Rodent Assemblages in the Mosaic of Habitat Types in the Zambezian Bioregion

 , ,
, ,  , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Data and Methods
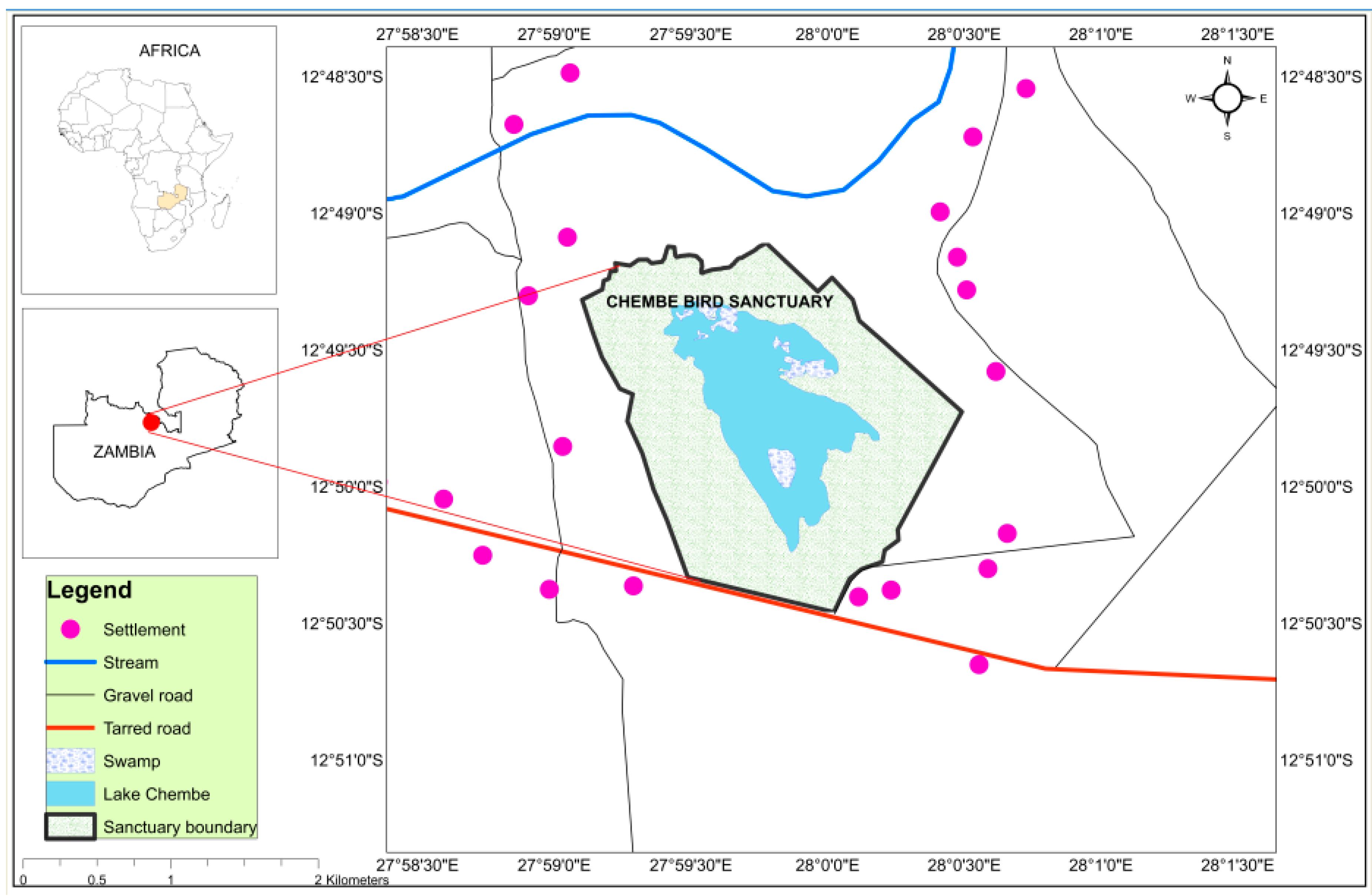
2.1. Study Area
2.2. Data Collection Protocols
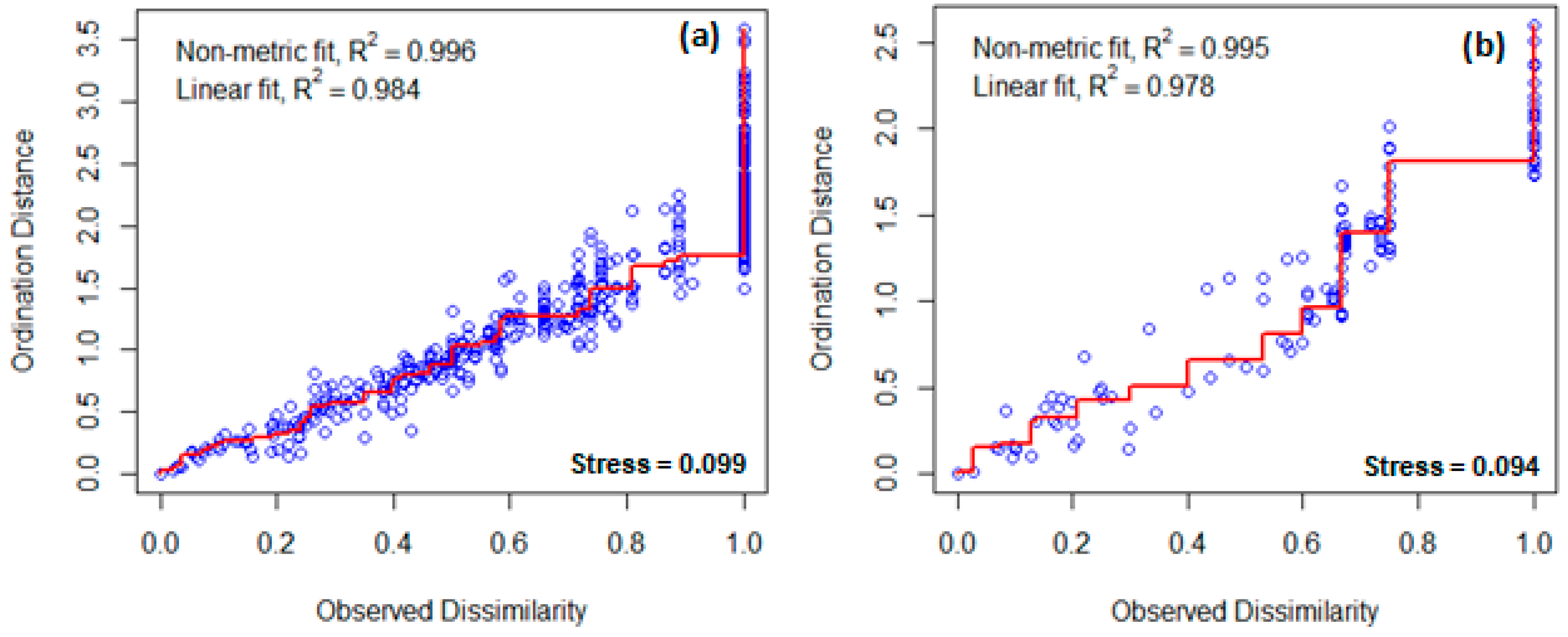
2.3. Data Analysis
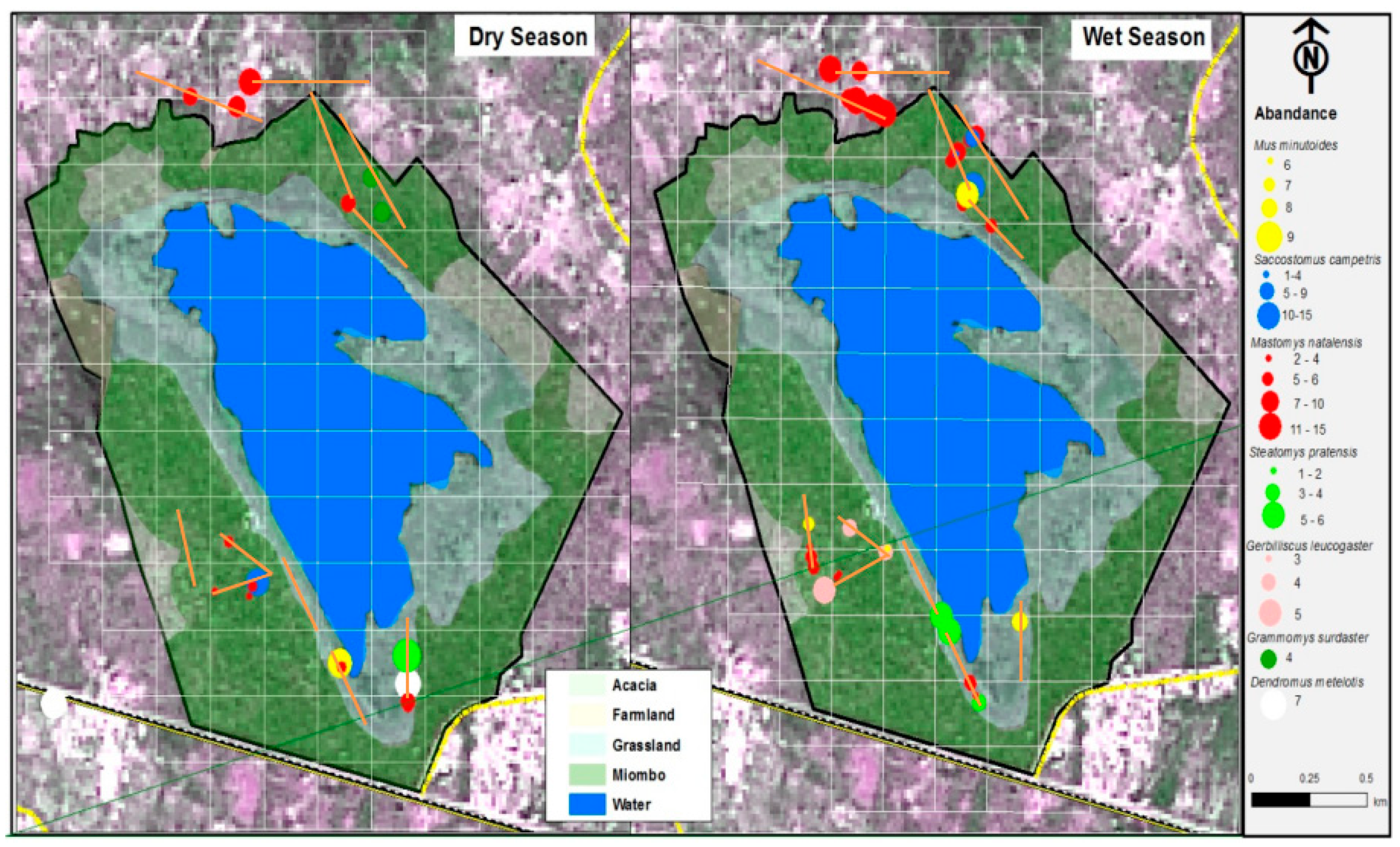
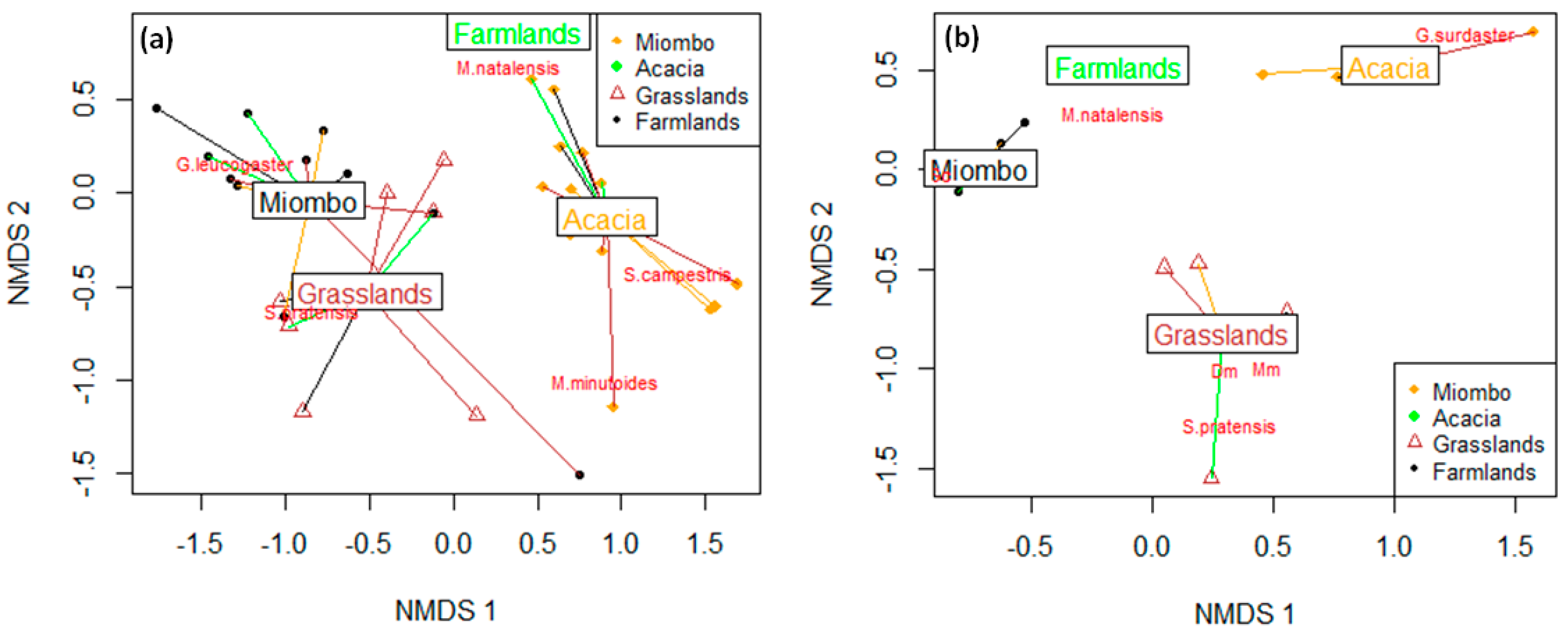
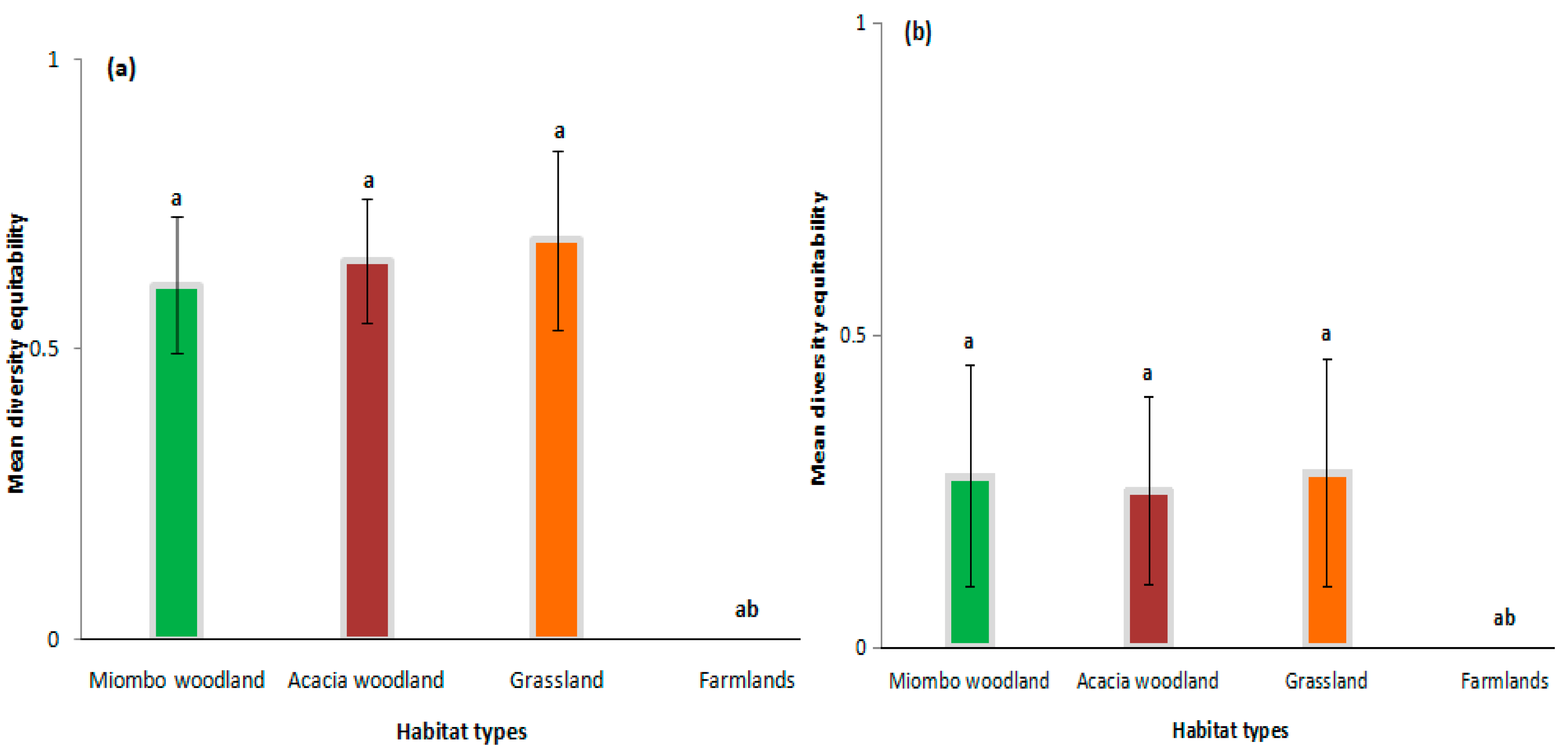
3. Results
4. Discussion
5. Conservation Implications
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kurten, E.L. Cascading effects of contemporaneous defaunation of tropical forest communities. Biol. Conserv. 2013, 163, 22–32. [Google Scholar] [CrossRef]
- Johnson, S.; Burgoyne, P.; Harder, L.D.; Dotterl, S. Mammal pollinators lured by the scent of a parasitic plant. Proc. R. Soc. B 2011, 278, 2303–2310. [Google Scholar] [CrossRef] [PubMed]
- Galiano, D.; Kubiak, B.B.; Overbeck, G.E.; de Freitas, T.R.O. Effects of rodents on plant cover, soil hardness, and soil nutrient content: A case study on tuco-tucos (Ctenomys minutes). Acta Theriol. 2014, 59, 583–587. [Google Scholar] [CrossRef]
- Yong, S.K.; Jalaludin, N.H.; Brau, E.; Shamsudin, N.N.; Heo, C.C. Changes in soil nutrients (ammonia, phosphate and nitrate) associated with rat carcass decomposition under tropical climate conditions. Soil Res. 2019, 57, 482–488. [Google Scholar] [CrossRef]
- Flores-Peredo, R.; Sánchez-Velásquez, L.; Galindo-González, J.; Morales-Mávil, J. Post-dispersed pine seed removal and its effect on seedling establishment in a Mexican Temperate Forest. Plant. Ecol. 2011, 212, 1037–1046. [Google Scholar] [CrossRef]
- Stephens, R.B.; Rowe, R.J. The underappreciated role of rodent generalists in fungal spore-dispersal networks. Ecology 2020. [Google Scholar] [CrossRef] [PubMed]
- Labuschagne, L.; Swanepoel, L.H.; Taylor, P.J.; Belmain, R.R.; Keith, M. Are avian predators effective biological control agents for rodent pest management in agricultural systems? Biol. Control 2016, 101, 94–102. [Google Scholar] [CrossRef]
- Moretti, T.; Benedito, R.O.; Thyssen, P.; Russ Solis, D. Insects on decomposing carcasses of small rodents in a secondary forest in Southeastern Brazil. Eur. J. Entomol. 2008, 105, 691–696. [Google Scholar] [CrossRef]
- Byrom, A.; Ruscoe, W.; Nkwabi, A.; Metzger, K.; Forrester, G.; Craft, M.; Durant, S.; Makacha, S.; Bukombe, J.; Mchetto, J.; et al. Small mammal diversity and population dynamics in the greater Serengeti ecosystem. In Serengeti IV: Sustaining Biodiversity in a Coupled Human-Natural System; Sinclair, A.R.E., Metzger, K.L., Mduma, S.A.R., Fryxell, J.M., Eds.; University of Chicago Press: Chicago, IL, USA, 2015; pp. 323–357. [Google Scholar] [CrossRef]
