A Comparison of the Population Genetic Structure and Diversity between a Common (Chrysemys p. picta) and an Endangered (Clemmys guttata) Freshwater Turtle

 ,
,
Abstract
1. Introduction
2. Materials and Methods
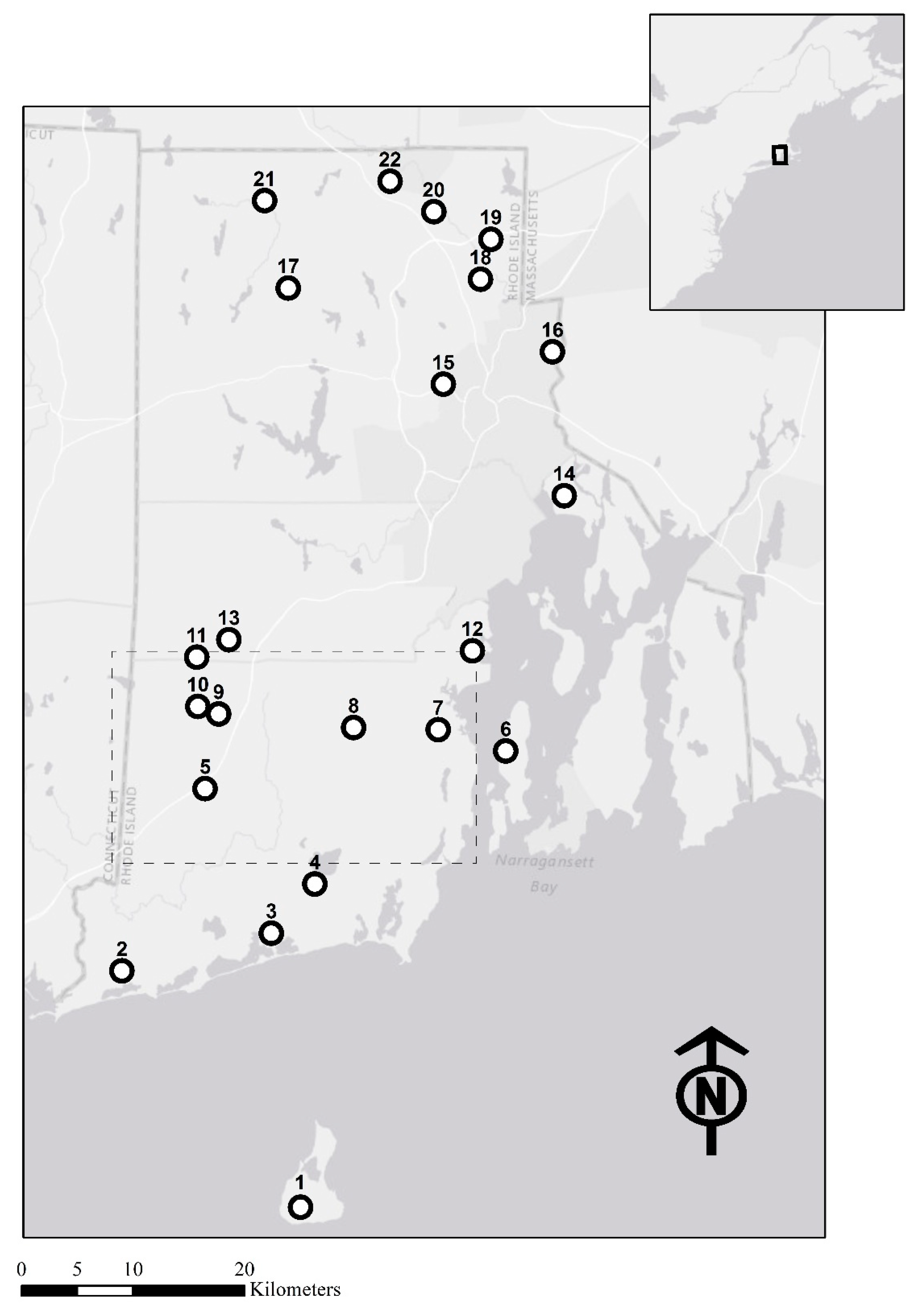
2.1. Study Area and Sampling
2.2. Microsatellite Genotyping
2.3. Genetic Diversity and Differentiation
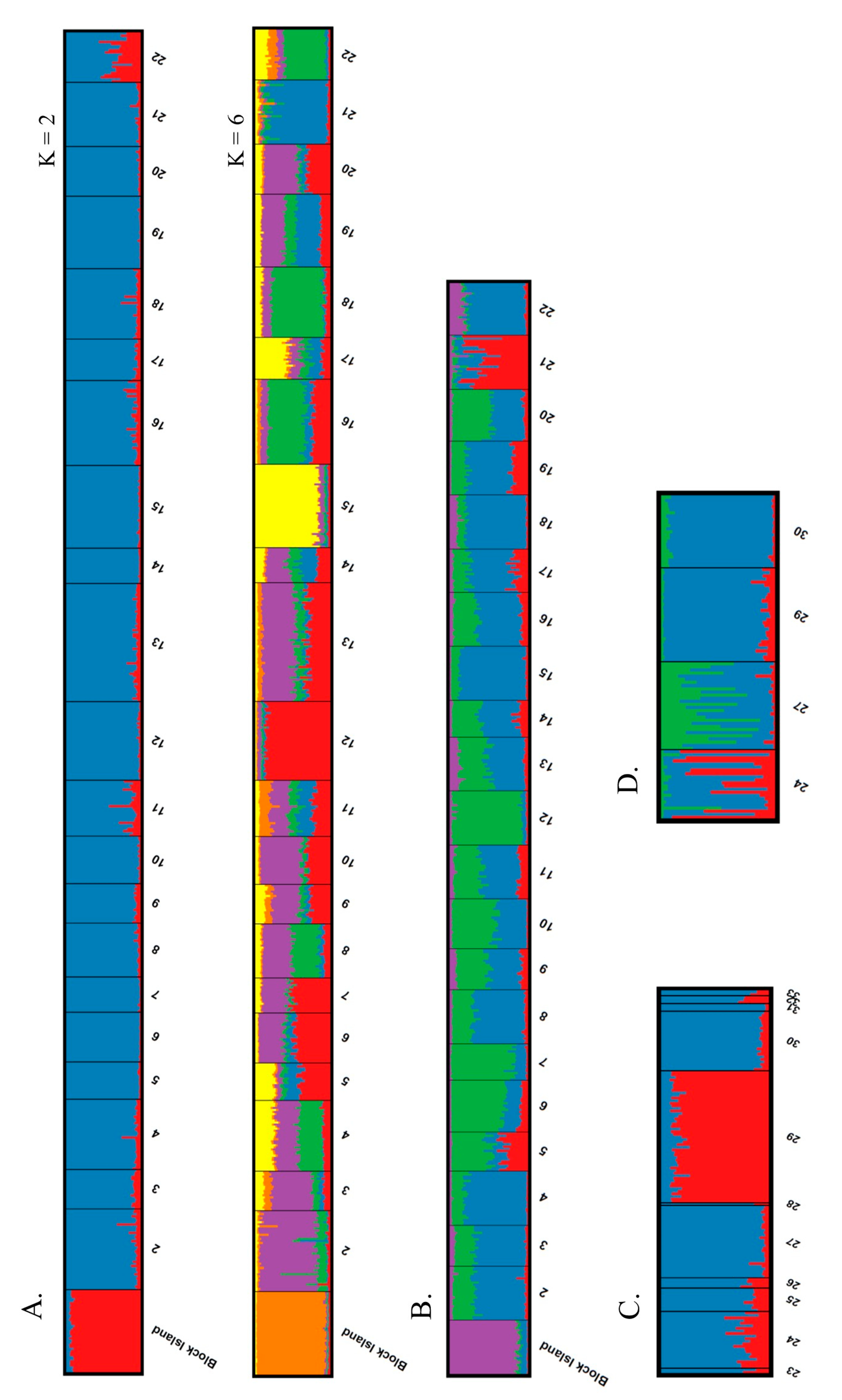
2.4. Population Structure
2.5. Comparison of Pooled Groups
3. Results
3.1. Sampling and Genotyping
3.2. Genetic Diversity and Population Structure
3.3. Comparison of Pooled Groups
4. Discussion
4.1. Genetic Diversity
4.2. Population Structure
4.3. Population Bottleneck and Effective Population Size
4.4. Scope and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foster, D.R.; Aber, J.D. Forests in Time: The Environmental Consequences of 1000 years of Change in New England; Yale University Press: New Haven, CT, USA, 2004; ISBN 9780300115376. [Google Scholar]
- Dahl, T.E. Wetlands Losses in the United States, 1780’s to 1980’s; U.S. Department of the Interior, Fish and Wildlife Service: Washington, DC, USA, 1990; 13p.
