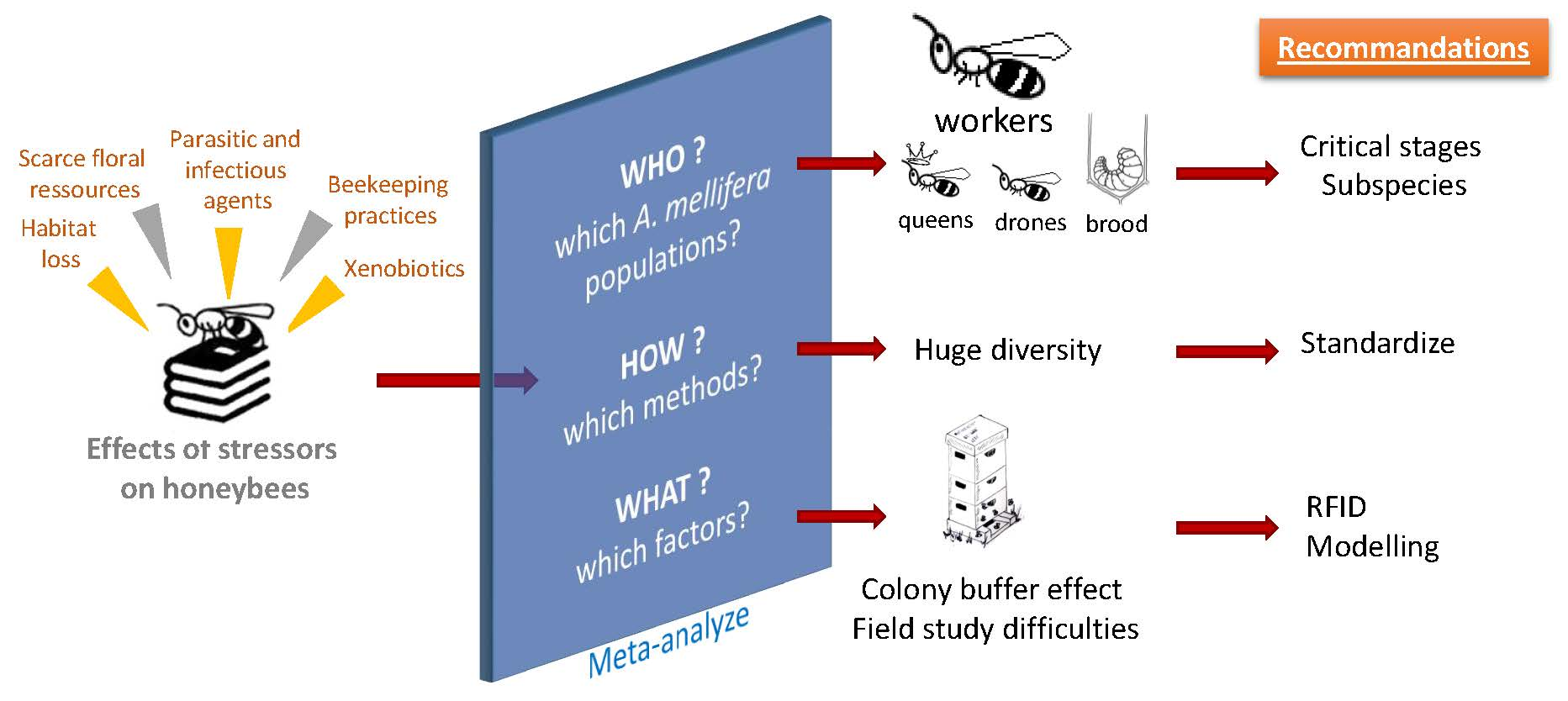
Impact of Stressors on Honey Bees (Apis mellifera; Hymenoptera: Apidae): Some Guidance for Research Emerge from a Meta-Analysis

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Identification of the Key Concepts and the Relevant Keywords
