Transferability of Microsatellite Markers Developed in Oenothera spp. to the Invasive Species Oenothera drummondii Hook. (Onagraceae)


 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
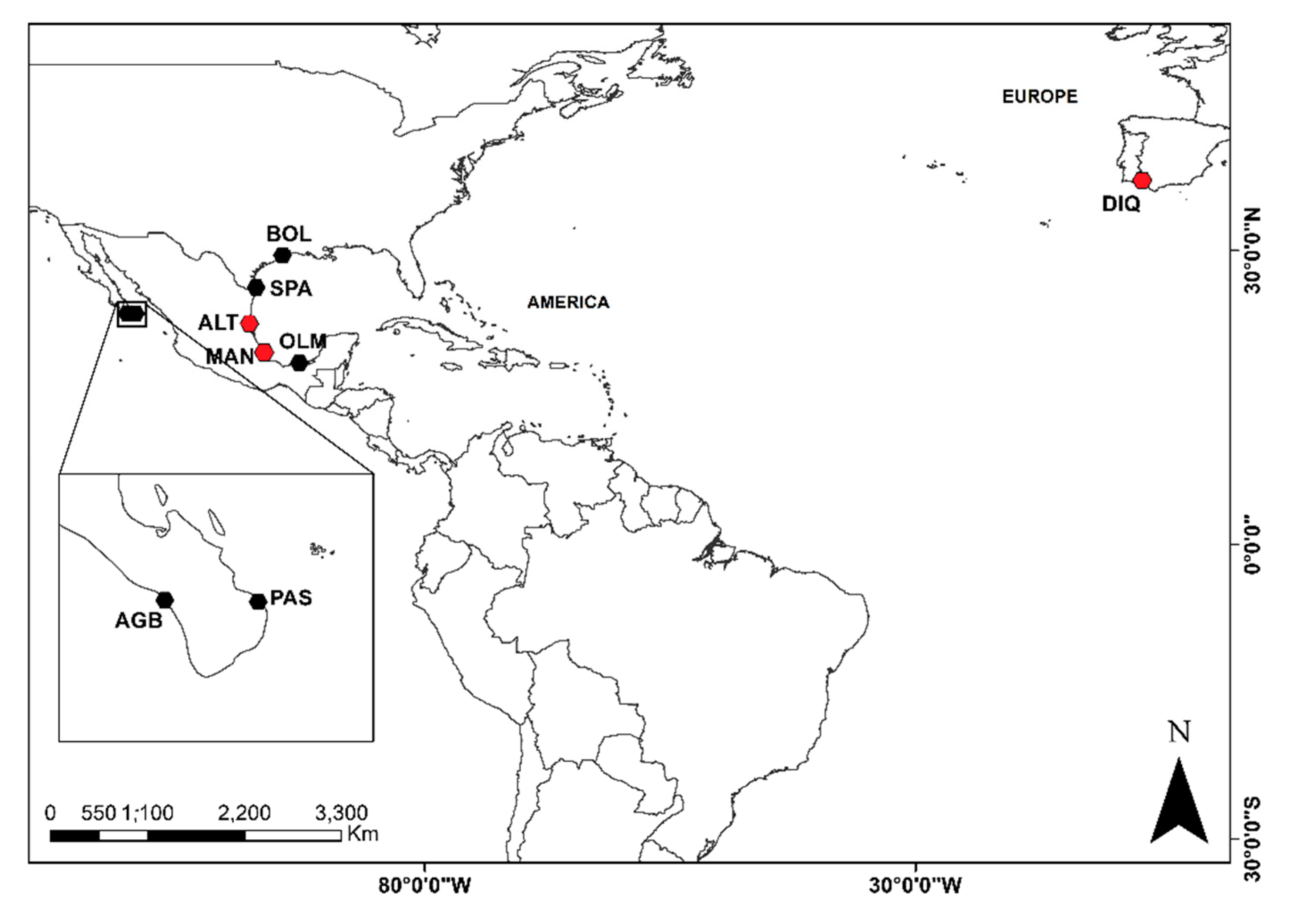
2.1. Sampling
2.2. DNA Extraction and Vegetative Material
2.3. Primer Selection
2.4. Microsatellite Amplification
2.5. Data Analysis
3. Results
3.1. Microsatellite Transferability
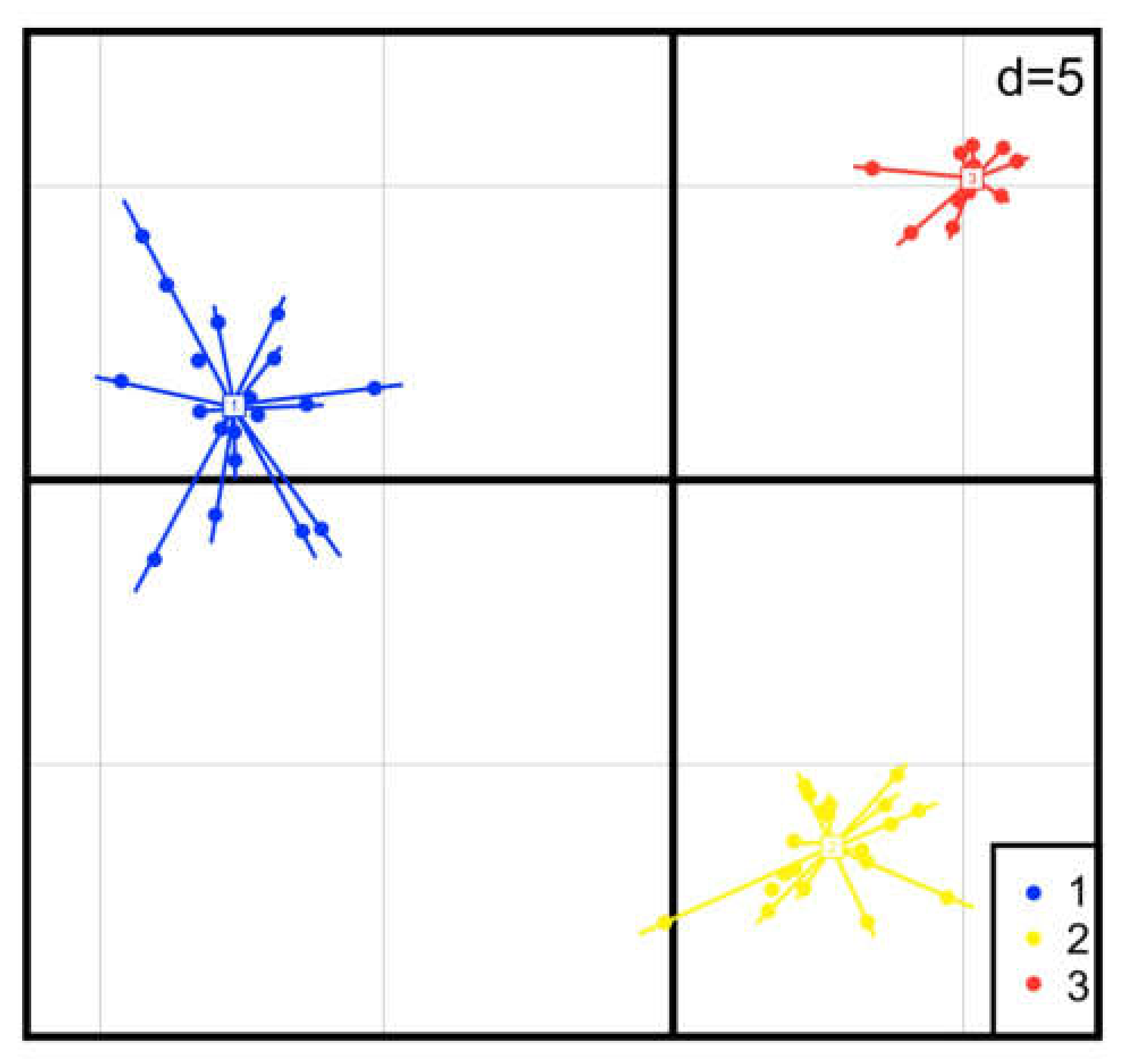
3.2. Characterization of Polymorphic Microsatellites in Three Populations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dietrich, W. Vol. VIII Haloragaceae-Euphorbiaceae. In Flora Ibérica; Castroviejo, S., Aedo, C., Benedi, C., Lainz, M., Garmendia, F., Feliner, G., Paiva, J., Eds.; Real Jardín Botánico: Madrid, Spain, 1997; p. 375. ISBN 84-00-06221-3. [Google Scholar]
- Frean, M.; Balkwill, K.; Gold, C.; Burt, S. The expanding distributions and invasiveness of Oenothera in southern Africa. South Afr. J. Bot. 1997, 63, 449–458. [Google Scholar] [CrossRef]
- Lonard, R.I.; Judd, F.W. Phenology of Native Angiosperms of South Padre Island, Texas. In Proceedings of the Eleventh North American Prairie Conference, Lincoln, NE, USA, 7–11 August 1988; University of Nebraska—Lincoln: Lincoln, NE, USA, 1989; Volume 32, pp. 217–222. [Google Scholar]
- Wagner, W.L.; Hoch, P.C.; Raven, P.H. Revised classification of the Onagraceae. Syst. Bot. Monogr. 2007, 37, 1–240. [Google Scholar] [CrossRef]
- Dietrich, W.; Wagner, W.L. Systematics of Oenothera Section Oenothera Subsection Raimannia and Subsection Nutantigemma (Onagraceae); American Society of Plant Taxonomists: Ann Arbor, MI, USA, 1988; Volume 24, pp. 1–91. [Google Scholar] [CrossRef]
- Dietrich, W.; Wagner, W.L. New Taxa of Oenothera L. Sect. Oenothera (Onagraceae). Ann. Mo. Bot. Gard. 1987, 74, 144–150. [Google Scholar] [CrossRef]
- García-de Lomas, J.; Fernández-Carrillo, L.; Saavedra, M.; Mangas, L.; Rodríguez, C.; Sánchez-Gullón, E.; Martínez, E. Invasión of Oenothera drummondii Hook. (Onagraceae) en el paraje natural Marismas del Odiel (Huelva, Sur de España): Bases para la gestión de una invasión avanzada. Rev. Soc. Gad. Hist. Nat. 2015, 9, 41–50. [Google Scholar]
- Campos, J.A.; Herrera, M.; Biurrun, I.; Loidi, J. The role of alien plants in the natural coastal vegetation in central-northern Spain. Biodivers. Conserv. 2004, 13, 2275–2293. [Google Scholar] [CrossRef]
- Xu, H.; Qiang, S.; Genovesi, P.; Ding, H.; Wu, J.; Meng, L.; Han, Z.; Miao, J.; Hu, B.; Guo, J.; et al. An inventory of invasive alien species in China. NeoBiota 2012, 15, 1–26. [Google Scholar] [CrossRef]