- Manning, J.; Edge, W. Small mammals responses to fine woody depris and forest fuel reduction in southwest Oregon. J. Wildl. Manag. 2008, 72, 625–632. [Google Scholar] [CrossRef]
- Mulungu, L.S.; Borremans, B.; Ngowo, V.; Mdangi, M.E.; Katakweba, A.S.; Tesha, P.; Mrosso, F.P.; Mchomvu, M.; Kilonzo, B.S. Comparative study of movement patterns of Mastomys natalensis in irrigated rice and fallow fields in eastern Tanzania. Afr. J. Ecol. 2015, 53, 473–479. [Google Scholar] [CrossRef]
- Sellers, L.; Long, R.; Baldwin, R.A.; Jay-Russell, M.; Li, X.; Atwill, E.R.; Engeman, R.M. Impact of field border plantings on rodents and food safety concerns. Proc. Vertebr. Pest. Conf. 2016, 27, 150–155. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, M.; Zhang, H.; Yao, N.; Wang, Y.; Zhang, Y.; Wang, Z. Complex spatial patterns delay the food discovery and locating of rodents. Integr. Zool. 2020. [Google Scholar] [CrossRef]
- Flores-Peredo, R.; Vázquez-Domínquez, Y.G. Influence of vegetation type and season on rodent assemblage in a Mexican temperate forest mosaic. Therya 2016, 3, 357–369. [Google Scholar] [CrossRef]
- Namukonde, N.; Daniel, K.; Jörg, U.G. Differential effects of fire on small mammal communities in Busanga plain, Zambia. Trop. Conserv. Sci. 2017, 10, 1–13. [Google Scholar] [CrossRef]
- Babale, A.; Joshua, A.; Elena, M.; Pierre-Michel, F. Forest disturbance and seasonal food availability influence a conditional seed dispersal mutualism. Biotropica 2018, 50, 750–757. [Google Scholar] [CrossRef]
- Garrido-García, J.A.; Nieto-Lugilde, D.; Alba-Sánchez, F.; Soriguer, R.C. Agricultural intensification during the late Holocene rather than climatic aridification drives the population dynamics and the current conservation status of Microtus cabrerae, an endangered Mediterranean rodent. J. Biogeogr. 2018, 45, 448–460. [Google Scholar] [CrossRef]
- Avenant, N. The potential utility of rodents and other small mammals of indicators of ecosystem integrity of South African grasslands. Wildl. Res. 2011, 38, 626–639. [Google Scholar] [CrossRef]
- Ofori, B.Y.; Attuquayefio, D.K.; Owusu, E.H.; Musah, Y.; Ntiamoa-Baidu, Y. Spatio-temporal variation in small mammal species richness, relative abundance and body mass reveal changes in a coastal wetland ecosystem in Ghana. Environ. Monit. Assess. 2016, 188, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Milstead, B.; Meserve, P.; Campanella, A.; Previtali, K.; Gutierrez, R. Spatial ecology of small mammals in north-central Chile: Role of precipitation and refuges. J. Mammal. 2007, 88, 1532–1538. [Google Scholar] [CrossRef]
