Spatial Response to Linear Infrastructures by the Endangered Golden Lion Tamarin

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Study Species
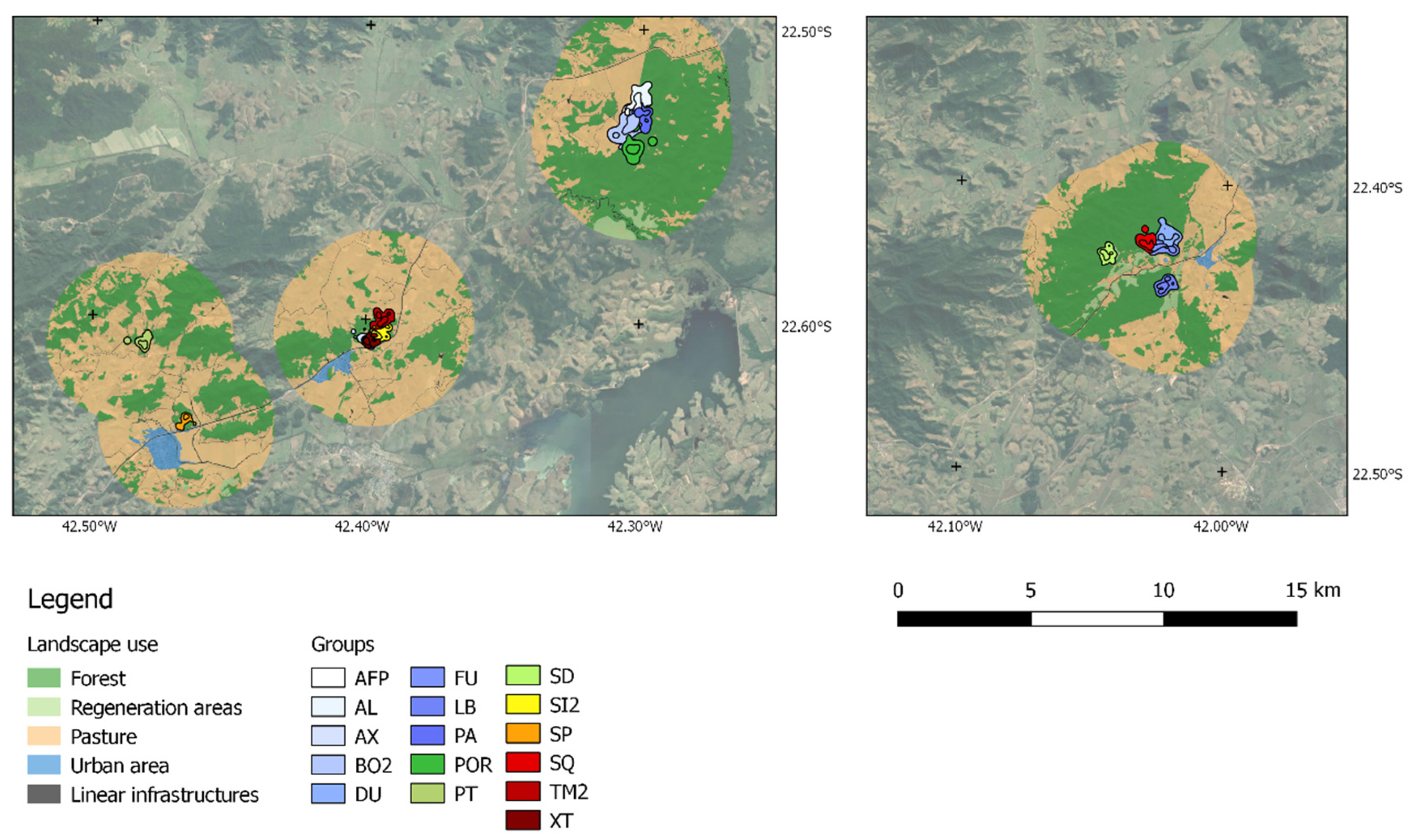
2.2. Study Area
2.3. Golden Lion Tamarin Capture and Radio Telemetry
2.4. Spatial Analysis
2.5. Explanatory Variables and Models
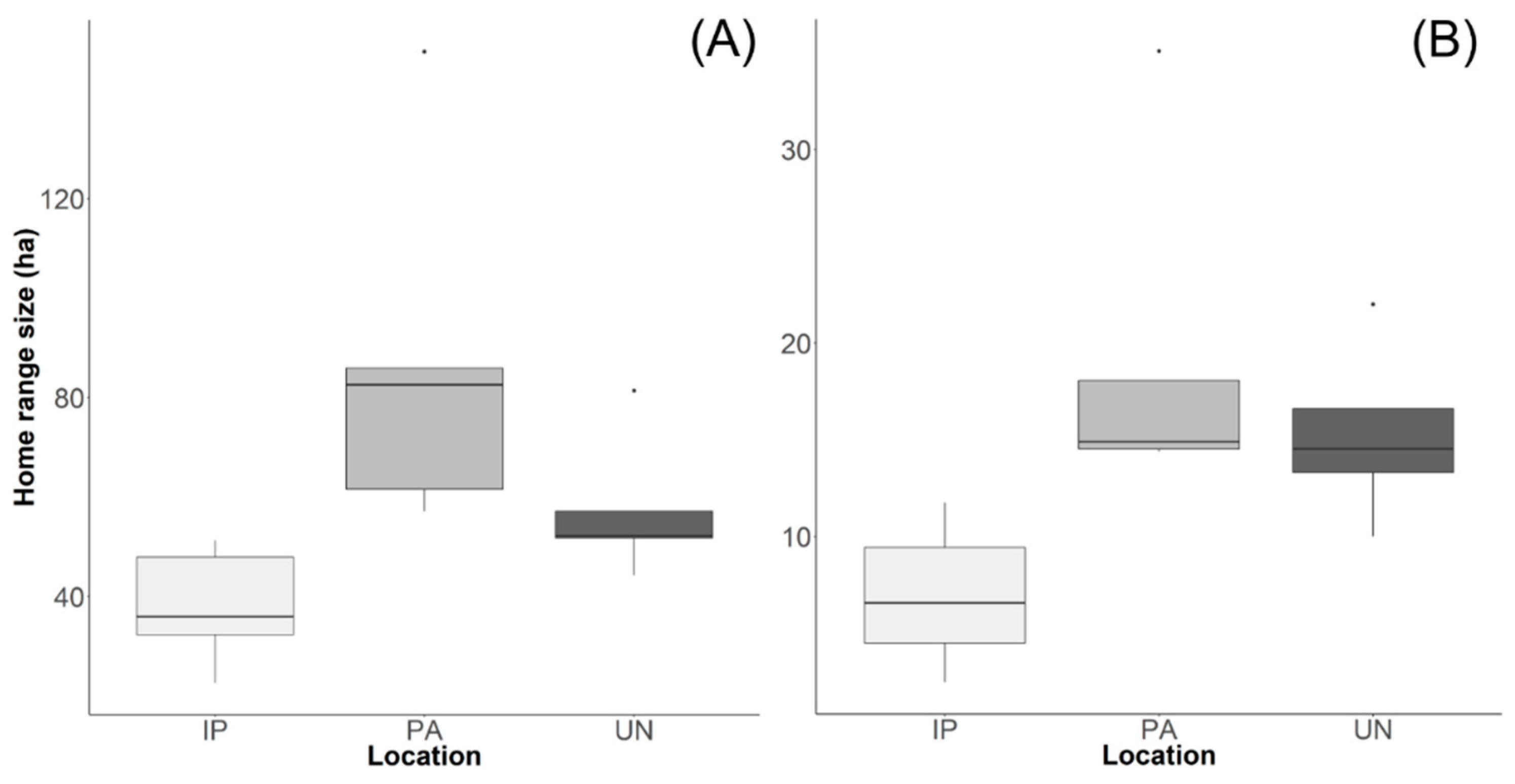
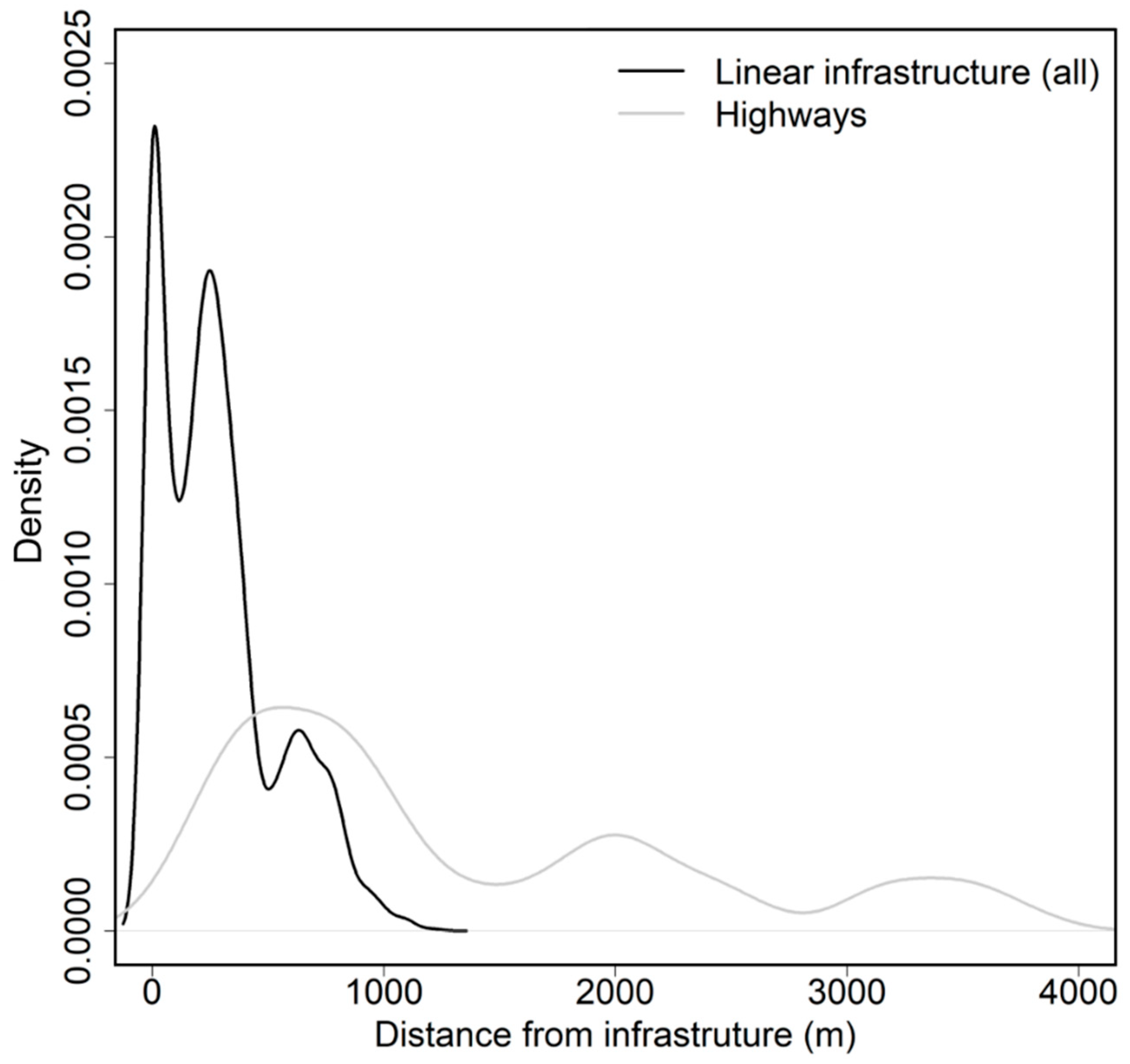
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Laurance, W.F.; Cochrane, M.A.; Bergen, S.; Fearnside, P.M.; Delamônica, P.; Barber, C.; D’Angelo, S.; Fernandes, T. The future of the Brazilian Amazon. Science 2001, 291, 438–439. [Google Scholar] [CrossRef] [PubMed]
- Geneletti, D. Some common shortcomings in the treatment of impacts of linear infrastructures on natural habitat. Environ. Impact Asses. 2006, 26, 257–267. [Google Scholar] [CrossRef]
- Perz, S.G.; Overdevest, C.; Caldas, M.M.; Walker, R.T.; Arima, E.Y. Unofficial road building in the Brazilian Amazon: Dilemmas and models for road governance. Environ. Conserv. 2007, 34, 112–121. [Google Scholar] [CrossRef]
- Roberts, K.; Sjölund, A. Incorporating biodiversity issues into road design: The road agency perspective. In Handbook of Road Ecology; van der Ree, R., Smith, D.J., Grilo, C., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 27–31. [Google Scholar]
- Fahrig, L.; Rytwinski, T. Effects of roads on animal abundance: An empirical review and synthesis. Ecol. Soc. 2009, 14, 21. Available online: http://www.ecologyandsociety.org/vol14/iss1/art21/ (accessed on 25 February 2019). [CrossRef]
- Da Rosa, C.A.; Bager, A. Review of the factors underlying the mechanisms and effects of roads on vertebrates. Oecol. Aust. 2013, 17, 208–221. [Google Scholar]