- Dufour-Dror, J.M. Israel’s Least Wanted Alien Ornamental Plant Species Ornamental Plants Potentially Invasive in Israel’s Natural Ecosystems; The Middle East Nature Conservation Promotion Association: Jerusalem, Israel, 2013. [Google Scholar]
- Heenan, P.B.; de Lange, P.J.; Cameron, E.K.; Champion, P.D. Checklist of dicotyledons, gymnosperms, and pteridophytes naturalized or casual in New Zealand: Additional records 1999–2000. N. Z. J. Bot. 2002, 40, 155–174. [Google Scholar] [CrossRef]
- Heyligers, P.C. Flora of the Stockton and Port Hunter sandy foreshores with comments on fifteen notable introduced species. Cuninghamia 2008, 10, 493–511. [Google Scholar]
- Gallego-Fernández, J.B.; Martínez, M.L.; García-Franco, J.G.; Zunzunegui, M. Multiple seed dispersal modes of an invasive plant species on coastal dunes. Biological Invasions 2020, 6. [Google Scholar] [CrossRef]
- Zunzunegui, M.; Ruiz-valdepeñas, E.; Sert, M.A.; Díaz-barradas, M.C.; Gallego-fernández, J.B. Field comparison of ecophysiological traits between an invader and a native species in a Mediterranean coastal dune. Plant Physiol. Biochem. 2020, 146, 278–286. [Google Scholar] [CrossRef]
- Gallego-Fernández, J.B.; Martínez, M.L.; García-Franco, J.G.; Zunzunegui, M. The impact on plant communities of an invasive alien herb, Oenothera drummondii, varies along the beach-coastal dune gradient. Flora 2019, 260, 151466. [Google Scholar] [CrossRef]
- García-De-Lomas, J.; Fernández-Carrillo, L.; Saavedra, C.; Dana, E.D.; Rodríguez, C.; Martínez, E. Feasibility of using glyphosate to control beach evening primrose Oenothera drummondii in heavily invaded coastal dunes, Odiel Marshes, Spain. Conserv. Evid. 2016, 13, 72–78. [Google Scholar]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.U.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evolut. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, H.; Richardson, D.M.; Le Roux, J.J. Introduction to the special issue: Tree invasions: Towards a better understanding of their complex evolutionary dynamics. AoB PLANTS 2017, 9, 1–8. [Google Scholar] [CrossRef][Green Version]
- Richards, A.J. Plant Breeding Systems; Chapman & Hall Ltd.: London, UK, 1997. [Google Scholar]
- Mazzolari, A.C.; Marrero, H.J.; Vázquez, D.P. Potential contribution to the invasion process of different reproductive strategies of two invasive roses. Biol. Invas. 2017, 19, 615–623. [Google Scholar] [CrossRef]
- Nei, M.; Maruyama, T.; Chakraborty, R. The Bottleneck Effect and Genetic Variability in Populations. Evolution 1975, 29, 1–10. [Google Scholar] [CrossRef]