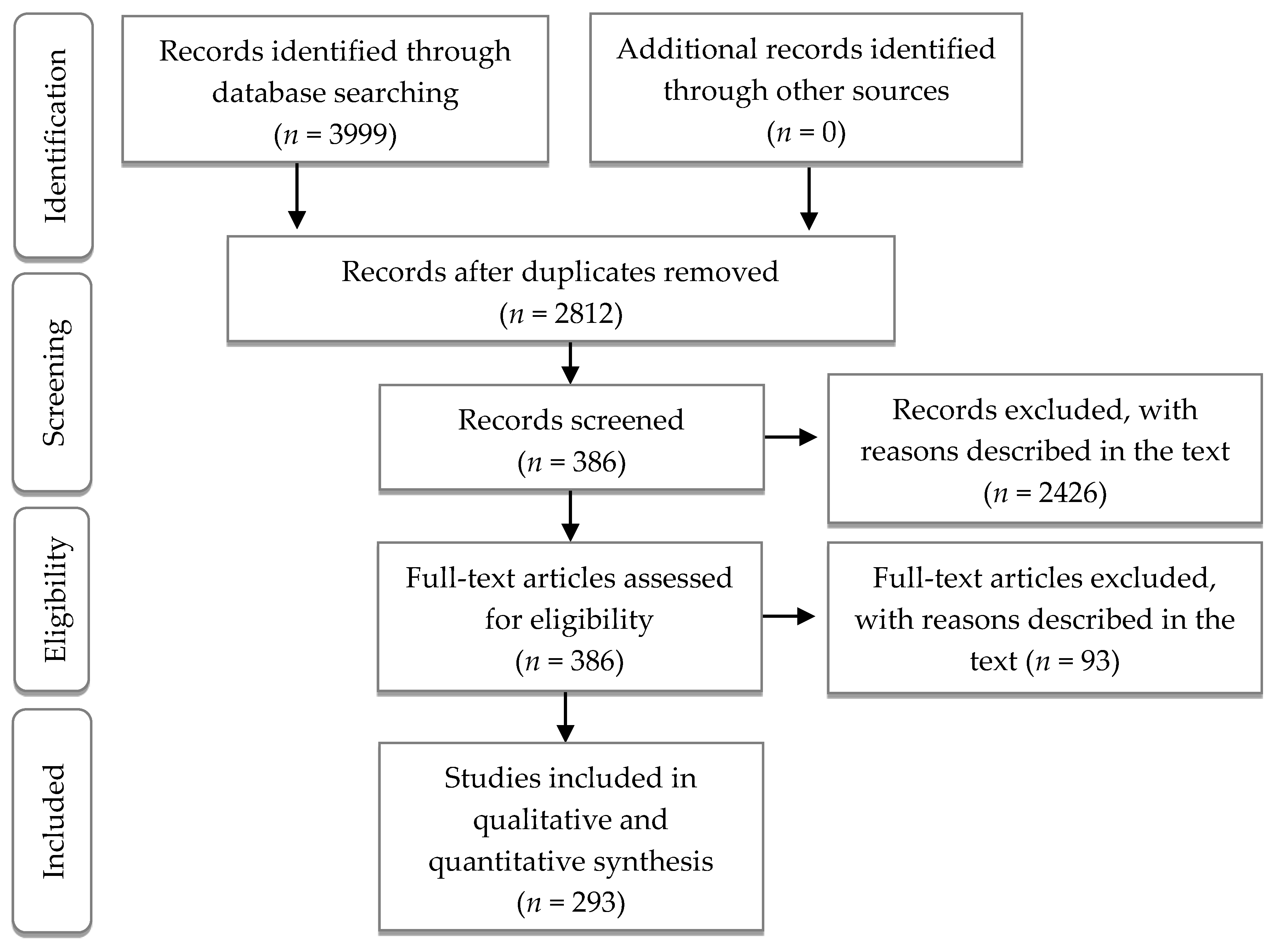
2.2. Literature Search
2.3. Data Extraction
2.4. Data Analysis
3. Results and Discussion
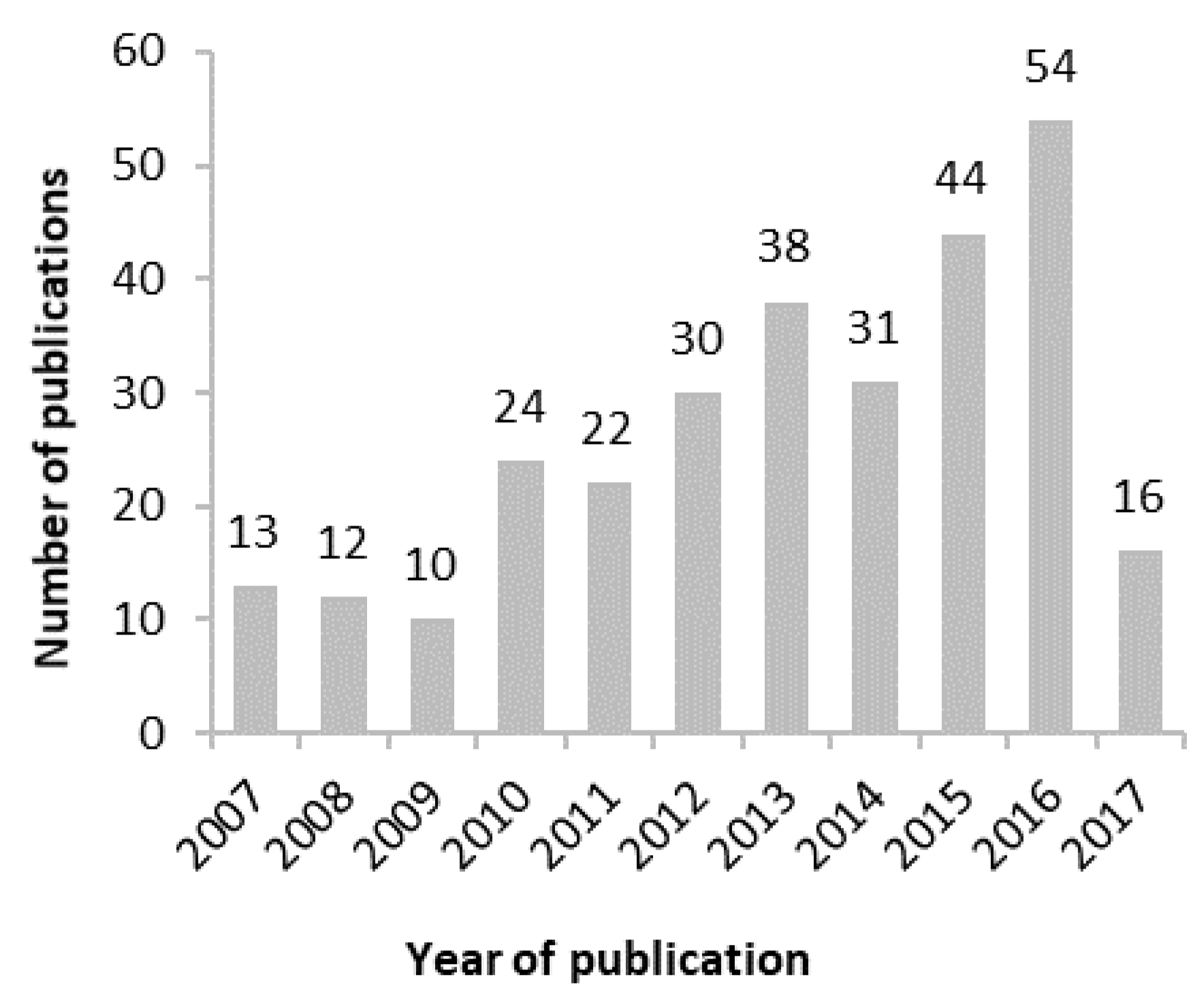
3.1. General Information
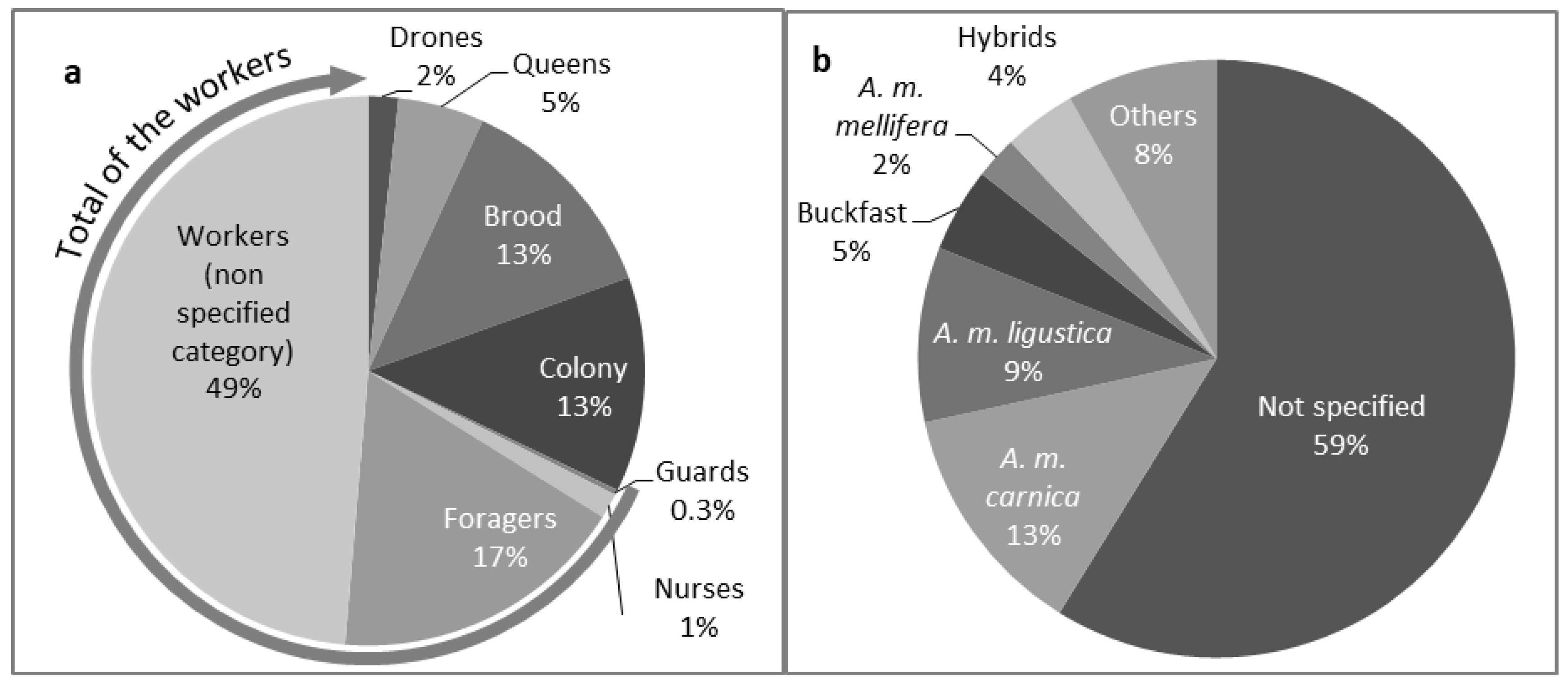
3.2. What Kind of Populations were Studied in the Articles?
3.3. What Stressors Were Studied and Did They Have an Impact on Bee Health?
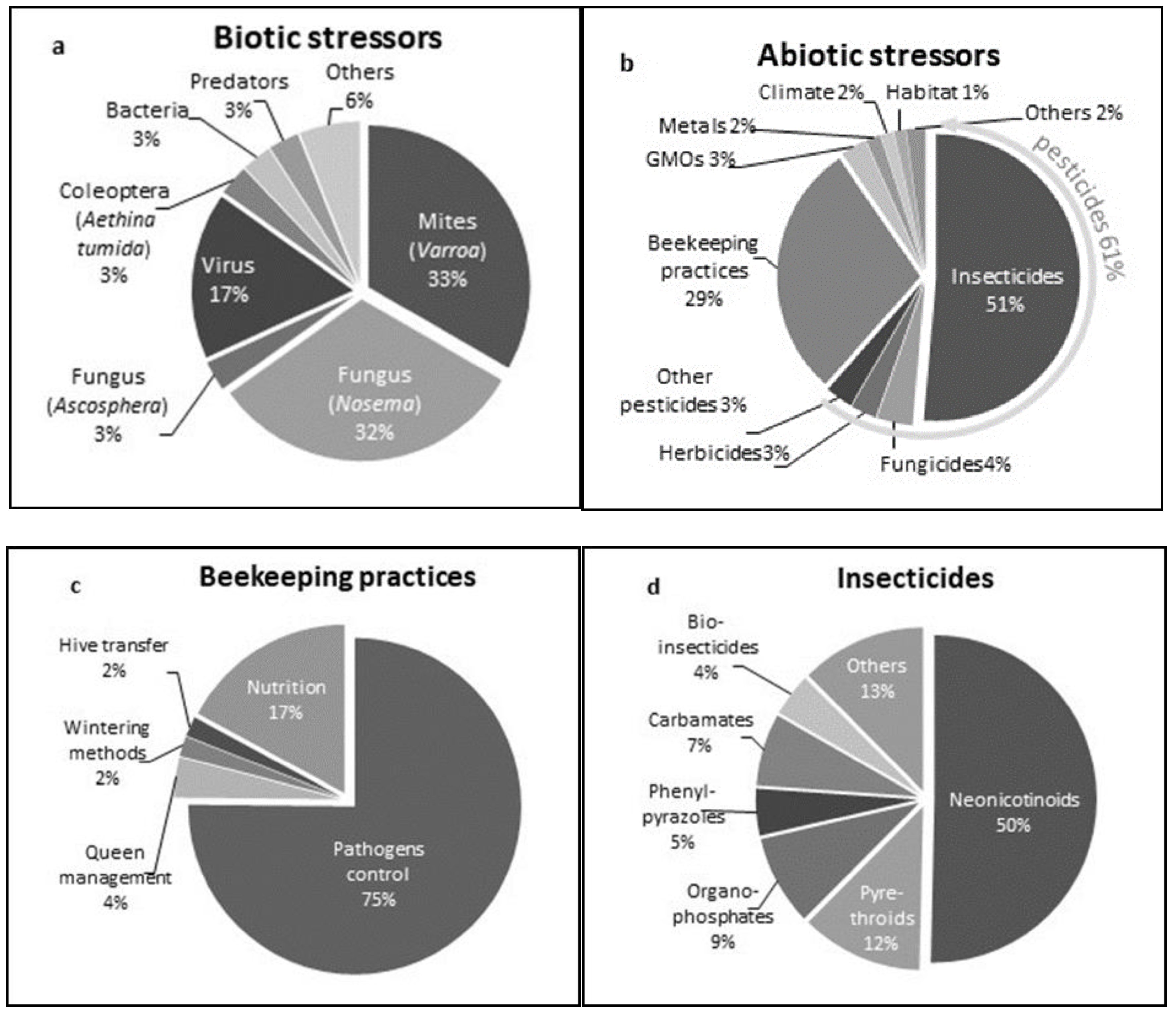
3.3.1. Biotic Stressors
3.3.2. Abiotic Stressors
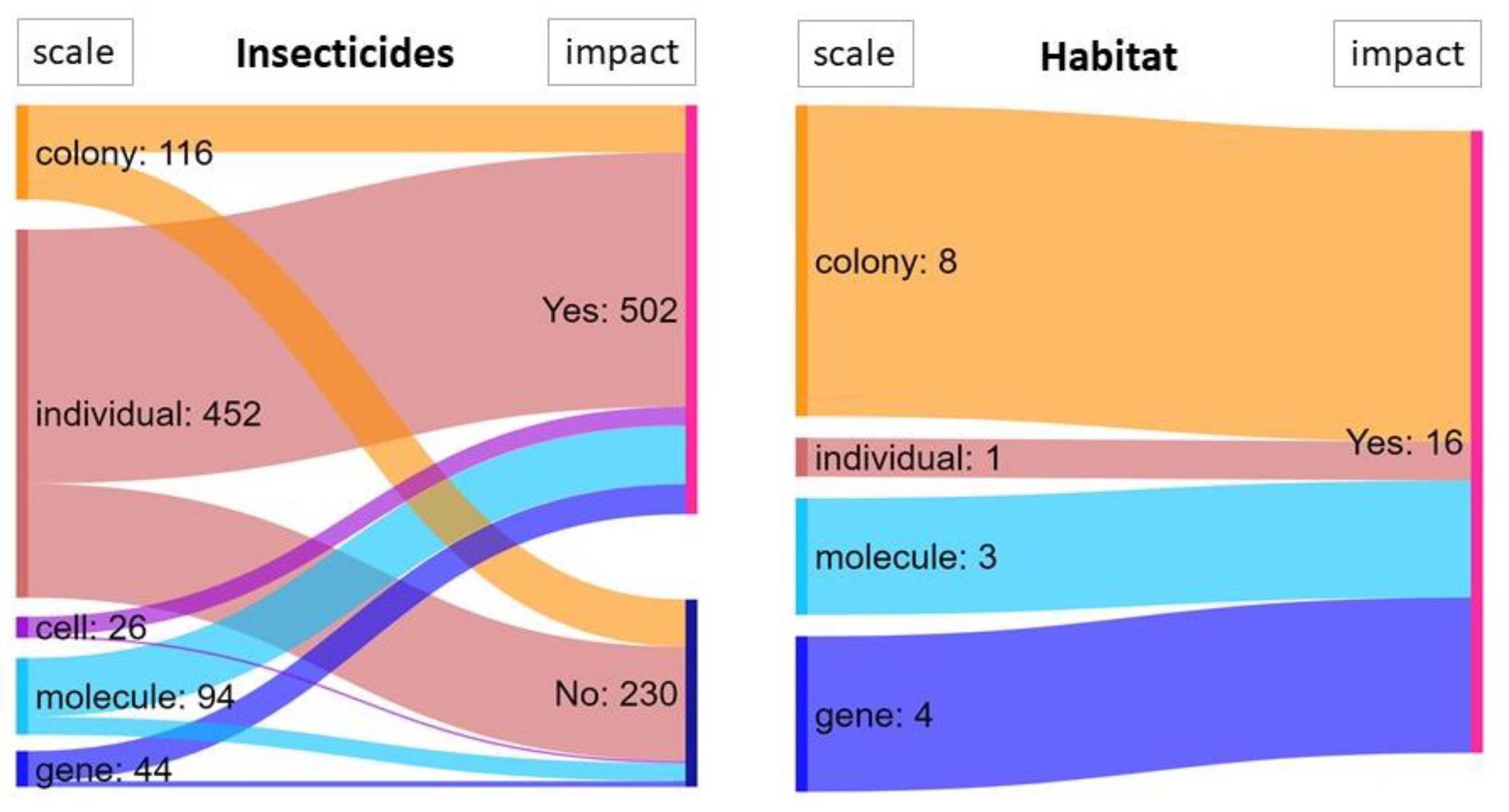
3.3.3. Stressors’ Impact
3.3.4. Co-Exposures
3.4. What Methods Were Used in the Articles to Measure Various Endpoints?
3.4.1. Colony Endpoints
3.4.2. Individual Endpoints
3.5. Did the Evidence of an Impact Vary According to the Type of Study or to the Scale of Study?
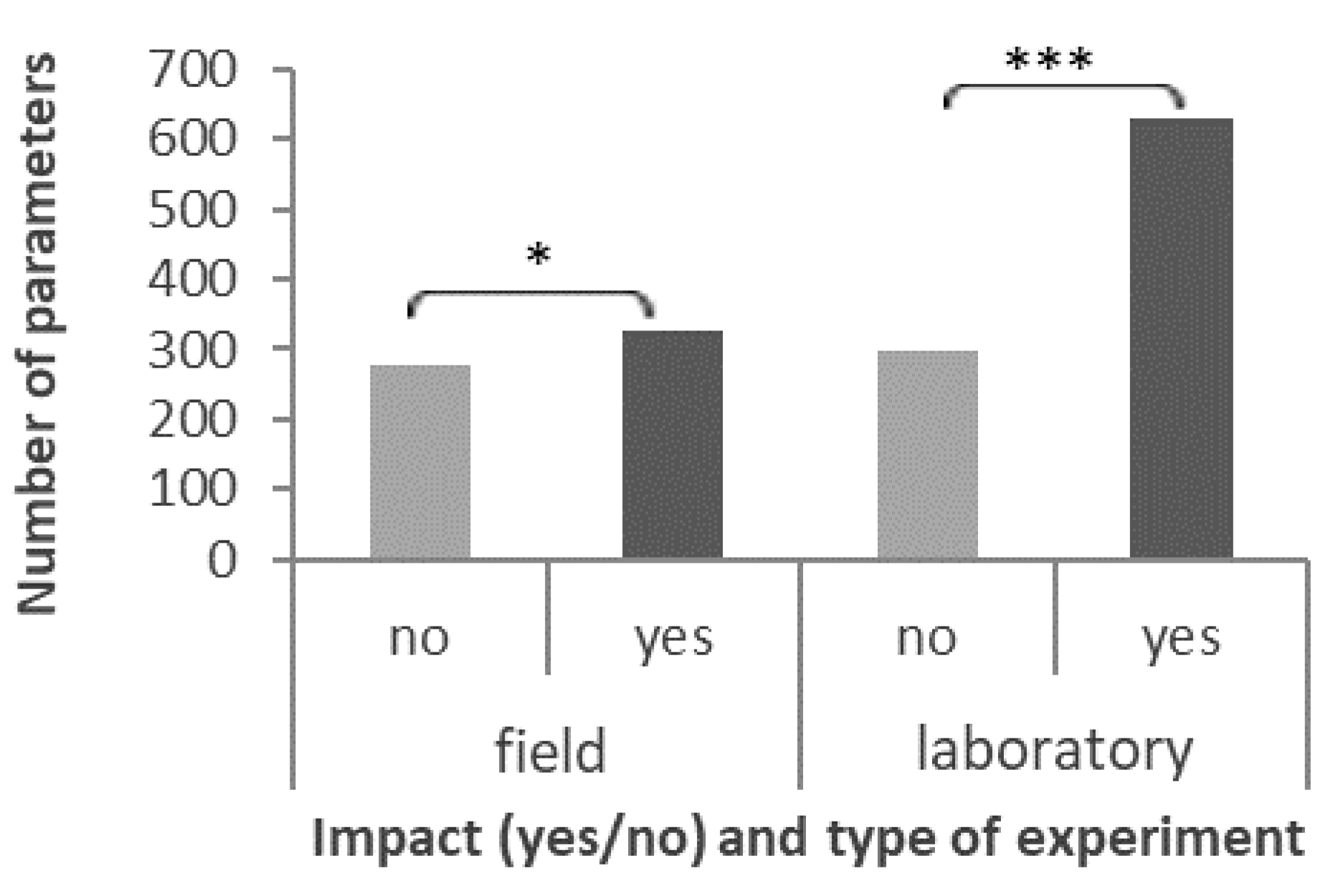
3.5.1. Type of Study
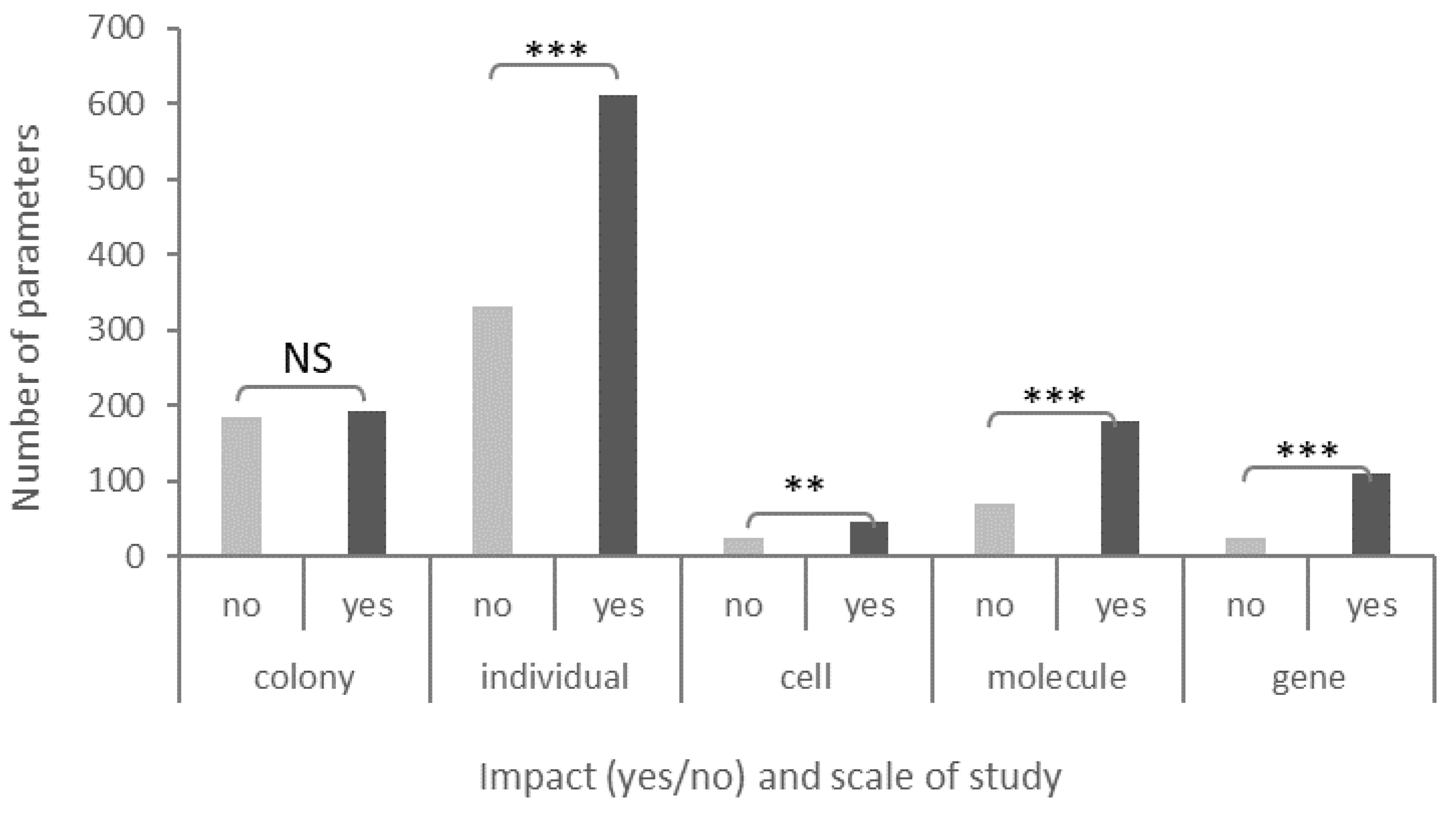
3.5.2. Scale of Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Pro. Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Decourtye, A.; Devillers, J. Ecotoxicity of neonicotinoid insecticides to bees. Adv. Exp. Med. Biol. 2010, 683, 85–95. [Google Scholar] [PubMed]
- Allen-Wardell, G.; Bernhardt, P.; Bitner, R.; Burquez, A.; Buchmann, S.; Cane, J.; Cox, P.; Dalton, V.; Feinsinger, P.; Ingram, M.; et al. The Potential Consequences of Pollinator Declines on the Conservation of Biodiversity and Stability of Food Crop Yields. Conserv. Biol. 1998, 12, 8–17. [Google Scholar]
- Vanengelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103 (Suppl. 1), S80–S95. [Google Scholar] [CrossRef] [PubMed]
- Benaets, K.; Van Geystelen, A.; Cardoen, D.; De Smet, L.; de Graaf, D.C.; Schoofs, L.; Larmuseau, M.H.D.; Brettell, L.E.; Martin, S.J.; Wenseleers, T. Covert deformed wing virus infections have long-term deleterious effects on honeybee foraging and survival. Proc. Biol. Sci. 2017, 284, 20162149. [Google Scholar] [CrossRef]
- Alburaki, M.; Cheaib, B.; Quesnel, L.; Mercier, P.; Chagnon, M.; Derome, N. Performance of honeybee colonies located in neonicotinoid-treated and untreated cornfields in Quebec. J. Appl. Entomol. 2016, 141, 112–121. [Google Scholar] [CrossRef]
- Fine, J.D.; Cox-Foster, D.L.; Mullin, C.A. An Inert Pesticide Adjuvant Synergizes Viral Pathogenicity and Mortality in Honey Bee Larvae. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 125–957. [Google Scholar] [CrossRef]
- Christen, V.; Bachofer, S.; Fent, K. Binary mixtures of neonicotinoids show different transcriptional changes than single neonicotinoids in honeybees (Apis mellifera). Environ. Pollut. 2017, 220, 1264–1270. [Google Scholar] [CrossRef]
- Le Conte, Y.; Navajas, M. Climate change: Impact on honey bee populations and diseases. Rev.-Off. Int. Epizoot. 2008, 27, 485–497. [Google Scholar] [PubMed]
- Henry, M.; Cerrutti, N.; Aupinel, P.; Decourtye, A.; Gayrard, M.; Odoux, J.-F.; Pissard, A.; Rüger, C.; Bretagnolle, V. Reconciling laboratory and field assessments of neonicotinoid toxicity to honeybees. Proc. Biol. Sci. 2015, 282, 20152110. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Eisenhardt, D.; Rademacher, E. Sublethal effects of oxalic acid on Apis mellifera (Hymenoptera: Apidae): Changes in behaviour and longevity. Apidologie 2012, 43, 218–225. [Google Scholar] [CrossRef]
- Aliouane, Y.; El Hassani, A.K.; Gary, V.; Armengaud, C.; Lambin, M.; Gauthier, M. Subchronic exposure of honeybees to sublethal doses of pesticides: Effects on behavior. Environ. Toxicol. Chem. 2009, 28, 113–122. [Google Scholar] [CrossRef]
- Bonnafé, E.; Alayrangues, J.; Hotier, L.; Massou, I.; Renom, A.; Souesme, G.; Marty, P.; Allaoua, M.; Treilhou, M.; Armengaud, C. Monoterpenoid-based preparations in beehives affect learning, memory, and gene expression in the bee brain. Environ. Toxicol. Chem. 2017, 36, 337–345. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Higes, M.; Sagastume, S.; Juarranz, Á.; Dias-Almeida, J.; Budge, G.E.; Meana, A.; Boonham, N. Microsporidia infection impacts the host cell’s cycle and reduces host cell apoptosis. PLoS ONE 2017, 12, e0170183. [Google Scholar] [CrossRef]
- Christen, V.; Mittner, F.; Fent, K. Molecular Effects of Neonicotinoids in Honey Bees (Apis mellifera). Environ. Sci. Technol. 2016, 50, 4071–4081. [Google Scholar] [CrossRef]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; McField, M.; et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2015, 22, 68–102. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Towards an integrated environmental risk assessment of multiple stressors on bees: Review of research projects in Europe, knowledge gaps and recommendations. EFSA J. 2014, 12, 3594. [Google Scholar]
