Diversity of a Large Collection of Natural Populations of Mango (Mangifera indica Linn.) Revealed by Agro-Morphological and Quality Traits

Abstract
1. Introduction
2. Materials and Methods
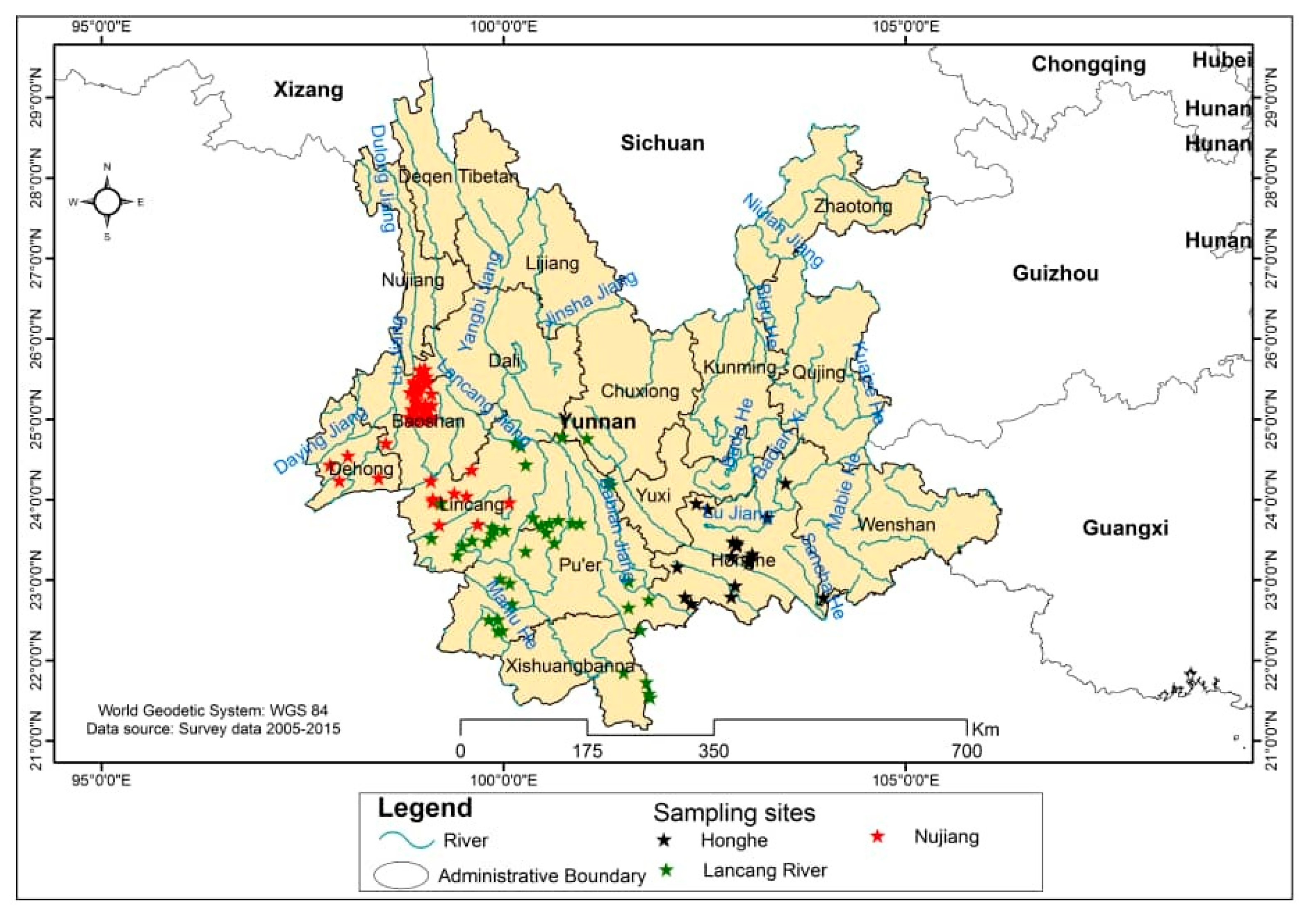
2.1. Description of the Mango Germplasm Collection Sites
2.1.1. Nujiang
2.1.2. Honghe
2.1.3. Lancang River
2.2. Mango Germplasm Resources Evaluation
2.2.1. Monitoring of the Tree Phenology
2.2.2. Measurement of Dendrometric Parameters
2.2.3. Measurement of the Leaf Parameters
2.2.4. Measurement of the Flower Parameters
2.2.5. Measurement of Fruit and Seed Parameters
2.3. Statistical Analysis
3. Results
3.1. Variation of Qualitative Traits in the Mango Tree Collection
3.2. Quantitative Traits Variation in the Germplasm
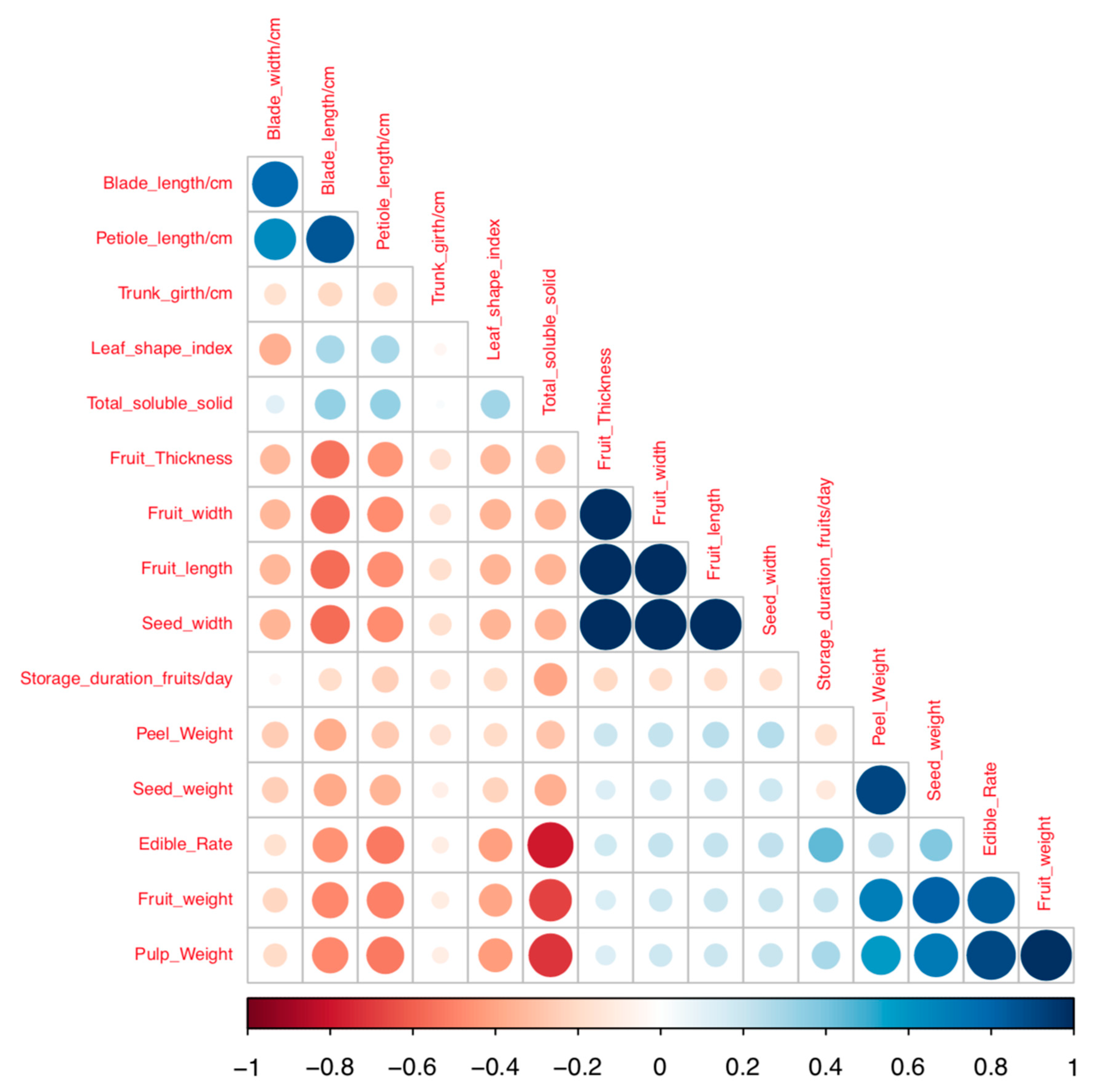
3.3. Correlation Analysis for Quantitative Traits
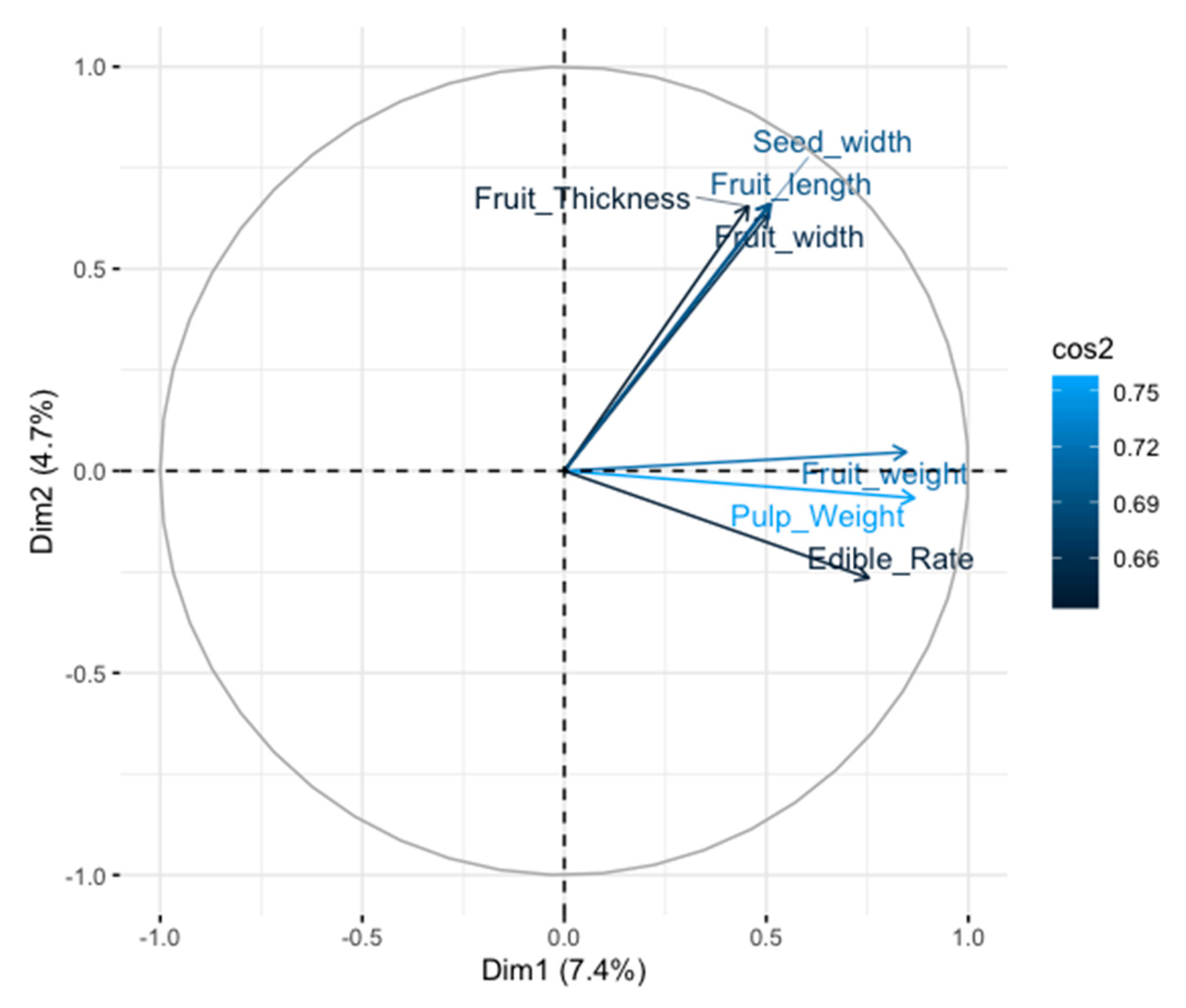
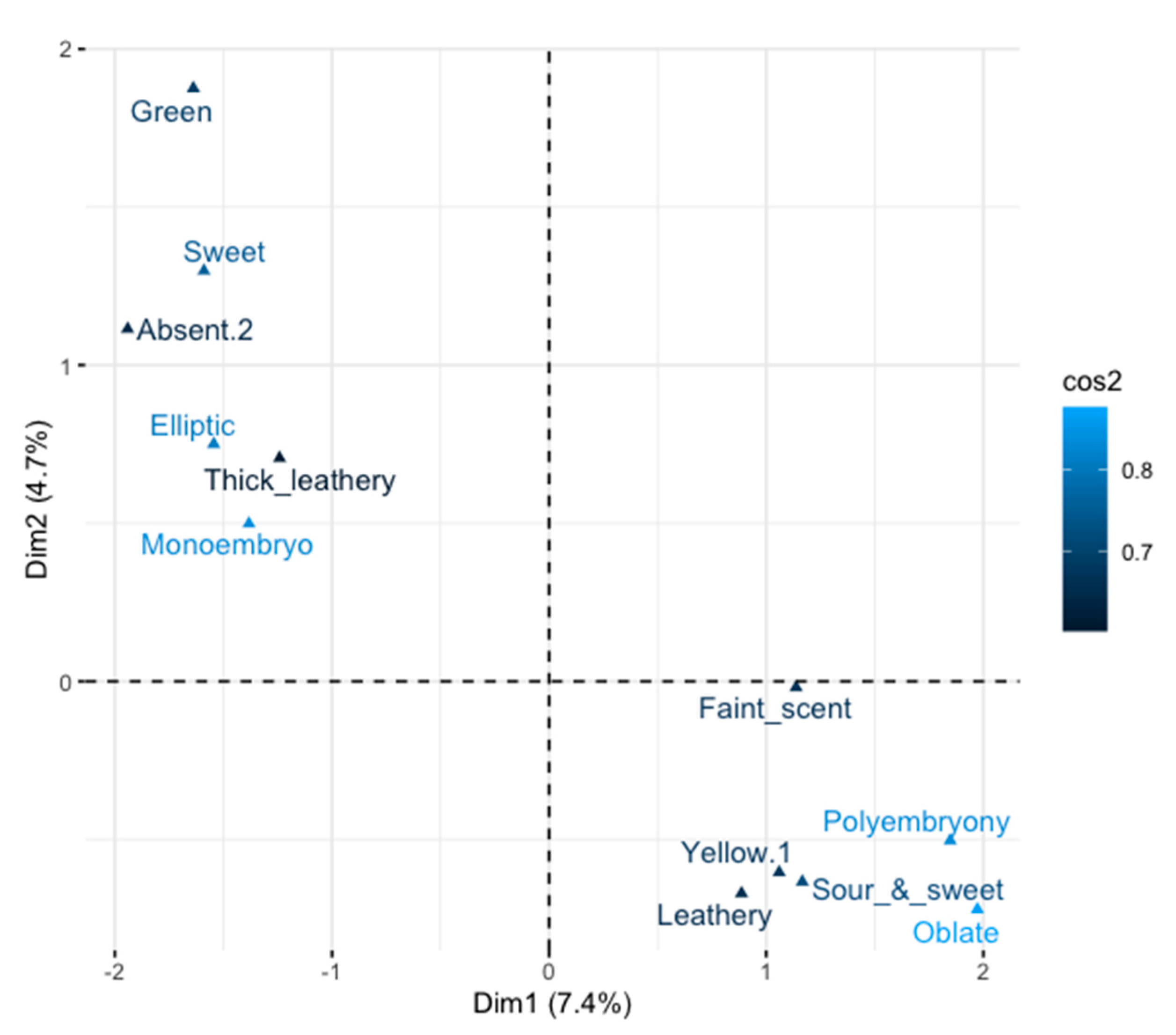
3.4. Relative Importance of Each Descriptor and Variability in the Germplasm
4. Discussion
4.1. Variability in the Mango Germplasm
4.2. Implications for Conservation and Utilization of Mango Genetic Resources
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Saúco, V.G. Trends in world mango production and marketing. Acta Hortic. 2017, 1183, 351–363. [Google Scholar] [CrossRef]
- Douthett, B.D.G. The mango: Asia’s king of fruits. Ethnobot. Leafl. 2001, 2000, 4. [Google Scholar]
- Huang, C.Y.; Kuo, C.H.; Wu, C.H.; Kuan, A.W.; Guo, H.R.; Lin, Y.H.; Wang, P.K. Free radical-scavenging, anti-inflammatory, and antibacterial activities of water and ethanol extracts prepared from compressional-puffing pretreated mango (Mangifera indica L.) peels. J. Food Qual. 2018, 2018, 1025387. [Google Scholar] [CrossRef]
- Schulze-Kaysers, N.; Feuereisen, M.M.; Schieber, A. Phenolic compounds in edible species of the Anacardiaceae family—A review. RSC Adv. 2015, 5, 73301–73314. [Google Scholar] [CrossRef]
- Shah, K.A.; Patel, M.B.; Patel, R.J.; Parmar, P.K. Mangifera indica (Mango). Pharmacogn. Rev. 2019, 4, 42–48. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAO Statistical Division. Available online: http://www.fao.org/statistics/en/ (accessed on 10 January 2020).