- Liu, L.; Pei, C.; Liu, S.; Guo, X.; Du, N.; Guo, W. Genetic and epigenetic changes during the invasion of a cosmopolitan species (Phragmites australis). Ecol. Evolut. 2018, 8, 6615–6624. [Google Scholar] [CrossRef]
- Dlugosch, K.M.; Anderson, S.R.; Braasch, J.; Cang, F.A.; Gillette, H.D. The devil is in the details: Genetic variation in introduced populations and its contributions to invasion. Mol. Ecol. 2015, 24, 2095–2111. [Google Scholar] [CrossRef]
- Dlugosch, K.M.; Hays, C.G. Genotypes on the move: Some things old and some things new shape the genetics of colonization during species invasions. Mol. Ecol. 2008, 17, 44583–44585. [Google Scholar] [CrossRef]
- Frankham, R. Resolving the genetic paradox in invasive species. Heredity 2005, 94, 385. [Google Scholar] [CrossRef]
- Refoufi, A.; Esnault, M.A. Genetic diversity and population structure of Elytrigia pycnantha (Godr.) (Triticeae) in Mont Saint-Michel Bay using microsatellite markers. Plant Biol. 2006, 8, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.; Robson, B.; Pearson, M.L. Population genetic structure of a colonizing, triploid weed, Hieracium lepidulum. Heredity 2004. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, S.; Mauricio, R. Neutral genetic variation among wild North American populations of the weedy plant Arabidopsis thaliana is not geographically structured. Mol. Ecol. 2004, 13, 3403–3413. [Google Scholar] [CrossRef] [PubMed]
- Durka, W.; Bossdorf, O.; Prati, D.; Auge, H. Molecular evidence for multiple introductions of garlic mustard (Alliaria petiolata, Brassicaceae) to North America. Mol. Ecol. 2005, 14, 1697–1706. [Google Scholar] [CrossRef]
- Meekins, J.F.; Ballard, H.E., Jr.; McCarthy, B.C. Genetic Variation and Molecular Biogeography of a North American Invasive Plant Species (Alliaria petiolata, Brassicaceae). Int. J. Plant Sci. 2001, 162, 161–169. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The Population Biology of Invasive Species. Ann. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef]
- Husband, B.C.; Barrett, S.C.H. Colonization history and population genetic structure of Eichornia paniculata in Jamaica. Heredity 1991, 66, 287–296. [Google Scholar] [CrossRef][Green Version]
- Marrs, R.A.; Sforza, R.; Hufbauer, R.A. When invasion increases population genetic structure: A study with Centaurea diffusa. Biol. Invas. 2008, 10, 561–572. [Google Scholar] [CrossRef]
- Novak, S.J.; Welfley, A.Y. Genetic Diversity in the introduced Clonal Grass Poa bulbosa (Bulbous Bluegrass). NorthWest Sci. 1997, 71, 271–280. [Google Scholar]