- Dorsey, B.; Olsson, M.; Rew, L.J. Ecological effects of railways on wildlife. In Handbook of Road Ecology; van der Ree, R., Smith, D.J., Grilo, C., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 219–227. [Google Scholar]
- Roy, M.; Sukumar, R. Railways and wildlife: A case study of train-elephant collisions in northern West Bengal, India. In Railway Ecology; Borda-de-Água, L., Barrientos, R., Beja, P., Pereira, H., Eds.; Springer: Cham, Switzerland, 2017; pp. 157–177. [Google Scholar]
- Santos, S.M.; Carvalho, F.; Mira, A. Current knowledge on wildlife mortality in railways. In Railway Ecology; Borda-de-Água, L., Barrientos, R., Beja, P., Pereira, H., Eds.; Springer: Cham, Switzerland, 2017; pp. 11–22. [Google Scholar]
- Barrientos, R.; Ascensão, F.; Beja, P.; Pereira, H.M.; Borda-de-Água, L. Railway ecology vs. road ecology: Similarities and differences. Eur. J. Wildlife Res. 2019, 65, 12. [Google Scholar] [CrossRef]
- Malo, J.E.; Suárez, F.; Díez, A. Can we mitigate animal–vehicle accidents using predictive models? J. Appl. Ecol. 2004, 41, 701–710. [Google Scholar] [CrossRef]
- Taylor, B.D.; Goldingay, R.L. Wildlife road-kills on three major roads in north-eastern New South Wales. Wildlife Res. 2004, 31, 83–91. [Google Scholar] [CrossRef]
- Olson, K.A.; van der Ree, R. Railways, Roads and Fences Across Kazakhstan and Mongolia Threaten the Survival of Wide-Ranging Wildlife. In Handbook of Road Ecology; van der Ree, R., Smith, D.J., Grilo, C., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 472–478. [Google Scholar]
- Lokschin, L.X.; Rodrigo, C.P.; Cabral, J.N.H.; Buss, G. Power lines and howler monkey conservation in Porto Alegre, Rio Grande do Sul, Brazil. Neotrop. Primates 2007, 14, 76–81. [Google Scholar] [CrossRef]
- Katsis, L.; Cunneyworth, P.M.; Turner, K.M.; Presotto, A. Spatial patterns of primate electrocutions in Diani, Kenya. Int. J. Primatol. 2018, 39, 493–510. [Google Scholar] [CrossRef]
