Living on the Edge: Variation in the Abundance and Demography of a Kelp Forest Epibiont

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
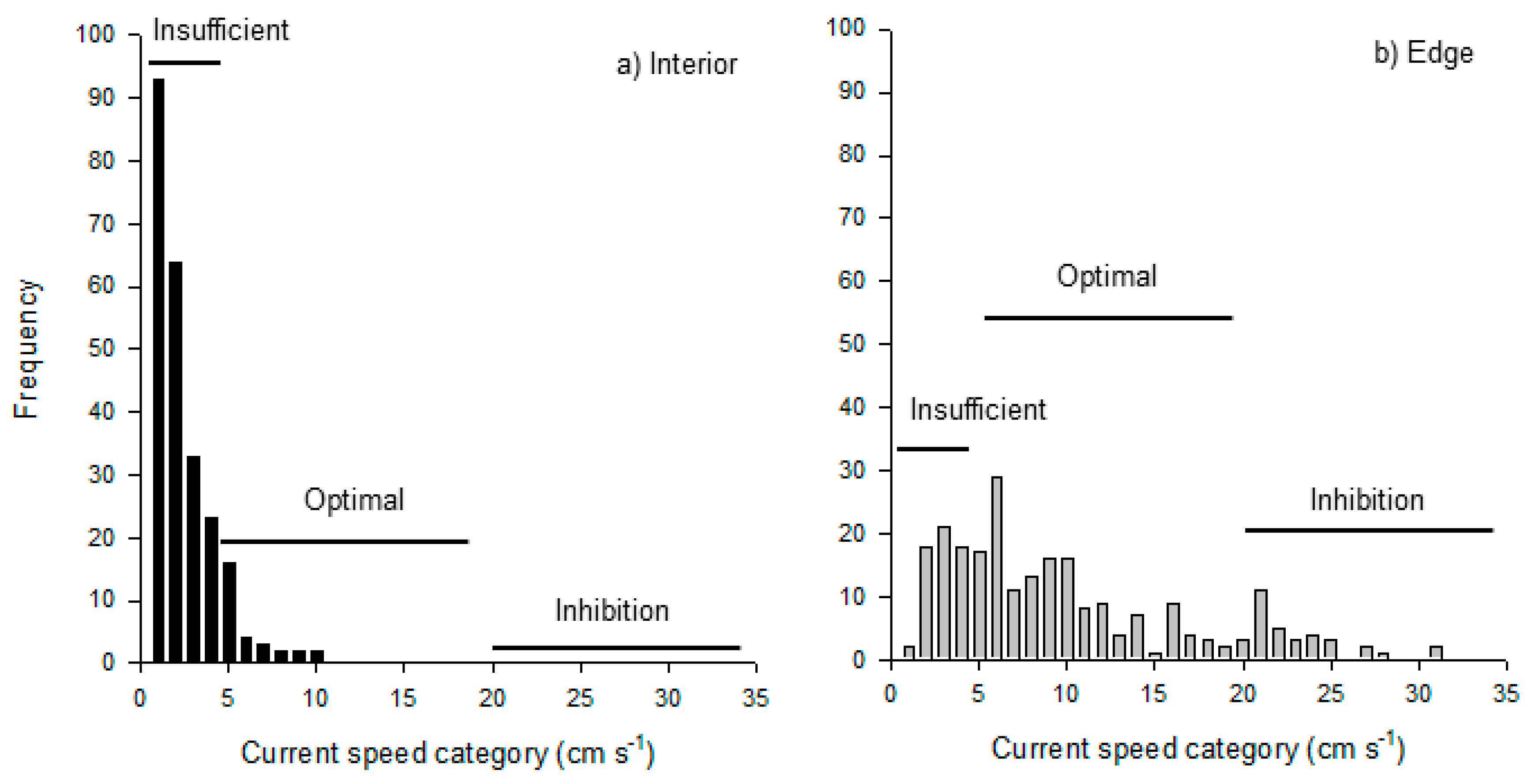{kind=link}
1. Introduction
2. Materials and Methods
2.1. Description of Study Sites and Species
2.2. Distribution and Abundance of Membranipora in Kelp Forests
2.3. Colony and Blade Mortality Within a Kelp Forest
2.4. Larval Recruitment in Kelp Forests
2.5. Clonal Reproduction in Kelp Forests
2.6. Relationships Between Kelp Forest Structure and Membranipora Demography
3. Results
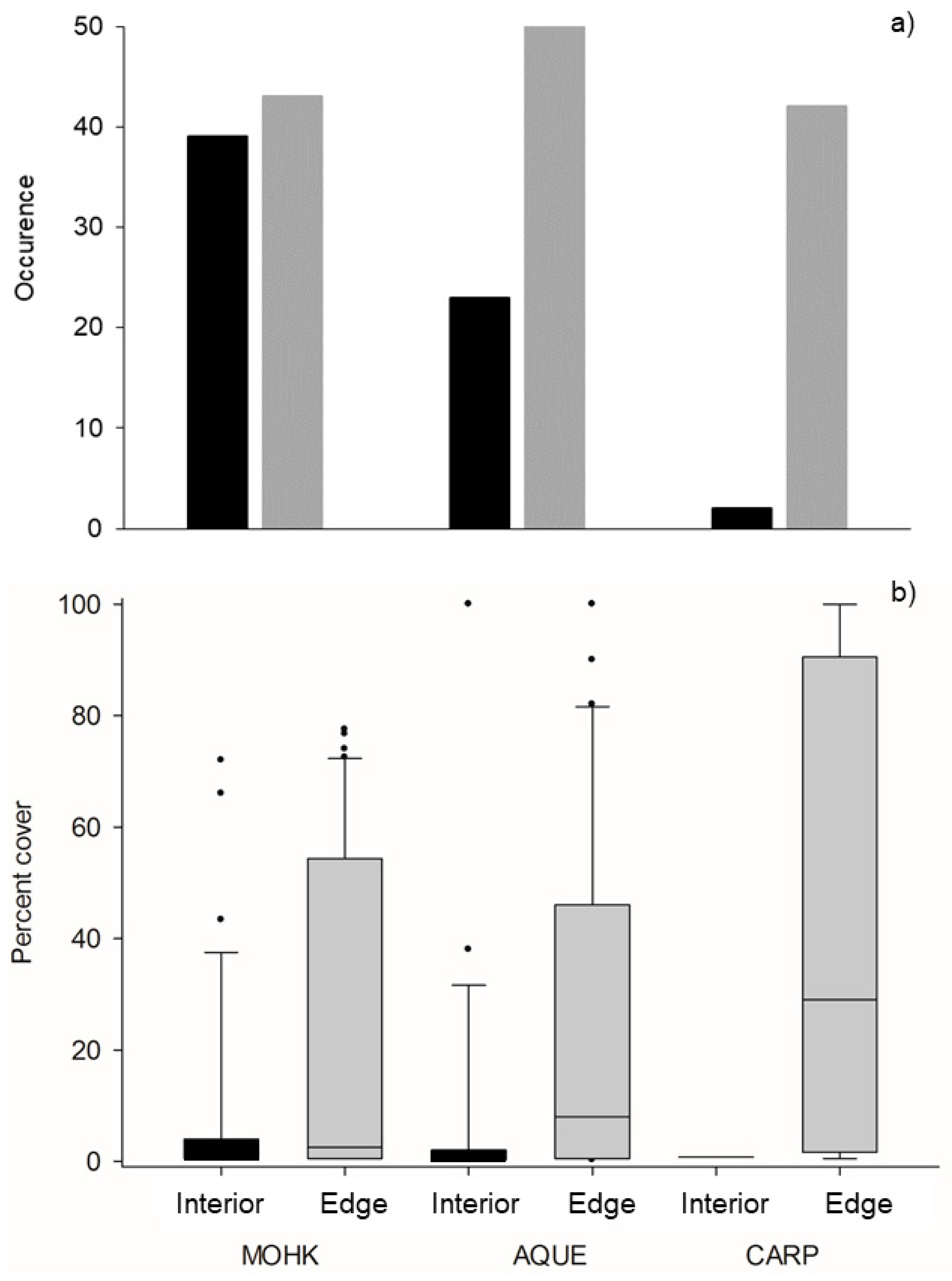
3.1. Distribution and Abundance of Membranipora in Kelp Forests
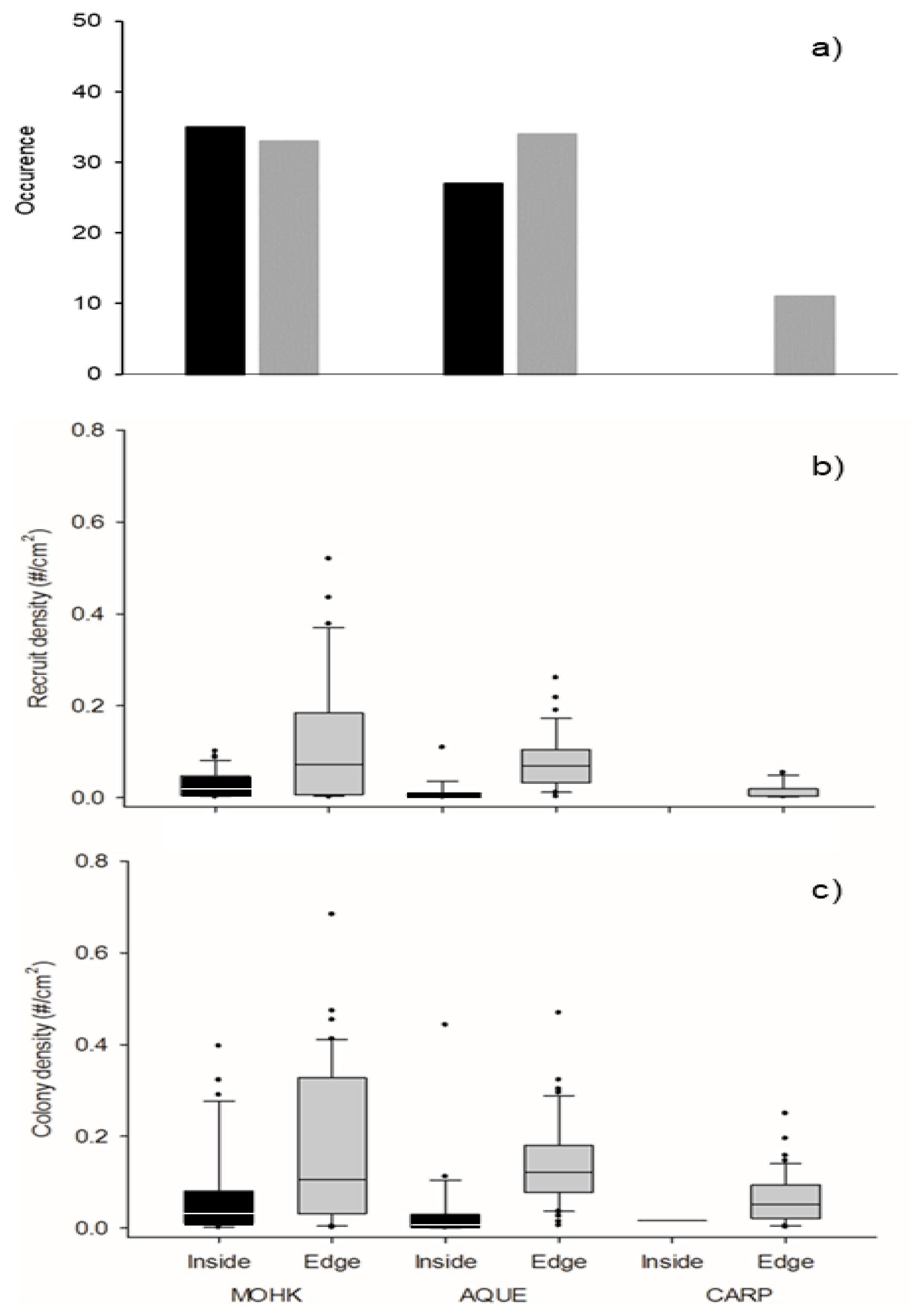
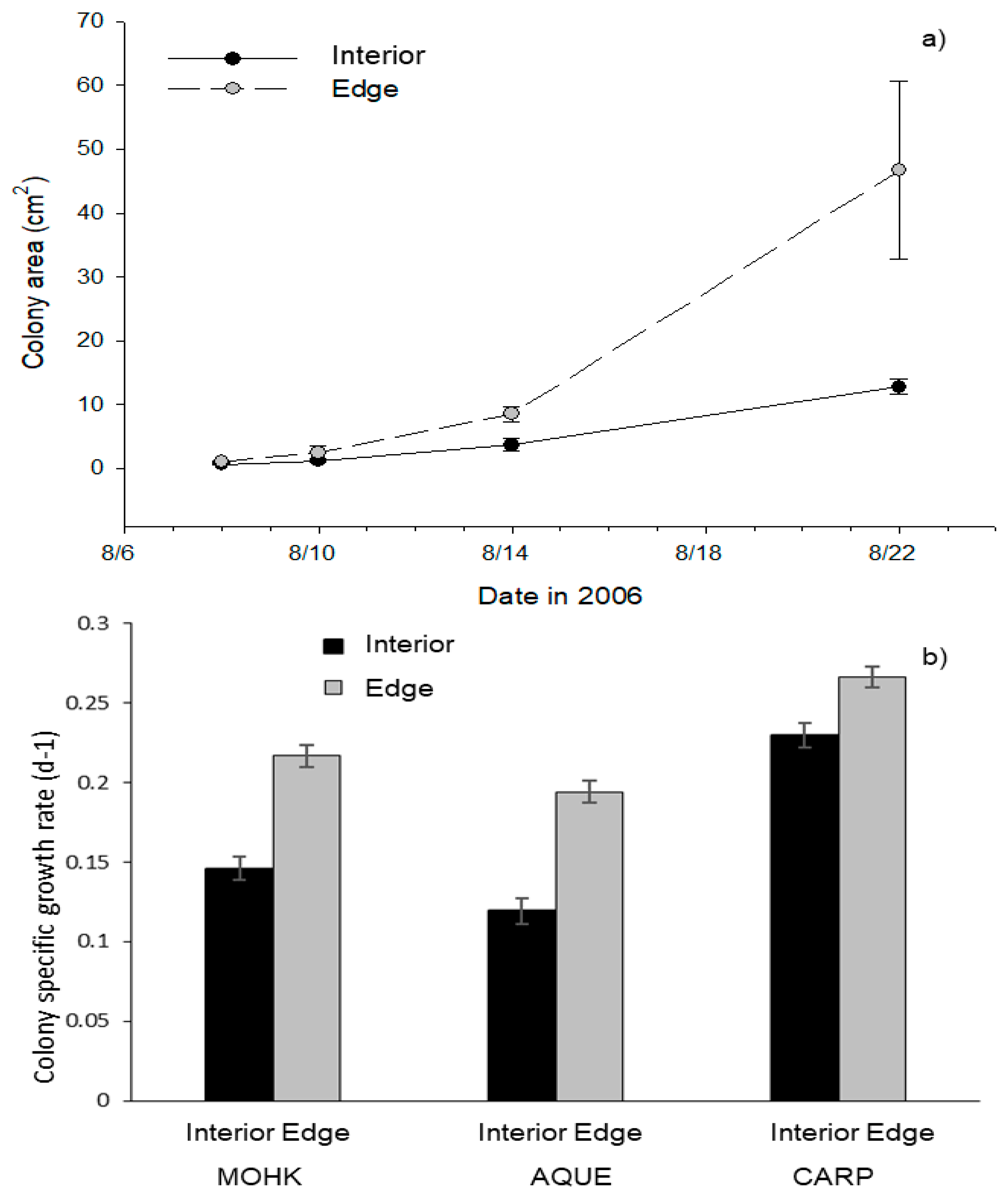
3.2. Colony and Blade Mortality Within a Kelp Forest
3.3. Larval Recruitment in Kelp Forests
3.4. Clonal Reproduction in Kelp Forests
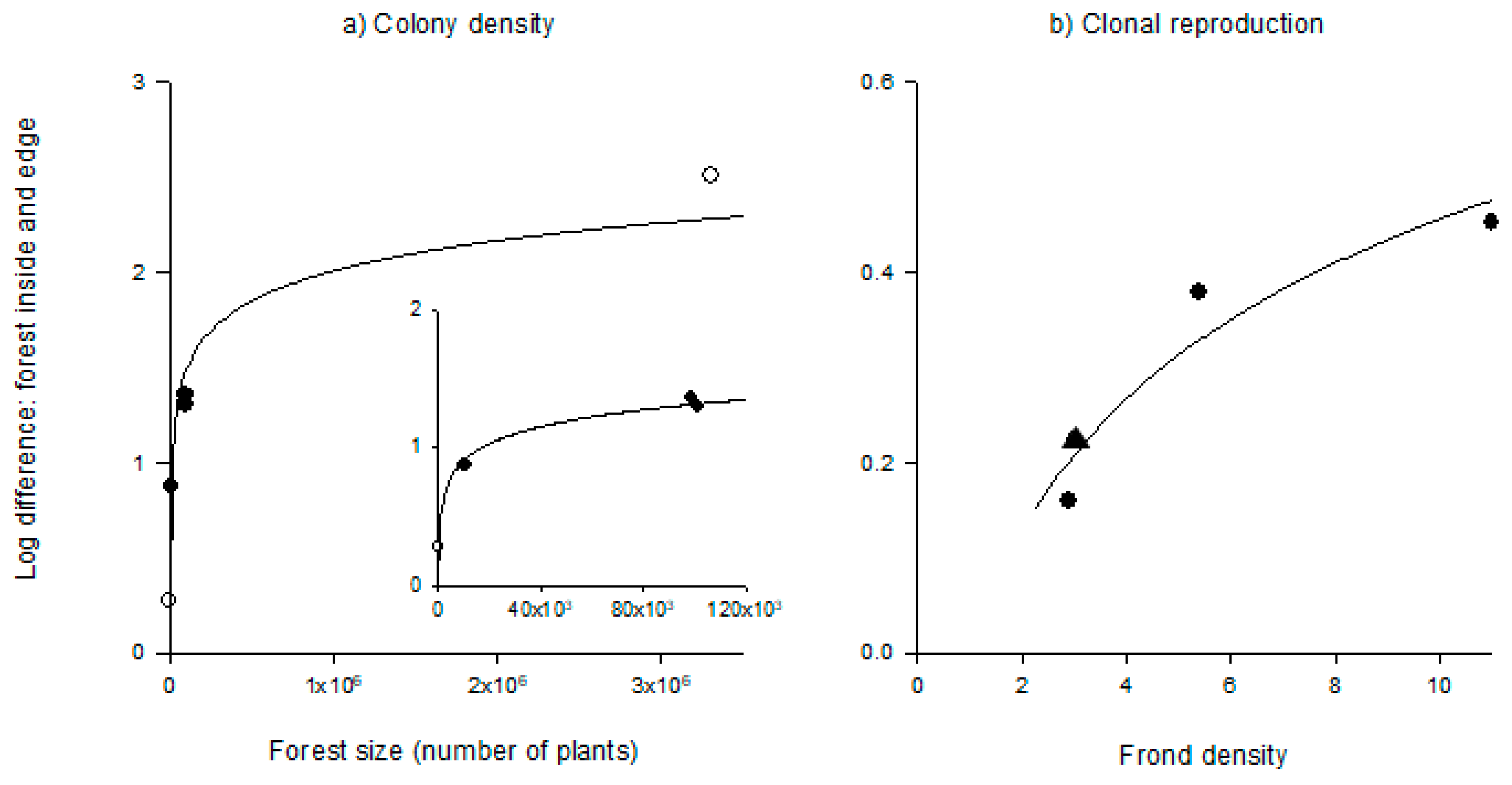
3.5. Relationships Between Kelp Forest Structure and Membranipora Demography
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Bernstein, B.B.; Jung, N. Selective Pressures and Coevolution in a Kelp Canopy Community in Southern California. Ecol. Monogr. 1979, 49, 335–355. [Google Scholar] [CrossRef]
- Dayton, P.K. Ecology of Kelp Communities. Annu. Rev. Ecol. Syst. 1985, 16, 215–245. [Google Scholar] [CrossRef]
- Irlandi, E.A.; Peterson, C.H. Modification of animal habitat by large plants: Mechanisms by which seagrasses influence clam growth. Oecologia 1991, 87, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Hixon, M.A.; Beets, J.P. Predation, Prey Refuges, and the Structure of Coral-Reef Fish Assemblages. Ecol. Monogr. 1993, 63, 77–101. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Positive and Negative Effects of Organisms as Physical Ecosystem Engineers. Ecology 1997, 78, 1946–1957. [Google Scholar] [CrossRef]
- Crain, C.M.; Bertness, M.D. Community Impacts of a Tussock Sedge: Is Ecosystem Engineering Important in Benign Habitats? Ecology 2005, 86, 2695–2704. [Google Scholar] [CrossRef]