- Vieira, M.L.C.; Santini, L.; Diniz, A.L.; de Freitas Munhoz, C. Microsatellite markers: What they mean and why they are so useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef]
- Daw, E.W.; Heath, S.C.; Lu, Y. Single-nucleotide polymorphism versus microsatellite markers in a combined linkage and segregation analysis of a quantitative trait. BMC Genetics 2005, 6, 4–6. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tsykun, T.; Rellstab, C.; Dutech, C.; Sipos, G.; Prospero, S. Comparative assessment of SSR and SNP markers for inferring the population genetic structure of the common fungus Armillaria cepistipes. Heredity 2017, 119, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Zalapa, J.E.; Cuevas, H.; Zhu, H.; Steffan, S.; Senalik, D.; Zeldin, E.; McCown, B.; Harbut, R.; Simon, P. Using next-generation sequencing approaches to isolate simple sequence repeat (SSR) loci in the plant sciences. Am. J. Bot. 2012, 99, 193–208. [Google Scholar] [CrossRef]
- Hoshino, A.A.; Bravo, J.P.; Nobile, P.M.; Morelli, K.A. Microsatellites as tools for genetic diversity analysis. In Genetic Diversity in Microorganisms; Caliskan, M., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Gou, X.; Shi, H.; Yu, S.; Wang, Z.; Li, C.; Liu, S.; Ma, J.; Chen, G.; Liu, T.; Liu, Y. SSRMMD: A Rapid and Accurate Algorithm for Mining SSR Feature Loci and Candidate Polymorphic SSRs Based on Assembled Sequences. Front. Genet. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Guichoux, E.; Lagache, L.; Wagner, S.; Chaumeil, P.; Léger, P.; Lepais, O.; Lepoittevin, C.; Malausa, T.; Revardel, E.; Salin, F.; et al. Current trends in microsatellite genotyping. Mol. Ecol. Resour. 2011, 11, 591–611. [Google Scholar] [CrossRef]
- Soares, T.N.; Sant’Ana, L.L.; de Oliveira, L.K.; Telles, M.P.C.; Collevatti, R.G. Transferability and characterization of microsatellite loci in Anacardium humile A. St. Hil. (Anacardiaceae). Genet. Mol. Res. 2013, 12, 3146–3149. [Google Scholar] [CrossRef]
- Lavor, P.; van den Berg, C.; Versieux, L.M. Transferability of 10 nuclear microsatellite primers to Vriesea minarum (Bromeliaceae), a narrowly endemic and threatened species from Brazil. Braz. J. Bot. 2013, 36, 165–168. [Google Scholar] [CrossRef]
- Gallego-Fernández, J.B.; Martínez, M.L. Environmental filtering and plant functional types on Mexican foredunes along the Gulf of Mexico. Ecoscience 2011, 18, 52–62. [Google Scholar] [CrossRef]
- González, D.; Vovides, A. Low Intralineage Divergence in Ceratozamia (Zamiaceae) Detected with Nuclear Ribosomal DNA ITS and Chloroplast DNA trnL-F Non-coding Region. Syst. Bot. 2002, 27, 654–661. [Google Scholar]