- Gao, A.; Chen, Y.; Luo, R.; Huang, J.; Zhao, Z.; Wang, W.; Wang, Y.; Dang, Z. Development status of Chinese mango industry in 2018. Adv. Agric. Hortic. Entomol. 2019, 2019, 1–6. [Google Scholar]
- Chen, Q.B. Perspectives on the mango industry in mainland China. In Proceedings of the IX International Mango Symposium, Sanya, China, 8–12 April 2013; pp. 25–36. [Google Scholar]
- Luo, C.; He, X.H.; Chen, H.; Hu, Y.; Ou, S.J. Genetic relationship and diversity of Mangifera indica L.: Revealed through SCoT analysis. Genet. Resour. Crop Evol. 2012, 59, 1505–1515. [Google Scholar] [CrossRef]
- Normand, F.; Lauri, P.; Legave, J.M. Climate change and its probable effects on mango production and cultivation. Acta Hortic. 2015, 1075, 21–32. [Google Scholar] [CrossRef]
- Khan, I.A.; Khan, A.S.; Rajwana, I.A.; Khan, A.A.; Azmat, M.A.; Raza, S.A. Premium quality mango genotypes for extended harvest season. HortScience 2016, 51, 1609–1612. [Google Scholar] [CrossRef]
- Bally, I.S.E.; Dillon, N.L. Mango (Mangifera indica L.). In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer International Publishing AG: Basel, Switzerland, 2018; pp. 811–896. ISBN 9783319919447. [Google Scholar]
- Rajan, S.; Hudedamani, U. Genetic resources of mango: Status, threats, and future prospects. In Conservation and Utilization of Horticultural Genetic Resources; Rajasekharan, P.E., Rao, V.R., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2019; pp. 217–250. ISBN 9789811336683. [Google Scholar]
- Warschefsky, E.J.; von Wettberg, E.J.B. Population genomic analysis of mango (Mangifera indica) suggests a complex history of domestication. New Phytol. 2019, 222, 2023–2037. [Google Scholar] [CrossRef]
- FAO. The Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2010. [Google Scholar]
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace germplasm for improving yield and abiotic stress adaptation. Trends Plant Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Sennhenn, A.; Prinz, K.; Gebauer, J.; Whitbread, A.; Jamnadass, R.; Kehlenbeck, K. Identification of mango (Mangifera indica L.) landraces from Eastern and Central Kenya using a morphological and molecular approach. Genet. Resour. Crop Evol. 2014, 61, 7–22. [Google Scholar] [CrossRef]