- Boström, C.; Jackson, E.L.; Simenstad, C.A. Seagrass landscapes and their effects on associated fauna: A review. Estuar. Coast. Shelf Sci. 2006, 68, 383–403. [Google Scholar] [CrossRef]
- Winkler, M.; Hülber, K.; Hietz, P. Population dynamics of epiphytic bromeliads: Life strategies and the role of host branches. Basic Appl. Ecol. 2007, 8, 183–196. [Google Scholar] [CrossRef]
- Larkum, A.W.D.; Orth, R.J.; Duarte, C. (Eds.) Seagrasses: Biology, Ecology and Conservation; Springer: Dordrecht, The Netherlands, 2006; ISBN 978-1-4020-2942-4. [Google Scholar]
- Jones, C.; Lawton, J.H.; Shachak, M. Organisms as Ecosystem Engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Pearson, L.C. Influence of Temperature and Humidity on Distribution of Lichens in a Minnesota Bog. Ecology 1969, 50, 740–746. [Google Scholar] [CrossRef]
- Esseen, P.-A.; Renhorn, K.-E. Edge Effects on an Epiphytic Lichen in Fragmented Forests. Conserv. Biol. 1998, 12, 1307–1317. [Google Scholar] [CrossRef]
- Coote, L.; Smith, G.; Kelly, D.; O’Donoghue, S.; Dowding, P.; Iremonger, S.; Mitchell, F. Epiphytes of Sitka spruce (Picea sitchensis) plantations in Ireland and the effects of open spaces. Biodivers. Conserv. 2008, 17, 953–968. [Google Scholar] [CrossRef]
- Saunders, J.E.; Attrill, M.J.; Shaw, S.M.; Rowden, A.A. Spatial variability in the epiphytic algal assemblages of Zostera marina seagrass beds. Mar. Ecol. Prog. Ser. 2003, 249, 107–115. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Lavery, P.S.; van Keulen, M. Epiphytes of Seagrasses. In Seagrasses: Biology, Ecologyand Conservation; Springer: Dordrecht, The Netherlands, 2007; pp. 441–461. [Google Scholar]
- Gutiérrez, J.L.; Jones, C.G.; Byers, J.E.; Arkema, K.K.; Berkenbusch, K.; Commito, J.A.; Duarte, C.M.; Hacker, S.D.; Lambrinos, J.G.; Hendriks, I.E.; et al. Physical Ecosystem Engineers and the Functioning of Estuaries and Coasts. In Treatise on Estuarine and Coastal Science; Elsevier: Amsterdam, The Netherlands, 2011; pp. 53–81. ISBN 978-0-08-087885-0. [Google Scholar]