- Yue, Q.; Da-li, F.; Meng-pei, L.; Yu-qin, L.; Shuo, W. Optimization of microsatellite DNA Gelred fluorescence imaging technology. Afr. J. Biotech. 2012, 11, 14749–14754. [Google Scholar] [CrossRef]
- Lewis, E.M.; Fant, J.B.; Moore, M.J.; Hastings, A.P.; Larson, E.L.; Agrawal, A.A.; Skogen, K.A. Microsatellites for Oenothera gayleana and O. hartwegii subsp. filifolia (Onagraceae), and Their Utility in Section Calylophus. Appl. Plant Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Skogen, K.A.; Hilpman, E.T.; Todd, S.L.; Fant, J.B. Microsatellite primers in Oenothera harringtonii (Onagraceae), an annual endemic to the shortgrass prairie of Colorado. Am. J. Bot. 2012, 99, e313–e316. [Google Scholar] [CrossRef] [PubMed]
- Larson, E.L.; Bogdanowicz, S.M.; Agrawal, A.A.; Johnson, M.T.J.; Harrison, R.G. Isolation and characterization of polymorphic microsatellite loci in common evening primrose (Oenothera biennis). Mol. Ecol. Resour. 2008, 8, 434–436. [Google Scholar] [CrossRef] [PubMed]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments. Nat. Biotech. 2000, 18, 233–234. [Google Scholar] [CrossRef]
- Missiaggia, A.; Grattapaglia, D. Plant microsatellite genotyping with 4-color fluorescent detection using multiple-tailed primers. Genet. Mol. Res. 2006, 5, 72–78. [Google Scholar]
- Oblessuc, P.R.; de Campos, T.; Cardoso, J.M.K.; Sforça, D.A.; Baroni, R.M.; de Souza, A.P.; Benchimol, L.L. Adaptation of fluorescent technique for genotyping with new microsatellite markers in common bean. Pesqui. Agropecu. Bras. 2009, 44, 638–644. [Google Scholar] [CrossRef]
- Culley, T.M.; Stamper, T.I.; Stokes, R.L.; Brzyski, J.R.; Hardiman, N.A.; Klooster, M.R.; Merritt, B.J. An Efficient Technique for Primer Development and Application that Integrates Fluorescent Labeling and Multiplex PCR. Appl. Plant Sci. 2013, 1, 1300027. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Hartl, D.; Clark, A. Principles of Population Genetics, 3rd ed.; Sinauer, A., Ed.; Sinauer Associates: Sunderland, MA, Canada, 1997; ISBN 0-87893-306-9. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Slatkin, M. Inbreeding coefficients and coalescence times. Genet. Res. 1991, 58, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. The Interpretation of Population Structure by F-Statistics with Special Regard to Systems of Mating. Evolution 1965, 19, 395–420. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. An Introduction to dplR. 2013. Available online: http://cran.univ-paris1.fr/web/packages/dplR/vignettes/intro-dplR.pdf (accessed on 31 May 2013).