- Hurst, Z.M.; McCleery, R.A.; Collier, B.A.; Fletcher, R.J., Jr.; Silvy, N.J.; Taylor, P.J.; Monadjem, A. Dynamic edge effects in small mammal communities across a conservation-agricultural interface in Swaziland. PLoS ONE 2013, 8, e74520. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, A.; Remont, L.; Morotti, L.; Saino, N.; Prigioni, C.; Guidali, F. Multilevel habitat preferences of Apodemus syvaticus and Clethrionomys glareolus in an intensively cultivated agricultural landscape. Ethol. Ecol. Evol. 2017, 29, 38–53. [Google Scholar] [CrossRef]
- Ellis, E.; Ramankutty, N. Putting people in the map: Anthropogenic biomes of the world. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef]
- Monadlem, A.; Mahlaba, T.; Dlamini, N.; Eiscb, S.; Belmain, S. Impact of crop cycle on movement patterns of pest rodent species between fields and houses in Africa. Wildl. Res. 2011, 38, 603–609. [Google Scholar] [CrossRef]
- Nikolić, T.; Radišić, D.; Ćosić, N.; Díaz-Delgado, R.; Milić, D.; Vujić, A.; Ćirović, D. Landscape heterogeneity effects on keystone rodent species: Agro-ecological zoning for conservation of open grasslands. Biodivers. Conserv. 2019. [Google Scholar] [CrossRef]
- Yadok, B.G.; Pech, R.; Chapman, H. Perception of predation risk by African giant pouched rats (Cricetomys sp. nov) is higher in forest-edge microhabitats. Behav. Process. 2019, 168, 103953. [Google Scholar] [CrossRef] [PubMed]
- Pyke, G.H. Optimal foraging theory: A critical review. Annu. Rev. Ecol. Evol. Syst. 1984, 15, 523–575. [Google Scholar] [CrossRef]
- Cooper, W.E., Jr.; Frederick, W.G. Optimal flight initiation distance. J. Theor. Biol. 2007, 224, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Ciuti, S.; Pipia, A.; Ghiandai, F.; Grignolio, S.; Apollonio, M. The key role of lamb presence in affecting flight response in Sardinian mouflon (Ovis orientalis musimon). Behav. Process. 2008, 77, 408–412. [Google Scholar] [CrossRef]
- Linder, H.P.; de Klerk, H.M.; Born, J.; Burgess, N.D.; Fjeldsa, J.; Rahbek, C. The partitioning of Africa: Statistically defined biogeographical regions in sub-Saharan Africa. J. Biogeogr. 2012, 39, 1189–1205. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Biodiversity Synthesis; World Resources Institute: Washington, DC, USA, 2005. [Google Scholar]
- Konečný, A.; Koubek, P.; Bryja, J. Indications of higher density and abundance of small rodents in human-influenced Sudanian savannah than in the Niokolo Koba National Park (Senagal). Afr. J. Ecol. 2010, 48, 718–726. [Google Scholar] [CrossRef]
- Nicolas, V.; Colyn, M. Seasonal variations in population and community structure of small rodents in a tropical forest of Gabon. Can. J. Zool. 2003, 81, 1034–1046. [Google Scholar] [CrossRef]
- Addisu, A.; Bekele, A. Habitat preferences, seasonal abundance and diets of rodents in Alage, southern Ethiopia. Afr. J. Ecol. 2013, 52, 284–291. [Google Scholar] [CrossRef]