- Khan, A.S.; Ali, S.; Khan, I.A. Morphological and molecular characterization and evaluation of mango germplasm: An overview. Sci. Hortic. (Amsterdam) 2015, 194, 353–366. [Google Scholar] [CrossRef]
- Luo, C.; He, X.H.; Chen, H.; Ou, S.J.; Gao, M.P.; Brown, J.S.; Tondo, C.T.; Schnell, R.J. Genetic diversity of mango cultivars estimated using SCoT and ISSR markers. Biochem. Syst. Ecol. 2011, 39, 676–684. [Google Scholar] [CrossRef]
- Rajwana, A.I.; Tabbasam, N.; Malik, A.U.; Malik, S.A.; Mehboob-ur-Rahman; Zafar, Y. Assessment of genetic diversity among mango (Mangifera indica L.) genotypes using RAPD markers. Sci. Hortic. (Amsterdam) 2008, 117, 297–301. [Google Scholar] [CrossRef]
- Ramessur, A.D.; Ranghoo-Sanmukhiya, V.M. RAPD marker-assisted identification of genetic diversity among mango (Mangifera indica) varieties in Mauritius. Int. J. Agric. Biol. 2011, 13, 167–173. [Google Scholar]
- Gitahi, R.; Kasili, R.; Kyallo, M.; Kehlenbeck, K. Diversity of threatened local mango landraces on smallholder farms in Eastern Kenya. For. Trees Livelihoods 2016, 25, 239–254. [Google Scholar] [CrossRef]
- Rajan, S.; Kumar, R.; Yadava, L.P.; Sharan, R.; Bhal, C.; Verma, J.P. Variability pattern in mango (Mangifera indica L.) accessions of diverse geographical origins. Acta Hortic. 2013, 992, 341–352. [Google Scholar] [CrossRef]
- Mussane, C.R.B.; Biljon, A.V.; Herselman, L. Morphological and genetic characterization of mango varieties in Mozambique. In Proceedings of the Second RUFORUM Biennial Meeting, Entebbe, Uganda, 20–24 September 2010; pp. 991–995. [Google Scholar]
- Ye, C.; Li, Y.; Li, Q.; Hua, W.; Bian, Z. Tropical Crop Germplasm Resources Data Standards (Chinese Edition); China Agriculture Press: Beijing, China, 2000; ISBN 978-7109135673. [Google Scholar]
- IPGRI. Descriptors for Mango (Mangifera indica); IPGRI: Rome, Italy, 2006; Volume 84, ISBN 9789290436522. [Google Scholar]
- Joshi, N.; Joshi, A. Urban tree canopy analysis. Pollut. Res. 2014, 33, 427–431. [Google Scholar]
- Cichy, K.A.; Snapp, S.S.; Blair, M.W. Plant growth habit, root architecture traits and tolerance to low soil phosphorus in an Andean bean population. Euphytica 2009, 165, 257–268. [Google Scholar] [CrossRef]