- Benuszak, J.; Laurent, M.; Chauzat, M.-P. The exposure of honey bees (Apis mellifera; Hymenoptera: Apidae) to pesticides: Room for improvement in research. Sci. Total Environ. 2017, 587, 423–438. [Google Scholar] [CrossRef]
- Johnson, R.M.; Dahlgren, L.; Siegfried, B.D.; Ellis, M.D. Effect of in-hive miticides on drone honey bee survival and sperm viability. J. Apic. Res. 2013, 52, 88–95. [Google Scholar] [CrossRef]
- Alaux, C.; Folschweiller, M.; McDonnell, C.; Beslay, D.; Cousin, M.; Dussaubat, C.; Brunet, J.-L.; Le Conte, Y. Pathological effects of the microsporidium Nosema ceranae on honey bee queen physiology (Apis mellifera). J. Invertebr. Pathol. 2011, 106, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Rangel, J.; Tarpy, D.R. The combined effects of miticides on the mating health of honey bee (Apis mellifera L.) queens. J. Apic. Res. 2015, 54, 275–283. [Google Scholar] [CrossRef]
- Williams, G.R.; Troxler, A.; Retschnig, G.; Roth, K.; Yañez, O.; Shutler, D.; Neumann, P.; Gauthier, L. Neonicotinoid pesticides severely affect honey bee queens. Sci. Rep. 2015, 5, 1–8. [Google Scholar] [CrossRef]
- Chaimanee, V.; Evans, J.D.; Chen, Y.; Jackson, C.; Pettis, J.S. Sperm viability and gene expression in honey bee queens (Apis mellifera) following exposure to the neonicotinoid insecticide imidacloprid and the organophosphate acaricide coumaphos. J. Insect Physiol. 2016, 89, 1–8. [Google Scholar] [CrossRef]