- Graham, M.H.; Vásquez, J.A.; Buschmann, A.H. Global ecology of the giant kelp Macrocystis: From ecotypes to ecosystems. Oceanogr. Mar. Biol. 2007, 45, 39–88. [Google Scholar]
- Krumhansl, K.A.; Okamoto, D.K.; Rassweiler, A.; Novak, M.; Bolton, J.J.; Cavanaugh, K.C.; Connell, S.D.; Johnson, C.R.; Konar, B.; Ling, S.D.; et al. Global patterns of kelp forest change over the past half-century. Proc. Natl. Acad. Sci. USA 2016, 113, 13785–13790. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.; Schiel, R. The ecology of giant kelp forests in California: A community profile. US Fish Wildl. Serv. Biol. Rep. 1985, 85. [Google Scholar]
- Graham, M.H. Effects of Local Deforestation on the Diversity and Structure of Southern California Giant Kelp Forest Food Webs. Ecosystems 2004, 7, 341–357. [Google Scholar] [CrossRef]
- Miller, R.J.; Lafferty, K.D.; Lamy, T.; Kui, L.; Rassweiler, A.; Reed, D.C. Giant kelp, Macrocystis pyrifera, increases faunal diversity through physical engineering. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172571. [Google Scholar] [CrossRef] [PubMed]
- Teagle, H.; Hawkins, S.J.; Moore, P.J.; Smale, D.A. The role of kelp species as biogenic habitat formers in coastal marine ecosystems. J. Exp. Mar. Biol. Ecol. 2017, 492, 81–98. [Google Scholar] [CrossRef]
- Jackson, G.A.; Winant, C.D. Effect of a kelp forest on coastal currents. Cont. Shelf Res. 1983, 2, 75–80. [Google Scholar] [CrossRef]
- Gaylord, B.; Rosman, J.H.; Reed, D.C.; Koseff, J.R.; Fram, J.; MacIntyre, S.; Arkema, K.; McDonald, C.; Brzezinski, M.A.; Largier, J.L.; et al. Spatial patterns of flow and their modification within and around a giant kelp forest. Limnol. Oceanogr. 2007, 52, 1838–1852. [Google Scholar] [CrossRef]