- Magilligan, F.J.; Graber, B.E.; Nislow, K.H.; Chipman, J.W.; Sneddon, C.S.; Fox, C.A. River restoration by dam removal: Enhancing connectivity at watershed scales. Elementa 2016, 4, 000108. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service. Bog Turtle (Clemmys muhlenbergii), Northern Population, Recovery Plan; U.S. Fish and Wildlife Service: Hadley, MA, USA, 2001; 103p.
- Rosenbaum, P.A.; Robertson, J.M.; Zamudio, K.R. Unexpectedly low genetic divergences among populations of the threatened bog turtle (Glyptemys muhlenbergii). Conserv. Genet. 2007, 8, 331–342. [Google Scholar] [CrossRef]
- Price, S.J.; Guzy, J.C.; Witczak, L.; Dorcas, M.E. Do ponds on golf courses provide suitable habitat for wetland-dependent animals in suburban areas? An assessment of turtle abundances. J. Herpetol. 2013, 47, 243–250. [Google Scholar] [CrossRef]
- Winchell, K.M.; Gibbs, J.P. Golf courses as habitat for aquatic turtles in urbanized landscapes. Landsc. Urban Plan. 2016, 147, 59–70. [Google Scholar] [CrossRef]
- Congdon, J.D.; Gibbons, J.W. Structure and Dynamics of a Turtle Community. In Long-Term Studies of Vertebrate Communities; Cody, M.L., Smallwood, J.A., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 137–159. [Google Scholar]
- Gamble, T.; Simons, A.M. Comparison of harvested and nonharvested painted turtle populations. Wildl. Soc. Bull. 2004, 32, 1269–1277. [Google Scholar] [CrossRef][Green Version]
- Ernst, C.H. Ecology of the spotted turtle, Clemmys guttata (Reptilia, Testudines, Testudinidae), in southeastern Pennsylvania. J. Herpetol. 1976, 10, 25–33. [Google Scholar] [CrossRef]
- Ernst, C.H.; Lovich, J.E. Turtles of the United States and Canada, 2nd ed.; John Hopkins University Press: Baltimore, MD, USA, 2009; ISBN 9780801891212. [Google Scholar]
- Wilbur, H.M. The evolutionary and mathematical demography of the turtle Chrysemys picta. Ecology 1975, 56, 64–77. [Google Scholar] [CrossRef]
- Zweifel, R.G. Long–term ecological studies on a population of painted turtles, Chrysemys picta, on Long Island, New York. Am. Mus. Novit. 1989, 2952, 1–55. [Google Scholar]
- Bowne, D.R. Terrestrial activity of Chrysemys picta in Northern Virginia. Copeia 2008, 2008, 306–310. [Google Scholar] [CrossRef]
- Tuberville, T.D.; Gibbons, J.W.; Greene, J.L. Invasion of new aquatic habitats by male freshwater turtles. Copeia 1996, 1996, 713–715. [Google Scholar] [CrossRef]
- Cosentino, B.J.; Schooley, R.L.; Phillips, C.A. Wetland hydrology, area, and isolation influence occupancy and spatial turnover of the painted turtle, Chrysemys picta. Landsc. Ecol. 2010, 25, 1589–1600. [Google Scholar] [CrossRef]
- Gibbons, J.W.; Scott, D.E.; Ryan, T.J.; Buhlmann, K.A.; Tuberville, T.D.; Metts, B.S.; Greene, J.L.; Mills, T.; Leiden, Y.; Poppy, S.; et al. The global decline of reptiles, déjà vu amphibians. BioScience 2000, 50, 653–666. [Google Scholar] [CrossRef]
- Lewis, T.L.; Ulmer, J.M.; Mazza, J.L. Threats to spotted turtle (Clemmys guttata) habitat in Ohio. Ohio J. Sci. 2004, 104, 65–71. [Google Scholar]
- Van Dijk, P.P. Clemmys guttata. The IUCN Red List of Threatened Species. e.T4968A97411228. 2011. Available online: http://dx.doi.org/10.2305/IUCN.UK.2011-1.RLTS.T4968A11103766.en (accessed on 15 January 2013).
