Patterns in Alpha and Beta Phytoplankton Diversity along a Conductivity Gradient in Coastal Mediterranean Lagoons

 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
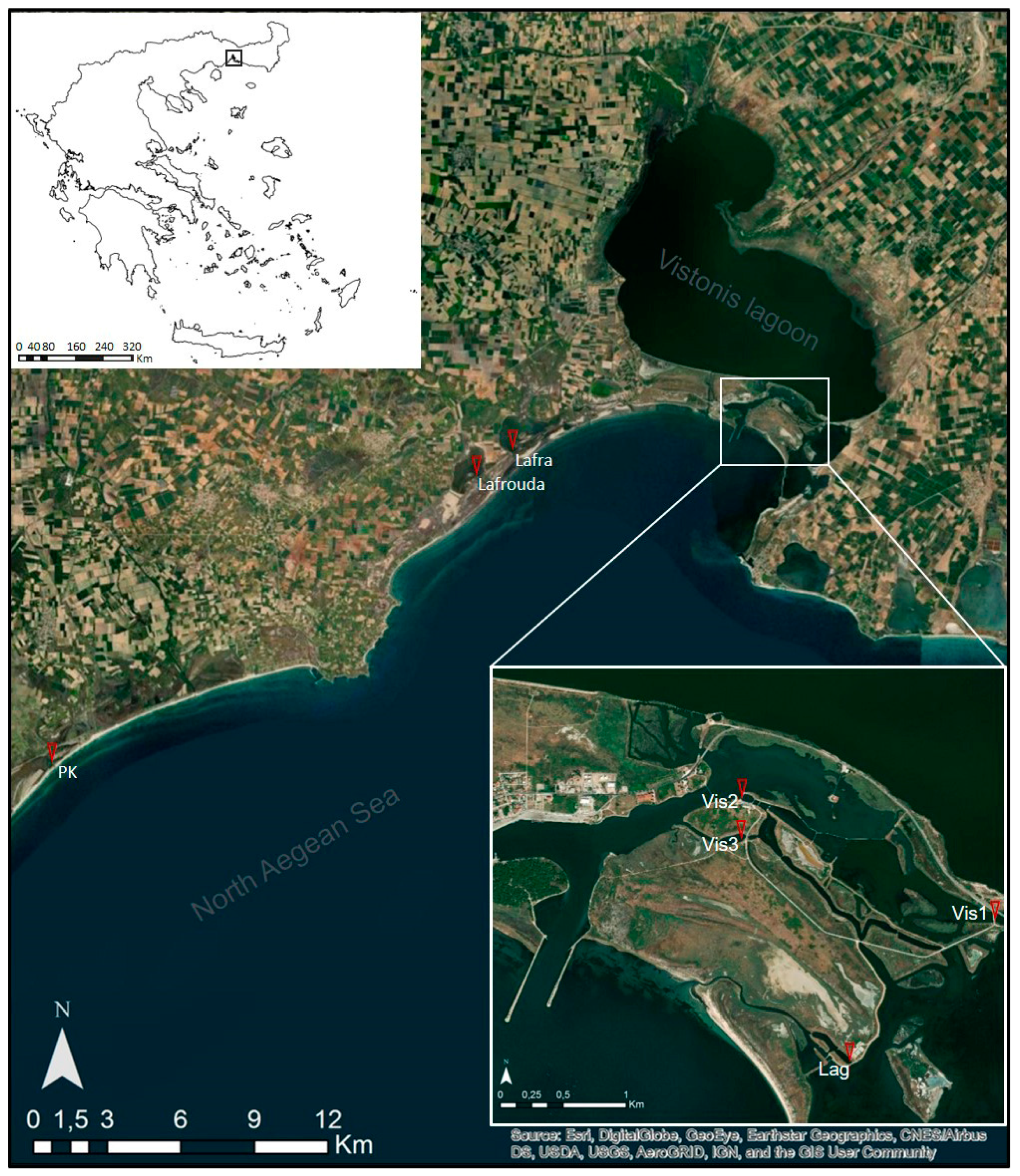
2.1. Sampling Sites and Sample Collection
2.2. Microscopy Analysis
2.3. Diversity
2.4. Non-Linear Regression Models
2.5. Statistical Analysis
3. Results
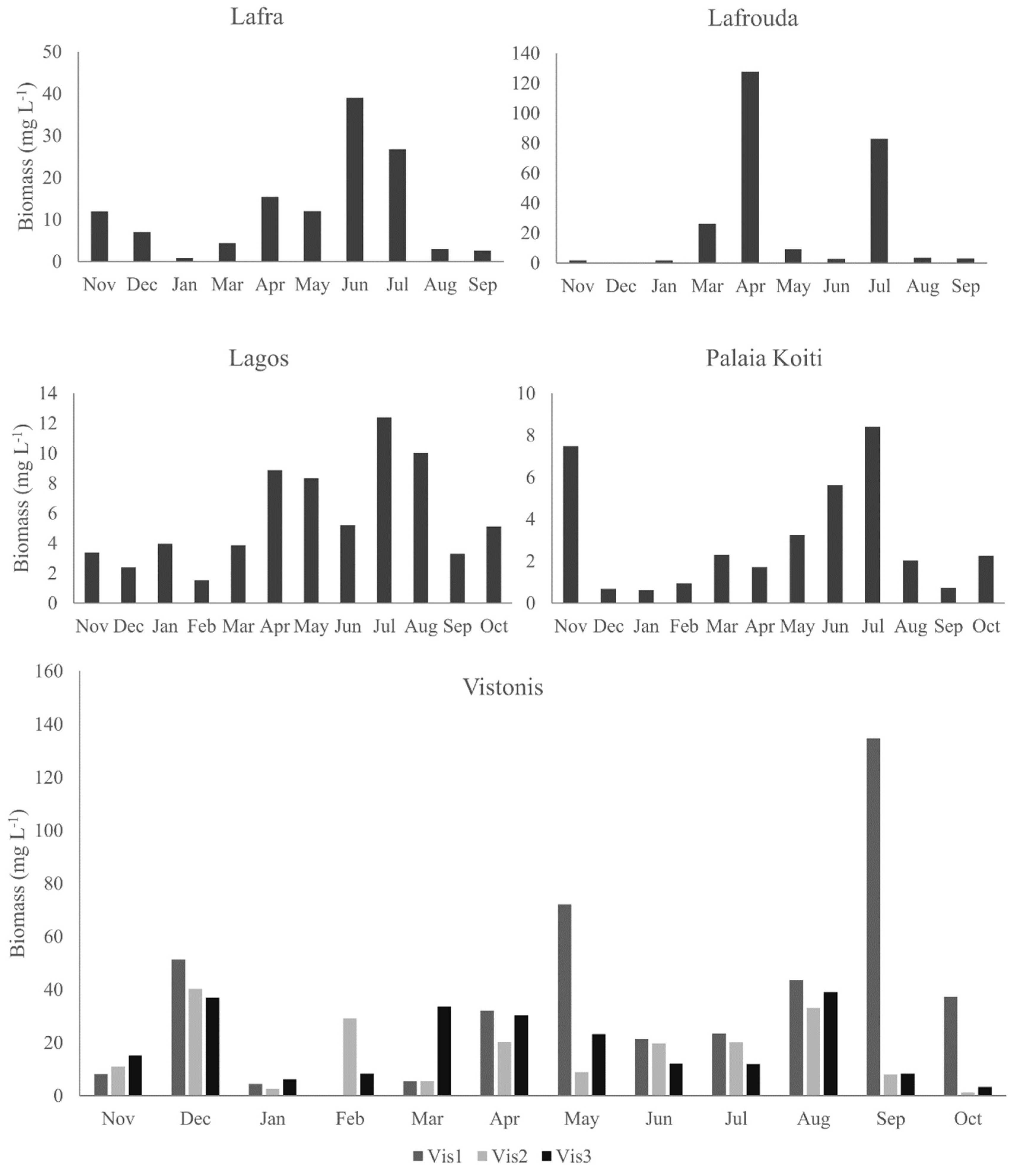
3.1. Phytoplankton Community Structure and Biomass
3.2. Phytoplankton Diversity
3.2.1. Alpha Diversity Estimators
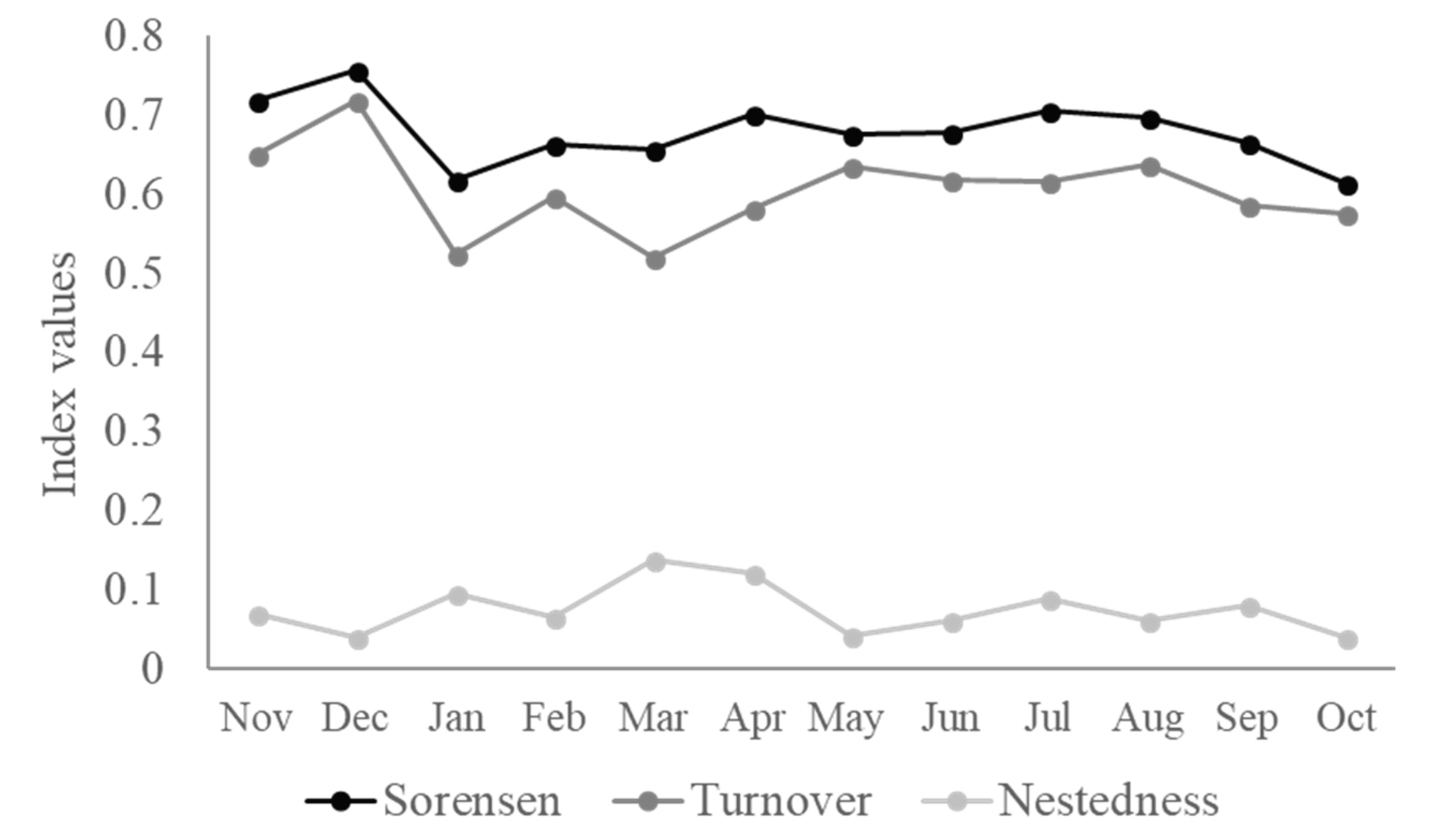
3.2.2. Beta Diversity
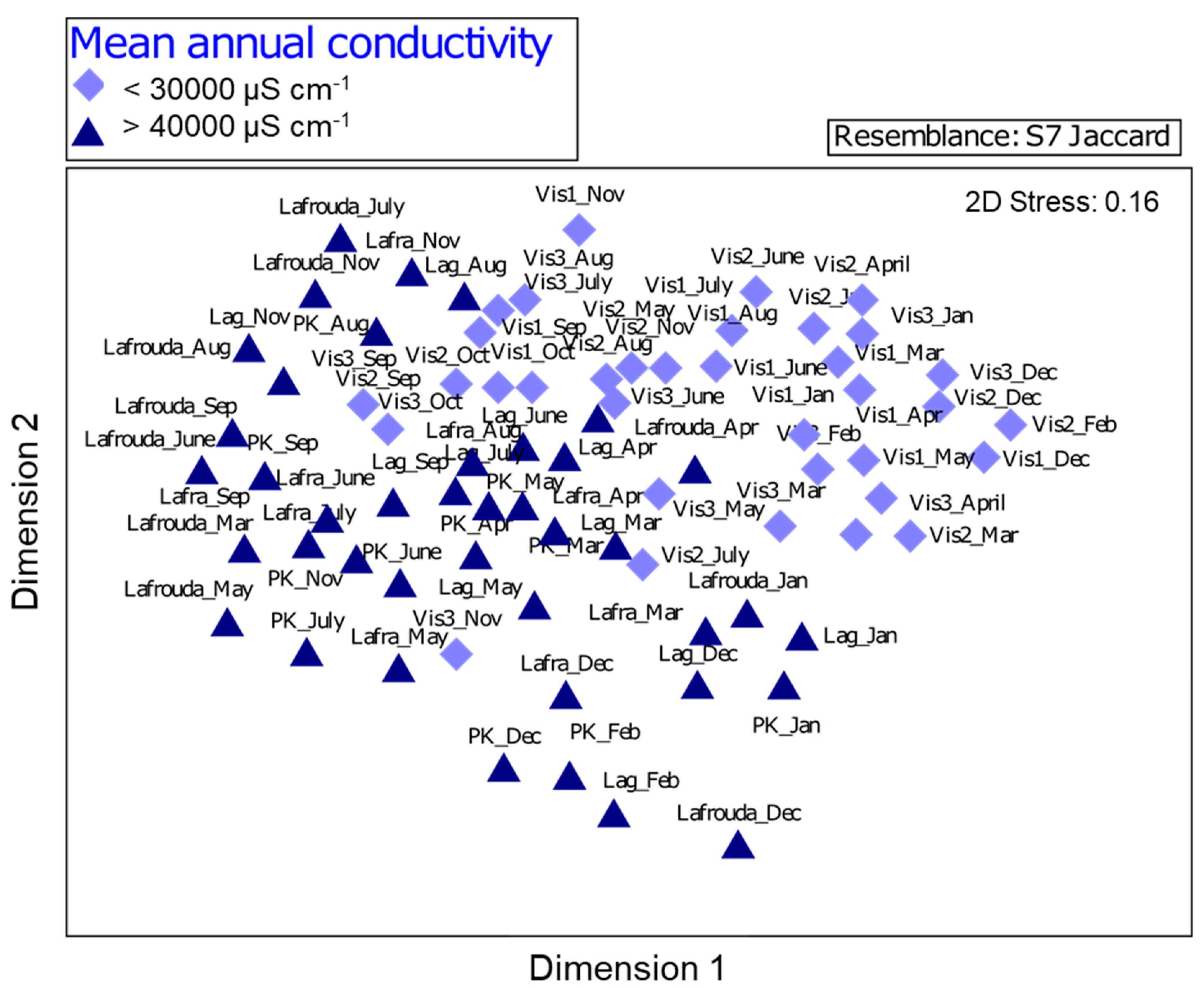
3.3. Phytoplankton Grouping
3.4. Non-linear Regression Models
4. Discussion
4.1. Phytoplankton Composition and Alpha Diversity
4.2. High Variation in Conductivity Promotes High Phytoplankton Beta Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kjerfve, B. Coastal Lagoon Processes; Elsevier Science Publishers: Amsterdam, The Netherland, 1994; p. 577. [Google Scholar]
- Knoppers, B. Aquatic primary production in coastal lagoons. In Coastal Lagoon Processes; Kjerfve, B., Ed.; Elsevier Science Publishers: Amsterdam, The Netherland, 1994; pp. 243–286. [Google Scholar]
- Levin, L.A.; Boesch, D.F.; Covich, A.; Dahm, C.; Erséus, C.; Ewel, K.C.; Kneib, R.T.; Moldenke, A.; Palmer, M.A.; Snelgrove, P.; et al. The function of marine critical transition zones and the importance of sediment biodiversity. Ecosystems 2000, 4, 430–451. [Google Scholar] [CrossRef]
- Mouillot, D.; Gaillard, S.; Aliaume, C.; Verlaque, M.; Belsher, T.; Troussellier, M.; Thang, D.C. Ability of taxonomic diversity indices to discriminate coastal lagoon environments based on macrophyte communities. Ecol. Indic. 2005, 5, 1–17. [Google Scholar] [CrossRef]
- Lloret, J.; Marín, A.; Marín-Guirao, L. Is coastal lagoon eutrophication likely to be aggravated by global climate change? Estuar. Coast. Shelf Sci. 2008, 78, 403–412. [Google Scholar] [CrossRef]
- De Casabianca, M.L.; Laugier, T.; Marinho-Soriano, E. Seasonal changes of nutrients in water and sediment in a Mediterranean lagoon with shellfish farming activity (Thau Lagoon, France). ICES Mar. Sci. 1997, 54, 905–916. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Fernández, A.I.; Concepción, M.; Gilabert, J.; Quispe, I.J.; García-Charton, J.A. Spatial and temporal variations of hydrological conditions, nutrients and chlorophyll a in a Mediterranean coastal lagoon (Mar Menor, Spain). Hydrobiologia 2005, 550, 11–27. [Google Scholar] [CrossRef]