- Gaylord, B.; Nickols, K.J.; Jurgens, L. Roles of transport and mixing processes in kelp forest ecology. J. Exp. Biol. 2012, 215, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Rosman, J.H.; Koseff, J.R.; Monismith, S.G.; Grover, J. A field investigation into the effects of a kelp forest (Macrocystis pyrifera) on coastal hydrodynamics and transport. J. Geophys. Res. Oceans 2007, 112. [Google Scholar] [CrossRef]
- Bray, R. Influence of water currents and zooplankton densities on daily foraging movements of blacksmith, Chromis punctipinnis, a planktivorous reef fish. Fish. Bull. 1980, 78, 829–841. [Google Scholar]
- Arkema, K.K.; Reed, D.C.; Schroeter, S.C. Direct and indirect effects of giant kelp determine benthic community structure and dynamics. Ecology 2009, 90, 3126–3137. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.J.; Page, H.M.; Reed, D.C. Trophic versus structural effects of a marine foundation species, giant kelp (Macrocystis pyrifera). Oecologia 2015, 179, 1199–1209. [Google Scholar] [CrossRef]
- Eckman, J.E.; Duggins, D.O. Life and death beneath macrophyte canopies: Effects of understory kelps on growth rates and survival of marine, benthic suspension feeders. Oecologia 1991, 87, 473–487. [Google Scholar] [CrossRef]
- Stewart, H.; Fram, J.; Reed, D.; Williams, S.; Brzezinski, M.; Macintyre, S.; Gaylord, B. Differences in growth, morphology and tissue carbon and nitrogen of Macrocystis pyrifera within and at the outer edge of a giant kelp forest in California, USA. Mar. Ecol. Prog. Ser. 2009, 375, 101–112. [Google Scholar] [CrossRef]
- Bell, T.W.; Cavanaugh, K.C.; Siegel, D.A. Remote monitoring of giant kelp biomass and physiological condition: An evaluation of the potential for the Hyperspectral Infrared Imager (HyspIRI) mission. Remote Sens. Environ. 2015, 167, 218–228. [Google Scholar] [CrossRef]
- Saunders, M.; Metaxas, A. High recruitment of the introduced bryozoan Membranipora membranacea is associated with kelp bed defoliation in Nova Scotia, Canada. Mar. Ecol. Prog. Ser. 2008, 369, 139–151. [Google Scholar] [CrossRef]