- Litzgus, J.D.; Brooks, R.J. Growth in a cold environment: Body size and sexual maturity in a northern population of spotted turtles, Clemmys guttata. Can. J. Zool. 1998, 76, 773–782. [Google Scholar] [CrossRef]
- COSEWIC. COSEWIC Assessment and Update Status Report on Spotted Turtle Clemmys guttata in Canada; Committee on the Status of Endangered Wildlife in Canada Ottawa: Ottawa, ON, USA, 2014; 74p. [Google Scholar]
- Beaudry, F.; deMaynadier, P.G.; Hunter, M.L. Seasonally dynamic habitat use by spotted (Clemmys guttata) and blanding’s turtles (Emydoidea blandingii) in Maine. J. Herpetol. 2009, 43, 636–645. [Google Scholar] [CrossRef]
- Milam, J.C.; Melvin, S.M. Density, habitat use, movements, and conservation of spotted turtles (Clemmys guttata) in Massachusetts. J. Herpetol. 2001, 35, 418–427. [Google Scholar] [CrossRef]
- Rasmussen, M.L.; Litzgus, J.D. Habitat selection and movement patterns of spotted turtles (Clemmys guttata): Effects of spatial and temporal scales of analyses. Copeia 2010, 2010, 86–96. [Google Scholar] [CrossRef]
- Buchanan, S.W.; Buffum, B.; Puggioni, G.; Karraker, N.E. Occupancy of freshwater turtles across a gradient of altered landscapes. J. Wildl. Manag. 2019, 83, 435–445. [Google Scholar]
- Haxton, T.; Berrill, M. Habitat selectivity of Clemmys guttata in central Ontario. Can. J. Zool. 1999, 77, 593–599. [Google Scholar] [CrossRef]
- Litzgus, J.D.; Costanzo, J.P.; Brooks, R.J.; Lee, J.R.E. Phenology and ecology of hibernation in spotted turtles (Clemmys guttata) near the northern limit of their range. Can. J. Zool. 1999, 77, 1348–1357. [Google Scholar]
- U.S. Fish and Wildlife Service. Petition to List Spotted Turtle in Connecticut, Delaware, Florida, Georgia, Illinois, Maine, Maryland, Massachusetts, Michigan, Pennsylvania, New Hampshire, New York, North Carolina, Ohio, South Carolina, Vermont, Virginia, and West Virginia under the Endangered Species Act of 1973; as Amended; Federal Register Docket ID FWS–R5–ES–2015–0064; U.S. Fish and Wildlife Service: Washington, DC, USA, 2015.
- Brook, B.W. Demographics Versus Genetics in Conservation Biology. In Conservation Biology: Evolution in Action; Carroll, S.P., Fox, C.W., Eds.; Oxford University Press: New York, NY, USA, 2008; pp. 35–49. ISBN 978-0195306781. [Google Scholar]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0521702713. [Google Scholar]
- Ralls, K.; Ballou, J.D.; Templeton, A. Estimates of lethal equivalents and the cost of inbreeding in mammals. Conserv. Biol. 1988, 2, 185–193. [Google Scholar] [CrossRef]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- O’Grady, J.J.; Brook, B.W.; Reed, D.H.; Ballou, J.D.; Tonkyn, D.W.; Frankham, R. Realistic levels of inbreeding depression strongly affect extinction risk in wild populations. Biol. Conserv. 2006, 133, 42–51. [Google Scholar] [CrossRef]
- U.S. Census Bureau. Population, Housing Units, Area, and Density: 2010. 2010. Available online: https://factfinder.census.gov (accessed on 15 January 2013).
- Butler, B.J. Rhode Island’s Forest Resources, 2012; Res. Note, NRS-190; U.S. Forest Service: Newton Square, PA, USA, 2013; 3p.
- Uchupi, E.; Driscoll, N.; Ballard, R.D.; Bolmer, S.T. Drainage of late Wisconsin glacial lakes and the morphology and late quaternary stratigraphy of the New Jersey–southern New England continental shelf and slope. Mar. Geol. 2001, 172, 117–145. [Google Scholar] [CrossRef]
- Boothroyd, J.C.; Sirkin, L. Quaternary geology and landscape development of Block Island and adjacent regions. In Proceedings of the Rhode Island Natural History Survey, Kingston, RI, USA, 28 October 2000; pp. 13–27. [Google Scholar]
- Hale, M.L.; Burg, T.M.; Steeves, T.E. Sampling for microsatellite-based population genetic studies: 25 to 30 individuals per population is enough to accurately estimate allele frequencies. PLoS ONE 2012, 7, e45170. [Google Scholar] [CrossRef] [PubMed]
- Rhode Island Geographic Information System Home Page. Available online: http://www.rigis.org/ (accessed on 1 February 2013).