- Ramírez-Bautista, A.; Williams, J.N. The importance of productivity and seasonality for structuring small rodent diversity across a tropical elevation gradient. Oecologia 2019, 190, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Magige, F.; Senzota, R. Abundance and diversity of rodents at the human-wildlife interface in Western Serengeti, Tanzania. Afr. J. Ecol. 2006, 44, 371–378. [Google Scholar] [CrossRef]
- Johnson, M.; De León, Y. Effect of an invasive plant and moonlight on rodent foraging behavior in a coastal dune ecosystem. PLoS ONE 2015, 10, e0117903. [Google Scholar] [CrossRef] [PubMed]
- Mazoch, V.; Mikula, O.; Bryja, J.; Konvičková, H.; Russo, I.-R.; Verheyen, E.; Šumbera, R. Phylogeography of a widespread sub-Saharan murid rodent Aethomys chrysophilus: The role of geographic barriers and paleoclimate in the Zambezian bioregion. Mammalia 2018, 82, 373–387. [Google Scholar] [CrossRef]
- Conservation Farming Unit. Conservation Farming and Conservation Agriculture Handbook for Hoe Farmers in Agro-Ecological Regions I and IIa; Conservation Farming Unit: Lusaka, Zambia, 2009. [Google Scholar]
- Nyirenda, V.R.; Musonda, F.; Kambole, S.; Tembo, S. Peasant farmer–raptor conflicts around Chembe Bird Sanctuary, Zambia, Central Africa: Poultry predation, ethno–biology, land use practices and conservation. Anim. Biodivers. Conserv. 2017, 40, 121–132. [Google Scholar] [CrossRef]
- Anthony, N.M.; Ribic, C.A.; Bautz, R.; Garland, T., Jr. Comparative effectiveness of Longworth and Sherman live traps. Wildl. Soc. Bull. 2005, 33, 1018–1026. [Google Scholar] [CrossRef]
- Andrzejewski, R. The home-range concept in rodents revised. Acta Theriol. 2002, 47, 81–101. [Google Scholar] [CrossRef]
- Foord, S.H.; Swanepoel, L.H.; Evans, S.W.; Schoeman, C.S.; Erasmus, B.; Schoeman, M.C.; Keith, M.; Smith, A.; Mauda, E.V.; Maree, N.; et al. Animal taxa contrast in their scale-dependent responses to land use change in rural Africa. PLoS ONE 2018, 13, e0194336. [Google Scholar] [CrossRef]
- Bergeron, P.; Réale, D.; Humphries, M.M.; Garant, D. Anticipation and tracking of pulsed resources drive population dynamics in eastern chipmunks. Ecology 2011, 92, 2027–2034. [Google Scholar] [CrossRef]
- Hansen, N.; Hughes, N.K.; Byrom, A.E.; Banks, P.B. Population recovery of alien black rats Rattus rattus: A test of reinvasion theory. Austral. Ecol. 2020, 45, 291–304. [Google Scholar] [CrossRef]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Anderson, M.J. Distance-based test for homogeneity of multivariate dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef]