- Grigorakis, K.; Rigos, G. Aquaculture effects on environmental and public welfare—The case of Mediterranean mariculture. Chemosphere 2011, 85, 899–919. [Google Scholar] [CrossRef]
- Eisenreich, S.J. (Ed.) Climate Change and the European Water Dimension; EU-Report 21553 of the European Commission; Joint Research Centre: Ispra, Italy, 2005; p. 253. [Google Scholar]
- Intergovernmental Panel on Climate Change [IPCC]. Climate Change 2014: Impacts, Adaptation and Vulnerability; IPCC Working Group II Contribution to the 5th Assessment Report of the International Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014; p. 169. [Google Scholar]
- Intergovernmental Panel on Climate Change [IPCC]. Summary for Policymakers. In Global Warming of 1.5 °C; An IPCC special report on the impacts of global warming of 1.5 °C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty; World Meteorological Organization: Geneva, Switzerland, 2018; p. 630. [Google Scholar]
- Stefanidou, N.; Genitsaris, S.; Lopez-Bautista, J.; Sommer, U.; Moustaka-Gouni, M. Effects of heat shock and salinity changes on coastal Mediterranean phytoplankton in a mesocosm experiment. Mar. Biol. 2018, 165, 154. [Google Scholar] [CrossRef]
- Stefanidou, N.; Genitsaris, S.; Lopez-Bautista, J.; Sommer, U.; Moustaka-Gouni, M. Unicellular eukaryotic community response to temperature and salinity variation in mesocosm experiments. Front. Microbiol. 2018, 9, 2444. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M. Mediterranean coastal lagoons in an ecosystem and aquatic resources management context. Phys. Chem. Earth Parts A B C 2011, 36, 160–166. [Google Scholar] [CrossRef]
- Blanchet, M.; Pringault, O.; Bouvy, M.; Catala, P.; Oriol, L.; Caparros, J.; Ortega-Retuerta, E.; Intertaglia, L.; West, N.; Agis, M.; et al. Changes in bacterial community metabolism and composition during the degradation of dissolved organic matter from the jellyfish Aurelia aurita in a Mediterranean coastal lagoon. Environ. Sci. Pollut. Res. 2015, 22, 13638. [Google Scholar] [CrossRef]
- Ghai, R.; Hernandez, C.M.; Picazo, A.; Mizuno, C.M.; Ininbergs, K.; Diez, B.; Valas, R.; DuPont, C.L.; McMahon, K.D.; Camacho, A.; et al. Metagenomes of Mediterranean coastal lagoons. Sci. Rep. 2012, 2, 490. [Google Scholar] [CrossRef] [PubMed]
- Riera, R.; Tuset, V.M.; Betancur-R, R.; Lombarte, A.; Marcos, C.; Pérez-Ruzafa, A. Modelling alpha-diversities of coastal lagoon fish assemblages from the Mediterranean Sea. Prog. Oceanogr. 2018, 165, 100–109. [Google Scholar] [CrossRef]