- Pearse, D.E.; Janzen, F.J.; Avise, J.C. Genetic markers substantiate long-term storage and utilization of sperm by female painted turtles. Heredity 2001, 86, 378–384. [Google Scholar] [CrossRef] [PubMed]
- King, T.L.; Julian, S.E. Conservation of microsatellite DNA flanking sequence across 13 Emydid genera assayed with novel bog turtle (Glyptemys muhlenbergii) loci. Conserv. Genet. 2004, 5, 719–725. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Resour. 2004, 4, 535–538. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 15 January 2018).
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed]
- Paradis, E. Pegas: An R package for population genetics with an integrated-modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [PubMed]
- Adamack, A.T.; Gruber, B. PopGenReport: Simplifying basic population genetic analyses in R. Methods Ecol. Evol. 2014, 5, 384–387. [Google Scholar] [CrossRef]
- Brookfield, J. A simple new method for estimating null allele frequency from heterozygote deficiency. Mol. Ecol. 1996, 5, 453–455. [Google Scholar] [CrossRef]
- Kalinowski, S.T. Counting alleles with rarefaction: Private alleles and hierarchical sampling designs. Conserv. Genet. 2004, 5, 539–543. [Google Scholar] [CrossRef]
- Keenan, K.; McGinnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. DiveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [PubMed]
- Jost, L. GST and its relatives do not measure differentiation. Mol. Ecol. 2008, 17, 4015–4026. [Google Scholar] [CrossRef]
- Gerlach, G.; Jueterbock, A.; Kraemer, P.; Deppermann, J.; Harmand, P. Calculations of population differentiation based on GST and D: Forget GST but not all of statistics! Mol. Ecol. 2010, 19, 3845–3852. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar]
- Kalinowski, S.T. Do polymorphic loci require large sample sizes to estimate genetic distances? Heredity 2005, 94, 33–36. [Google Scholar] [CrossRef]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Porras-Hurtado, L.; Ruiz, Y.; Santos, C.; Phillips, C.; Carracedo, Á.; Lareu, M.V. An overview of STRUCTURE: Applications, parameter settings, and supporting software. Front. Genet. 2013, 4, 98. [Google Scholar] [CrossRef]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Puechmaille, S.J. The program STRUCTURE does not reliably recover the correct population structure when sampling is uneven: Subsampling and new estimators alleviate the problem. Mol. Ecol. Resour. 2016, 16, 608–627. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Rosenberg, N.A. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Resour. 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.M. BOTTLENECK: A computer program for detecting recent reductions in the effective size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Luikart, G.; Cornuet, J.M. Empirical evaluation of a test for identifying recently bottlenecked populations from allele frequency data. Conserv. Biol. 1998, 12, 228–237. [Google Scholar] [CrossRef]
- Peery, M.Z.; Kirby, R.; Reid, B.N.; Stoelting, R.; Doucet-Beer, E.; Robinson, S.; Vasquez-Carrillo, C.; Pauli, J.N.; Palsboll, P. Reliability of genetic bottleneck tests for detecting recent population declines. Mol. Ecol. 2012, 21, 3403–3418. [Google Scholar] [CrossRef]
- Williamson-Natesan, E.G. Comparison of methods for detecting bottlenecks from microsatellite loci. Conserv. Genet. 2005, 6, 551–562. [Google Scholar] [CrossRef]
