Afromontane Forest Diversity and the Role of Grassland-Forest Transition in Tree Species Distribution



Abstract
1. Introduction
2. Materials and Methods
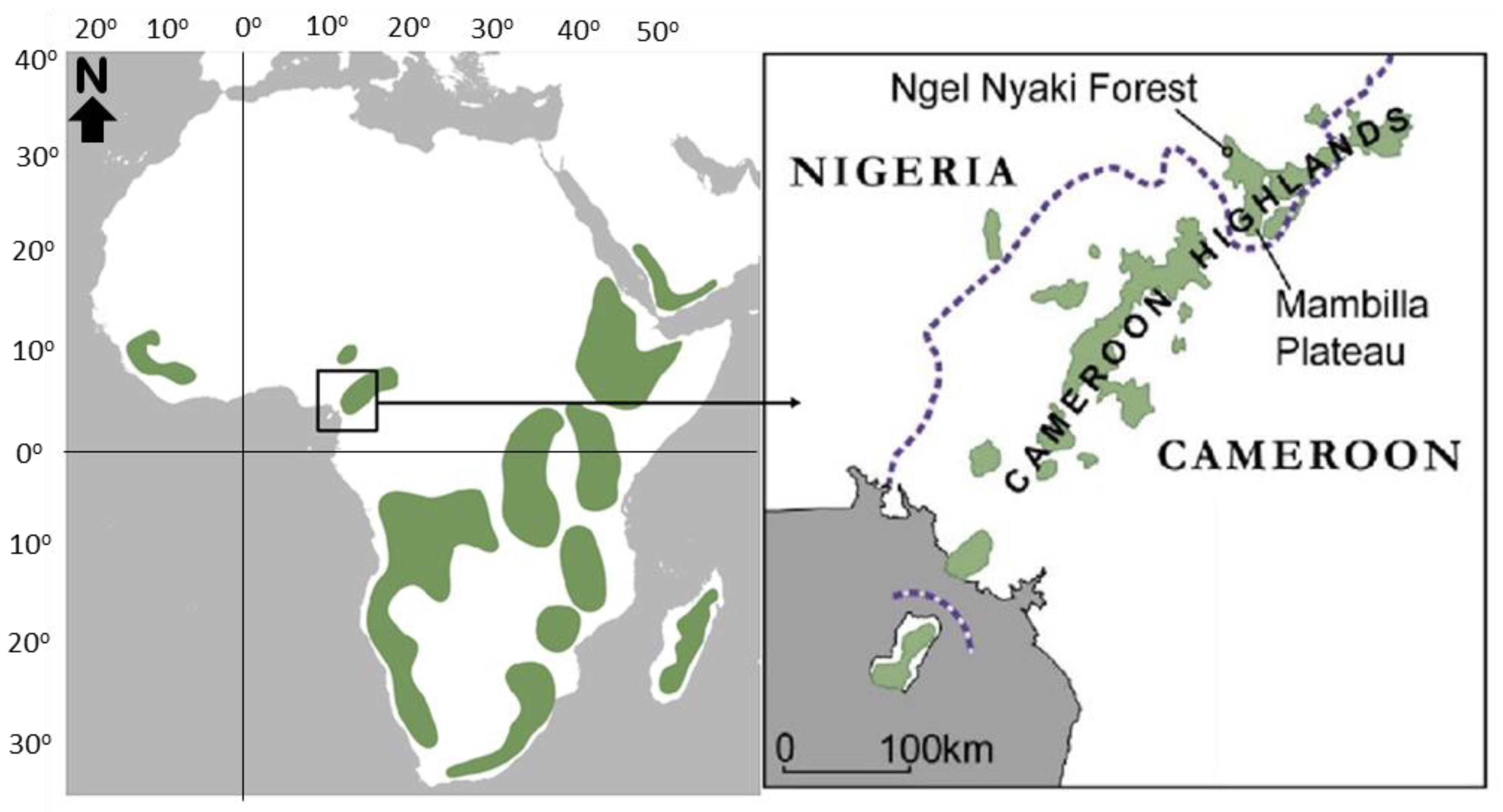
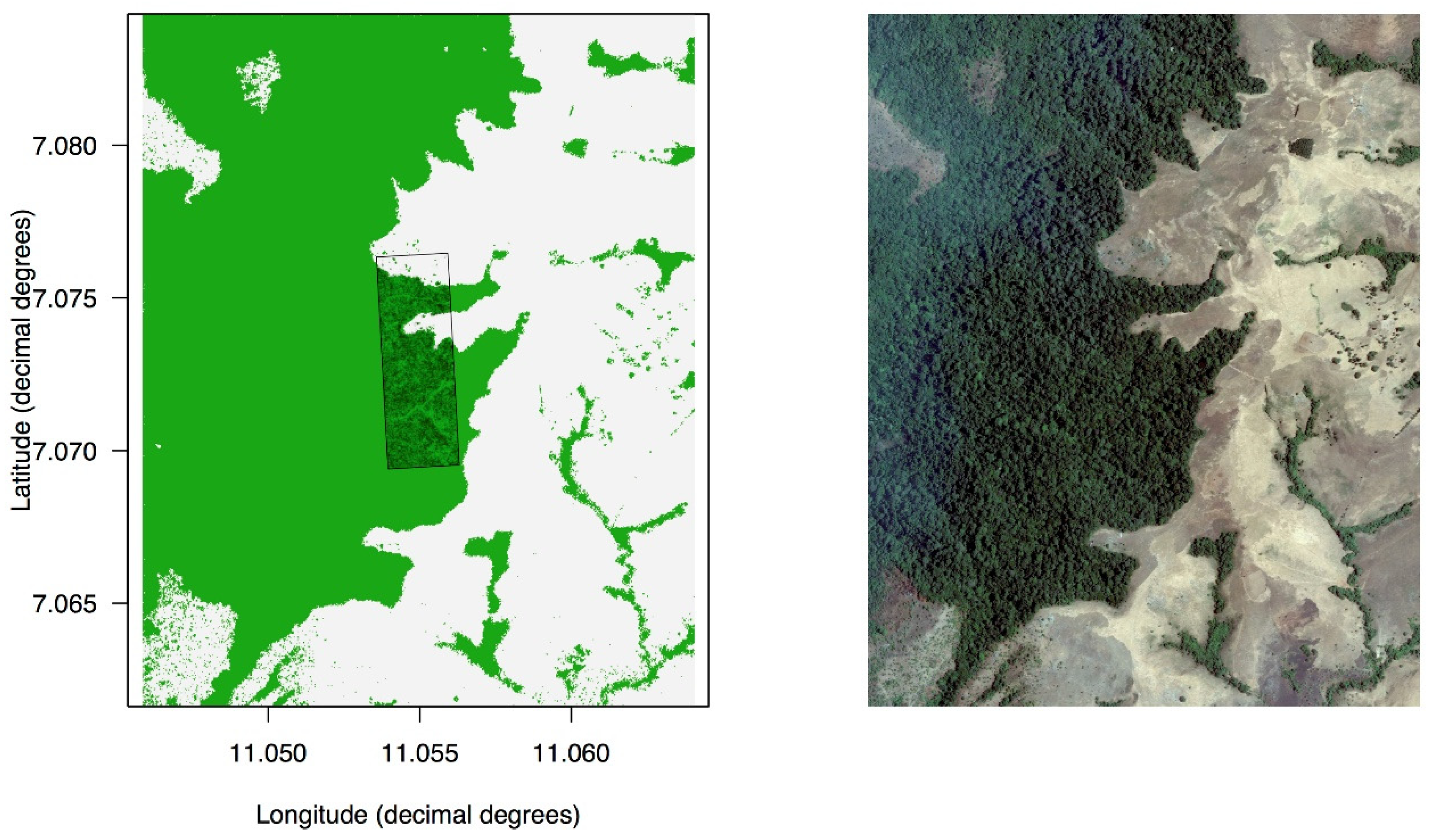
2.1. Study Area and Study Site
2.2. Census
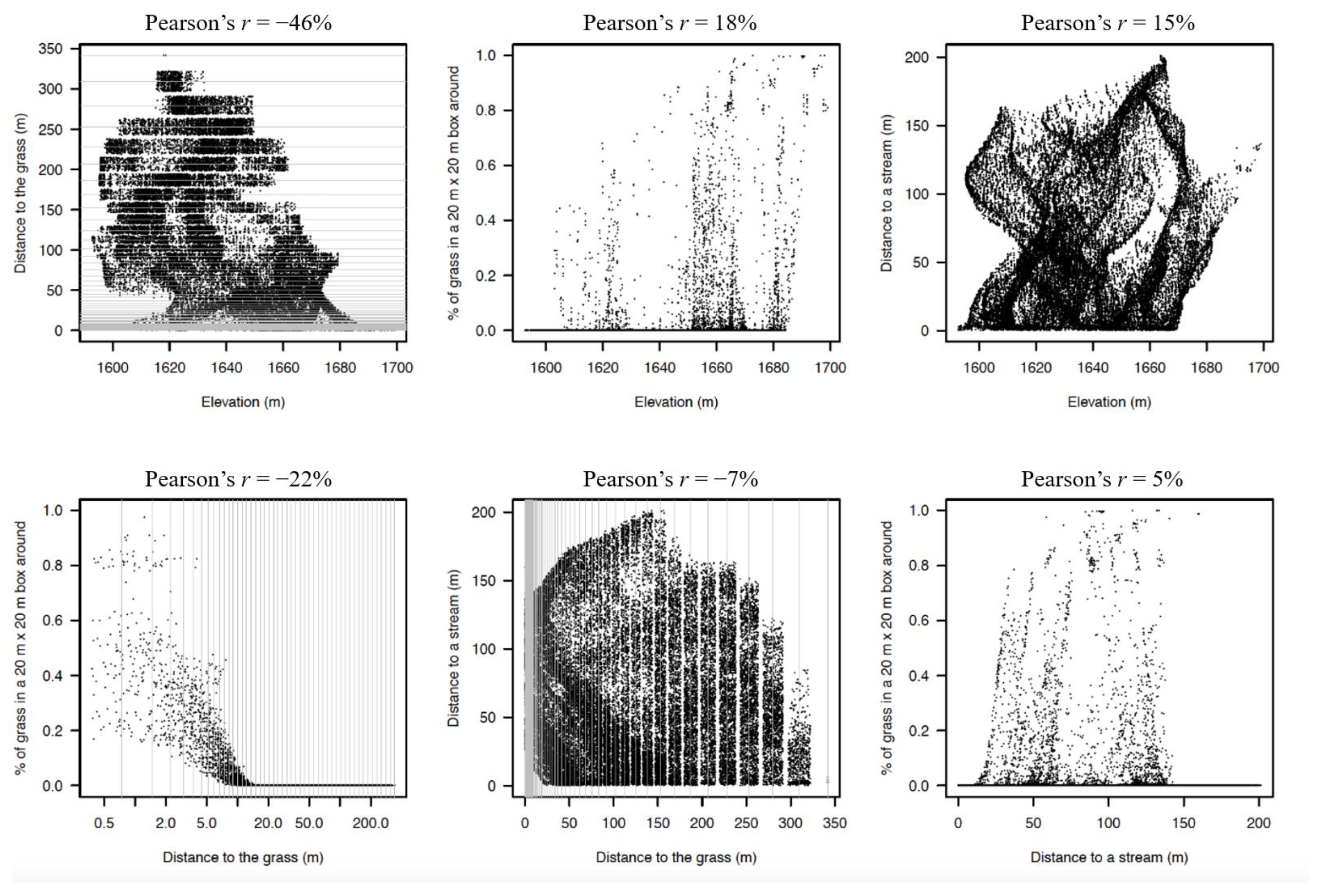
2.3. Environmental Variables and Gradients Evaluated
- (1)
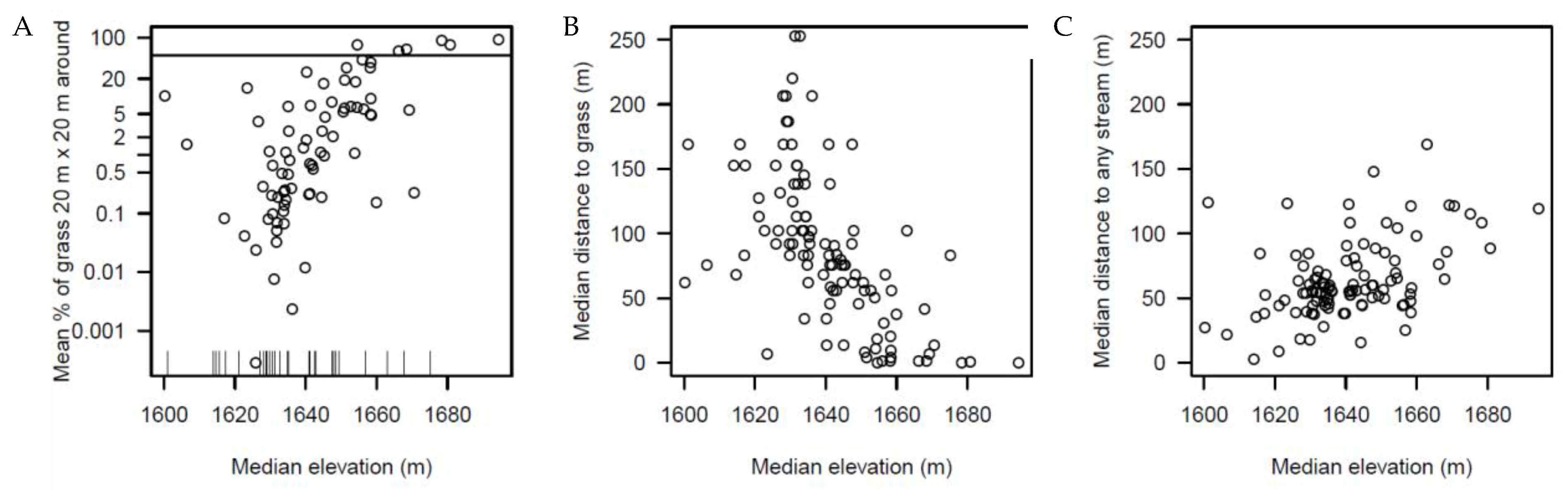
- The proportion of grass in a 20 × 20 m area surrounding the individual tree. Rather than count the grass vs. forest pixels in a 20 m diameter circle around each tree, we used a 20 × 20 m square window around each tree because it was computationally more efficient than defining a circle around each tree. Within this square we counted the number of red (grass) vs. other 1 × 1 m pixels and from this calculated the proportion of “grass” vs. forest pixels. We used this metric to indicate whether a species was a forest or grassland species. Our 20 × 20 m squares overlapped and were not necessarily independent of each other.
- (2)
- The distance to the closest grass pixel. This was calculated from the coordinates of each individual and the grass vs. forest binary map. This metric indicates if a species prefers the forest core (=far from grass) or the forest edge or the grassland itself (=close to the grass).
- (3)
- The shortest distance to any of two streams in the plot.
- (4)
- The local elevation.
2.4. Species Guilds
2.5. Data Analyses
3. Results
3.1. Diversity and Abundance
3.2. Basal Area
3.3. Species Distribution and Preferences
4. Discussion
4.1. Diversity and Composition of the Woody Plant Species
4.2. The Role of Environmental Variables
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
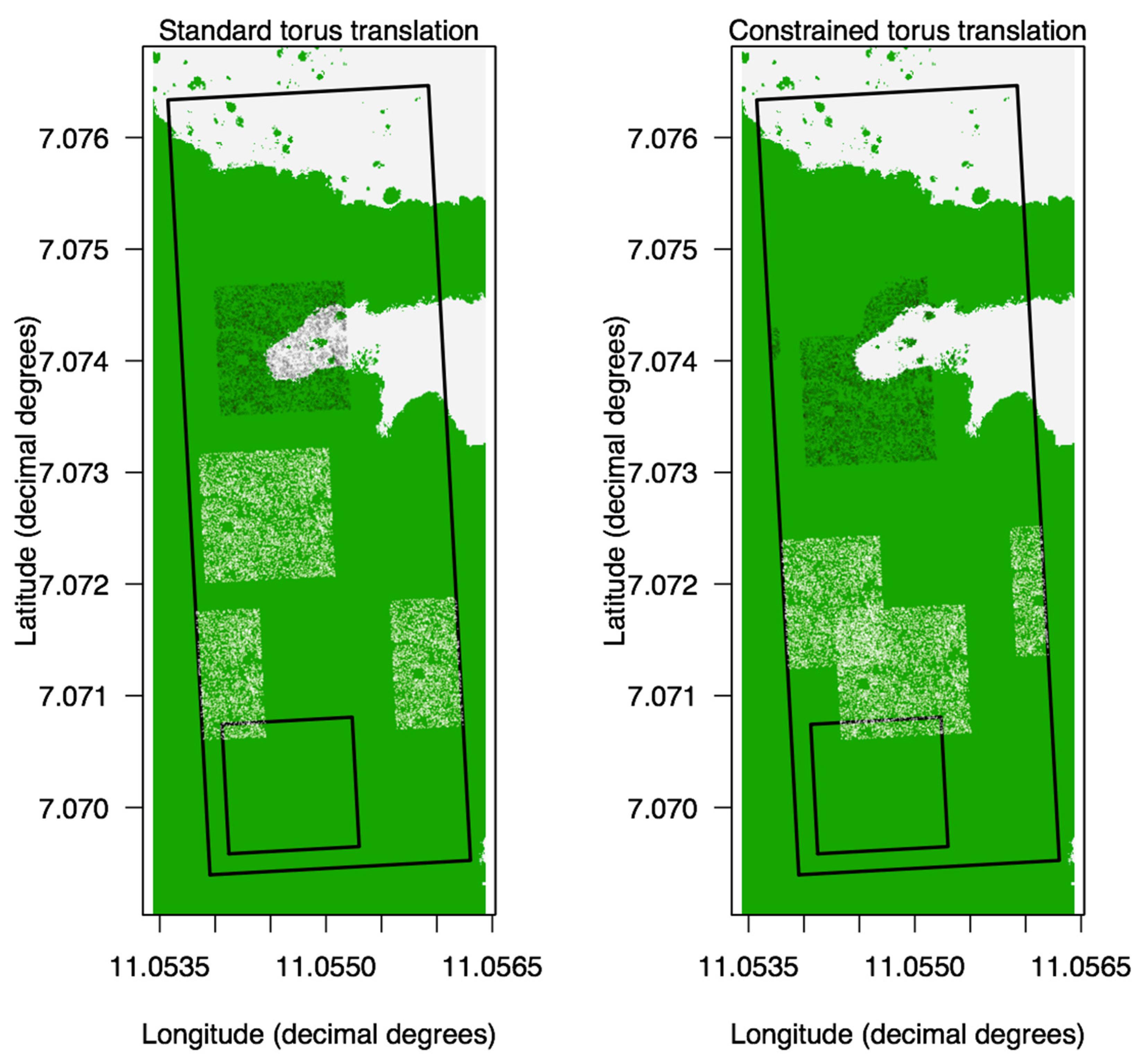
Appendix A.1. Torus Translations
Appendix A.2. Translating the Map vs. the Individuals
Appendix A.3. Constrained Translations
References
- Linder, H.P. The evolution of African plant diversity. Front. Ecol. Evol. 2014, 2, 38. [Google Scholar] [CrossRef]