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genetics 2010, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, M. Sourcing of SSR markers from related plant species. In Plant Genotyping: The DNA Fingerprinting of Plants; Henry, R.J., Ed.; Centre of Agriculture and Bioscience International: Wallingford, UK, 2009; pp. 211–224. [Google Scholar]
- Barbará, T.; Palma-Silva, C.; Paggi, G.M.; Bered, F.; Fay, M.F.; Lexer, C. Cross-species transfer of nuclear microsatellite markers: Potential and limitations. Mol. Ecol. 2007, 16, 3759–3767. [Google Scholar] [CrossRef]
- Squirrell, J.; Hollingsworth, P.M.; Woodhead, M.; Russell, J.; Lowe, A.J.; Gibby, M.; Powell, W. How much effort is required to isolate nuclear microsatellites from plants? Mol. Ecol. 2003, 12, 1339–1348. [Google Scholar] [CrossRef]
- Zane, L.; Bargelloni, L.; Patarnello, T. Strategies for microsatellite isolation: A review. Mol. Ecol. 2002, 11, 1–16. [Google Scholar] [CrossRef]
- Singleton, J.J.; Mangat, P.K.; Shim, J.; Vavra, C.; Coldren, C.; Angeles-Shim, R.B. Cross-species transferability of Solanum spp. DNA markers and their application in assessing genetic variation in silverleaf nightshade (Solanum elaeagnifolium ) populations from Texas, USA. Weed Sci. 2020, 68, 396–404. [Google Scholar] [CrossRef]
- Zhu, X.; Raman, H.; Wu, H.; Lemerle, D.; Burrows, G.; Stanton, R. Development of SSR Markers for Genetic Analysis of Silverleaf Nightshade (Solanum elaeagnifolium) and Related Species. Plant Mol. Biol. Rep. 2012, 31. [Google Scholar] [CrossRef]
- Lobato-deMagalhães, T.; Cabrera-Toledo, D.; Martínez, M. Microsatellite loci transferability and genetic diversity of the aquatic plant Nimphoides fallax Ornduff (Menyanthaceae), endemic to the Mexican and Guatemalan highlands. Limnology 2019, 20, 223–241. [Google Scholar]
- Senan, S.; Kizhakayil, D.; Sasikumar, B.; Sheeja, T.E. Methods for Development of Microsatellite Markers: An Overview. Notulae Sci. Biol. 2014, 6, 1–13. [Google Scholar] [CrossRef]
- Levin, D.A. Genic heterozygosity and protein polymorphism among local populations of Oenothera biennis. Genetics 1974, 79, 477–491. [Google Scholar]
- Mather, K. Polygenic inheritance and natural selection. Biol. Rev. 1943, 18, 32–64. [Google Scholar] [CrossRef]
- Stebbins, G. Self-fertilization and population variability in plants. Am. Nat. 1957, 41, 337–352. [Google Scholar] [CrossRef]
- Martínez, M.L.; Psuty, N.; Lubke, R. A perspective on coastal dunes. In Coastal Dunes. Ecology and Conservation; Martínez, M.L., Psuty, N., Eds.; Springer-Verlag: Berlin, Germany, 2004; pp. 3–10. [Google Scholar]