- Bellino, A.; Mangano, M.C.; Baldantoni, D.; Russell, B.D.; Mannino, A.M.; Mazzola, A.; Vizzini, A.; Sarà, G. Seasonal patterns of biodiversity in Mediterranean coastal lagoons. Divers. Distrib. 2019, 25, 1512–1526. [Google Scholar] [CrossRef]
- Orfanidis, S.; Pinna, M.; Sabetta, L.; Stamatis, N.; Nakou, K. Variation of structural and functional metrics in macrophyte communities within two habitats of eastern Mediterranean coastal lagoons: Natural versus human effects. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, S45–S61. [Google Scholar] [CrossRef]
- Koutrakis, E.T.; Tsikliras, A.C.; Sinis, A.I. Temporal variability of the ichthyofauna in a Northern Aegean coastal lagoon (Greece). Influence of environmental factors. Hydrobiologia 2005, 543, 245. [Google Scholar] [CrossRef]
- Carlier, A.; Riera, P.; Amouroux, J.M.; Bodiou, J.Y.; Desmalades, M.; Grémare, A. Food web structure of two Mediterranean lagoons under varying degree of eutrophication. J. Sea Res. 2008, 60, 264–275. [Google Scholar] [CrossRef]
- Roselli, L.; Stanca, E.; Ludovisi, A.; Durante, G.; Souza, J.S.D.; Dural, M.; Alp, T.; Bulent, S.; Gjoni, V.; Ghinis, S.; et al. Multi-scale biodiverity patterns in phytoplankton from coastal lagoons: The Eastern Mediterranean. Transit. Waters Bull. 2013, 7, 202–219. [Google Scholar]
- Bazin, P.; Jouenne, F.; Deton-Cabanillas, A.F.; Pérez-Ruzafa, A.; Véron, B. Complex patterns in phytoplankton and microeukaryote diversity along the estuarine continuum. Hydrobiologia 2014, 726, 155. [Google Scholar] [CrossRef]
- Leruste, A.; Pasqualini, V.; Garrido, M.; Malet, N.; De Wit, R.; Bec, B. Physiological and behavioral responses of phytoplankton communities to nutrient availability in a disturbed Mediterranean coastal lagoon. Estuar. Coast. Shelf Sci. 2019, 219, 176–188. [Google Scholar] [CrossRef]
- Trombetta, T.; Vidussi, F.; Mas, S.; Parin, D.; Simier, M.; Mostajir, B. Water temperature drives phytoplankton blooms in coastal waters. PLoS ONE 2019, 14, e0214933. [Google Scholar] [CrossRef]
- Pavloudi, C.; Oulas, A.; Vasileiadou, K.; Sarropoulou, E.; Kotoulas, G.; Arvanitidis, C. Salinity is the major factor influencing the sediment bacterial communities in a Mediterranean lagoonal complex (Amvrakikos Gulf, Ionian Sea). Mar. Genom. 