- White, F. The Vegetation of Africa, a Descriptive Memoir to Accompany the UNESCO/AETFAT/UNSO Vegetation Map of Africa; UNESCO: Paris, France, 1983; 356p. [Google Scholar]
- White, F. The afromontane region. In Biogeography and Ecology of Southern Africa; Springer: Berlin/Heidelberg, Germany, 1978; pp. 463–513. [Google Scholar]
- Gehrke, B.; Linder, H.P. Species richness, endemism and species composition in the tropical Afroalpine flora. Alp. Bot. 2014, 124, 165–177. [Google Scholar] [CrossRef]
- Linder, H.P.; Lovett, J.; Mutke, J.M.; Barthlott, W.; Jürgens, N.; Rebelo, T.; Küper, W. A numerical re-evaluation of the sub-Saharan phytochoria of mainland Africa. Biol. Skr. 2005, 55, 229–252. [Google Scholar]
- Burgess, N.D.; Balmford, A.; Cordeiro, N.J.; Fjeldsa, J.; Küper, W.; Rahbek, C.; Sanderson, E.W.; Scharlemann, J.P.; Sommer, J.H.; Williams, P.H. Correlations among species distributions, human density and human infrastructure across the high biodiversity tropical mountains of Africa. J. Biol. Conserv. 2007, 134, 164–177. [Google Scholar] [CrossRef]
- Cordeiro, N.J.; Burgess, N.D.; Dovie, D.B.; Kaplin, B.A.; Plumptre, A.J.; Marrs, R. Conservation in areas of high population density in sub-Saharan Africa. Biol. Conserv. 2007, 134, 155–163. [Google Scholar] [CrossRef]
- Meadows, M.; Linder, H. Special Paper: A Palaeoecological perspective on the origin of afromontane grasslands. J. Biogeogr. 1993, 20, 345–355. [Google Scholar] [CrossRef]
- Spracklen, D.; Righelato, R. Tropical montane forests are a larger than expected global carbon store. Biogeosciences 2014, 11, 2741–2754. [Google Scholar] [CrossRef]
- Schröter, D.; Cramer, W.; Leemans, R.; Prentice, I.C.; Araújo, M.B.; Arnell, N.W.; Bondeau, A.; Bugmann, H.; Carter, T.R.; Gracia, C.A. Ecosystem service supply and vulnerability to global change in Europe. Science 2005, 310, 1333–1337. [Google Scholar] [CrossRef]