- Ingvarsson, P.K. A metapopulation perspective on genetic diversity and differentiation in partially self-fertilizing plants. Evolution 2002, 56, 2368–2373. [Google Scholar] [CrossRef]
- Nordborg, M. Linkage Disequilibrium, Gene Trees and Selfing: An Ancestral Recombination Graph With Partial Self-Fertilization. Genetics 2000, 154, 923–929. [Google Scholar]
- Ward, S.M.; Jasieniuk, M. Review: Sampling Weedy and Invasive Plant Populations for Genetic Diversity Analysis. Weed Sci. 2009, 57, 593–602. [Google Scholar] [CrossRef]
- Lowe, A.; Harris, S.; Ashton, P. Ecological genetics: Design, Analysis and Application; Blackwell: Oxford, UK, 2004; ISBN 1-4051-0033-8. [Google Scholar]
- Taheri, S.; Abdullah, T.L.; Yusop, M.R.; Hanafi, M.M.; Sahebi, M.; Azizi, P.; Shamshiri, R.R. Mining and development of novel SSR markers using Next Generation Sequencing (NGS) data in plants. Molecules 2018, 23, 399. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Country | Locality | Code | Coordinates | |
|---|---|---|---|---|
| Latitude | Longitude | |||
| Spain | Andalucía; Huelva, El Dique | DIQ | 37.09 | −6.54 |
| USA | Texas; Bolívar | BOL | 29.30 | −94.30 |
| USA | Texas; South Padre Island | SPA | 26.13 | −97.11 |
| Mexico | Baja California Sur; Punta Arena del Sur | PAS | 23.55 | −109.47 |
| Mexico | Baja California Sur; Agua Blanca | AGB | 23.66 | −110.41 |
| Mexico | Tamaulipas; Altamira | ALT | 22.40 | −97.84 |
| Mexico | Veracruz; La Mancha | MAN | 19.59 | −96.38 |
| Mexico | Tabasco; Olmeca | OLM | 18.23 | −94.06 |
| Primer Name | Repeat Motif | Primer Sequences (5′3′) | Size (bp) | Ta (°C) | GenBank Access | Ref. * |
|---|---|---|---|---|---|---|
| OenhaB105 | (TTG)17 | F: TAGCCTCCTCAAGAGACACA R: CGATGCTGGAACTTCAAAG | 150–160 | 47 | JQ266359 | [48] |
| OenhaD102 | (ATG)8 | F: GCACAAATCCGAAACTCA R: TGCCGCATCTAACCATAC | 255–265 | 47 | JQ266363 | [48] |
| Oenbi2diA_C10 | (GA)20 | F: AGGAGCAAACTGAAGCAGGA R:TTGCAGAACCCAGAATCTGTT | 232–269 | 47 | KT762972 | [47] |
| Oenbi2triA_A1 | (TTC)8 | F: CCACAGCATCACCAAATTCTTACTT R: GGGGCGCCAGGTATTGTCG | 336–355 | 47 | KT762970 | [47] |
| Oenbi2triA_D3 | (ATG)9 | F: CAGATTACGGCGAAAGGAGACAAC R: CGCTCAGGCATCGCATCTC | 300–315 | 47 | KT762967 | [47] |
| Oenbi39tri10 | (CTT)6 | F: AACAAATTTATGCGATTTCGCC R: CTGGAAGGGGCGACTGAAAC | 170–185 | 47 | KT900894 | [47] |
| Oenbi2triA_F5 | (GAT)8 | F: GGGACGCGACCTCAGATTC R: CGCTCAGGCATCGCATCTC | 235–250 | 47 | KT762965 | [47] |
| Oenbi2triA_H1 | (GCT)14 | F: GAGCCGGAATAAACTGATACCACT R: AGCAGAGAAGGCGTCAACCATAAT | 200–217 | 47 | KT762964 | [47] |