- Barrientos, R.; Borda-de-Água, L. Railways as Barriers for Wildlife: Current Knowledge. In Railway Ecology; Borda-de-Água, L., Barrientos, R., Beja, P., Pereira, H., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Goosem, M.; Harding, E.K.; Chester, G.; Tucker, N.; Harriss, C.; Oakley, K. Roads in Rainforest: Best Practice Guidelines for Planning, Design and Management. 2010. Available online: https://researchonline.jcu.edu.au/12113/1/goosem_guidelines.pdf (accessed on 7 June 2019).
- Ascensão, F.; Lucas, P.S.; Costa, A.; Bager, A. The effect of roads on edge permeability and movement patterns for small mammals: A case study with Montane Akodont. Landsc. Ecol. 2017, 32, 781–790. [Google Scholar] [CrossRef]
- Jaeger, J.A.; Bowman, J.; Brennan, J.; Fahrig, L.; Bert, D.; Bouchard, J.; Charbonneau, N.; Frank, K.; Gruber, B.; von Toschanowitz, K.T. Predicting when animal populations are at risk from roads: An interactive model of road avoidance behavior. Ecol. Modell. 2005, 185, 329–348. [Google Scholar] [CrossRef]
- Lucas, P.S.; de Carvalho, R.G.; Grilo, C. Railway disturbances on wildlife: Types, effects, and mitigation measures. In Railway Ecology; Borda-de-Água, L., Barrientos, R., Beja, P., Pereira, H., Eds.; Springer: Cham, Switzerland, 2017; pp. 81–99. [Google Scholar]
- Forman, R.T.; Deblinger, R.D. The ecological road-effect zone of a Massachusetts (USA) suburban highway. Biol. Conserv. 2000, 14, 36–46. [Google Scholar] [CrossRef]
- Eigenbrod, F.; Hecnar, S.J.; Fahrig, L. Quantifying the road-effect zone: Threshold effects of a motorway on anuran populations in Ontario, Canada. Ecol Soc. 2009, 14, 24. Available online: https://eprints.soton.ac.uk/id/eprint/181359 (accessed on 2 March 2019). [CrossRef]