- Nadkarni, N.M.; Wheelwright, N.T. Monteverde: Ecology and Conservation of a Tropical Cloud Forest; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Chapman, H.M.; Olson, S.M.; Trumm, D. An assessment of changes in the montane forests of Taraba State, Nigeria, over the past 30 years. Oryx 2004, 38, 282–290. [Google Scholar] [CrossRef]
- Lézine, A.-M.; Izumi, K.; Kageyama, M.; Achoundong, G. A 90,000-year record of Afromontane forest responses to climate change. Science 2019, 363, 177–181. [Google Scholar] [CrossRef]
- Lawes, M.J. The Distribution of the Samango Monkey (Cercopithecus mitis erythrarchus Peters, 1852 and Cercopithecus mitis labiatus I. Geoffroy, 1843) and Forest History in Southern Africa. J. Biogeogr. 1990, 17, 669–680. [Google Scholar] [CrossRef]
- White, G.C.; Burnham, K.P.; Anderson, D.R. Advanced features of Program Mark. In Wildlife, Land, and People: Priorities for the 21st Century; Field, R., Warren, R.J., Okarma, H., Sievert, P.R., Eds.; The Wildlife Society: Bethesda, MD, USA, 2001. [Google Scholar]
- Lézine, A.-M.; Assi-Kaudjhis, C.; Roche, E.; Vincens, A.; Achoundong, G. Towards an understanding of West African montane forest response to climate change. J. Biogeogr. 2013, 40, 183–196. [Google Scholar] [CrossRef]
- Eeley, H.A.; Lawes, M.J.; Piper, S. The influence of climate change on the distribution of indigenous forest in KwaZulu-Natal, South Africa. J. Biogeogr. 1999, 26, 595–617. [Google Scholar] [CrossRef]
- Dowsett-Lemaire, F.; Dowsett, R.J.; Dyer, M. Important Bird Areas in Africa and Associated Islands; Pisces Publications and BirdLife International: Cambridge, UK, 2001. [Google Scholar]
- Lebamba. Forest-savannah dynamics on the Adamawa plateau (Central Cameroon) during the “African humid period” termination: A new high-resolution pollen record from Lake Tizong. Rev. Palaeobot. Palynol. 2016, 235. [Google Scholar] [CrossRef]
- Venter, Z.S.; Cramer, M.D.; Hawkins, H.J. Drivers of woody plant encroachment over Africa. Nat. Commun. 2018, 9, 2272. [Google Scholar] [CrossRef]