| Oenbi39tri4 | (CTT)16 | F: TTTCCTCTCCTTATCTCCTCAATTCTTCTCA R: CTTGCGTTATCTTATGCGTAGCCGTTAGTTTAT | 252–278 | 47 | EF988090 | [48] |
| Altamira (ALT) | La Mancha (MAN) | El Dique (DIQ) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Primer name | N | A | Ap | Ho | He b | N | A | Ap | Ho | He b | N | A | Ap | Ho | He b |
| OenhaB105 | 20 | 2 | - | 0.40 | 0.33 | 20 | 2 | - | 0.30 | 0.50 | 20 | 1 | - | 0 | 0 |
| OenhaD102 | 20 | 2 | - | 0.60 | 0.47 | 20 | 1 | - | 0 | 0 | 20 | 1 | - | 0 | 0 |
| OenbidiA_C10 | 20 | 7 | 4 | 0.45 | 0.85 * | 20 | 5 | 1 | 0.35 | 0.75 * | 20 | 2 | - | 0.65 | 0.45 |
| Oenbi2triA_A1 | 20 | 3 | - | 0.55 | 0.62 * | 20 | 2 | - | 0.20 | 0.43 * | 20 | 2 | - | 0 | 0.33 * |
| Oenbi2triA_D3 | 20 | 2 | - | 0.20 | 0.33 | 20 | 2 | 1 | 0.20 | 0.18 | 20 | 2 | - | 0.30 | 0.26 |
| Oenbi39tri10 | 20 | 3 | 1 | 0.55 | 0.44 | 20 | 3 | 1 | 0.55 | 0.44 | 20 | 1 | - | 0 | 0 |
| Oenbi2triA_F5 | 20 | 2 | - | 0 | 0.38 * | 20 | 3 | 1 | 0.30 | 0.26 | 20 | 1 | - | 0 | 0 |
| Oenbi2triA_H1 | 20 | 3 | 1 | 0.55 | 0.50 | 20 | 2 | - | 0.25 | 0.48 | 20 | 1 | - | 0 | 0 |
| Oenbi39tri4 | 20 | 2 | - | 0.40 | 0.50 | 20 | 5 | 1 | 0.75 | 0.66 | 20 | 3 | - | 0.40 | 0.34 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Espinosa, R.; González-Astorga, J.; Espinosa de los Monteros, A.; Cabrera-Toledo, D.; Gallego-Fernández, J.B. Transferability of Microsatellite Markers Developed in Oenothera spp. to the Invasive Species Oenothera drummondii Hook. (Onagraceae). Diversity 2020, 12, 387. https://doi.org/10.3390/d12100387
Hernández-Espinosa R, González-Astorga J, Espinosa de los Monteros A, Cabrera-Toledo D, Gallego-Fernández JB. Transferability of Microsatellite Markers Developed in Oenothera spp. to the Invasive Species Oenothera drummondii Hook. (Onagraceae). Diversity. 2020; 12(10):387. https://doi.org/10.3390/d12100387
Chicago/Turabian StyleHernández-Espinosa, Raquel, Jorge González-Astorga, Alejandro Espinosa de los Monteros, Dánae Cabrera-Toledo, and Juan B. Gallego-Fernández. 2020. "Transferability of Microsatellite Markers Developed in Oenothera spp. to the Invasive Species Oenothera drummondii Hook. (Onagraceae)" Diversity 12, no. 10: 387. https://doi.org/10.3390/d12100387
APA StyleHernández-Espinosa, R., González-Astorga, J., Espinosa de los Monteros, A., Cabrera-Toledo, D., & Gallego-Fernández, J. B. (2020). Transferability of Microsatellite Markers Developed in Oenothera spp. to the Invasive Species Oenothera drummondii Hook. (Onagraceae). Diversity, 12(10), 387. https://doi.org/10.3390/d12100387