2016, 28, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Grzebyk, D.; Audic, S.; Lasserre, B.; Abadie, E.; de Vargas, C.; Bec, B. Insights into the harmful algal flora in northwestern Mediterranean coastal lagoons revealed by pyrosequencing metabarcodes of the 28S rRNA gene. Harmful Algae 2017, 68, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Aires, T.; Muyzer, G.; Serrão, E.A.; Engelen, A.H. Seaweed Loads Cause Stronger Bacterial Community Shifts in Coastal Lagoon Sediments Than Nutrient Loads. Front. Microbiol. 2019, 9, 3283. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 28, 453–470. [Google Scholar] [CrossRef]
- Koleff, P.; Gaston, K.J.; Lennon, J.J. Measuring beta diversity for presence–absence data. J. Anim. Ecol. 2003, 72, 367–382. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. Betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Papadakis, E.M.; Tsaboula, A.; Kotopoulou, A.; Kintzikoglou, K.; Vryzas, Z.; Papadopoulou-Mourkidou, E. Pesticides in the surface waters of Lake Vistonis Basin, Greece: Occurrence and environmental risk assessment. Sci. Total Environ. 2015, 536, 793–802. [Google Scholar] [CrossRef]
- Arabatzis, G.D.; Kokkinakis, A.K. Typology of the lagoons of Northern Greece according to their environmental characteristics and fisheries production. Oper. Res. Int. J. 2005, 5, 21–34. [Google Scholar] [CrossRef]
- Koutrakis, E. Northern Greece Lagoon Management, Suggestions and Problems; Institute of Plant Breeding and Genetic Resources, Hellenic Agricultural Organization-Inale: Kavala, Greece, 2000; p. 50. [Google Scholar]
- Weyl, P.K. On the change in electrical conductance of seawater with temperature. Limnol. Oceanogr. 1964, 9, 75–78. [Google Scholar] [CrossRef]
- Mazaris, A.D.; Moustaka-Gouni, M.; Michaloudi, E.; Bobori, D.C. Biogeographical patterns of freshwater micro- and macroorganisms: A comparison between phytoplankton, zooplankton and fish in the eastern Mediterranean. J. Biogeogr. 2010, 37, 1341–1351. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.-D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Menden-Deuer, S.; Lessard, E.J. Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton. Limnol. Oceanogr. 2000, 45, 569–579. [Google Scholar] [CrossRef]
- Sommer, U.; Lengfellner, K.; Lewandowska, A. Experimental induction of a coastal spring bloom early in the year by intermittent high-light episodes. Mar. Ecol. Prog. Ser. 2012, 446, 61–71. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9–10. [Google Scholar]
- Baselga, A. Multiplicative partition of true diversity yields independent alpha and beta components; additive partition does not. Ecology 2010, 91, 1974–1981. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 20 May 2019).