- Benítez-López, A.; Alkemade, R.; Verweij, P.A. The impacts of roads and other infrastructure on mammal and bird populations: A meta-analysis. Biol. Conserv. 2010, 143, 1307–1316. [Google Scholar] [CrossRef]
- Clauzel, C.; Xiqing, D.; Gongsheng, W.; Giraudoux, P.; Li, L. Assessing the impact of road developments on connectivity across multiple scales: Application to Yunnan snub-nosed monkey conservation. Biol. Conserv. 2015, 192, 207–217. [Google Scholar] [CrossRef]
- van der Ree, R.; Smith, D.J.; Grilo, C. The ecological effects of linear infrastructure and traffic: Challenges and opportunities of rapid global growth. In Handbook of Road Ecology; van der Ree, R., Smith, D.J., Grilo, C., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 1–9. [Google Scholar]
- Ceia-Hasse, A.; Navarro, L.M.; Borda-de-Água, L.; Pereira, H.M. Population persistence in landscapes fragmented by roads: Disentangling isolation, mortality, and the effect of dispersal. Ecol. Model. 2018, 375, 45–53. [Google Scholar] [CrossRef]
- Laurance, W.F.; Goosem, M.; Laurance, S.G. Impacts of roads and linear clearings on tropical forests. Trends Ecol. Evol. 2009, 24, 659–669. [Google Scholar] [CrossRef]
- Laurance, W.F. Bad roads, good roads. In Handbook of Road Ecology; van der Ree, R., Smith, D.J., Grilo, C., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 10–15. [Google Scholar]
- Soanes, K.; van der Ree, R. Reducing road impacts on tree-dwelling animals. In Handbook of Road Ecology; van der Ree, R., Smith, D.J., Grilo, C., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 334–340. [Google Scholar]
- Laurance, W.F.; Croes, B.M.; Tchignoumba, L.; Lahm, S.A.; Alonso, A.; Lee, M.E.; Campbell, P.; Ondzeano, C. Impacts of roads and hunting on central African rainforest mammals. Conserv. Biol. 2006, 20, 1251–1261. [Google Scholar] [CrossRef]