- Lenard, E. Habits of trees and shrubs in landscape design. Archit. Civ. Eng. Environ. 2008, 1, 13–20. [Google Scholar]
- SAC. Fresh Fruits and Vegetables—Sampling; GB/T 8855-2008; The Standardization Administration of China: Beijing, China, 2008.
- Jha, S.K.; Sethi, S.; Srivastav, M.; Dubey, A.K.; Sharma, R.R.; Samuel, D.V.K.; Singh, A.K. Firmness characteristics of mango hybrids under ambient storage. J. Food Eng. 2010, 97, 208–212. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Technol. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Abdi, H. Bonferroni test. In Encyclopedia of Measurement and Statistics; Salkind, J.N., Ed.; SAGE: Thousand Oaks, CA, USA, 2007; pp. 103–107. [Google Scholar]
- Pagès, J. Multiple Factor Analysis by Example Using R; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Husson, F.; Lê, S.; Pagès, J. Exploratory Multivariate Analysis by Example Using R, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Sebastien, L.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix, Version 0.84; CRAN: 2017. Available online: https://github.com/taiyun/corrplot (accessed on 10 January 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Toili, M.; Rimberia, F.; Nyende, A.; Sila, D. Morphological diversity of mango germplasm from the Upper Athi River region of eastern Kenya: An analysis based on non- fruit descriptors. African J. Food, Agric. Nutr. Dev. 2016, 16, 10913–10935. [Google Scholar] [CrossRef]
- Wang, S.B.; Wu, H.X.; Ma, W.H.; Ma, X.W.; Zhan, R.L.; Yao, Q.S.; Sun, G.M.; Xie, J.H. Evaluation of sixteen introduced mango cultivars in Zhanjiang, China. Acta Hortic. 2013, 992, 211–220. [Google Scholar] [CrossRef]
- Rymbai, H.; Laxman, R.H.; Dinesh, M.R.; Sunoj, V.S.J.; Ravishankar, K.V.; Jha, A.K. Diversity in leaf morphology and physiological characteristics among mango (Mangifera indica) cultivars popular in different agro-climatic regions of India. Sci. Hortic. (Amsterdam) 2014, 176, 189–193. [Google Scholar] [CrossRef]
- Rajan, S.; Kumar, R.; Negi, S.S. Variation in canopy characteristics of mango (Mangifera indica L.) cultivars from diverse eco-geographical regions. J. Appl. Hort 2001, 3, 95–97. [Google Scholar]