- Benoit, K. Linear Regression Models with Logarithmic Transformations; London Methodology Institute, London School of Economics: London, UK, 2011; p. 25. [Google Scholar]
- Hemraj, D.A.; Hossain, M.A.; Ye, Q.; Qin, J.G.; Leterme, S.C. Plankton bioindicators of environmental conditions in coastal lagoons. Estuar. Coast. Shelf Sci. 2017, 184, 102–114. [Google Scholar] [CrossRef]
- Yang, J.; Ma, L.A.; Jiang, H.; Wu, G.; Dong, H. Salinity shapes microbial diversity and community structure in surface sediments of the Qinghai-Tibetan Lakes. Sci. Rep. 2016, 6, 25078. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; De Pascalis, F.; Ghezzo, M.; Quispe-Becerra, J.L.; Hernández-García, R.; Muñoz, I.; Vergara, C.; Pérez-Ruzafa, I.M.; Umgiesser, G.; Marcos, C. Connectivity between coastal lagoons and sea: Asymmetrical effects on assemblages’ and populations’ structure. Estuar. Coast. Shelf Sci. 2019, 216, 171–186. [Google Scholar] [CrossRef]
- Carstensen, J.; Klais, R.; Cloern, J.E. Phytoplankton blooms in estuarine and coastal waters: Seasonal patterns and key species. Estuar. Coast. Shelf Sci. 2015, 162, 98–109. [Google Scholar] [CrossRef]
- Bussard, A.; Corre, E.; Hubas, C.; Duvernois-Berthet, E.; Le Corguillé, G.; Jourdren, L.; Coulpier, F.; Claquin, P.; Lopez, P.J. Physiological adjustments and transcriptome reprogramming are involved in the acclimation to salinity gradients in diatoms. Environ. Microbiol. 2017, 19, 909–925. [Google Scholar] [CrossRef] [PubMed]
- Michaloudi, E.; Moustaka-Gouni, M.; Gkelis, S.; Pantelidakis, K. Plankton community structure during an ecosystem disruptive algal bloom of Prymnesium parvum. J. Plankton Res. 2009, 31, 301–309. [Google Scholar] [CrossRef]
- Adolf, J.E.; Burns, J.; Walker, J.K.; Gamiao, S. Near shore distributions of phytoplankton and bacteria in relation to submarine groundwater discharge-fed fishponds, Kona coast, Hawaii, USA. Estuar. Coast. Shelf Sci. 2019, 219, 341–353. [Google Scholar] [CrossRef]
- Draredja, M.A.; Frihi, H.; Boualleg, C.; Gofart, A.; Abadie, E.; Laabir, M. Seasonal variations of phytoplankton community in relation to environmental factors in a protected meso-oligotrophic southern Mediterranean marine ecosystem (Mellah lagoon, Algeria) with an emphasis of HAB species. Environ. Monit. Assess. 2019, 191, 603. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.C.; Sheng, Y.; MacDonald, G.M.; Hinzman, L.D. Disappearing Arctic lakes. Science 2005, 308, 1429. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Best, R.J.; Bracken, M.E.S.; Graham, M.H. Complementarity in marine biodiversity manipulations: Reconciling divergent evidence from field and mesocosm experiments. Proc. Natl. Acad. Sci. USA 2008, 105, 18842–18847. [Google Scholar] [CrossRef]