- Gijbels, I.; Omelka, M. Testing for homogeneity of multivariate dispersions using dissimilarity measures. Biometrics 2013, 69, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Brady, M.J.; Slade, N.A. Diversity of a grassland rodent community at varying temporal scales: The role of ecologically dominant species. J. Mammal. 2001, 82, 974–983. [Google Scholar] [CrossRef]
- Mortelliti, A.; Boitani, L. Patterns of rodent species diversity and abundance in a Kenyan relict tropical rainforest. Biodivers. Conserv. 2006, 15, 1425–1440. [Google Scholar] [CrossRef]
- Dag, O.; Dolgun, A.; Konar, N.M. Onewaytests: An R package for one-way tests in independent group designs. R J. 2018, 10, 175–199. [Google Scholar] [CrossRef]
- Krebs, C.J. Demographic changes in fluctuating populations of Microtus californicus. Ecol. Monogr. 1966, 36, 239–273. [Google Scholar] [CrossRef]
- Sunyer, P.; Munoz, A.; Mazerolle, M.; Bonal, R.; Espelta, J.M. Wood mouse population dynamics: Interplay among seed abundance seasonality, shrub cover and wild boar interference. Mamm. Biol. 2016, 81, 372–379. [Google Scholar] [CrossRef]
- Massawe, A.W.; Makundi, R.H.; Mulungu, L.S.; Katakweba, A.; Shayo, T.N. Breeding dynamics of rodent species inhabiting farm–fallow mosaic fields in Central Tanzania. Afr. Zool. 2012, 47, 128–137. [Google Scholar] [CrossRef]
- Gasperini, S.; Mortelliti, A.; Bartolommei, P.; Bonacchi, A.; Manzo, E.; Cozzolino, R. Effects of forest management on diversity and survival in three forest rodent species. For. Ecol. Manag. 2016, 382, 151–160. [Google Scholar] [CrossRef]
- Hoset, K.S.; Villers, A.; Wistbacka, R.; Selonen, V. Pulsed food resources, but not forest cover, determine lifetime reproductive success in a forest-dwelling rodent. J. Anim. Ecol. 2017, 86, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Ellison, G.T.H.; Skinner, J.D. The influence of ambient temperature on spontaneous daily torpor in pouched mice (Saccostomus campestris: Rodentia: Cricetidae) from southern Africa. J. Therm. Biol. 1992, 17, 25–31. [Google Scholar] [CrossRef]
- Smith, S.E.; Ramos, F.A.; Refinetti, R.; Farthing, J.P.; Paterson, P.G. Proteinenergy malnutrition induces an aberrant acute-phase response and modifies the circadian rhythm of core temperature. Appl. Physiol. Nutr. Metab. 2013, 38, 844–853. [Google Scholar] [CrossRef]
- Charron, I.; Cabanac, M. Influence of pellet size on rat’s hoarding behaviour. Physiol. Behav. 2004, 82, 447–451. [Google Scholar] [CrossRef]
- Yadok, B.G.; Forget, P.-M.; Gerhard, D.; Chapman, H. Low fruit-crop years of Carapa oreophila drive increased seed removal and predation by scatter hoarding rodents in a West African forest. Acta Oecol. 2019, 99, 103448. [Google Scholar] [CrossRef]