- Müller, S.C.; Overbeck, G.E.; Pfadenhauer, J.; Pillar, V.D. Woody species patterns at forest–grassland boundaries in southern Brazil. Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 586–598. [Google Scholar] [CrossRef]
- Gignoux, J.; Konaté, S.; Lahoreau, G.; Le Roux, X.; Simioni, G. Allocation strategies of savanna and forest tree seedlings in response to fire and shading: Outcomes of a field experiment. Sci. Rep. 2016, 6, 38838. [Google Scholar] [CrossRef]
- Kotze, D.J.; Lawes, M.J. Viability of ecological processes in small Afromontane forest patches in South Africa. J. Austral Ecol. 2007, 32, 294–304. [Google Scholar] [CrossRef]
- Andela, N.; Morton, D.; Giglio, L.; Chen, Y.; Van Der Werf, G.; Kasibhatla, P.; DeFries, R.; Collatz, G.; Hantson, S.; Kloster, S. A human-driven decline in global burned area. Science 2017, 356, 1356–1362. [Google Scholar] [CrossRef]
- Van Langevelde, F.; Van De Vijver, C.A.; Kumar, L.; Van De Koppel, J.; De Ridder, N.; Van Andel, J.; Skidmore, A.K.; Hearne, J.W.; Stroosnijder, L.; Bond, W.J. Effects of fire and herbivory on the stability of savanna ecosystems. Ecology 2003, 84, 337–350. [Google Scholar] [CrossRef]
- Adie, H.; Kotze, D.J.; Lawes, M.J. Small fire refugia in the grassy matrix and the persistence of Afrotemperate forest in the Drakensberg Mountains. Sci. Rep. 2017, 7, 6549. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.; Chapman, H. The Forests of Taraba and Adamawa States, Nigeria an Ecological Account and Plant Species Checklist; University of Canterbury: Christchurch, New Zealand, 2001; p. 221. [Google Scholar]
- Beck, J.; Chapman, H. A population estimate of the endangered chimpanzee Pan troglodytes vellerosus in a Nigerian montane forest: Implications for conservation. Oryx 2008, 42, 448. [Google Scholar] [CrossRef]
- Nigerian Montane Forest Project. Available online: https://www.canterbury.ac.nz/afromontane/ (accessed on 6 November 2019).