- Lewandowska, A.M.; Breithaupt, P.; Hillebrand, H.; Hoppe, H.G.; Jürgens, K.; Sommer, U. Responses of primary productivity to increased temperature and phytoplankton diversity. J. Sea Res. 2012, 72, 87–93. [Google Scholar] [CrossRef]
- Török, P.; Krasznai, E.T.; Béres, V.B.; Bácsi, I.; Borics, B.G.; Tóthmérész, B. Functional diversity supports the biomass-diversity humped-back relationship in phytoplankton assemblages. Funct. Ecol. 2016, 30, 1593–1602. [Google Scholar] [CrossRef]
- Dodson, S.I.; Arnott, S.E.; Cottingham, K.L. The relationship in lake communities between primary productivity and species richness. Ecology 2000, 81, 2662–2679. [Google Scholar] [CrossRef]
- Heil, C.A. Influence of humic, fulvic and hydrophilic acids on the growth, photosynthesis and respiration of the dinoflagellate Prorocentrum minimum (Pavillard) Schiller. Harmful Algae 2005, 4, 603–618. [Google Scholar] [CrossRef]
- Anderson, D.M. Bloom dynamics of toxic Alexandrium species in the northeastern US. Limnol. Oceanogr. 1997, 42, 1009–1022. [Google Scholar] [CrossRef]
- Genitsaris, S.; Stefanidou, N.; Sommer, U.; Moustaka-Gouni, M. Phytoplankton Blooms, Red Tides and Mucilaginous Aggregates in the Urban Thessaloniki Bay, Eastern Mediterranean. Diversity 2019, 11, 136. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Stoll, S.; Jähnig, S.C.; Haase, P. Contrasting metacommunity structure and beta diversity in an aquatic-floodplain system. Oikos 2016, 125, 686–697. [Google Scholar] [CrossRef]
- Heino, J.; Soininen, J.; Alahuhta, J.; Lappalainen, J.; Virtanen, R. Metacommunity ecology meets biogeography: Effects of geographical region, spatial dynamics and environmental filtering on community structure in aquatic organisms. Oecologia 2017, 183, 121. [Google Scholar] [CrossRef]
- Maloufi, S.; Catherine, A.; Mouillot, D.; Louvard, C.; Couté, A.; Bernard, C.; Troussellier, M. Environmental heterogeneity among lakes promotes hyper β-diversity across phytoplankton communities. Freshw. Biol. 2016, 61, 633–645. [Google Scholar] [CrossRef]