- Bryson-Morrison, N.; Tzanopoulos, J.; Matsuzawa, T.; Humle, T. Activity and habitat use of chimpanzees (Pan troglodytes verus) in the anthropogenic landscape of Bossou, Guinea, West Africa. Int. J. Primatol. 2017, 38, 282–302. [Google Scholar] [CrossRef]
- Decker, B.S. Endangered primates in the Selous Game Reserve and an imminent threat to their habitat. Oryx 1994, 28, 183–190. [Google Scholar] [CrossRef]
- Donaldson, A.; Cunneyworth, P. Case Study: Canopy bridges for primate conservation. In Handbook of Road Ecology; van der Ree, R., Smith, D.J., Grilo, C., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 341–343. [Google Scholar]
- Kioko, J.; Kiffner, C.; Jenkins, N.; Collinson, W.J. Wildlife roadkill patterns on a major highway in northern Tanzania. Afr. Zool. 2015, 50, 17–22. [Google Scholar] [CrossRef]
- Asensio, N.; Murillo-Chacon, E.; Schaffner, C.M.; Aureli, F. The effect of roads on spider monkeys’ home range and mobility in a heterogeneous regenerating forest. Biotropica 2017, 49, 546–554. [Google Scholar] [CrossRef]
- Coimbra-Filho, A.F.; Mittermeier, R.A. Distribution and ecology of the genus Leontopithecus Lesson, 1840 in Brazil. Primates 1973, 14, 47–66. [Google Scholar] [CrossRef]
- Kleiman, D.G.; Beck, B.B.; Dietz, J.M.; Dietz, L.A.; Ballou, J.D.; Coimbra-Filho, A.F. Conservation program for the golden lion tamarin: Captive research and management, ecological studies, educational strategies, and reintroduction. In Primates: The Road to Self-Sustaining Populations; Benirschke, K., Ed.; Springer: New York, NY, USA, 1986; pp. 960–979. [Google Scholar]
- Kierulff, M.C.; Rylands, A.B.; Procópio-de-Oliveira, M.M. Leontopithecus rosalia. The IUCN Red List of Threatened Species 2008: E T11506A3287321. Available online: https://www.iucnredlist.org/species/11506/3287321 (accessed on 7 January 2019).