- Do, C.; Waples, R.S.; Peel, D.; MacBeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator v2: Re-implementation of software for the estimation of contemporary effective population sixe (Ne) from genetic data. Mol. Ecol. Resour. 2014, 14, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Ramirez, M.; Stuckas, H.; Castano-Mora, O.V.; Fritz, U. Extremely low genetic diversity and weak population differentiation in the endangered Colombian river turtle Podocnemis lewyana (Testudines: Podocnemididae). Conserv. Genet. 2012, 13, 65–77. [Google Scholar] [CrossRef]
- Kuo, C.H.; Janzen, F.J. Genetic effects of a persistent bottleneck on a natural population of ornate box turtles (Terrapene ornata). Conserv. Genet. 2004, 5, 425–437. [Google Scholar] [CrossRef]
- Parker, P.G.; Whiteman, H.H. Genetic diversity in fragmented populations of Clemmys guttata and Chrysemys picta marginata as shown by DNA fingerprinting. Copeia 1993, 1993, 841–846. [Google Scholar] [CrossRef]
- Reid, B.N.; Mladenoff, D.J.; Peery, M.Z. Genetic effects of landscape, habitat preference and demography on three co-occurring turtle species. Mol. Ecol. 2017, 26, 781–798. [Google Scholar] [CrossRef]
- Anthonysamy, W.B. Spatial Ecology, Habitat Use, Genetic Diversity, and Reproductive Success: Measures of Connectivity of a Sympatric Freshwater Turtle Assemblage in a Fragmented Landscape. Ph.D. Dissertation, University of Illinois at Urbana–Champaign, Champaign, IL, USA, 2012. [Google Scholar]
- Rubinsztein, D.C.; Amos, W.; Leggo, J.; Goodburn, S.; Jain, S.; Li, S.H.; Margolis, R.L.; Ross, C.A.; Ferguson-Smith, M.A. Microsatellite evolution—Evidence for directionality and variation in rate between species. Nat. Genet. 1995, 10, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Väli, Ü.; Einarsson, A.; Waits, L.; Ellegren, H. To what extent do microsatellite markers reflect genome-wide genetic diversity in natural populations? Mol. Ecol. 2008, 17, 3808–3817. [Google Scholar] [CrossRef] [PubMed]
- Sirkin, L. Block Island Geology; Book and Tackle Shop: Westerly, RI, USA, 1996; 203p. [Google Scholar]
- Starkey, D.E.; Shaffer, H.B.; Burke, R.L.; Forstner, M.R.; Iverson, J.B.; Janzen, F.J.; Rhodin, A.G.; Ultsch, G.R. Molecular systematics, phylogeography, and the effects of Pleistocene glaciation in the painted turtle (Chrysemys picta) complex. Evolution 2003, 57, 119–128. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M.; Brooks, S.; Churchill, T.A.; Brooks, R.J. Hatchling turtles survive freezing during winter hibernation. Proc. Natl. Acad. Sci. USA 1988, 85, 8350–8354. [Google Scholar] [CrossRef] [PubMed]
- Churchill, T.A.; Storey, K.B. Natural freezing survival by painted turtles Chrysemys picta marginata and C. picta bellii. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1992, 262, R530–R537. [Google Scholar] [CrossRef] [PubMed]
- Holman, J.; Andrews, K.D. North American Quaternary cold–tolerant turtles: Distributional adaptations and constraints. Boreas 1994, 23, 44–52. [Google Scholar] [CrossRef]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–9013. [Google Scholar] [CrossRef]
- Weisrock, D.W.; Janzen, F.J. Comparative molecular phylogeography of North American softshell turtles (Apalone): Implications for regional and wide-scale historical evolutionary forces. Mol. Phylogenet. Evol. 2000, 14, 152–164. [Google Scholar] [CrossRef]
- Buchanan, S.W.; Buffum, B.; Karraker, N.E. Responses of a spotted turtle (Clemmys guttata) population to creation of early-successional habitat. Herpetol. Conserv. Biol. 2017, 12, 688–700. [Google Scholar]