- Chaparro, G.; Horváth, Z.; O’Farrell, I.; Ptacnik, R.; Hein, T. Plankton metacommunities in floodplain wetlands under contrasting hydrological conditions. Freshw. Biol. 2018, 63, 380–391. [Google Scholar] [CrossRef]
- Zhang, Y.; Peng, C.; Huang, S.; Wang, J.; Xiong, X.; Li, D. The relative role of spatial and environmental processes on seasonal variations of phytoplankton beta diversity along different anthropogenic disturbances of subtropical rivers in China. Environ. Sci. Pollut. Res. 2019, 26, 1422. [Google Scholar] [CrossRef]
- Bozelli, R.L.; Thomaz, S.M.; Padial, A.A.; Lopes, P.M.; Bini, L.M. Floods decrease zooplankton beta diversity and environmental heterogeneity in an Amazonian floodplain system. Hydrobiologia 2015, 753, 233–241. [Google Scholar] [CrossRef]
- Gao, Y.; Sassenhagen, I.; Richlen, M.L.; Donald, M.; Anderson, D.M.; Martin, J.L.; Erdner, D.L. Spatiotemporal genetic structure of regional-scale Alexandrium catenella dinoflagellate blooms explained by extensive dispersal and environmental selection. Harmful Algae 2019, 86, 46–54. [Google Scholar] [CrossRef]
- Katsiapi, M.; Genitsaris, S.; Stefanidou, N.; Tsavdaridou, A.; Giannopoulou, I.; Stamou, G.; Michaloudi, E.; Mazaris, A.; Moustaka-Gouni, M. Ecological connectivity in two ancient lakes: Impact upon planktonic cyanobacteria and water quality. Water 2020, 12, 18. [Google Scholar] [CrossRef]
- Soininen, J.; Heino, J.; Wang, J. A meta-analysis of nestedness and turnover components of beta diversity across organisms and ecosystems. Glob. Ecol. Biogeogr. 2018, 27, 96–109. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Sommer, U.; Economou-Amilli, A.; Arhonditsis, G.B.; Katsiapi, M.; Papastergiadou, E.; Kormas, A.K.; Vardaka, E.; Karayanni, H.; Papadimitriou, T. Implementation of the Water Framework Directive: Lessons Learned and Future Perspectives for an Ecologically Meaningful Classification Based on Phytoplankton of the Status of Greek Lakes, Mediterranean Region. Environ. Manag. 2019, 64, 675. [Google Scholar] [CrossRef] [PubMed]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Tunney, T.D.; McCann, K.S.; Lester, N.P.; Brian, J.; Shuter, B.J. Differential warming affects complex communities. Proc. Natl. Acad. Sci. USA 2014, 22, 8077–8082. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Lagoon | Latitude | Longitude | Mean Annual Conductivity (μS cm−1) | Mean Annual Salinity (psu) |
|---|---|---|---|---|---|
| Lafra | Lafra | 40°59′32.08″ N | 25° 2′29.49″ E | 4,3334.9 | 28.2 |
| Lafrouda | Lafrouda | 40°59′11.98″ N | 25° 1′51.57″ E | 4,7201.4 | 31.1 |
