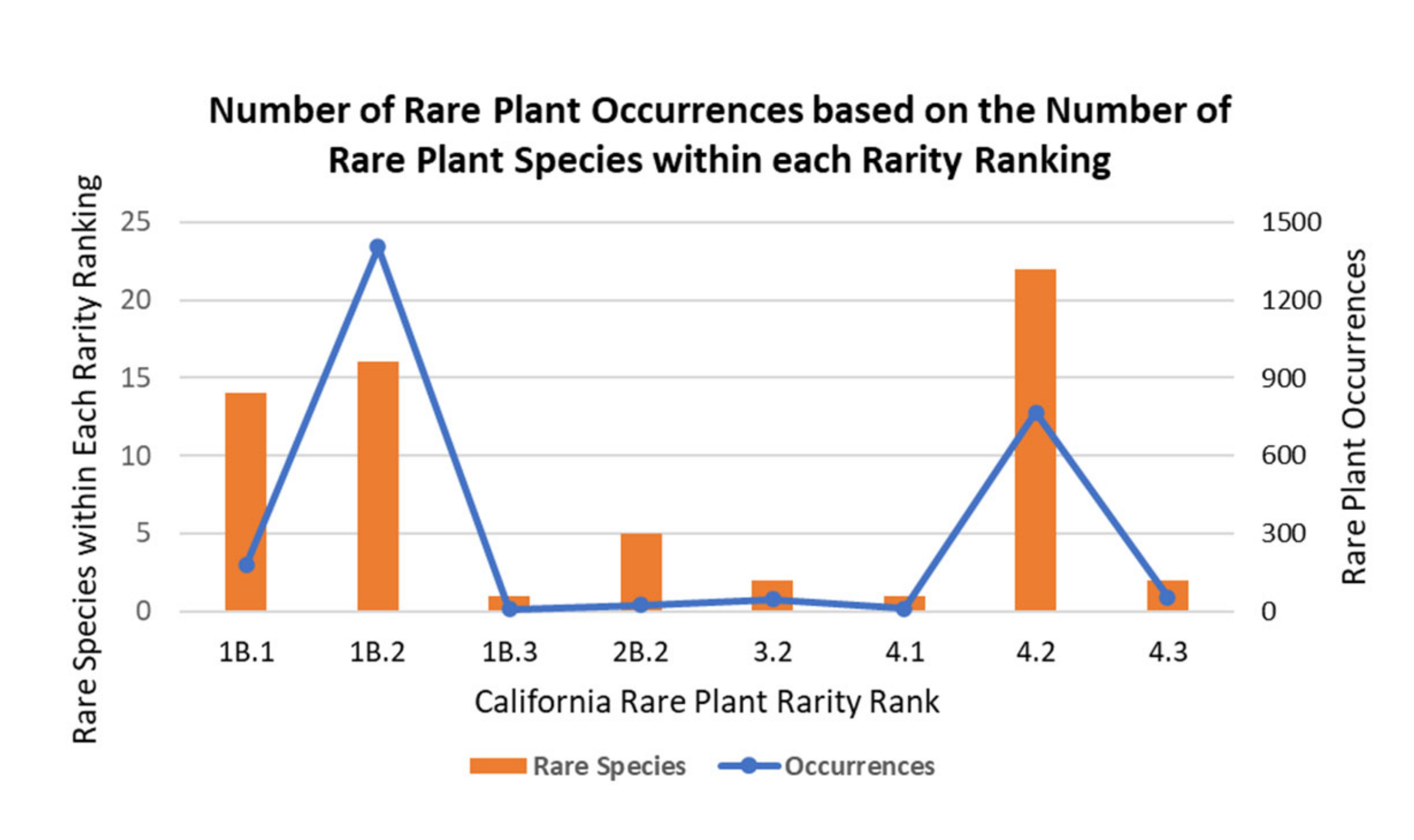
Ninety-two (92) rare species were selected based on the California Native Plant Society (CNPS) Inventory of Rare Plants database search for Orange County, California and species considered locally rare within Orange County, California. The CNPS Inventory of Rare Plants provides species with a California Rare Plant Rank (CRPR) from one to four, with A or B providing a designation for likely extirpated in California (A), or not extirpated within California (B; CNPS 2019). The last number, 1 through 3, is the threatened status, one indicating the highest threat level. For example, Brodiaea filifolia (thread-leaved brodiaea), CRPR 1B.1, is rare, threatened, and endangered in California, currently has an established extent, and is seriously threatened in California with more than 80% of occurrences at a high threat to extirpation. 1B species are already listed or pending state and federal listing under the California Endangered Species Act (CESA) or the Federal Endangered Species Act (ESA). Species with a CRPR of 2 are rare, threatened, or endangered in California but are more common elsewhere, like Pseudognaphalium leucocephyllum (white cudweed), which also has occurrences in New Mexico and Arizona. CRPR 3 species are species that CNPS is unsure of sorting into a group due to lack of information about their distribution, threats, ecology, and taxonomy. Lastly, CRPR 4 species are infrequent species with limited distribution, whose populations should be monitored regularly.