- Scheibling, R.E.; Gagnon, P. Temperature-mediated outbreak dynamics of the invasive bryozoan Membranipora membranacea in Nova Scotian kelp beds. Mar. Ecol. Prog. Ser. 2009, 390, 1–13. [Google Scholar] [CrossRef]
- Krumhansl, K.A.; Lee, J.M.; Scheibling, R.E. Grazing damage and encrustation by an invasive bryozoan reduce the ability of kelps to withstand breakage by waves. J. Exp. Mar. Biol. Ecol. 2011, 407, 12–18. [Google Scholar] [CrossRef]
- Dixon, J.; Schroeter, S.C.; Kastendiek, J. Effects of the Encrusting Bryozoan, Membranipora membranacea, on the Loss of Blades and Fronds by the Giant Kelp, Macrocystis pyrifera (Laminariales)1. J. Phycol. 1981, 17, 341–345. [Google Scholar] [CrossRef]
- Watanabe, S.; Scheibling, R.E.; Metaxas, A. Contrasting patterns of spread in interacting invasive species: Membranipora membranacea and Codium fragile off Nova Scotia. Biol. Invasions 2010, 12, 2329–2342. [Google Scholar] [CrossRef]
- Levin, P.S.; Coyer, J.A.; Petrik, R.; Good, T.P. Community-Wide Effects of Nonindigenous Species on Temperate Rocky Reefs. Ecology 2002, 83, 3182–3193. [Google Scholar] [CrossRef]
- Hepburn, C.D.; Hurd, C.L.; Frew, R.D. Colony Structure and Seasonal Differences in Light and Nitrogen Modify the Impact of Sessile Epifauna on the Giant Kelp Macrocystis pyrifera (L.) C Agardh. Hydrobiologia 2006, 560, 373–384. [Google Scholar] [CrossRef]
- Arkema, K.K. Flow-mediated feeding in the field: Consequences for the performance and abundance of a sessile marine invertebrate. Mar. Ecol. Prog. Ser. 2009, 388, 207–220. [Google Scholar] [CrossRef]
- Arkema, K.K. Consequences of Kelp Forest Structure and Dynamics for Epiphytes and Understory Communities. Ph.D. Thesis, University of California, Santa Barbara, CA, USA, 2008. [Google Scholar]
- Marzinelli, E.M.; Underwood, A.J.; Coleman, R.A. Modified Habitats Influence Kelp Epibiota via Direct and Indirect Effects. PLoS ONE 2011, 6, e21936. [Google Scholar] [CrossRef]