- Krishnapillai, N.; Wilson Wijeratnam, R.S. Morphometric analysis of mango varieties in Sri Lanka. Aust. J. Crop Sci. 2016, 10, 784–792. [Google Scholar] [CrossRef]
- Ahmed, T.H.M.; Mohamed, Z.M.A. Diversity of mango (Mangifera indica L.) cultivars in Shendi Area: Morphological fruit characterization. Int. J. Res. Agric. Sci. 2015, 2, 2348–3997. [Google Scholar]
- Kulkarni, M.M.; Burondkar, M.M.; Dalvi, N.V.; Salvi, B.R.; Haldankar, P.M.; Bhattacharyya, T. Mango fruit size diversity found in Konkan. Adv. Agric. Res. Technol. J. 2019, 3, 43–46. [Google Scholar]
- Skinner, D.Z.; Bauchan, G.R.; Auricht, G.; Hughes, S. A method for the efficient management and utilization of large germplasm collections. Crop Sci. 1999, 39, 1237–1242. [Google Scholar] [CrossRef]
- Maia, M.C.C.; de Araújo, L.B.; dos Santos Dias, C.T.; de Oliveira, L.C.; Vasconcelos, L.F.L.; de Carvalho Júnior, J.E.V.; Simeão, M.; Bastos, Y.G.M. Selection of mango rosa genotypes in a breeding population using the multivariate-biplot method. Ciência Rural 2016, 46, 1689–1694. [Google Scholar] [CrossRef]
- Jolliffe, I.T. Principal Component Analysis, 2nd ed.; Bickel, P., Diggle, P., Fienberg, S., Krickeberg, K., Olkin, I., Wermuth, N., Zeger, S., Eds.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Prohens, J.; Blanca, J.M.; Nuez, F. Morphological and molecular variation in a collection of eggplants from a secondary center of diversity: Implications for conservation and breeding. J. Am. Soc. Hortic. Sci. 2005, 130, 54–63. [Google Scholar] [CrossRef]
- Cobb, J.N.; Juma, R.U.; Biswas, P.S.; Arbelaez, J.D.; Rutkoski, J.; Atlin, G.; Hagen, T.; Quinn, M.; Ng, E.H. Enhancing the rate of genetic gain in public-sector plant breeding programs: Lessons from the breeder’s equation. Theor. Appl. Genet. 2019, 132, 627–645. [Google Scholar] [CrossRef] [PubMed]
- Negi, S.S.; Rajan, S.; Kumar, R. Developing new mango varieties through hybridization. Acta Horticulturae 2000, 509, 159–160. [Google Scholar] [CrossRef]