- Dietz, J.M.; Baker, A.J.; Miglioretti, D. Seasonal variation in reproduction, juvenile growth, and adult body mass in golden lion tamarins (Leontopithecus rosalia). Am. J. Primatol. 1994, 34, 115–132. [Google Scholar] [CrossRef]
- Baker, A.J.; Bales, K.; Dietz, J.M. Mating system and group dynamics in Lion Tamarins. In Lion Tamarins: Biology and Conservation; Kleiman, D.G., Rylands, A.B., Eds.; Smithsonian Institution Press: Washington DC, USA, 2002; pp. 188–212. [Google Scholar]
- Stevenson, M.F.; Rylands, A.B. The marmosets, genus Callithrix. In Ecology end Behavior of Neotropical Primates; Mittermier, R.A., Rylands, A.B., Coimbra-Filho, A., Fonseca, G.A.B., Eds.; World Wildlife Fund: Washington, DC, USA, 1988; pp. 131–222. [Google Scholar]
- Dietz, J.M.; Baker, A.J. Polygyny and female reproductive success in golden lion tamarins, Leontopithecus rosalia. Anim. Behav. 1993, 46, 1067–1078. [Google Scholar] [CrossRef]
- Romano, V.; Martins, A.F.; Ruiz-Miranda, C.R. Unraveling the dispersal patterns and the social drivers of natal emigration of a cooperative breeding mammal, the golden lion tamarin. Am. J. Primatol. 2019, 81, e22959. [Google Scholar] [CrossRef] [PubMed]
- Rylands, A.B.; Faria, D.S. Habitats, feeding, and home range size in the genus Callithrix. In Marmosets and Tamarins (Systematics, Behaviour, and Ecology); Rylands, A.B., Ed.; Oxford Univ. Press: New York, NY, USA, 1993; p. 396. [Google Scholar]
- Peres, C.A. Costs and benefits of territorial defense in wild golden lion tamarins, Leontopithecus rosalia. Behav. Ecol. Sociobiol. 1989, 25, 227–233. [Google Scholar] [CrossRef]
- Dietz, J.M.; Peres, C.A.; Pinder, L. Foraging ecology and use of space in wild golden lion tamarins (Leontopithecus rosalia). Am. J. Primatol. 1997, 41, 289–305. [Google Scholar] [CrossRef]
- Miller, K.E.; Dietz, J.M. Effects of individual and group characteristics on feeding behaviors in Leontopithecus rosalia. Int. J. Primatol. 2005, 26, 1291–1319. [Google Scholar] [CrossRef]
- Coelho, A.S.; Ruiz Miranda, C.R.; Beck, B.B.; Martins Oliveira, A.C.R.; Sabatini, V. Comportamento do mico-leão-dourado (Leontopithecus rosalia, Linnaeus 1766) em relação a fragmentação do habitat. In Conservação do Mico-Leão-Dourado: Enfrentando os Desafios de uma Paisagem Fragmentada; Oliveira, P.P., Grativol, A.D., Ruiz Miranda, C.R., Eds.; Campos dos Goytacazes, Uniuversidade Estadual do Norte Fluminense: Rio de Janeiro, Brazil, 2008; pp. 58–85. [Google Scholar]
- Lapenta, M.J.; Procópio-de-Oliveira, P. Some aspects of seed dispersal effectiveness of golden lion tamarins (Leontopithecus rosalia) in a Brazilian Atlantic forest. Trop. Conserv. Sci. 2008, 1, 122–139. [Google Scholar] [CrossRef]
- Lima, H.C.; Pessoa, S.V.A.; Guedes-Bruni, R.R.; Moraes, L.F.D.; Granzotto, S.V.; Iwamoto, S.; Ciero, J.D. Caracterização fisionômico-florística e mapeamento da vegetação da Reserva Biológica de Poço das Antas, Silva Jardim, Rio de Janeiro, Brasil. Rodriguésia 2006, 53, 369–389. [Google Scholar] [CrossRef]
- ICMBio—Instituto Chico Mendes de Conservação da Biodiversidade. Plano de Manejo da Área de Proteção Ambiental da Bacia do Rio São João/Mico-Leão-Dourado; Instituto Chico Mendes de Conservação da Biodiversidade: Brasília, Brazil, 2008.