- Dussaubat, C.; Maisonnasse, A.; Crauser, D.; Tchamitchian, S.; Bonnet, M.; Cousin, M.; Kretzschmar, A.; Brunet, J.-L.; Le Conte, Y. Combined neonicotinoid pesticide and parasite stress alter honeybee queens’ physiology and survival. Sci. Rep. 2016, 6, 31430. [Google Scholar] [CrossRef]
- Kairo, G.; Provost, B.; Tchamitchian, S.; Ben Abdelkader, F.; Bonnet, M.; Cousin, M.; Sénéchal, J.; Benet, P.; Kretzschmar, A.; Belzunces, L.P.; et al. Drone exposure to the systemic insecticide Fipronil indirectly impairs queen reproductive potential. Sci. Rep. 2016, 6, 31904. [Google Scholar] [CrossRef]
- Sturup, M.; Baer-Imhoof, B.; Nash, D.R.; Boomsma, J.J.; Baer, B. When every sperm counts: Factors affecting male fertility in the honeybee Apis mellifera. Behav. Ecol. 2013, 24, 1192–1198. [Google Scholar] [CrossRef]
- Suchail, S.; Guez, D.; Belzunces, L. Characteristics of imidacloprid toxicity in two Apis mellifera subspecies. Environ. Toxicol. Chem. 2000, 19, 1901–1905. [Google Scholar] [CrossRef]
- Rinkevich, F.D.; Margotta, J.W.; Pittman, J.M.; Danka, R.G.; Tarver, M.R.; Ottea, J.A.; Healy, K.B. Genetics, Synergists, and Age Affect Insecticide Sensitivity of the Honey Bee, Apis mellifera. PLoS ONE 2015, 10, e0139841. [Google Scholar] [CrossRef]
- Giangaspero, M.; Turno, P. Aethina tumida, an Exotic Parasite of Bees. Clin. Microbiol. 2015, 4, 1000e128. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103 (Suppl. 1), S96–S119. [Google Scholar] [CrossRef]