- Vasanthaiah, H.K.N.; Ravishankar, K.V.; Mukunda, G.K. Mango. In Genome Mapping and Molecular Breeding in Plants. Volume 4: Fruits and Nuts; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Luo, C.; Shu, B.; Yao, Q.; Wu, H.; Xu, W.; Wang, S. Construction of a high-density genetic map based on large-scale marker development in mango using specific-locus amplified fragment sequencing (SLAF-seq). Front. Plant Sci. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Ortiz, R. A mini core subset for capturing diversity and promoting utilization of chickpea genetic resources in crop improvement. Theor. Appl. Genet. 2001, 102, 1292–1298. [Google Scholar] [CrossRef]
- Archak, S.; Tyagi, R.K.; Harer, P.N.; Mahase, L.B.; Singh, N.; Dahiya, O.P.; Nizar, M.A.; Singh, M.; Tilekar, V.; Kumar, V.; et al. Characterization of chickpea germplasm conserved in the Indian National Genebank and development of a core set using qualitative and quantitative trait data. Crop J. 2016, 4, 417–424. [Google Scholar] [CrossRef]
- Malosetti, M.; Abadie, T. Sampling strategy to develop a core collection of Uruguayan maize landraces based on morphological traits. Genet. Resour. Crop Evol. 2001, 48, 381–390. [Google Scholar] [CrossRef]
- Bhattacharjee, R.; Dumet, D.; Ilona, P.; Folarin, S.; Franco, J. Establishment of a cassava (Manihot esculenta Crantz) core collection based on agro-morphological descriptors. Plant Genet. Resour. 2012, 10, 119–127. [Google Scholar] [CrossRef]
- Girma, G.; Bhattacharjee, R.; Lopez-Montes, A.; Gueye, B.; Ofodile, S.; Franco, J.; Abberton, M. Re-defining the yam (Dioscorea spp.) core collection using morphological traits. Plant Genet. Resour. 2018, 16, 193–200. [Google Scholar] [CrossRef]
- Gajanana, T.; Dinesh, M.; Rajan, S.; Vasudeva, R.; Singh, S.K.; Lamers, H.A.; Parthasarathy, V.; Sthapit, B.; Rao, V.R. Motivation for on-farm conservation of mango (Mangifera indica) Diversity in India—A case study. Indian J. Plant Genet. Resour. 2015, 28, 1. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Shannon–Weaver Diversity Index |
|---|---|
| Crown shape | 0.3 |
| Leaf blade shape | 0.8 |
| Leaf surface | 0.7 |
| Leaf texture | 1.0 |
| Leaf apex shape | 0.1 |
| Leaf margin | 0.6 |
| Inflorescence shape | 0.4 |
| Type flower | 1.1 |
| Flesh color | 1.5 |
| Petals color | 0.1 |
| Anther color | 0.6 |
| Fruit shape | 1.2 |
| Fruit beak type | 0.6 |
| Fruit sinus type | 0.6 |