- Borokini, T. A Systematic Compilation of IUCN Red-listed Threatened Plant Species in Nigeria. Int. J. Environ. Sci. 2014, 3, 104–133. [Google Scholar]
- Thia, J.A. The Plight of Trees in Disturbed Forest: Conservation of Montane Trees, Nigeria. Master’s Thesis, University of Canterbury, Christchurch, New Zealand, 2014. [Google Scholar]
- Condit, R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and A Comparison with Other Plots; Springer Science & Business Media: Berlin, Germany, 1998. [Google Scholar]
- Barcode of Life Data System. Available online: https://www.boldsystems.org/ (accessed on 6 November 2019).
- The World’s Most Detailed Globe. Available online: https://www.google.com/earth/ (accessed on 6 November 2019).
- Hawthorne, W. Ecological profiles of Ghanaian forest trees. In Tropical Forestry Papers; Oxford Forestry Institute: Oxford, UK, 1995; Volume 29, pp. 1–345. [Google Scholar]
- Hawthorne, W.D. Holes and the sums of parts in Ghanaian forest: Regeneration, scale and sustainable use. Proc. R. Soc. Edinb. Sect. B Biol. Sci. 1996, 104, 75–176. [Google Scholar] [CrossRef]
- Global Plants. Available online: https://plants.jstor.org/ (accessed on 6 November 2019).
- Barnes, A.D.; Chapman, H.M. Dispersal traits determine passive restoration trajectory of a Nigerian montane forest. Acta Oecol. 2014, 56, 32–40. [Google Scholar] [CrossRef]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha Neotropical forest plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point-Pattern Analysis in Ecology; CRC: Boca Raton, FL, USA, 2013. [Google Scholar]
- Arellano, G.; Medina, N.G.; Tan, S.; Mohamad, M.; Davies, S.J. Crown damage and the mortality of tropical trees. New Phytol. 2019, 221, 169–179. [Google Scholar] [CrossRef]
- RCoreTeam. R: A Language and Environment for Statistical Computing; Version 3.3.3; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- The CTFS R Package. Available online: http://ctfs.si.edu/Public/CTFSRPackage/ (accessed on 6 November 2019).