- Marzinelli, E.M.; Underwood, A.J.; Coleman, R.A. Modified habitats change ecological processes affecting a non-indigenous epibiont. Mar. Ecol. Prog. Ser. 2012, 446, 119–129. [Google Scholar] [CrossRef][Green Version]
- Yoshioka, P.M. Role of Planktonic and Benthic Factors in the Population Dynamics of the Bryozoan Membranipora membranacea. Ecology 1982, 63, 457–468. [Google Scholar] [CrossRef]
- Schwaninger, H.R. Population structure of the widely dispersing marine bryozoan Membranipora membranacea (Cheilostomata): Implications for population history, biogeography, and taxonomy. Mar. Biol. 1999, 135, 411–423. [Google Scholar] [CrossRef]
- Dick, M.H.; Grischenko, A.V.; Mawatari, S.F. Intertidal Bryozoa (Cheilostomata) of Ketchikan, Alaska. J. Nat. Hist. 2005, 39, 3687–3784. [Google Scholar] [CrossRef]
- Temkin, M.H. Gamete spawning and fertilization in the gymnolaemate bryozoan Membranipora membranacea. Biol. Bull. 1994, 187, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.C.; Rassweiler, A.; Arkema, K.K. Biomass rather than growth rate determines variation in net primary production by giant kelp. Ecology 2008, 89, 2493–2505. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.C.; Rassweiler, A.R.; Miller, R.J.; Page, H.M.; Holbrook, S.J. The value of a broad temporal and spatial perspective in understanding dynamics of kelp forest ecosystems. Mar. Freshw. Res. 2016. [Google Scholar] [CrossRef]
- Cavanaugh, K.; Siegel, D.; Kinlan, B.; Reed, D. Scaling giant kelp field measurements to regional scales using satellite observations. Mar. Ecol. Prog. Ser. 2010, 403, 13–27. [Google Scholar] [CrossRef]
- Rodriguez, G.E.; Rassweiler, A.; Reed, D.C.; Holbrook, S.J. The importance of progressive senescence in the biomass dynamics of giant kelp (Macrocystis pyrifera). Ecology 2013, 94, 1848–1858. [Google Scholar] [CrossRef]