| Pericarp color | 1.3 |
| Flowering habit | 0.4 |
| Fruit maturity characteristics | 0.5 |
| Kernel shape | 0.8 |
| Type embryos | 0.8 |
| Fruit aroma | 1.3 |
| Fruit flavor | 1.2 |
| Tree growth habit | 0.5 |
| Quantity fiber pulp | 1 |
| Fiber length pulp | 1 |
| Edible quality | 0.7 |
| Average | 0.8 ± 0.3 |
| Quantitative Descriptors | Mean ± SD | Coefficient of Variation (%) | Range |
|---|---|---|---|
| Dendrometric traits | |||
| Trunk girth (cm) | 426 ± 101 | 34 | 314–753 |
| Blade length (cm) | 21 ± 3.7 | 17 | 12.7–37.7 |
| Blade width (cm) | 5.6 ± 1 | 17 | 3.2–9.7 |
| Leaf shape index | 3.8 ± 0.5 | 14 | 2.3–6.7 |
| Petiole length (cm) | 4.1 ± 1 | 23 | 2.1–12.6 |
| Fruit traits | |||
| Storage duration of fruits (days) | 3.7 ± 1.1 | 30 | 2–9 |
| Fruit weight (g) | 88.9 ± 47.6 | 53 | 10.4–272 |
| Peel weight (g) | 13.4 ± 5.5 | 41 | 2.1–33.9 |
| Pulp weight (g) | 56.8 ± 40.4 | 71 | 3.2–225 |
| Fruit length (cm) | 7.6 ±1.8 | 23 | 2.5–12.4 |
| Fruit width (cm) | 4.5 ± 0.9 | 21 | 1.2–7.5 |
| Fruit thickness (cm) | 4 ± 0.7 | 18 | 1–6 |
| Total soluble solid | 17.4 ± 4.0 | 23 | 1.8–61.6 |
| Edible rate (%) | 58.3 ± 14.1 | 24 | 14–86.5 |
| Seed traits | |||
| Seed weight (g) | 19.1 ± 6.9 | 36 | 1–51 |
| Seed width (cm) | 6.7 ± 1.6 | 24 | 2.4–11.1 |
| Quantitative Descriptors | Mean ± SD | p-Value | ||
|---|---|---|---|---|
| Nujiang | Lancang River | Honghe | ||
| Leaf traits | ||||
| Blade width (cm) | 5.7 ± 0.9 | 5.4 ± 1.2 | 5.8 ± 1.1 | 0.034 |
| Leaf shape index | 3.8 ± 0.5 | 4.0 ±0.6 | 3.9 ± 0.4 | 0.001 |
| Petiole length (cm) | 4.0 ± 0.9 | 4.4 ± 1.3 | 4.5 ± 0.6 | <0.001 |
| Fruit traits | ||||
| Storage duration of fruits (days) | 3.9 ± 1.2 | 3.2 ± 0.6 | 3.0 ± 0.6 | 0.000 |
| Fruit weight (g) | 93.3 ± 51.7 | 81.2 ± 33.6 | 65.7 ± 14.8 | 0.001 |
| Peel weight (g) | 12.5 ± 5.6 | 16.4 ± 4.7 | 14.3 ± 3.5 | 0.000 |
| Pulp weight (g) | 63.1 ± 43.2 | 42.5 ± 26.6 | 30.4 ± 9.5 | 0.000 |
| Seed weight (g) | 18.1 ± 7.1 | 22.4 ± 5.6 | 21.1 ± 4.2 | 0.000 |
| Total soluble solid | 17.0 ± 4.2 | 18.4 ± 3.0 | 19.0 ± 3.0 | <0.001 |
| Edible rate (%) | 61.9 ± 13.7 | 49.2 ± 9.2 | 44.9 ± 8.3 | 0.000 |
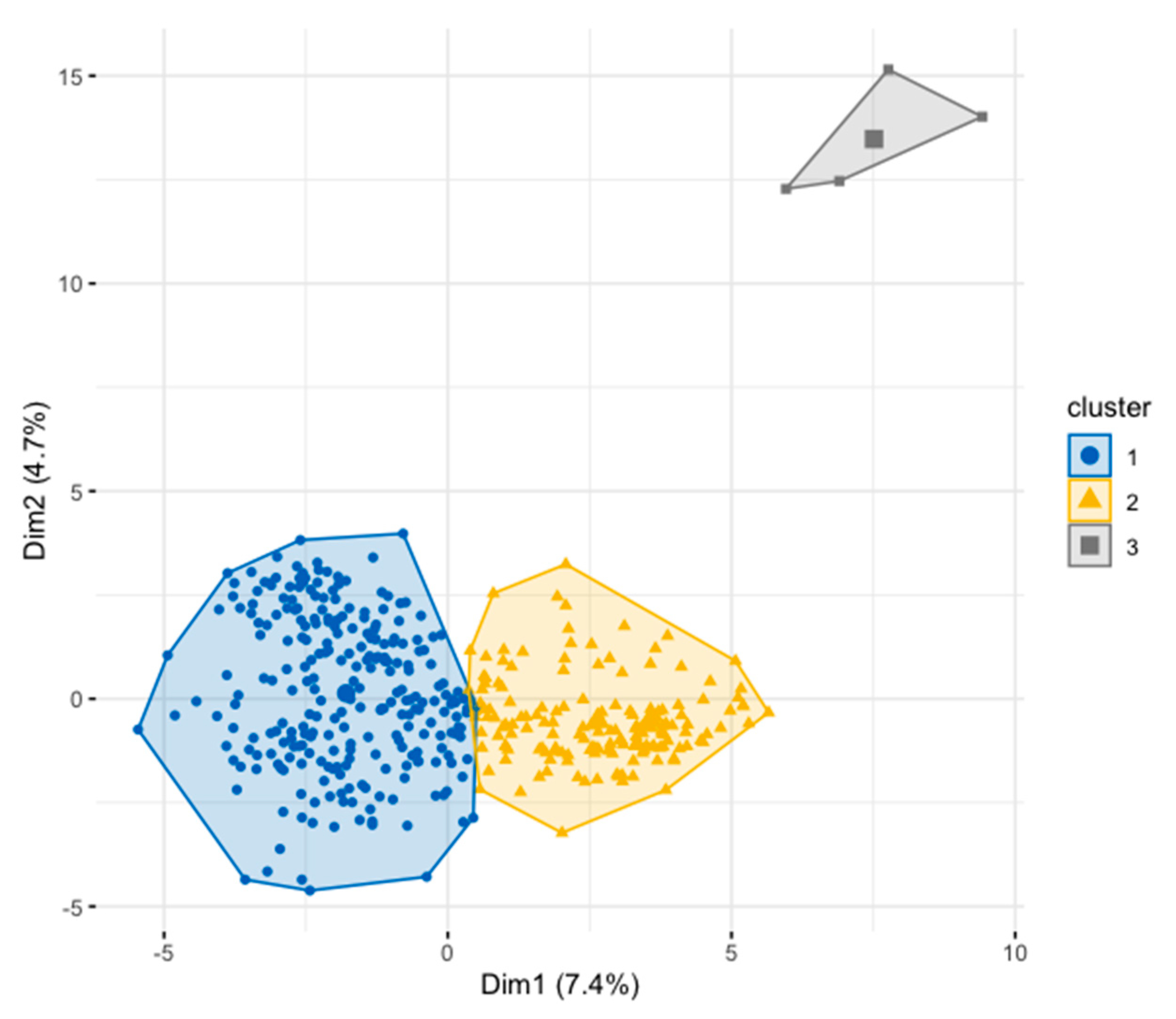
| Quantitative Descriptors | Cluster 1 | Cluster 2 | Cluster 3 | p Value |