- Hamilton, A.C.; Bensted-Smith, R. Forest Conservation in the East Usambara Mountains, Tanzania; IUCN: Dar es Salaam, Tanzania, 1989; Volume 15. [Google Scholar]
- Dowsett-Lemaire, F. The flora and phytogeography of the evergreen forests of Malawi I: Afromontane and mid-altitude forests. Bull. Jard. Bot. Natl. Belg./Bull. Natl. Plantentuin Belg. 1989, 59, 3–131. [Google Scholar] [CrossRef]
- Schmitt, C.B.; Denich, M.; Demissew, S.; Friis, I.; Boehmer, H.J. Floristic diversity in fragmented Afromontane rainforests: Altitudinal variation and conservation importance. Appl. Veg. Sci. 2010, 13, 291–304. [Google Scholar] [CrossRef]
- Kenfack, D.; Thomas, D.W.; Chuyong, G.B.; Condit, R. Rarity and abundance in a diverse African forest. Biodivers. Conserv. 2007, 16, 2045–2074. [Google Scholar] [CrossRef]
- Memiaghe, H.; Lutz, J.; Korte, L.; Alonso, A.; Kenfack, D. Ecological importance of small-diameter trees to the structure, diversirty and biomass of a Tropical Evergreen Forest at Rabi, Gabon. PLoS ONE 2016, 11, e0154988. [Google Scholar] [CrossRef] [PubMed]
- Makana, J.; Ewango, C.; McMahon, S.; Thomas, S.; Hart, T.; Condit, R. Demography and biomass change in monodominant and mixed old-growth forest of the Congo. J. Trop. Ecol. 2011, 27, 447–461. [Google Scholar] [CrossRef]
- Valencia, R.; Foster, R.B.; Villa, G.; Condit, R.; Svenning, J.C.; Hernandez, C.; Romoleroux, K.; Losos, E.; Magard, E.; Balslev, H. Tree species distributions and local habitat variation in the Amazon: Large forest plot in eastern Ecuador. J. Ecol. 2004, 92, 214–229. [Google Scholar] [CrossRef]
- Condit, R.; Ashton, P.; Baslev, H.; Brokaw, N.; Bunyavejchewin, S.; Chuyong, G.; Co, L.; Dattaraja, H.; Davies, S.; Esufali, S.; et al. Tropical tree alpha-diversity: Results from a worldwide network of large plots. Biol. Skr. 2005, 55, 565–582. [Google Scholar]
- Beavon, M.A.; Chapman, H.M. Andromonoecy and high fruit abortion in Anthonotha noldeae in a West African montane forest. Plant Syst. Evol. 2011, 296, 217–224. [Google Scholar] [CrossRef]
- Jackson, J.K. The vegetation of the Imatong Mountains, Sudan. J. Ecol. 1956, 341–374. [Google Scholar] [CrossRef]
- Grubb, P.J. Rainforest dynamics: The need for new paradigms. In Tropical Rainforest Research—Current Issues; Edwards, D.S., Booth, W.E., Choy, S.C., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 215–233. [Google Scholar]
- Diabate, M.; Munive, A.; De Faria, S.M.; Ba, A.; Dreyfus, B.; Galiana, A. Occurrence of nodulation in unexplored leguminous trees native to the West African tropical rainforest and inoculation response of native species useful in reforestation. New Phytol. 2005, 166, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Babale, A. The Interplay of Habitat and Seed Size on the Shift in Species Composition in a Fragmented Afromontane Forest Landscape: Implications for the Management of Forest Restoration. Ph.D. Thesis, University of Canterbury, Christchurch, New Zealand, 2014. [Google Scholar]
- Ndah, N.R.; Andrew, E.E.; Bechem, E. Species composition, diversity and distribution in a disturbed Takamanda Rainforest, South West, Cameroon. Afr. J. Plant Sci. 2013, 7, 577–585. [Google Scholar]
- Bässler, C.; Cadotte, M.W.; Beudert, B.; Heibl, C.; Blaschke, M.; Bradtka, J.H.; Langbehn, T.; Werth, S.; Müller, J. Contrasting patterns of lichen functional diversity and species richness across an elevation gradient. Ecography 2016, 39, 689–698. [Google Scholar] [CrossRef]
- Murphy, S.J.; Audino, L.D.; Whitacre, J.; Eck, J.L.; Wenzel, J.W.; Queenborough, S.A.; Comita, L.S. Species associations structured by environment and land-use history promote beta-diversity in a temperate forest. Ecology 2015, 96, 705–715. [Google Scholar] [CrossRef]
- Comita, L.S.; Engelbrecht, B.M. Seasonal and spatial variation in water availability drive habitat associations in a tropical forest. Ecology 2009, 90, 2755–2765. [Google Scholar] [CrossRef] [PubMed]
- Scalley, T.H.; Crowl, T.A.; Thompson, J. Tree species distributions in relation to stream distance in a mid-montane wet forest, Puerto Rico. Caribb. J. Sci. 2009, 45, 52–64. [Google Scholar] [CrossRef]
- Koffi, K.; Hardy, O.; Doumenge, C.; Cruaud, C.; Heuertz, M. Diversity gradients and phylogeographic patterns in Santiria trimera (Burseraceae), a widespread African tree typical of mature rainforests. Am. J. Bot. 2011, 98, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.D. The species-rich, paleotropical genus Memecylon (Melastomataceae): Molecular phylogenetics and revised infrageneric classification of the African species. Taxon 2014, 63, 539–561. [Google Scholar] [CrossRef]
- Brailsford, L.E. Evidence for Genetic Decline within Afromontane Forest Fragments on the Mambilla Plateau, Nigeria. Master’s Thesis, University of Canterbury, Christchurch, New Zealand, 2018. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | dbh ≥ 1 cm | dbh ≥ 10 cm | dbh ≥ 30 cm |
|---|---|---|---|