- Landguth, E.; Cushman, S.; Schwartz, M.; McKelvey, K.; Murphy, M.; Luikart, G. Quantifying the lag time to detect barriers in landscape genetics. Mol. Ecol. 2010, 19, 4179–4191. [Google Scholar] [CrossRef] [PubMed]
- Avise, J.C.; Bowen, B.W.; Lamb, T.; Meylan, A.B.; Bermingham, E. Mitochondrial DNA evolution at a turtle’s pace: Evidence for low genetic variability and reduced microevolutionary rate in the Testudines. Mol. Biol. Evol. 1992, 9, 457–473. [Google Scholar] [PubMed]
- Shaffer, H.B.; Minx, P.; Warren, D.E.; Shedlock, A.M.; Thomson, R.C.; Valenzuela, N.; Abramyan, J.; Amemiya, C.T.; Badenhorst, D.; Biggar, K.K. The western painted turtle genome, a model for the evolution of extreme physiological adaptations in a slowly evolving lineage. Genome Biol. 2013, 14, R28. [Google Scholar] [CrossRef]
- Bennett, A.M.; Keevil, M.; Litzgus, J.D. Spatial ecology and population genetics of northern map turtles (Graptemys geographica) in fragmented and continuous habitats in Canada. Chelonian Conserv. Biol. 2010, 9, 185–195. [Google Scholar] [CrossRef]
- Kolbe, J.J.; Leal, M.; Schoener, T.W.; Spiller, D.A.; Losos, J.B. Founder effects persist despite adaptive differentiation: A replicated field experiment in a Caribbean lizard. Science 2012, 335, 1086–1089. [Google Scholar] [CrossRef]
- Blair, C.; Arcos, V.H.J.; de la Cruz, F.R.M.; Murphy, R.W. Landscape genetics of leaf-toed geckos in the tropical dry forest of Northern Mexico. PLoS ONE 2013, 8, e57433. [Google Scholar] [CrossRef] [PubMed]
- Delaney, K.S.; Riley, S.P.; Fisher, R.N. A rapid, strong, and convergent genetic response to urban habitat fragmentation in four divergent and widespread vertebrates. PLoS ONE 2010, 5, e12767. [Google Scholar] [CrossRef]
- Spinks, P.Q.; Thomson, R.C.; Shaffer, H.B. The advantages of going large: Genome-wide SNPs clarify the complex population history and systematics of the threatened western pond turtle. Mol. Ecol. 2014, 23, 2228–2241. [Google Scholar] [CrossRef]
- Elbers, J.P.; Clostio, R.W.; Taylor, S.S. Population genetic inferences using immune gene SNPs mirror patterns inferred by microsatellites. Mol. Ecol. Resour. 2016, 17, 481–491. [Google Scholar] [CrossRef]
- Kautz, R.; Kawula, R.; Hoctor, T.; Comiskey, J.; Jansen, D.; Jennings, D.; Kasbohm, J.; Mazzotti, F.; McBride, R.; Richardson, L. How much is enough? Landscape-scale conservation for the Florida panther. Biol. Conserv. 2006, 130, 118–133. [Google Scholar] [CrossRef]
- Shoemaker, K.T.; Gibbs, J.P. Genetic connectivity among populations of the threatened bog turtle (Glyptemys muhlenbergii) and the need for a regional approach to turtle conservation. Copeia 2013, 2013, 324–331. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Site | Geographic Coordinates (Latitude, Longitude) | No. of Individuals | He | Ho | Private Alleles | Mean Allelic Richness | FIS |
|---|---|---|---|---|---|---|---|
| Painted turtles | |||||||
| 1 | 41.153247, −71.604493 | 40 | 0.63 | 0.63 | 2 | 5.83 | 0.014 |
| 2 | 41.346108, −71.789074 | 39 | 0.61 | 0.60 | 0 | 6.21 | −0.012 |
| 3 | 41.380141, −71.630523 | 19 | 0.61 | 0.67 | 1 | 5.82 | −0.101 |
| 4 | 41.42052, −71.585525 | 34 | 0.63 | 0.66 | 0 | 6.29 | −0.062 |
| 5 * | 41.494763, −71.706265 | 18 | 0.61 | 0.66 | 1 | 5.69 | −0.076 |
| 6 | 41.53227, −71.385252 | 24 | 0.59 | 0.59 | 0 | 5.54 | −0.003 |
| 7 * | 41.548007, −71.458184 | 17 | 0.59 | 0.63 | 0 | 5.75 | −0.076 |
| 8 * | 41.547675, −71.549071 | 26 | 0.63 | 0.64 | 2 | 6.59 | −0.022 |
| 9 * | 41.555359, −71.694297 | 19 | 0.63 | 0.71 | 1 | 6.33 | −0.127 |
| 10 * | 41.560943, −71.716856 | 23 | 0.63 | 0.64 | 0 | 6.22 | −0.018 |
| 11 * | 41.600674, −71.719651 | 27 | 0.65 | 0.68 | 0 | 6.21 | −0.058 |