- Mamba, M.; Fasel, N.; Mahlaba, T.A.M.; Austin, J.D.; McCleery, R.A.; Monadjem, A. Influence of sugarcane plantations on the population dynamics and community structure of small mammals in a savanna-agricultural landscape. Glob. Ecol. Conserv. 2019, 82, 250–260. [Google Scholar] [CrossRef]
- Mulungu, L.S.; Mahlaba, T.; Massawe, A.; Kennis, J.; Crauwels, D.; Eiseb, E.; Monadjem, A.; Makundi, R.; Katakweba, A.; Leirs, H.; et al. Dietary preferences of the multimammate mouse (Mastomys natalensis, Smith 1832) across different habitats and seasons in Tanzania and Swaziland. Wildl. Res. 2011, 38, 640–646. [Google Scholar] [CrossRef]
- Taylor, P.J.; Downs, S.; Monadjem, A.; Eiseb, S.J.; Mulungu, L.S.; Massawe, A.W.; Mahlaba, T.A.; Kirsten, F.; Maltitz von, E.; Malebane, P.; et al. Experimental treatment-control studies of ecologically based rodent management in Africa: Balancing conservation and pest management. Wildl. Res. 2012, 39, 51–61. [Google Scholar] [CrossRef]
- Püttker, T.; Barros, C.S.; Pinotti, B.T.; Bueno, A.A.; Pardini, R. Co-occurrence patterns of rodents at multiple spatial scales: Competitive release of generalists following habitat loss? J. Mammal. 2019, 100, 1229–1242. [Google Scholar] [CrossRef]
- Rocha, C.R.; Ribeiro, R.; Marinho-Filho, J. Influence of temporal variation and seasonality on population dynamics of three sympatric rodents. Mamm. Biol. 2017, 84, 20–29. [Google Scholar] [CrossRef]
- Plavsic, M.J. Seasonal dynamics of macrohabitat use by small mammals in the Okavango Delta, Boswana: Implications for landscape-level disturbance resilience. Afr. J. Ecol. 2015, 53, 44–53. [Google Scholar] [CrossRef]
- Mikula, O.; Šumbera, R.; Aghová, T.; Mbau, J.S.; Katakweba, A.S.; Sabuni, C.A.; Bryja, J. Evolutionary history and species diversity of African pouched mice (Rodentia: Nesomyidae: Saccostomus). Zool. Scripta 2016, 45, 595–617. [Google Scholar] [CrossRef]
- Bryja, J.; Šumbera, R.; Peterhans, J.C.K.; Aghová, T.; Bryjová, A.; Mikula, O.; Nicholas, V.; Denys, C.; Verheyen, E.K. Evolutionary history of the thicket rats (genus Grammomys) mirrors the evolution of African forests since the late Miocene. J. Biogeogr. 2017, 44, 182–194. [Google Scholar] [CrossRef]
- Rusch, U.D.; Midgley, J.J.; Anderson, B. Seasonal fluctuations in rodent seed catching and consumption behavior in fynbos shrublands: Implications for fire management. S. Afr. J. Bot. 2014, 93, 217–221. [Google Scholar] [CrossRef]
- McCain, C.M.; King, S.R.B.; Szewczyk, T.; Beck, J. Small mammal species richness is directly linked to regional productivity, but decoupled from food resources, abundance, or habitat complexity. J. Biogeogr. 2018, 45, 2533–2545. [Google Scholar] [CrossRef]
- Makundi, R.H.; Massawe, A.W.; Mulungu, L.S. Breeding seasonality and population dynamics of three rodent species in the Magamba Forest Reserve, Western Usambara Mountains, north-eastern Tanzania. Afr. J. Ecol. 2006, 45, 17–21. [Google Scholar] [CrossRef]