This study includes occurrence data from open source websites, password-protected databases, researchers, and consultants. Data for all 92 species were organized into a single geodatabase using ArcGIS 10.5 software by Esri [
29]. Data without geographic coordinate information was excluded, as well as all data from before 1900, because those observations were either extirpated or confirmed in a more recent survey. Data were projected using the NAD 1983 UTM Zone 11 coordinate system. The occurrence data was then clipped based on the NCC Reserve System boundary, and only 64 of the 92 species had occurrences within the NCC Reserve System. One of the 65 species is extirpated from the area and was not included in the analyses. Occurrence data were quality controlled by local botanical experts (see Acknowledgements).
2.3. Data Analysis
To answer our first question of spatial occurrence for rare plants within the NCC Reserve, we used ArcGIS to map rare plant species occurrence, rare plant species richness, and distance of rare plants from the coastal edge, the urban edge, and the trail edge.
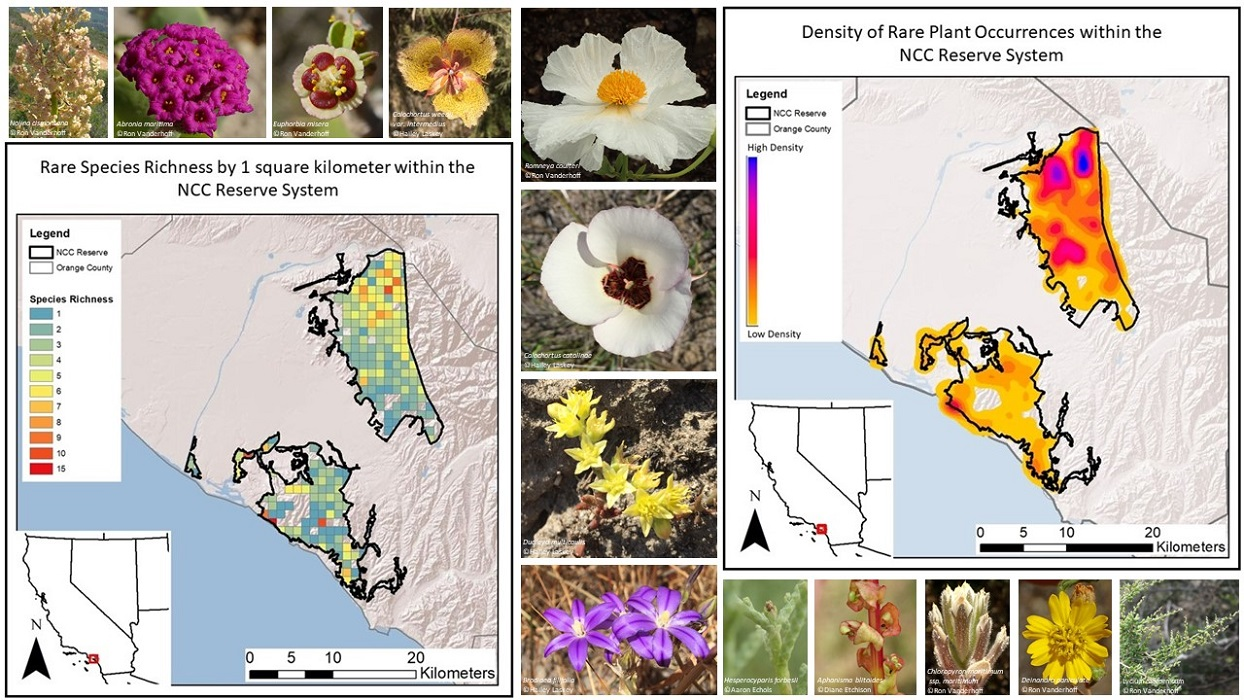
A kernel density tool was used to understand the distribution and density of occurrences throughout the NCC Reserve. Kernel density estimation (KDE) is a non-parametric statistical method for estimating the probability of points per unit area—we chose 100 square meters as the unit area. The KDE calculates a normal distribution for each data point first, and then all the y-values along this distribution are added up. Bumps are created from the added y-values, and the size of the bumps indicates the density of points in that area (larger bumps = higher density) along with the distribution. Probable density distribution is then visualized using a 250-bin color ramp.