| 12 | 41.612399, −71.42399 | 38 | 0.64 | 0.69 | 0 | 5.96 | −0.073 |
| 13 | 41.615407, −71.685647 | 57 | 0.62 | 0.63 | 1 | 6.34 | −0.029 |
| 14 | 41.739527, −71.329793 | 17 | 0.62 | 0.67 | 0 | 5.79 | −0.065 |
| 15 | 41.826773, −71.463335 | 40 | 0.62 | 0.60 | 2 | 6.02 | 0.034 |
| 16 | 41.855324, −71.346786 | 41 | 0.64 | 0.63 | 1 | 6.54 | 0.016 |
| 17 | 41.900587, −71.633623 | 20 | 0.63 | 0.69 | 0 | 6.25 | −0.100 |
| 18 | 41.912204, −71.426565 | 34 | 0.61 | 0.61 | 0 | 6.18 | −0.005 |
| 19 | 41.944757, −71.416485 | 35 | 0.65 | 0.71 | 0 | 6.29 | −0.090 |
| 20 | 41.965696, −71.478915 | 24 | 0.62 | 0.67 | 0 | 6.12 | −0.078 |
| 21 | 41.970512, −71.66193 | 31 | 0.64 | 0.68 | 0 | 5.94 | −0.064 |
| 22 | 41.98915, −71.527097 | 24 | 0.59 | 0.61 | 2 | 5.98 | −0.018 |
| Global values | 607 | - | - | - | - | −0.022 | |
| Pooled group | - | 130 | 0.64 | 0.66 | - | 10.27 | −0.026 |
| Spotted turtles | |||||||
| 23 | - | 2 | - | - | 1 | - | - |
| 24 * | - | 22 | 0.65 | 0.63 | 2 | 4.78 | 0.040 |
| 25 * | - | 9 | 0.67 | 0.68 | 2 | 4.81 | −0.028 |
| 26 * | - | 4 | - | - | 1 | - | - |
| 27 * | - | 28 | 0.67 | 0.67 | 3 | 4.86 | −0.008 |
| 28 | - | 1 | - | - | 1 | - | - |
| 29 * | - | 51 | 0.67 | 0.64 | 5 | 4.90 | 0.033 |
| 30 * | - | 23 | 0.68 | 0.67 | 6 | 4.97 | 0.007 |
| 31 | - | 3 | - | - | 1 | - | - |
| 32 | - | 3 | - | - | 1 | - | - |
| 33 | - | 2 | - | - | 1 | - | - |
| Global values | - | 133 | - | - | - | - | 0.036 |
| Pooled group | - | 137 | 0.68 | 0.66 | - | 8.59 | 0.039 |
| Proportion of Multi-Step Mutation Model | ||||||||
|---|---|---|---|---|---|---|---|---|
| 0.05 | 0.15 | 0.25 | 0.35 | |||||
| Species | Sign Test | Wilcoxon Test | Sign Test | Wilcoxon Test | Sign Test | Wilcoxon Test | Sign Test | Wilcoxon Test |
| Painted turtles | 0.586 | 0.545 | 0.337 | 0.425 | 0.331 | 0.395 | 0.322 | 0.259 |
| Spotted turtles | 0.505 | 0.550 | 0.478 | 0.161 | 0.477 | 0.072 | 0.048 * | 0.017 * |
| Critical Value | Ne | Parametric 95% CI | Jack-Knife 95% CI | Ne | Parametric 95% CI | Jack-Knife 95% CI |
|---|---|---|---|---|---|---|
| Painted turtles | Spotted turtles | |||||
| 0.05 | 283.3 | 174–642.3 | 132–4387.4 | 242.1 | 175.3–371.9 | 141.9–611.5 |
| All alleles | 692.9 | 395.6–2354.3 | 237.2–Infinite | 334.9 | 255.7–473.8 | 206.7–756.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buchanan, S.W.; Kolbe, J.J.; Wegener, J.E.; Atutubo, J.R.; Karraker, N.E. A Comparison of the Population Genetic Structure and Diversity between a Common (Chrysemys p. picta) and an Endangered (Clemmys guttata) Freshwater Turtle. Diversity 2019, 11, 99. https://doi.org/10.3390/d11070099
Buchanan SW, Kolbe JJ, Wegener JE, Atutubo JR, Karraker NE. A Comparison of the Population Genetic Structure and Diversity between a Common (Chrysemys p. picta) and an Endangered (Clemmys guttata) Freshwater Turtle. Diversity. 2019; 11(7):99. https://doi.org/10.3390/d11070099
Chicago/Turabian StyleBuchanan, Scott W., Jason J. Kolbe, Johanna E. Wegener, Jessica R. Atutubo, and Nancy E. Karraker. 2019. "A Comparison of the Population Genetic Structure and Diversity between a Common (Chrysemys p. picta) and an Endangered (Clemmys guttata) Freshwater Turtle" Diversity 11, no. 7: 99. https://doi.org/10.3390/d11070099
APA StyleBuchanan, S. W., Kolbe, J. J., Wegener, J. E., Atutubo, J. R., & Karraker, N. E. (2019). A Comparison of the Population Genetic Structure and Diversity between a Common (Chrysemys p. picta) and an Endangered (Clemmys guttata) Freshwater Turtle. Diversity, 11(7), 99. https://doi.org/10.3390/d11070099