- Massawe, A.W.; Mulungu, L.S.; Makundi, R.H.; Dlamini, N.; Eiseb, S.; Kirsten, F.; Mahlaba, T.; Malebane, P.; von Maltitz, E.; Monadjem, A.; et al. Belmain. Spatial and temporal population dynamics of rodents in three geographically different regions: Implications for ecologically-based rodent management. Afr. Zool. 2011, 46, 393–405. [Google Scholar] [CrossRef]
- Adams, A.; Yihune, M. Species composition, relative abundance and habitat association of rodents in Yekoche Forest, East Gojjam, Ethiopia. Int. J. Biodivers. Conserv. 2016, 8, 216–223. [Google Scholar] [CrossRef]
- Larsen, A.L.; Homyack, J.A.; Bently-Wigley, T.; Miller, D.A.; Kalcounis-Rueppell, M.C. Effects of habitat modification on cotton rat population dynamics and rodent community structure. For. Ecol. Manag. 2016, 376, 238–246. [Google Scholar] [CrossRef]
- Fleming, P.A.; Loveridge, J.P. Miombo woodland termite mounds: Resource islands for small vertebrates? J. Zool. 2003, 259, 161–168. [Google Scholar] [CrossRef]
- Mlyashimbi, E.C.M.; Marien, J.; Kimaro, D.N.; Tarimo, A.J.P.; Isabirye, M.; Makundi, R.H.; Massawe, A.W.; Mdangi, M.E.; Kifumba, D.; Nakiyemba, A.; et al. Relationships between seasonal changes in diet of Multimammate rat and its breeding patterns in sermi-arid areas in Tanzania. Cogent Food Agric. 2018, 4, 1507–1553. [Google Scholar] [CrossRef]
- Martin, J.M.; Branch, L.C.; Rain, R.N.; Beyeler, S.C. Temporal instability of agricultural habitat reduces reproductive success of Barn owls (Tyto alba). Auk 2010, 127, 909–916. [Google Scholar] [CrossRef]
- Madsen, T.; Ujvari, B.; Shine, R.; Buttemer, W.; Olsson, M. Size matters: Extraordinary rodent abundance on an Australian tropical flood plain. Austral. Ecol. 2006, 31, 361–365. [Google Scholar] [CrossRef]
- Damschen, E.I.; Brudvig, L.A.; Burt, M.A.; Fletcher, R.J.; Haddad, N.M.; Levey, D.J.; Orrock, J.L.; Resasco, J.; Tewksbury, J.J. Ongoing accumulation of plant diversity through habitat connectivity in an 18-year experiment. Science 2019, 365, 1478–1480. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 2001; Volume 1. [Google Scholar]
- Allan, J.R.; Possingham, H.P.; Atkinson, S.C.; Waldron, A.; Di Marco, M.; Adams, V.M.; Butchart, S.H.M.; Venter, O.; Maron, M.; Williams, B.A.; et al. Conservation attention necessary across at least 44% of Earth’s terrestrial area to safeguard biodiversity. bioRxiv 2019, 839977. [Google Scholar] [CrossRef]
- Dinerstein, E.; Vynne, C.; Sala, E.; Joshi, A.R.; Fernando, S.; Lovejoy, T.E.; Mayorga, J.; Olson, D.; Asner, G.P.; Baillie, J.E.M. A global deal for nature: Guiding principles, milestones, and targets. Sci. Adv. 2019, 5, eaaw2869. [Google Scholar] [CrossRef] [PubMed]