- IBGE—Instituto Brasileiro de Geografia e Estatística. Manual Técnico da Vegetação Brasileira: Sistema Fitogeográfico, Inventário das Formações Florestais e Campestres, Técnicas e Manejo de Coleções Botânicas, Procedimentos para Mapeamentos, 2nd ed.; Coordenação de Recursos Naturais e Estudos Ambientais; IBGE: Rio de Janeiro, Brazil, 2012; p. 276.
- Köppen, W. Climatologia: Con un Estudio de los Climas de la Tierra; Fondo de Cultura Economica: Pánuco, Mexico, 1948; p. 478. [Google Scholar]
- SEMA. Secretaria de Estado de Meio Ambiente e Desenvolvimento Sustentável, Governo do Estado do Rio de Janeiro. In Atlas das Unidades de Conservação da Natureza do Estado do Rio de Janeiro; Metalivros: Rio de Janeiro, Brazil, 2001; p. 48. [Google Scholar]
- Valladares-Padua, C.; Cullen Junior, L.; Padua, S. A pole bridge to avoid primate road kills. Neotrop. Primates 1995, 3, 13–15. [Google Scholar]
- Torres, A.; Jaeger, J.A.G.; Alonso, J.C. Assessing large-scale wildlife responses to human infrastructure development. Proc. Natl. Acad. Sci. USA 2016, 113, 8472–8477. [Google Scholar] [CrossRef] [PubMed]
- Walkup, D.K.; Leavitt, D.J.; Fitzgerald, L.A. Effects of habitat fragmentation on population structure of dune-dwelling lizards. Ecosphere 2017, 8, e01729. [Google Scholar] [CrossRef]
- Procópio-de-Oliveira, P.; Grativol, A.; Ruiz-Miranda, C.R. Conservação do Mico-leão-dourado: Enfrentando os Desafios de uma Paisagem Fragmentada; Universidade Estadual do Norte Fluminense Darcy Ribeiro Press: Campos dos Goytacazes, Brazil, 2008. [Google Scholar]
- Baker, A.J.; Dietz, J.M.; Kleiman, D.G. Behavioural evidence for monopolization of paternity in multi-male groups of golden lion tamarins. Anim. Behav. 1993, 46, 1091–1103. [Google Scholar] [CrossRef]
- Calenge, C. The package “adehabitat” for the R software: A tool for the analysis of space and habitat use by animals. Ecol. Model. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; ISBN 3-900051-07-0. [Google Scholar]
- Worton, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2015. [Google Scholar]
- Turchin, P. Quantitative Analysis of Movement; Sinuaer Associates, Inc. Publishers: Sunderland, UK, 1998. [Google Scholar]
- Calenge, C. Analysis of Animal Movements in R: The adehabitatLT Package. 2016. Available online: https://cran.r-project.org/web/packages/adehabitatLT/vignettes/adehabitatLT.pdf (accessed on 7 June 2019).
- Grilo, C.; Sousa, J.; Ascensão, F.; Matos, H.; Leitão, I.; Pinheiro, P.; Costa, M.; Bernardo, J.; Reto, D.; Lourenço, R.; et al. Individual Spatial Responses towards Roads: Implications for Mortality Risk. PLoS ONE 2012, 7, e43811. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Interface: A Practical Information Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 1998. [Google Scholar]
- Raboy, B.E.; Neves, L.G.; Zeigler, S.; Saraiva, N.A.; Cardoso, N.; dos Santos, G.R.; Ballou, J.D.; Leimgruber, P. Strength of habitat and landscape metrics in predicting golden-headed lion tamarin presence or absence in forest patches in southern Bahia, Brazil. Biotropica 2010, 42, 388–397. [Google Scholar] [CrossRef]
- Moraes, A.M.; Ruiz-Miranda, C.R.; Galetti Jr, P.M.; Niebuhr, B.B.; Alexandre, B.R.; Muylaert, R.L.; Grativol, A.D.; Ribeiro, J.W.; Ferreira, A.N.; Ribeiro, M.C. Landscape resistance influences effective dispersal of endangered golden lion tamarins within the Atlantic Forest. Biol. Conserv. 2018, 224, 178–187. [Google Scholar] [CrossRef]
- Amorim, A.P. Avaliação dos Recursos Alimentares dos Micos Leões Dourados (Leontopithecus Rosalia, LINNAEUS, 1766) e sua Relação com Medidas Corporais: Uma Aplicação da Lógica Fuzzy; Universidade Estadual do Norte Fluminense Darcy Ribeiro: Rio de Janeiro, Brazil, 2007. [Google Scholar]