| Number of species | 105 | 88 (83%) | 61 (58%) |
| Number of genera | 88 | 76 (86%) | 56 (64%) |
| Number of families | 47 | 43 (92%) | 34 (72%) |
| Abundance | 41,031 | 6931 (17%) | 1686 (4%) |
| Total basal area (m2) | 553.49 | 498.24 (90.0%) | 356.60 (64%) |
| Fisher’s α per hectare | 12.42 ± 1.51 | 12.03 ± 2.20 | 9.55 ± 2.74 |
| Mean density per hectare (N/ha) | 2061.86 ± 697.03 | 351.79 ± 112.74 | 85.04 ± 28.00 |
| Mean basal area per hectare (m2/ha) | 27.52 ± 8.64 | 24.70 ± 7.87 | 17.62 ± 5.58 |
| Species | Family | Density (Trees/ha) | Basal Area (m2/ha) |
|---|---|---|---|
| Garcinia smeathmanii (Planch. & Triana) Oliv. | Clusiaceae | 607.63 | 1.20 (7) |
| Deinbollia pinnata Schumach. &Thonn. | Sapindaceae | 158.68 | 0.49 (11) |
| Pleiocarpa pycnantha (K.Schum.) Stapf | Apocynaceae | 107.51 | 0.48 (12) |
| Leptaulus zenkeri Engl. | Icacinaceae | 69.19 | 1.25 (6) |
| Carapa oreophila Kenfack | Meliaceae | 56.92 | 2.14 (3) |
| Chrysophyllum albidum G. Don | Sapotaceae | 56.75 | 0.21 (18) |
| Sorindeia sp. | Anacardiaceae | 53.10 | 0.25 (15) |
| Strombosia scheffleri Engl. | Olacaceae | 51.13 | 2.76 (2) |
| Drypetes gosweilleri S. Moore | Putranjivaceae | 50.42 | 0.63 (10) |
| Newtonia buchannani (Baker f.) G.C.C. Gilbert & Boutique | Fabaceae | 47.95 | 0.87 (8) |
| Dicranolepis grandiflora Engl. | Thymelaeceae | 47.54 | 0.04 (19) |
| Anthonotha noldeae (Rossberg) Exell and Hillc. | Fabaceae | 46.24 | 3.42 (1) |
| Voacanga africana Stapf | Apocynaceae | 44.52 | 0.22 (17) |
| Tabernaemontana contorta Stapf | Apocynaceae | 40.38 | 0.23 (16) |
| Santiria trimera (Oliv.) Aubrév. | Burseraceae | 40.36 | 1.76 (4) |
| Oxyanthus speciosus DC. | Rubiaceae | 39.21 | 0.47 (13) |
| Psychotria peduncularis (Salisb.) Steyerm. | Rubiaceae | 37.58 | 0.01 (20) |
| Macaranga occidentalis (Müll. Arg.) Müll. Arg. | Euphorbiaceae | 34.90 | 0.84 (9) |
| Trichilia monadelpha (Thonn.) J.J. de Wilde | Meliaceae | 28.18 | 1.40 (5) |
| Millettia conraui Harms | Fabaceae | 23.04 | 0.27 (14) |
| Species | % of Grass in 20 × 20 m Around | Distance to the Grass | Elevation | Distance to Streams | Guild | Growth Form | Seed Size (g) | Dispersal Mode | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Prop. of obs. > Null | Association | Prop. of obs. > Null | Association | Prop. of obs. > Null | Association | Prop. of obs. > Null | Association | |||||
| Leptaulus zenkeri | 0 | forest | 1 | core | 0.159 | NA | 0.47 | NA | S | CT | 0.12 | animal |
| Newtonia buchanannii | 0 | forest | 1 | core | 0.272 | NA | 0.157 | NA | S | ET | 0.14 | wind |
| Chrysophylum albidum | 0 | forest | 0.998 | core | 0.143 | NA | 0.818 | NA | S | CT | 0.35 | animal |
| Voacanga africana | 0 | forest | 0.992 | core | 0.154 | NA | 0.518 | NA | S | UT | 0.24 | animal |
| Dicranolepis grandifolia | 0 | forest | 0.992 | core | 0.157 | NA | 0.51 | NA | S | UT | 0.38 | animal |
| Chionanthus africanus | 0 | forest | 0.989 | core | 0.144 | NA | 0.988 | dry | S | CT | - | animal |
| Drypetes gossweileri | 0 | forest | 0.978 | core | 0.133 | NA | 0.028 | wet | S | CT | 0.01 | animal |
| Oxyanthus speciosus | 0 | forest | 0.975 | core | 0.326 | NA | 0.726 | NA | S | UT | 1.60 | animal |
| Pavetta corymbosa | 0 | forest | 0.974 | core | 0.479 | NA | 0.495 | NA | S | USh | 0.02 | animal |
| Discoclaoxylon hexandrum | 0 | forest | 0.963 | core | 0.119 | NA | 0.843 | NA | S | USh | 0.04 | animal |
| Campylospermum flavum | 0 | forest | 0.961 | core | 0.234 | NA | 0.035 | wet | S | UT | - | animal |
| Dasylepis racemosa | 0 | forest | 0.925 | NA | 0.680 | NA | 0.441 | NA | S | UT | 0.43 | animal |
| Santiria trimera | 0 | forest | 0.922 | NA | 0.024 | low | 0.085 | NA | S | CT&UT | 1.32 | animal |
| Garcinia smeathmannii | 0 | forest | 0.905 | NA | 0.141 | NA | 0.712 | NA | S | UT | 2.57 | animal |
| Strombosia scheffleri | 0 | forest | 0.872 | NA | 0.027 | low | 0.099 | NA | S | CT | - | animal |
| Trichilia monadelpha | 0 | forest | 0.862 | NA | 0.220 | NA | 0.046 | wet | S | CT | 0.50 | animal |
| Sapium ellipticum | 0 | forest | 0.841 | NA | 0.121 | NA | 0.014 | wet | P | CT | - | animal |
| Diospyros monbuttensis | 0 | forest | 0.743 | NA | 0.353 | NA | 0.913 | NA | S | UT | 0.68 | animal |
| Kigellia africana | 0 | forest | 0.606 | NA | 0.272 | NA | 0.397 | NA | NPLD | CT | - | animal |
| Psychotria peduncularis | 0 | forest | 0.547 | NA | 0.263 | NA | 0.831 | NA | P | USh | 0.03 | animal |
| Antidesma vogelianum | 0 | forest | 0.515 | NA | 0.196 | NA | 0.675 | NA | P | USh | 0.03 | animal |
| Trilepisium madagascariense | 0 | forest | 0.451 | NA | 0.479 | NA | 0.640 | NA | - | CT | 0.97 | animal |
| Symphonia globulifera | 0 | forest | 0.444 | NA | 0.056 | NA | 0.409 | NA | S | CT | 0.45 | animal |
| Polyscias fulva | 0 | forest | 0.442 | NA | 0.228 | NA | 0.536 | NA | P/CP | UT | 0.01 | animal |
| Celtis gomphophylla | 0 | forest | 0.430 | NA | 0.154 | NA | 0.662 | NA | P/CP | UT | 0.01 | animal |