- Snyman, S. The role of ecotourism employment in poverty reduction and community perceptions of conservation and tourism in Southern Africa. J. Sustain. Tour. 2012, 20, 395–416. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitats | No. Transects | Trap Nights | Total No. Rodents Captured * | Total No. Rodents Captured * | Mean Diversity (H’) | Mean Diversity Equitability of Pielou (J’) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mastomys Natalensis | Mus Minutoides | Saccostomus Campestris | Steatomys Pratensis | Gerbilliscus Leucogaster | Grammomys Surdaster | Dendromus Melanotis | ||||||
| Miombo woodland | 3 | 840 | 27 (15) [34] | 13 (0) [13] | 0 (6) [6] | 0 (0) [0] | 16 (0) [16] | 0 (0) [0] | 0 (0) [0] | 56 (21) [69] | 0.611 ± 0.12 (0.274 ± 0.18) | 0.856 (0.881) |
| Acacia woodland | 2 | 840 | 59 (7) [61] | 10 (0) [10] | 24 (0) [24] | 0 (0) [0] | 0 (0) [0] | 0 (8) [8] | 0 (0) [0] | 93 (15) [103] | 0.653 ± 0.11 (0.250 ± 0.15) | 0.799 (0.998) |
| Grasslands | 3 | 840 | 14 (13) [19] | 8 (7) [11] | 0 (0) [0] | 20 (10) [23] | 0 (0) [0] | 0 (0) [0] | 0 (7) [7] | 42 (37) [60] | 0.688 ± 0.15 (0.280 ± 0.18) | 0.086 (0.980) |
| Farmlands | 2 | 840 | 85 (28) [94] | 0 (0) [0] | 0 (0) [0] | 0 (0) [0] | 0 (0) [0] | 0 (0) [0] | 0 (0) [0] | 85 (28) [94] | 0 (0) | 0 (0) |
| Total | 10 | 3360 | 185 (63) [208] | 31 (7) [34] | 24 (6) [30] | 20 (10) [23] | 16 (0) [16] | 0 (8) [8] | 0 (7) [7] | 276 (101) [326] | - | - |
| No. & percentage of rodents recaptured | - | - | n = 14; 7.0% (n = 2; 3.1%) | n = 3; 8.8% (n = 1; 12.5%) | n = 3; 11.1% (n = 1; 14.3%) | n = 1; 4.8% (n = 1; 9.1%) | n = 2; 11.1% (NA) | NA (n = 1; 11.1%) | NA (n = 1; 12.5%) | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nyirenda, V.R.; Namukonde, N.; Simwanda, M.; Phiri, D.; Murayama, Y.; Ranagalage, M.; Milimo, K. Rodent Assemblages in the Mosaic of Habitat Types in the Zambezian Bioregion. Diversity 2020, 12, 365. https://doi.org/10.3390/d12100365
Nyirenda VR, Namukonde N, Simwanda M, Phiri D, Murayama Y, Ranagalage M, Milimo K. Rodent Assemblages in the Mosaic of Habitat Types in the Zambezian Bioregion. Diversity. 2020; 12(10):365. https://doi.org/10.3390/d12100365
Chicago/Turabian StyleNyirenda, Vincent R., Ngawo Namukonde, Matamyo Simwanda, Darius Phiri, Yuji Murayama, Manjula Ranagalage, and Kaula Milimo. 2020. "Rodent Assemblages in the Mosaic of Habitat Types in the Zambezian Bioregion" Diversity 12, no. 10: 365. https://doi.org/10.3390/d12100365
APA StyleNyirenda, V. R., Namukonde, N., Simwanda, M., Phiri, D., Murayama, Y., Ranagalage, M., & Milimo, K. (2020). Rodent Assemblages in the Mosaic of Habitat Types in the Zambezian Bioregion. Diversity, 12(10), 365. https://doi.org/10.3390/d12100365