- Hankerson, S.J.; Franklin, S.P.; Dietz, J.M. Tree and forest characteristics influence sleeping site choice by golden lion tamarins. Am. J. Primatol. 2007, 69, 976–988. [Google Scholar] [CrossRef] [PubMed]
- Didham, R.K. The influence of edge effects and forest fragmentation on leaf litter invertebrates in central Amazonia. In Tropical Forest Remnants: Ecology, Management and Conservation of Fragmented Communities; Laurance, W.F., Bierregaard, J.R.R.O., Eds.; University of Chicago Press: Chicago, IL, USA, 1997; pp. 55–70. [Google Scholar]
- Eldegard, K.; Totland, Ø.; Moe, S.R. Edge effects on plant communities along power line clearings. J. Appl. Ecol. 2015, 52, 871–880. [Google Scholar] [CrossRef]
- Lewis, S.L. Tropical forests and the changing earth system. Philos. Trans. R Soc. B Biol. Sci. 2005, 361, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Malhi, Y. The productivity, metabolism and carbon cycle of tropical forest vegetation. J. Ecol. 2012, 100, 65–75. [Google Scholar] [CrossRef]
- Moraes, A.M.; Ruiz-Miranda, C.R.; Ribeiro, M.C.; Grativol, A.D.; Carvalho, C.D.S.; Dietz, J.M.; Kierulff, M.C.F.; Freitas, L.A.; Galetti, P.M. Temporal genetic dynamics of reintroduced and translocated populations of the endangered golden lion tamarin (Leontopithecus rosalia). Conserv. Genet. 2017, 18, 995–1009. [Google Scholar] [CrossRef]
- Holst, B.; Medici, E.; Marinho-Filho, O.J.; Kleiman, D.; Leus, K.; Pissinatti, A.; Vivekanda, G.; Ballou, J.; Traylor-Holzer, K.; Raboy, B.; et al. Lion Tamarin Population and Habitat Viability Assessment Workshop 2005, Final Report; IUCN/SSC Conservation Breeding Specialist Group: Apple Valley, MN, USA, 2006; p. 193. [Google Scholar]
- Soanes, K.; Lobo, M.C.; Vesk, P.A.; McCarthy, M.A.; Moore, J.L.; van der Ree, R. Movement re-established but not restored: Inferring the effectiveness of road-crossing mitigation for a gliding mammal by monitoring use. Biol. Conserv. 2013, 159, 434–441. [Google Scholar] [CrossRef]
- Teixeira, F.Z.; Printes, R.C.; Fagundes, J.C.G.; Alonso, A.C.; Kindel, A. Canopy bridges as road overpasses for wildlife in urban fragmented landscapes. Biota Neotrop. 2013, 13, 117–123. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Model | ΔAICc | Weight |
|---|---|---|
| step ~ inf + land | 0 | 0.2992 |
| step ~ dist_inf + inf + land | 0.1 | 0.2850 |
| step ~ dist_inf + inf | 0.3 | 0.2566 |
| step ~ land + dist_inf + inf + dist_hig | 1.3 | 0.1528 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucas, P.d.S.; Alves-Eigenheer, M.; Francisco, T.M.; Dietz, J.M.; Ruiz-Miranda, C.R. Spatial Response to Linear Infrastructures by the Endangered Golden Lion Tamarin. Diversity 2019, 11, 100. https://doi.org/10.3390/d11070100
Lucas PdS, Alves-Eigenheer M, Francisco TM, Dietz JM, Ruiz-Miranda CR. Spatial Response to Linear Infrastructures by the Endangered Golden Lion Tamarin. Diversity. 2019; 11(7):100. https://doi.org/10.3390/d11070100
Chicago/Turabian StyleLucas, Priscila da Silva, Milene Alves-Eigenheer, Talitha Mayumi Francisco, James M. Dietz, and Carlos Ramón Ruiz-Miranda. 2019. "Spatial Response to Linear Infrastructures by the Endangered Golden Lion Tamarin" Diversity 11, no. 7: 100. https://doi.org/10.3390/d11070100
APA StyleLucas, P. d. S., Alves-Eigenheer, M., Francisco, T. M., Dietz, J. M., & Ruiz-Miranda, C. R. (2019). Spatial Response to Linear Infrastructures by the Endangered Golden Lion Tamarin. Diversity, 11(7), 100. https://doi.org/10.3390/d11070100