- Rodriguez, G.E.; Reed, D.C.; Holbrook, S.J. Blade life span, structural investment, and nutrient allocation in giant kelp. Oecologia 2016, 182, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: A Practical Information-Theoretic Approach; Springer-Verlag: New York, NY, USA, 1998; ISBN 978-1-4757-2917-7. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Hermansen, P.; Larsen, P.S.; Riisgård, H.U. Colony growth rate of encrusting marine bryozoans (Electra pilosa and Celleporella hyalina). J. Exp. Mar. Biol. Ecol. 2001, 263, 1–23. [Google Scholar] [CrossRef]
- Dayton, P.K.; Tegner, M.J. Catastrophic storms, el nino, and patch stability in a southern california kelp community. Science 1984, 224, 283–285. [Google Scholar] [CrossRef] [PubMed]
- Hepburn, C.D.; Hurd, C.L. Conditional mutualism between the giant kelp Macrocystis pyrifera and colonial epifauna. Mar. Ecol. Prog. Ser. 2005, 302, 37–48. [Google Scholar] [CrossRef]
- Hepburn, C.; Frew, R.; Hurd, C. Uptake and transport of nitrogen derived from sessile epifauna in the giant kelp Macrocystis pyrifera. Aquat. Biol. 2012, 14, 121–128. [Google Scholar] [CrossRef]
- Bologna, P.; Heck, K. Impacts of Seagrass Habitat Architecture on Bivalve Settlement. Estuaries Coasts 2012, 23, 449–457. [Google Scholar] [CrossRef]
- Saunders, M.; Metaxas, A. Effects of temperature, size, and food on the growth of Membranipora membranacea in laboratory and field studies. Mar. Biol. 2009, 156, 2267–2276. [Google Scholar] [CrossRef]
- Pratt, M.C. Living where the flow is right: How flow affects feeding in bryozoans. Integr. Comp. Biol. 2008, 48, 808–822. [Google Scholar] [CrossRef]
- Williams, S.L.; Heck, K.L. Seagrass community ecology. In Marine Community Ecology; Sinauer Associates: Sunderland, MA, USA, 2001; pp. 317–337. [Google Scholar]
- Witman, J.D.; Dayton, P.K. Rocky subtidal communities. In Marine Community Ecology; Sinauer Associates: Sunderland, MA, USA, 2001; pp. 339–366. [Google Scholar]
- Wildish, D.; Kristmanson, D. Benthic Suspension Feeders and Flow; Cambridge University Press: Cambridge, UK, 2005; ISBN 978-0-521-02347-4. [Google Scholar]