- Jacques, A.; Laurent, M.; Ribière-Chabert, M.; Saussac, M.; Bougeard, S.; Budge, G.E.; Hendrikx, P.; Chauzat, M.-P. A pan-European epidemiological study reveals honey bee colony survival depends on beekeeper education and disease control. PLoS ONE 2017, 12, e0172591. [Google Scholar] [CrossRef]
- Yi, D.; Fang, Z.; Yang, L. Effects of Bt cabbage pollen on the honeybee Apis mellifera L. Sci. Rep. 2018, 8, 1–6. [Google Scholar] [CrossRef]
- Henry, M.; Becher, M.A.; Osborne, J.L.; Kennedy, P.J.; Aupinel, P.; Bretagnolle, V.; Brun, F.; Grimm, V.; Horn, J.; Requier, F. Predictive systems models can help elucidate bee declines driven by multiple combined stressors. Apidologie 2017, 48, 328–339. [Google Scholar] [CrossRef]
- OECD Test No. 213: Honeybees, Acute Oral Toxicity Test; OECD: Paris, France, 1998.
- OECD Test No. 214: Honeybees, Acute Contact Toxicity Test; OECD: Paris, France, 1998.
- OECD Test No. 245: Honey Bee (Apis mellifera L.), Chronic Oral Toxicity Test (10-Day Feeding); OECD: Paris, France, 2017.
- OECD Test No. 237: Honey Bee (Apis mellifera) Larval Toxicity Test, Single Exposure; OECD: Paris, France, 2013.
- OECD Series on Testing & Assessment No. 239: Guidance Document on Honey Bee (Apis mellifera) Larval Toxicity Test following Repeated Exposure; OECD: Paris, France, 2016.
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.-F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A common pesticide decreases foraging success and survival in honey bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef]