| Lag | Lagos | 40°59′36.13″ N | 25° 8′43.93″ E | 4,9274.1 | 29.8 |
| PK | Palaia Koiti | 40°54′23.90″ N | 24°52′24.32″ E | 4,9468.2 | 30.7 |
| Vis1 | Vistonis | 41°0′3.58″ N | 25° 9′20.89″ E | 1,1664.6 | 7.3 |
| Vis2 | Vistonis | 41°0′27.24″ N | 25° 8′15.77″ E | 2,4811.4 | 13.9 |
| Vis3 | Vistonis | 41°0′19.28″ N | 25° 8′16.10″ E | 3,0070.9 | 18.2 |
| Lafra | Lafrouda | Lagos | Palaia Koiti | Vis1 | Vis2 | Vis3 | |
|---|---|---|---|---|---|---|---|
| Cyanobacteria | 5 | 5 | 7 | 4 | 8 | 7 | 8 |
| Bacillariophyceae | 18 | 21 | 16 | 28 | 13 | 18 | 20 |
| Dinophyceae | 12 | 10 | 15 | 13 | 9 | 8 | 12 |
| Chlorophyceae | 4 | 2 | 4 | 3 | 8 | 5 | 7 |
| Others | 9 | 8 | 10 | 9 | 8 | 9 | 8 |
| Response Variable | Predictor | Multiple R2 | F-Statistic | dF | p-Value |
|---|---|---|---|---|---|
| Phytoplankton abundance | Conductivity | 0.400 | 22.80 | 67 | 3.810−8 |
| Phytoplankton biomass | Conductivity | 0.208 | 8.80 | 67 | 0.0004 |
| Phytoplankton C | Conductivity | 0.269 | 12.30 | 67 | 2.764−5 |
| Phytoplankton species richness | Conductivity | 0.187 | 7.70 | 67 | 0.943−8 |
| Shannon | Conductivity | 0.039 | 1.35 | 67 | 0.270 |
| Evenness | Conductivity | 0.100 | 3.60 | 67 | 0.053 |
| Pair-wise bSOR | Difference in conductivity | 0.100 | 73.69 | 2412 | 2.210−16 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefanidou, N.; Katsiapi, M.; Tsianis, D.; Demertzioglou, M.; Michaloudi, E.; Moustaka-Gouni, M. Patterns in Alpha and Beta Phytoplankton Diversity along a Conductivity Gradient in Coastal Mediterranean Lagoons. Diversity 2020, 12, 38. https://doi.org/10.3390/d12010038
Stefanidou N, Katsiapi M, Tsianis D, Demertzioglou M, Michaloudi E, Moustaka-Gouni M. Patterns in Alpha and Beta Phytoplankton Diversity along a Conductivity Gradient in Coastal Mediterranean Lagoons. Diversity. 2020; 12(1):38. https://doi.org/10.3390/d12010038
Chicago/Turabian StyleStefanidou, Natassa, Matina Katsiapi, Dimitris Tsianis, Maria Demertzioglou, Evangelia Michaloudi, and Maria Moustaka-Gouni. 2020. "Patterns in Alpha and Beta Phytoplankton Diversity along a Conductivity Gradient in Coastal Mediterranean Lagoons" Diversity 12, no. 1: 38. https://doi.org/10.3390/d12010038
APA StyleStefanidou, N., Katsiapi, M., Tsianis, D., Demertzioglou, M., Michaloudi, E., & Moustaka-Gouni, M. (2020). Patterns in Alpha and Beta Phytoplankton Diversity along a Conductivity Gradient in Coastal Mediterranean Lagoons. Diversity, 12(1), 38. https://doi.org/10.3390/d12010038