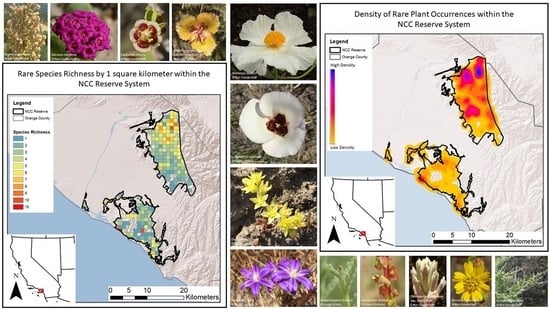
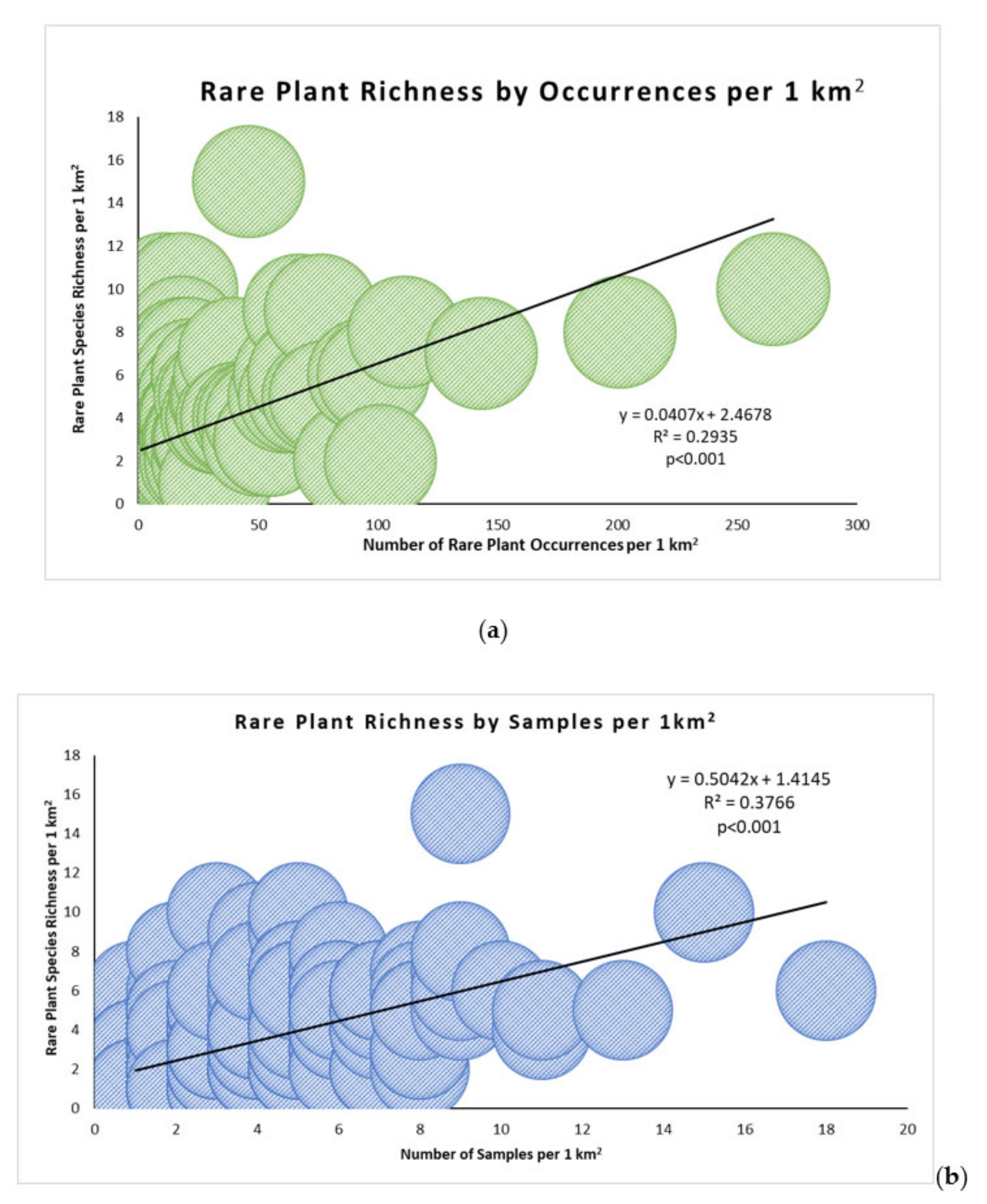
To understand the pattern of rare plant species richness across the NCC, we created a grid with an area of 1 square kilometer over the NCC Reserve and joined it to the rare plant occurrence layer by area to obtain a count of the total number of species for each grid polygon. Using the delete identical tool, we removed the species with multiple occurrences per grid cell to bring the value to the number of unique, rare plants per square kilometer. We completed the same steps as richness to obtain samples per square kilometer and occurrence density per square kilometer. To test if rare plant species richness was associated with either the number of samples or the number of occurrences, we used Spearman’s rank correlation. We then performed two linear regression analyses to understand if richness could be predicted by the number of samples or number of occurrences.