- Dietemann van der Zee, V.; Ellis, J.; Neumann, J. The COLOSS BEEBOOK, Volume I: Standard methods for Apis mellifera research. J. Apic. Res. 2013, 52. [Google Scholar] [CrossRef]
- Woodcock, B.A.; Heard, M.S.; Jitlal, M.S.; Rundlöf, M.; Bullock, J.M.; Shore, R.F.; Pywell, R.F. Replication, effect sizes and identifying the biological impacts of pesticides on bees under field conditions. J. Appl. Ecol. 2016, 53, 1358–1362. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havard, T.; Laurent, M.; Chauzat, M.-P. Impact of Stressors on Honey Bees (Apis mellifera; Hymenoptera: Apidae): Some Guidance for Research Emerge from a Meta-Analysis. Diversity 2020, 12, 7. https://doi.org/10.3390/d12010007
Havard T, Laurent M, Chauzat M-P. Impact of Stressors on Honey Bees (Apis mellifera; Hymenoptera: Apidae): Some Guidance for Research Emerge from a Meta-Analysis. Diversity. 2020; 12(1):7. https://doi.org/10.3390/d12010007
Chicago/Turabian StyleHavard, Tiphaine, Marion Laurent, and Marie-Pierre Chauzat. 2020. "Impact of Stressors on Honey Bees (Apis mellifera; Hymenoptera: Apidae): Some Guidance for Research Emerge from a Meta-Analysis" Diversity 12, no. 1: 7. https://doi.org/10.3390/d12010007
APA StyleHavard, T., Laurent, M., & Chauzat, M.-P. (2020). Impact of Stressors on Honey Bees (Apis mellifera; Hymenoptera: Apidae): Some Guidance for Research Emerge from a Meta-Analysis. Diversity, 12(1), 7. https://doi.org/10.3390/d12010007