- Buss, L.W.; Jackson, J.B.C. Planktonic food availability and suspension-feeder abundance: Evidence of in situ depletion. J. Exp. Mar. Biol. Ecol. 1981, 49, 151–161. [Google Scholar] [CrossRef]
- Peterson, C.H.; Black, R. Resource depletion by active suspension feeders on tidal fiats: Influence of local density and tidal elevation1. Limnol. Oceanogr. 1987, 32, 143–166. [Google Scholar] [CrossRef]
- Eckman, J.E.; Duggins, D.O. Effects of Flow Speed on Growth of Benthic Suspension Feeders. Biol. Bull. 1993, 185, 28–41. [Google Scholar] [CrossRef]
- Gutiérrez, J.L.; Iribarne, O.O. Conditional responses of organisms to habitat structure: An example from intertidal mudflats. Oecologia 2004, 139, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.C.; Rassweiler, A.; Carr, M.H.; Cavanaugh, K.C.; Malone, D.P.; Siegel, D.A. Wave disturbance overwhelms top-down and bottom-up control of primary production in California kelp forests. Ecology 2011, 92, 2108–2116. [Google Scholar] [CrossRef] [PubMed]
- Samhouri, J.F.; Levin, P.S.; Ainsworth, C.H. Identifying Thresholds for Ecosystem-Based Management. PLoS ONE 2010, 5, e8907. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arkema, K.K.; Samhouri, J.F. Living on the Edge: Variation in the Abundance and Demography of a Kelp Forest Epibiont. Diversity 2019, 11, 120. https://doi.org/10.3390/d11080120
Arkema KK, Samhouri JF. Living on the Edge: Variation in the Abundance and Demography of a Kelp Forest Epibiont. Diversity. 2019; 11(8):120. https://doi.org/10.3390/d11080120
Chicago/Turabian StyleArkema, Katie K., and Jameal F. Samhouri. 2019. "Living on the Edge: Variation in the Abundance and Demography of a Kelp Forest Epibiont" Diversity 11, no. 8: 120. https://doi.org/10.3390/d11080120
APA StyleArkema, K. K., & Samhouri, J. F. (2019). Living on the Edge: Variation in the Abundance and Demography of a Kelp Forest Epibiont. Diversity, 11(8), 120. https://doi.org/10.3390/d11080120