We used Chi-Square goodness-of-fit and Bonferroni testing to analyze the likelihood of rare plant occurrences within a certain distance from a trail [
30], urban edge [
31], and or coastal mean high tide line [
32]. To answer whether a species was more likely to be found within a specified distance to an edge than not, we created a 50 square meter grid over the NCC Reserve System polygon. We used the total number of squares as the expected value and the number of squares with a rare plant occurrence as the observed value. The distance in meters from an edge (trail, coastline, urban edge) was merged to each 50 square meter polygon. The distance data for each type of edge was then sorted into 11 distance categories. To see the significance between each group and to adjust for Type I error, we calculated the adjusted residuals and compared those values to a new critical level, the z criteria. The z criteria are the inverse of the standard normal cumulative distribution function. If the adjusted residual is greater than or less than the positive and negative z criteria, respectively, we can say that the observed or expected was more or less likely than normal. The Bonferroni test was then verified using a chi-square test to compare one distance category to the compilation of all categories.
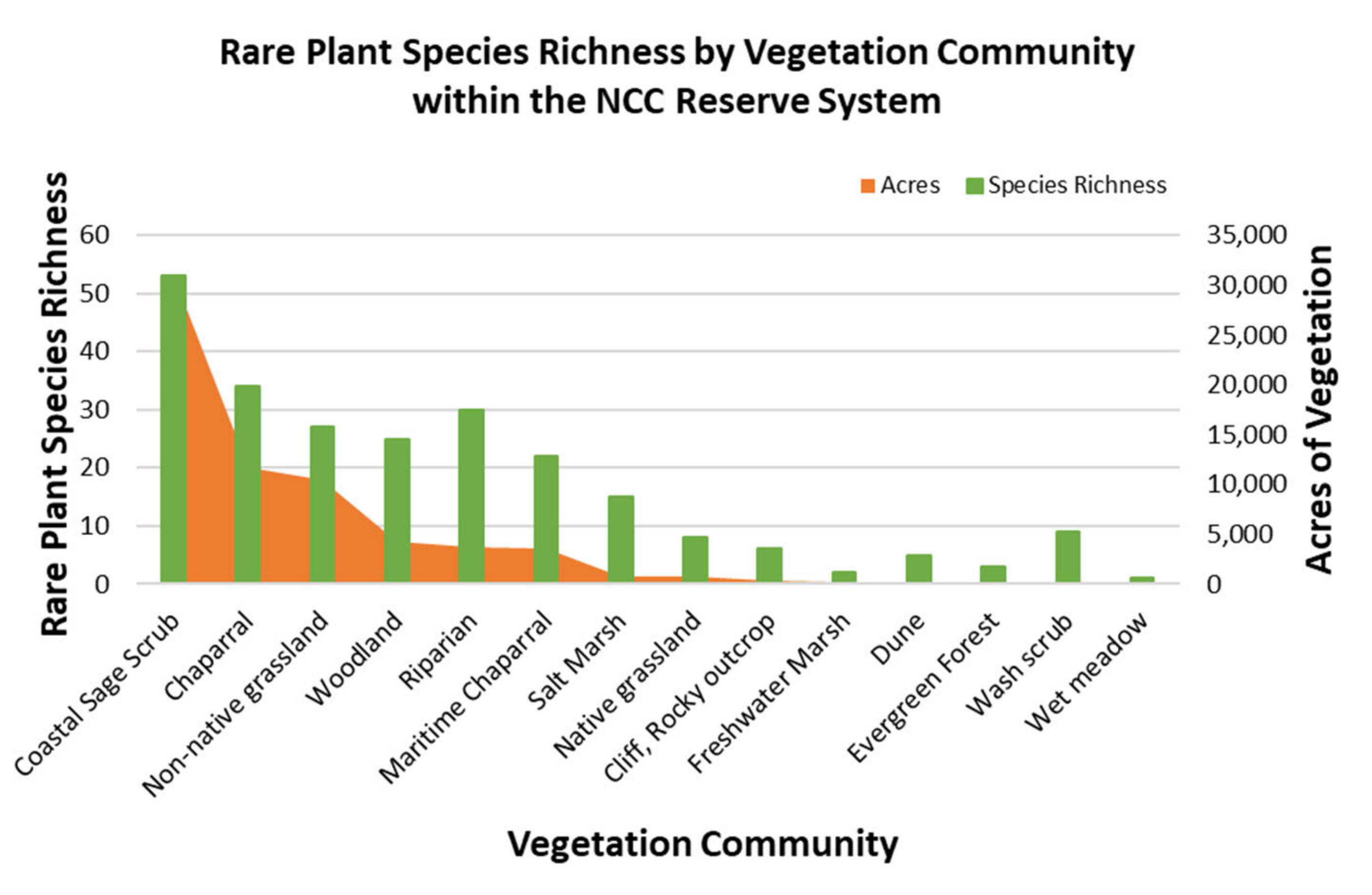
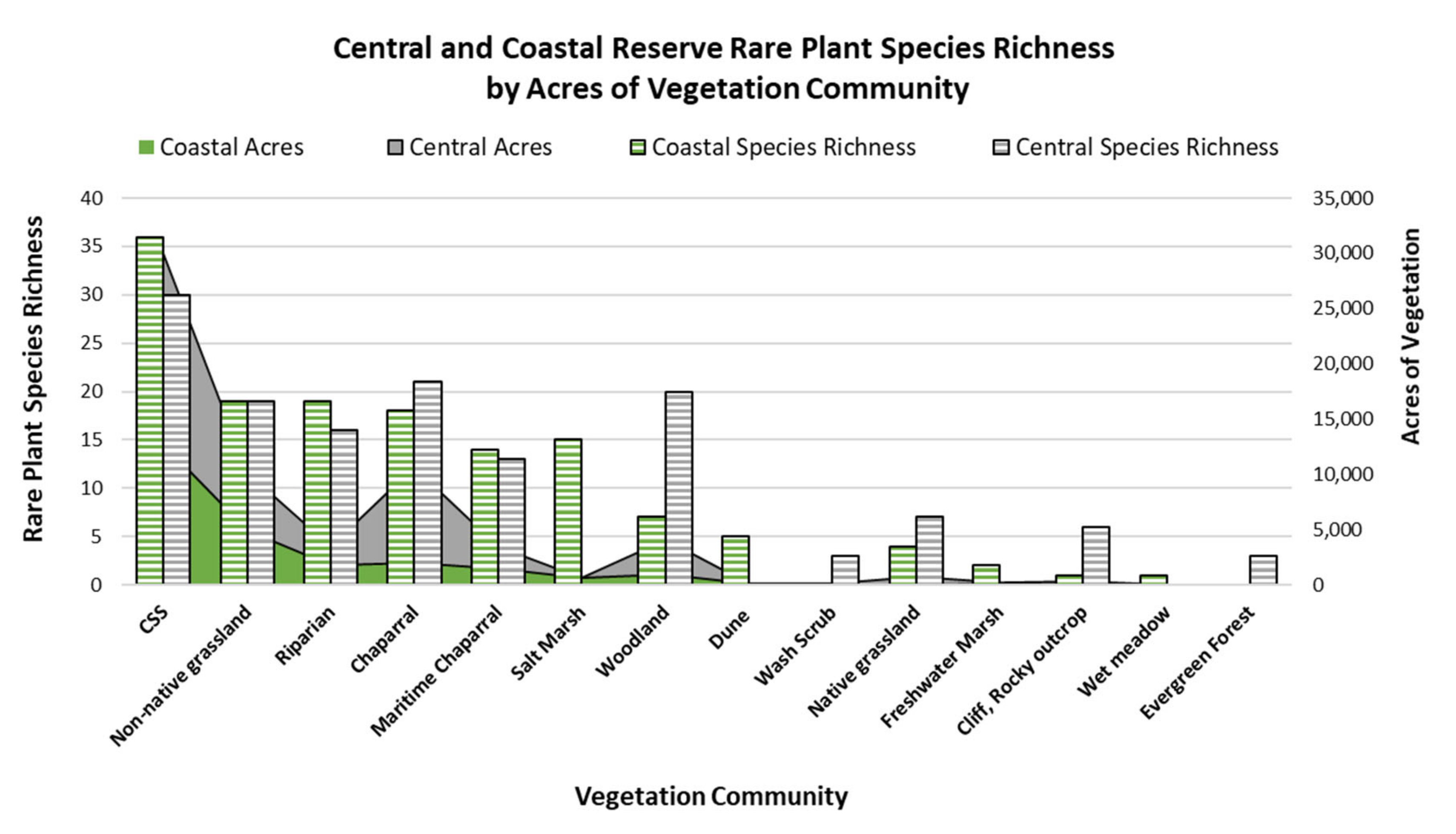
We analyzed rare plant species richness by vegetation community for the NCC Reserve System by first intersecting the occurrence data with the vegetation data for the NCC Reserve [
31]. To show the density of occurrences, we generated a table that included the count of species occurrences by vegetation community, which was used to obtain species richness. To analyze the association between the acres of each vegetation community and species richness within each vegetation community, we summed the count of species by each vegetation community and performed a Spearman’s rank correlation analysis. We ran this test for the entire reserve, and separately for both the coastal the central reserve. We also used Chi-Square goodness-of-fit and Bonferroni testing to calculate whether the probability that rare species were found in any vegetation type was greater than if the species were randomly distributed across the reserve. We used the polygon count of each vegetation community within the vegetation map to generate the expected values.