| Carapa oreophila | 0 | forest | 0.411 | NA | 0.032 | low | 0.063 | NA | NPLD | UT | 18.51 | gravity |
| Xymalos monospora | 0 | forest | 0.398 | NA | 0.667 | NA | 0.172 | NA | S | UT | 0.17 | animal |
| Parkia filicoidea | 0 | forest | 0.355 | NA | 0.048 | low | 0.939 | NA | NPLD | CT | 0.77 | animal |
| Mallotus oppositifolius | 0 | forest | 0.317 | NA | 0.418 | NA | 0.115 | NA | S | USh | 0.02 | animal |
| Zanthoxylum leprieurii | 0 | forest | 0.215 | NA | 0.803 | NA | 0.540 | NA | CP | UT | 0.02 | animal |
| Ficus sur | 0 | forest | 0.211 | NA | 0.351 | NA | 0.165 | NA | Strangler | Strangler | - | animal |
| Leea guineensis | 0 | forest | 0.202 | NA | 0.620 | NA | 0.569 | NA | CP | USh | 0.04 | animal |
| Psychotria succulenta | 0 | forest | 0.189 | NA | 0.358 | NA | 0.855 | NA | P | USh | - | animal |
| Trema orientalis | 0 | forest | 0.169 | NA | 0.011 | low | 0.236 | NA | P/ CP | UT | 0.01 | animal |
| Nuxia congesta | 0 | forest | 0.167 | NA | 0.017 | low | 0.382 | NA | P | UT | <0.001 | animal |
| Albizia gummifera | 0 | forest | 0.136 | NA | 0.980 | high | 0.664 | NA | P/CP | CT | 0.55 | wind |
| Entandophragma angolense | 0 | forest | 0.133 | NA | 0.063 | NA | 0.885 | NA | NPLD | ET | 1.73 | wind |
| Macaranga occidentalis | 0 | forest | 0.107 | NA | 0.327 | NA | 0.008 | wet | P | UT | 0.01 | animal |
| Deinbollia pinnata | 0 | forest | 0.104 | NA | 0.855 | NA | 0.841 | NA | S | UT | 0.62 | animal |
| Pouteria altissima | 0 | forest | 0.103 | NA | 0.833 | NA | 0.226 | NA | NPLD | ET | 3.19 | animal |
| Ficus lutea | 0 | forest | 0.051 | NA | 0.729 | NA | 0.213 | NA | Strangler | Strangler | - | animal |
| Pleiocarpa pycnantha | 0 | forest | 0.030 | edge | 0.892 | NA | 0.530 | NA | S | UT | 0.25 | animal |
| Beilschmiedia mannii | 0 | forest | 0.013 | edge | 0.816 | NA | 0.464 | NA | S | UT | 1.16 | animal |
| Isolona sp. | 0 | forest | 0.010 | edge | 0.870 | NA | 0.649 | NA | S | UT | 1.62 | animal |
| Millettia conraui | 0 | forest | 0.007 | edge | 0.796 | NA | 0.326 | NA | P/CP | UT | 0.20 | ballistic |
| Warneckea cinnamomoides | 0 | forest | 0.005 | edge | 0.664 | NA | 0.623 | NA | P/CP | USh | - | animal |
| Rauvolfia vomitoria | 0 | forest | 0.002 | edge | 0.520 | NA | 0.829 | NA | P/CP | USh | 0.07 | animal |
| Rothmania urcelliformis | 0 | forest | 0.002 | edge | 0.999 | high | 0.551 | NA | S | UT | 0.31 | animal |
| Eugenia gilgii | 0 | forest | 0.001 | edge | 0.806 | NA | 0.457 | NA | P | UT | 0.11 | animal |
| Ritchiea albersii | 0 | forest | 0.001 | edge | 0.996 | high | 0.774 | NA | S | UT | - | animal |
| Clausena anisata | 0 | forest | 0 | edge | 0.966 | high | 0.783 | NA | P/CP | USh | 0.07 | animal |
| Sorindeia sp. | 0 | forest | 0 | edge | 0.754 | NA | 0.713 | NA | NPLD | CT | 0.30 | animal |
| Tabanaemontana contorta | 0 | forest | 0 | edge | 0.971 | high | 0.922 | NA | P/CP | UT | 0.35 | animal |
| Anthonotha noldeae | 0 | forest | 0 | edge | 0.968 | high | 0.696 | NA | P/CP | CT | 6.57 | ballistic |
| Bridelia speciosa | 0 | forest | 0 | edge | 0.510 | NA | 0.847 | NA | P/CP | UT | - | wind |
| Ficus sp. | 0 | forest | 0 | edge | 0.945 | NA | 0.440 | NA | Strangler | Strangler | - | animal |
| Psychotria umbellata | 0 | forest | 0 | edge | 0.602 | NA | 0.923 | NA | P | USh | - | animal |
| Guarea cedrata | 0 | forest | 0 | edge | 0.300 | NA | 0.896 | NA | P/CP | UT | - | animal |
| Psorospermum aurantiacum | 1 | savanna | 0 | edge | 0.751 | NA | 0.907 | NA | G | Sh | 0.01 | animal |
| Entada abyssinica | 1 | savanna | 0 | edge | 0.999 | high | 0.852 | NA | G | ST | 0.25 | wind |
| Dombeya ledermannii | 1 | savanna | 0 | edge | 0.840 | NA | 0.467 | NA | G | ST | - | ballistic |
| Rytigynia sp. | 1 | savanna | 0 | edge | 0.797 | NA | 0.804 | NA | G | Sh | - | animal |
| Rubiaceae unidentified | 0 | forest | 0.998 | core | 0.359 | NA | 0.518 | NA | S | UT | - | animal |
| Unidentified | 0 | forest | 0.033 | edge | 0.475 | NA | 0.553 | NA | - | - | - | |
| Unidentified | 0 | forest | 0.002 | edge | 0.997 | high | 0.350 | NA | P | U | - | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abiem, I.; Arellano, G.; Kenfack, D.; Chapman, H. Afromontane Forest Diversity and the Role of Grassland-Forest Transition in Tree Species Distribution. Diversity 2020, 12, 30. https://doi.org/10.3390/d12010030
Abiem I, Arellano G, Kenfack D, Chapman H. Afromontane Forest Diversity and the Role of Grassland-Forest Transition in Tree Species Distribution. Diversity. 2020; 12(1):30. https://doi.org/10.3390/d12010030
Chicago/Turabian StyleAbiem, Iveren, Gabriel Arellano, David Kenfack, and Hazel Chapman. 2020. "Afromontane Forest Diversity and the Role of Grassland-Forest Transition in Tree Species Distribution" Diversity 12, no. 1: 30. https://doi.org/10.3390/d12010030
APA StyleAbiem, I., Arellano, G., Kenfack, D., & Chapman, H. (2020). Afromontane Forest Diversity and the Role of Grassland-Forest Transition in Tree Species Distribution. Diversity, 12(1), 30. https://doi.org/10.3390/d12010030