|---|---|---|---|---|
| Leaf traits | ||||
| Blade length (cm) | 21.7 ± 4 | 20.8 ± 3.1 | 18 ± 2.0 | 0.012 |
| Leaf shape index | 3.9 ± 0.6 | 3.7 ± 0.4 | 3.5 ± 0.2 | 0.000 |
| Petiole length (cm) | 4.3 ± 1 | 3.9 ± 0.8 | 3.7 ± 0.3 | 0.001 |
| Fruit traits | ||||
| Storage duration of fruits (days) | 3.4 ± 1 | 4.1 ± 1.2 | 3.0 ± 0.0 | 0.000 |
| Fruit weight (g) | 59.8 ± 21.9 | 135.1 ± 37.4 | 82.5 ± 25.3 | 0.000 |
| Peel weight (g) | 11.9 ± 4.7 | 15.8 ± 5.8 | 17.2 ± 2.5 | 0.000 |
| Pulp weight (g) | 31.1 ± 14.5 | 97 ± 32.39 | 50.1 ± 20.3 | 0.000 |
| Fruit length (cm) | 6.6 ± 1.2 | 9.4 ± 1.7 | 7 ± 2.09 | 0.001 |
| Fruit width (cm) | 3.9 ± 0.6 | 5.4 ± 0.8 | 4 ± 1.02 | 0.000 |
| Fruit thickness (cm) | 3.6 ± 0.5 | 4.3 ± 3.2 | 3.6 ± 0.8 | 0.000 |
| Seed weight (g) | 17.1 ± 6.5 | 22.6 ± 6 | 19.7 ± 3.5 | 0.000 |
| Seed width (cm) | 5.8 ± 1.1 | 8.3 ± 1.7 | 6.2 ± 1.88 | 0.001 |
| Total soluble solid | 18.4 ± 4.5 | 15.8 ± 2.5 | 16.9 ± 1.5 | 0.000 |
| Edible rate (%) | 50.6 ± 10.6 | 70.5 ± 8.3 | 58.9 ± 13.3 | 0.000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Xie, D.; Bai, T.; Luo, X.; Zhang, F.; Ni, Z.; Chen, Y. Diversity of a Large Collection of Natural Populations of Mango (Mangifera indica Linn.) Revealed by Agro-Morphological and Quality Traits. Diversity 2020, 12, 27. https://doi.org/10.3390/d12010027
Zhang C, Xie D, Bai T, Luo X, Zhang F, Ni Z, Chen Y. Diversity of a Large Collection of Natural Populations of Mango (Mangifera indica Linn.) Revealed by Agro-Morphological and Quality Traits. Diversity. 2020; 12(1):27. https://doi.org/10.3390/d12010027
Chicago/Turabian StyleZhang, Cuixian, Dehong Xie, Tianqi Bai, Xinping Luo, Faming Zhang, Zhangguang Ni, and Yufu Chen. 2020. "Diversity of a Large Collection of Natural Populations of Mango (Mangifera indica Linn.) Revealed by Agro-Morphological and Quality Traits" Diversity 12, no. 1: 27. https://doi.org/10.3390/d12010027
APA StyleZhang, C., Xie, D., Bai, T., Luo, X., Zhang, F., Ni, Z., & Chen, Y. (2020). Diversity of a Large Collection of Natural Populations of Mango (Mangifera indica Linn.) Revealed by Agro-Morphological and Quality Traits. Diversity, 12(1), 27. https://doi.org/10.3390/d12010027