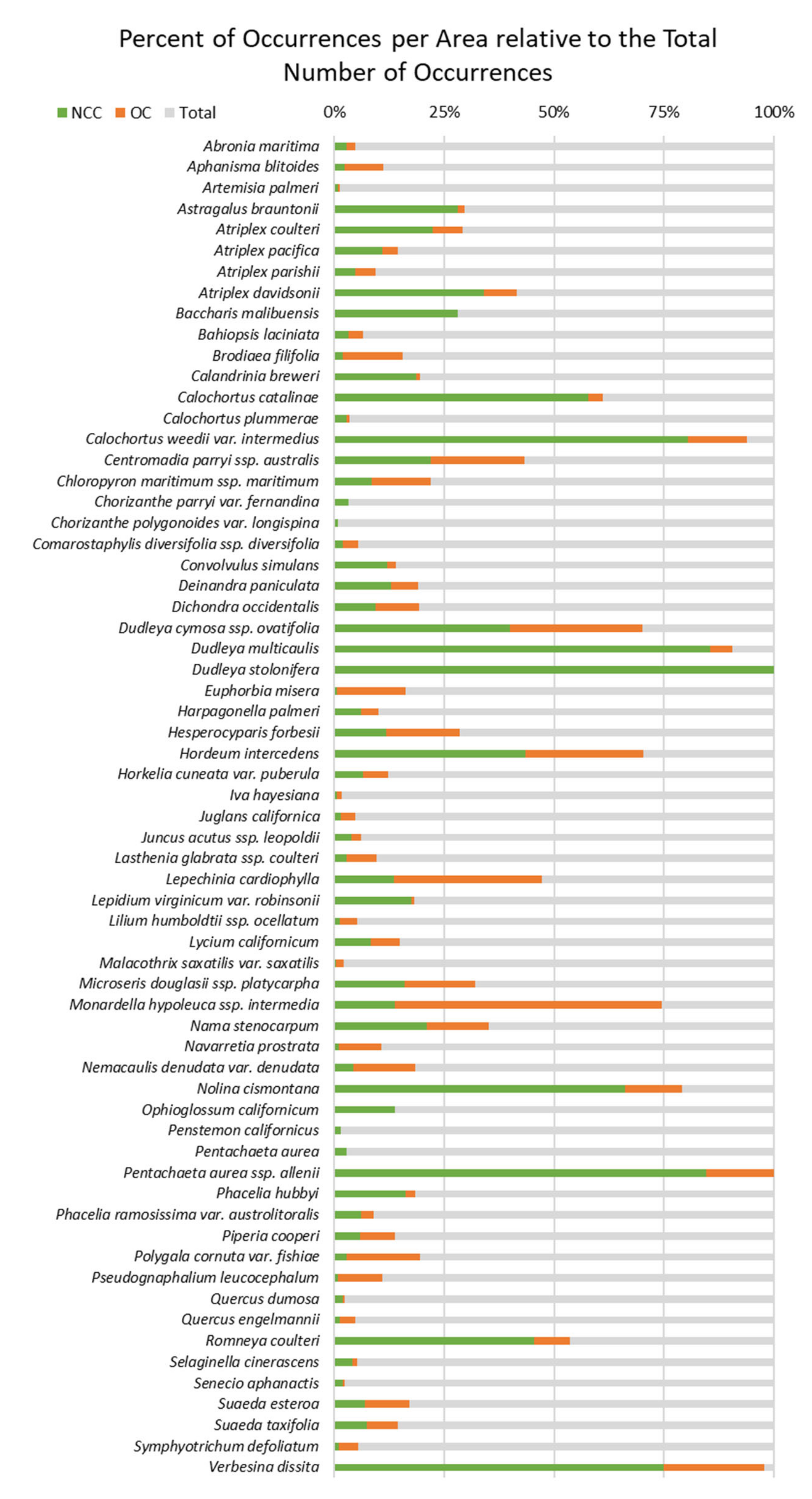
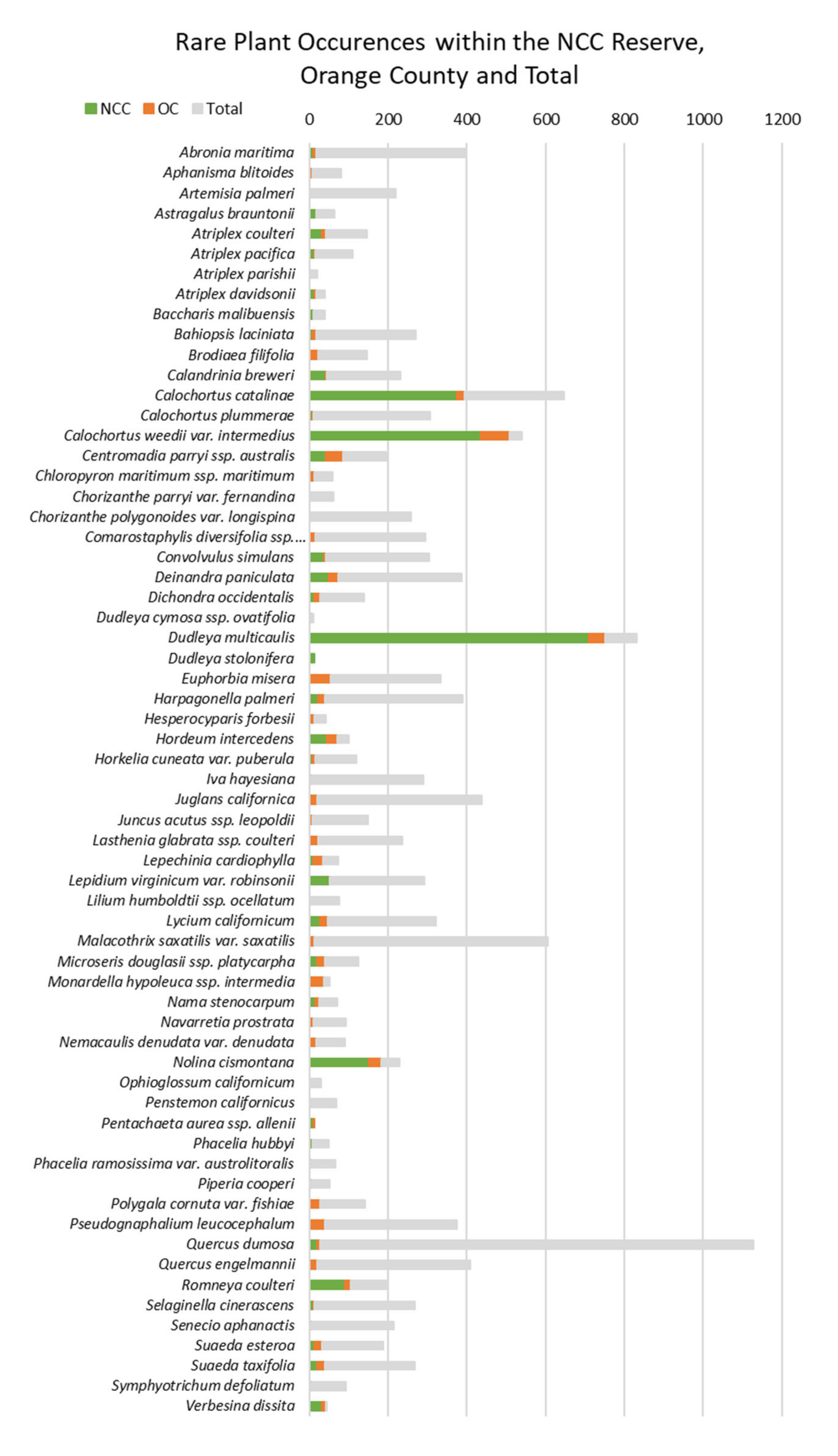
To analyze the geographic distribution of the 92 species, we compiled a table with the number of occurrences and the descriptive distribution of the species ranges (
Appendix A). We sorted the occurrence data by the total number of occurrences throughout the species’ entire range, within Orange County, and within the NCC Reserve. The kernel density maps were used to describe the species range and density of occurrences by area. Rare species ranges were described by the extent of a geographic region like the San Francisco Bay Area, Point Conception, and Baja California. Species ranges were also described with regards to the position of Orange County within the broader range.
5. Conclusions
Comprehensive analysis of the rare plant occurrences within our study region revealed geographic locations (e.g., coastal bluffs and farther away from trails) and vegetation types (e.g., invaded grasslands, coastal sage scrub, and salt marsh) with high numbers of rare plants, as well as, species that are endemic (e.g., Dudleya stolonifera and Pentachaeta aurea ssp. allenii) or whose ranges fall mostly within the Reserve (e.g., Dudleya multicaulis and Calochortus weedii var. intermedius). The data also revealed undocumented bias by species and uneven sampling throughout our study region. Although the species within our project area are protected, they are still vulnerable to threats, such as sea-level rise, increased disturbance regime, and habitat fragmentation. Land managers can use rare plant occurrence data to assess species distributions across multiple scales to prioritize species monitoring based on priority threats. However, occurrence data can include sampling bias and focused survey efforts may be necessary to address data gaps. We recommend that rare plant occurrence data be continually updated, and that long-term monitoring plots be established for the rarest species.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







